A Phytosociological Study on Andean Rainforests of Peru, and a Comparison with the Surrounding Countries

 ,
,  and
and 
Abstract
1. Introduction
2. Results
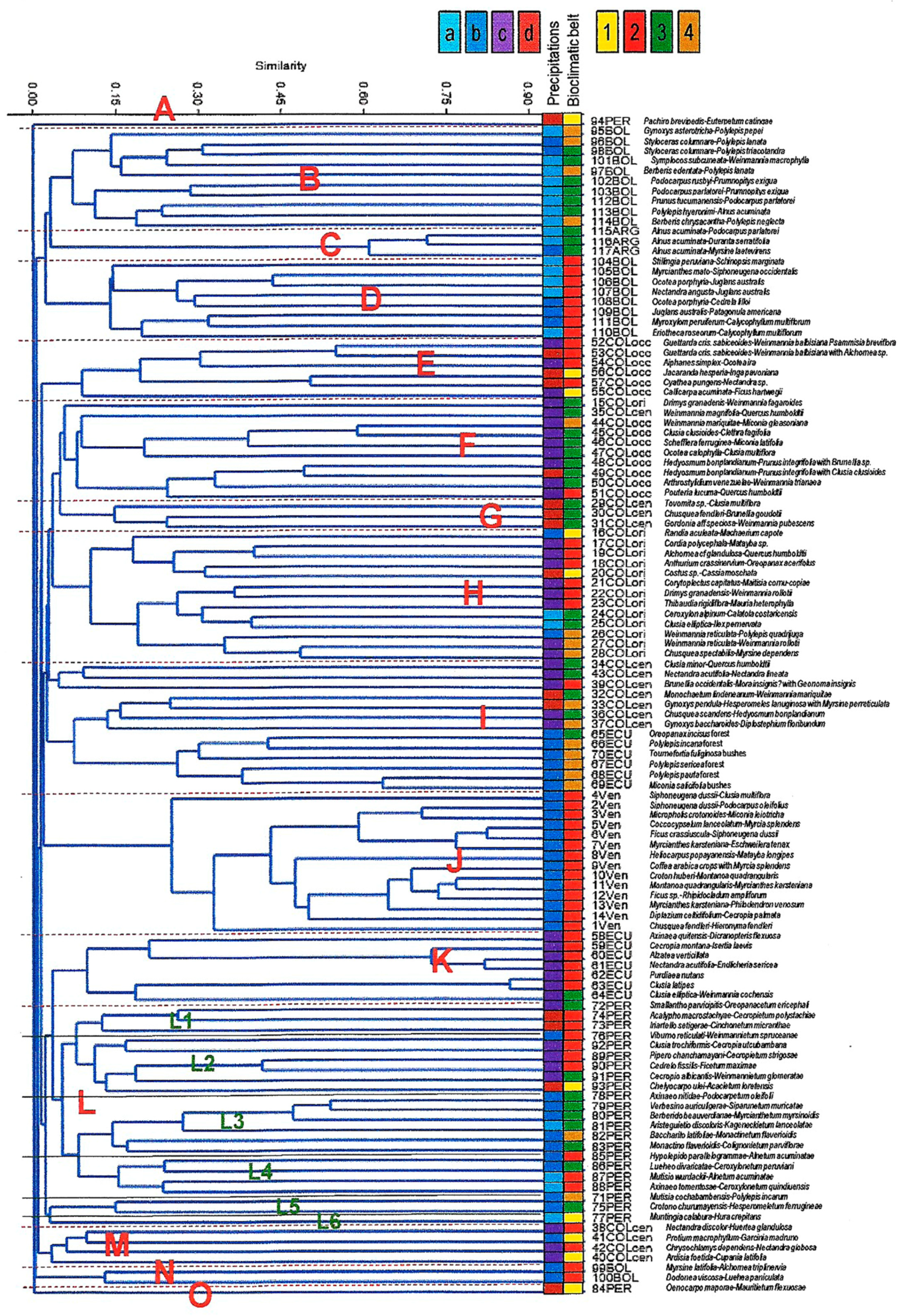
2.1. Relationships among South American Montane Rainforests
2.2. Phytosociological Study in Peru
2.2.1. Describing the New Associations and Plant Communities
Mutisia cochabambensis Hieron.-Polylepis incarum community
Smallantho parvicipitis-Oreopanacetum eriocephali ass. nov.
Iriartello setigerae-Cinchonetum micranthae ass. nov.
Acalypho macrostachyae-Cecropietum polystachyae ass. nov.
Crotono churumayensis-Hesperomeletum ferrugineae ass. nov.
Viburno reticulati-Weinmannietum spruceanae ass. nov.
Muntingia calabura-Hura crepitans community
Hypolepido parallelogrammae-Alnetum acuminatae ass. nov.
Lueheo divaricatae-Ceroxylonetum peruviani ass. nov.
Mutisio wurdackii-Alnetum acuminatae ass. nov.
Axinaeo tomentosae-Ceroxylonetum quindiuensis ass. nov.
Piperi chanchamayani-Cecropietum strigosae ass. nov.
Cedrelo fissilis-Ficetum maximae ass. nov.
Cecropio albicantis-Weinmannietum glomeratae ass. nov.
Clusia trochiformis-Cecropia utcubambana Community
Chelyocarpo ulei-Acacietum loretensis ass. nov.
2.2.2. Describing New Alliances, Orders and Classes
3. Discussion
3.1. Relationships among South American Montane Rainforests
3.2. Phytosociological Units Previously Described
3.3. Syntaxonomical Checklist for the Montane Rainforests of Peru
4. Materials and Methods
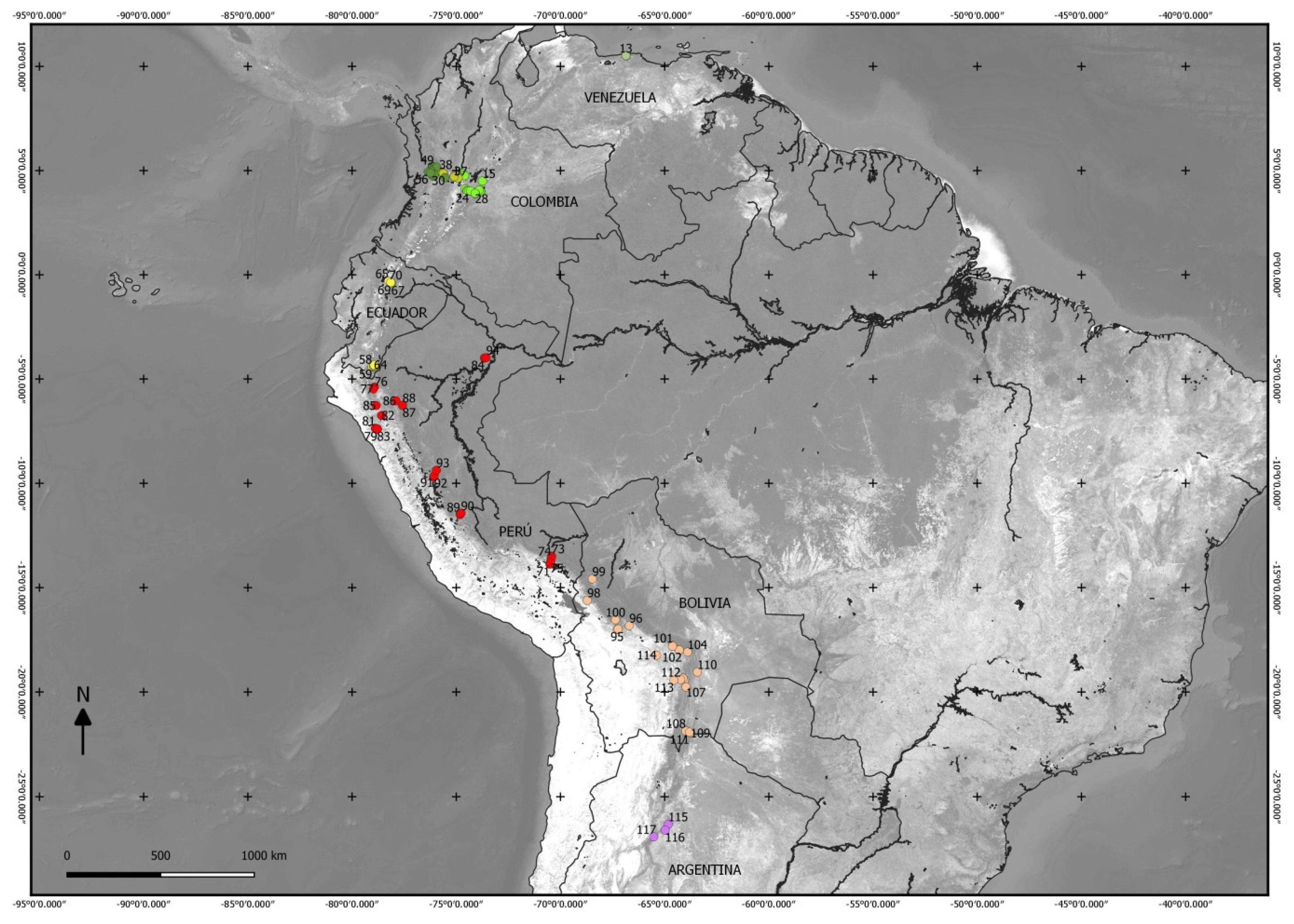
4.1. Study Area
4.2. Plots and Flora of Peru
4.3. Relationships among South American Rainforests
4.4. Vegetation Classification
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Column Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Columns of Table S3 | 71 | 72 | 73 | 74 | 75 | 76 | 77 | 78 | 79 | 80 | 81 | 82 | 83 | 84 | 85 | 86 | 87 | 88 | 89 | 90 | 91 | 92 | 93 | 94 |
| Number of plots | 1 | 5 | 7 | 7 | 3 | 6 | 1 | 5 | 3 | 5 | 4 | 7 | 2 | 2 | 8 | 5 | 2 | 4 | 3 | 6 | 5 | 1 | 4 | 10 |
| Reference of plot intervals of Tables S2 and S3 | 214 | 215–219 | 220–226 | 227–233 | 234–236 | 237–242 | 243 | 244–248 | 249–251 | 252–256 | 257–260 | 261–267 | 268–269 | 270–271 | 272–279 | 280–284 | 285–286 | 287–290 | 291–293 | 294–299 | 300–304 | 305 | 306–309 | 310–319 |
| Average altitude (m) | 3786 | 2273 | 752 | 1440 | 2754 | 1713 | 575 | 2519 | 2719 | 2540 | 3277 | 3260 | 2559 | 150 | 2474 | 1818 | 2337 | 2400 | 2313 | 1355 | 2654 | 1762 | 865 | 124 |
| Mutisia cochabambensis-Polylepis incarum community | ||||||||||||||||||||||||
| Polylepis incarum(T) | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Mutisia cochabambensis(C) | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Citharexylum dentatum(S) | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Elaphoglossum engelii(H) | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Gaultheria vaccinioides(SS) | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Smallantho parvicipitis-Oreopanacetum eriocephali ass. nov. | ||||||||||||||||||||||||
| Smallanthus parviceps(T) | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Fuchsia boliviana(S) | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Begonia bracteosa(H) | - | 100 | - | 14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Melica scabra(H) | - | 80 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Miconia aff adinantha(S) | - | 100 | - | 29 | - | - | - | - | - | - | - | - | - | - | 25 | - | - | - | - | - | - | - | - | - |
| Phenax angustifolius(T) | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Solanum aphyodendron(T) | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 17 | - | - | - | - |
| Rubus boliviensis(S) | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pennisetum latifolium(H) | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Croton regelianus(S) | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Canna iridiflora(H) | - | 40 | - | 14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Sambucus peruviana(T) | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Iriartello setigerae-Cinchonetum micranthae ass. nov. | ||||||||||||||||||||||||
| Cinchona micrantha(T) | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Iriartella setigera(S) | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cyathea subincisa(T) | - | - | 71 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Philodendron rudgeanum(Ep) | - | - | 71 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Heliconia hirsuta(H) | - | - | 86 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - |
| Ruagea insignis(T) | - | - | 57 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Dieffenbachia humilis(H) | - | - | 57 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Miconia lourteigiana(T) | - | - | 57 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Aphelandra aurantiaca(S) | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Proteaceae_sg(T) | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Smilax purhampuy(C) | - | - | 43 | 14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 17 | - | - | 25 | - |
| Strychnos sp._sg(S) | - | 20 | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Acalypho macrostachyae-Cecropietum polystachyae ass. nov. | ||||||||||||||||||||||||
| Cecropia polystachya(T) | - | 20 | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Maranta arundinacea(H) | - | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Acalypha macrostachya(T) | - | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 17 | - | - | 25 | - |
| Crotono churumayensis-Hesperomeletum ferrugineae ass. nov. | ||||||||||||||||||||||||
| Croton churumayensis(T) | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cuphea cordata(S) | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Tagetes elliptica(S) | - | - | - | - | 67 | - | - | - | - | - | - | 29 | - | - | - | - | - | - | - | - | - | - | - | - |
| Senna birostris(S) | - | - | - | - | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Adiantum digitatum(H) | - | - | - | - | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Peperomia sp._sg158(H) | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Miconia alpina(S) | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Gaultheria reticulata(S) | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Ditassa sp._sg(Ep) | - | 20 | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Baccharis genistelloides(SS) | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Viburno reticulati-Weinmannietum spruceanae ass. nov. | ||||||||||||||||||||||||
| Viburnum reticulatum(T) | - | - | - | - | - | 83 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pouzolzia poeppigiana(T) | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Weinmannia spruceana(T) | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Jatropha sp.(S) | - | - | - | 14 | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Anthurium hamiltonii(H) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Mandevilla fragrans(C) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Clusia crenata(T) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pitcairnia paniculata(H) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Rubus urticifolius(S) | - | - | - | 14 | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Gleichenella pectinata(H) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Muntingia calabura-Hura crepitans community | ||||||||||||||||||||||||
| Hura crepitans(T) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Ruellia brevifolia(S) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Albizia multiflora(T) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Piper peltatum(H) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Piper nudilimbum(S) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Ichnanthus nemorosus(H) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Leucaena trichodes(T) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Muntingia calabura(T) | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Paullinia alata(C) | - | - | 29 | 14 | - | 17 | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Axinaeo nitidae-Podocarpetum oleifolii | ||||||||||||||||||||||||
| Axinaea nitida(S) | - | - | - | - | - | - | - | 100 | 33 | - | - | - | - | - | 38 | 40 | - | - | - | - | - | - | - | - |
| Podocarpus oleifolius(T) | - | - | - | - | - | - | - | 100 | 20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Trichilia tomentosa(T) | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Chrysophyllum contumazense(T) | - | - | - | - | - | - | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Miconia firma(S) | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Viola argute(C) | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Verbesino auriculigerae-Siparunetum muricatae | ||||||||||||||||||||||||
| Siparuna muricata(T) | - | - | - | - | - | - | - | 40 | 100 | 100 | - | - | - | - | 25 | - | - | - | 33 | 50 | - | - | - | - |
| Verbesina auriculigera(S) | - | - | - | - | - | - | - | - | 100 | 40 | 25 | - | 100 | - | - | - | - | - | - | - | - | - | - | - |
| Cervantesia tomentosa(T) | - | - | - | - | - | - | - | - | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Meliosma frondosa(T) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Valeriana asterothrix(H) | - | - | - | - | - | - | - | - | 33 | 20 | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Solanum caripense(C) | - | - | - | - | - | - | - | - | 33 | 20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Calceolaria calycina(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Piper barbatum(C) | - | - | - | - | - | - | - | - | 33 | - | - | 29 | - | - | 13 | - | - | - | - | - | - | - | - | - |
| Berberis buceronis(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Munnozia ferreyrii(H) | - | - | - | - | - | - | - | - | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pappobolus sp._cjvs8(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Phenax laxiflorus(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lepechinia lamiifolia(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Scutia spicata(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Siphocampylus macropodoides(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Wedelia latifolia(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Symplocos sandemanii(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Fabaceae_cjvs6(T) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Begonia acerifolia(H) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Rubus sp._cjvs6(S) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Luffa operculata(C) | - | - | - | - | - | - | - | - | 33 | - | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Mikania sp._cjvs6(C) | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Paspalum candidum(H) | - | - | - | - | - | - | - | - | 33 | - | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Berberido beauverdianae-Myrcianthetum myrsinoidis | ||||||||||||||||||||||||
| Maytenus boarioides(S) | - | - | - | - | - | - | - | - | - | 80 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Viburnum mathewsii(T) | - | - | - | - | - | - | - | - | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Bidens squarrosa(H) | - | - | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Bomarea cornuta(C) | - | - | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - |
| Tecoma stans(T) | - | - | - | - | - | - | - | 20 | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Salvia oppositiflora(S) | - | - | - | - | - | - | - | - | - | 40 | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Liabum solidagineum(S) | - | - | - | - | - | - | - | 20 | - | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Thalictrum longistylum(H) | - | 20 | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Aristeguietio discoloris-Kageneckietum lanceolatae | ||||||||||||||||||||||||
| Kageneckia lanceolata(T) | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Aristeguietia discolor(S) | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Baccharis emarginata(S) | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Calceolaria rugulosa(S) | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Colletia spinosissima(S) | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cronquistianthus urubambensis(S) | - | - | - | 14 | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Coreopsis fasciculate(SS) | - | - | - | - | - | - | - | - | - | - | 75 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Scutellaria scutellarioides(SS) | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Passiflora peduncularis(C) | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Eryngium weberbaueri(H) | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Baccharito latifoliae-Monactinetum flaverioidis | ||||||||||||||||||||||||
| Passiflora tarminiana(C) | - | - | - | - | - | - | - | - | - | - | - | 57 | - | - | - | - | - | - | - | - | - | - | - | - |
| Salvia alborosea(S) | - | - | - | - | - | - | - | - | - | - | - | 57 | - | - | - | 20 | - | - | - | - | - | - | - | - |
| Cronquistianthus glomeratus(S) | - | - | - | - | - | - | - | - | - | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - |
| Calceolaria bicrenata(S) | - | - | - | - | - | - | - | - | - | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - |
| Minthostachys mollis(S) | - | 20 | - | - | - | - | - | - | - | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - |
| Monactino flaverioidis-Colignonietum parviflorae | ||||||||||||||||||||||||
| Colignonia parviflora(S) | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - |
| Alternanthera villosa(H) | - | - | - | - | - | - | - | - | - | 20 | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - |
| Heliotropium incanum(H) | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Urtica peruviana(H) | - | - | - | - | - | - | - | 20 | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Viguiera procumbens(S) | - | - | - | - | - | - | - | - | - | - | 25 | 14 | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Oenocarpo maporae-Mauritietum flexuosae | ||||||||||||||||||||||||
| Oenocarpus mapora(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Mauritia flexuosa(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Buchenavia oxycarpa(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Ficus americana subsp. guianensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Dialium guianense(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Virola decorticans(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Symphonia globulifera(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Geonoma macrostachys(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Rinorea racemosa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Agonandra sylvatica(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Planchonella obovata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Duguetia quitarensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Bactris concinna(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - |
| Inga sp. 01_jhom11(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Costus scaber(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Virola calophylla(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Ceiba pentandra(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Anthurium apaporanum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Terminalia amazonia(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Chrysobalanaceae_jhom11(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Euterpe precatoria(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Hevea guianensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Socratea exorrhiza(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - |
| Hypolepido parallelogrammae-Alnetum acuminatae ass. nov. | ||||||||||||||||||||||||
| Cyathea herzogii(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - |
| Hypolepis parallelogramma(H) | - | - | - | - | - | 17 | - | - | - | - | - | - | - | - | 63 | - | - | - | - | - | - | - | - | - |
| Hypolepis obtusata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - |
| Aulonemia longiaristata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Miconia aff asperrima(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Gurania sp. 01_cut(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Solanum sp. 02_cut(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Acanthus ilicifolius(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Inga acuminata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Miconia sp. 03_cut(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Hedyosmum sprucei(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Miconia aggregata(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Asplenium auriculatum(H) | - | - | - | - | - | - | - | - | - | - | - | 14 | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Cyathea lechleri(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | 20 | - | - | - |
| Geonoma orbignyana(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Saccoloma inaequale(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | - | - | - | - | - | - |
| Serpocaulon fraxinifolium(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | 20 | - | - | - | - | - | - | - | - |
| Lueheo divaricatae-Ceroxylonetum peruviani ass. nov. | ||||||||||||||||||||||||
| Luehea divaricata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - |
| Ceroxylon peruvianum(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - | - |
| Styloceras laurifolium(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 80 | - | - | - | - | - | - | - | - |
| Pueraria phaseoloides(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 | - | - | - | - | - | - | - | - |
| Solanum sp. 03(S) | - | - | - | - | - | 17 | - | - | - | - | - | 14 | - | - | 13 | 60 | - | - | - | - | - | - | - | - |
| Austroeupatorium inulaefolium(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - |
| Schistocarpha sinforosi(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 13 | 40 | - | - | - | - | - | - | - | - |
| Viburnum incarum(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 13 | 40 | - | - | - | - | - | - | - | - |
| Ladenbergia oblongifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - | 17 | - | - | 25 | - |
| Mutisio wurdackii-Alnetum acuminatae ass. nov. | ||||||||||||||||||||||||
| Mutisia wurdackii(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 25 | - | - | - | - | - | - |
| Weinmannia costulata(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - | - |
| Cortaderia jubata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - | - |
| Axinaeo tomentosae-Ceroxylonetum quindiuensis ass. nov. | ||||||||||||||||||||||||
| Ceroxylon quindiuense(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - | - |
| Cedrela odorata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | 100 | - | - | - | - | - | - |
| Axinaea tomentosa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 75 | - | - | - | - | - | - |
| Cronquistianthus chachapoyensis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 13 | 20 | - | 75 | - | - | - | - | - | - |
| Citronella incarum(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Vismia pozuzoensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Erythrina aff edulis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Solanum barbulatum(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Hedyosmum lechleri(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | 20 | - | - | - |
| Viburnum pichinchense(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Cornus peruviana(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Piperi chanchamayani-Cecropietum strigosae ass. nov. | ||||||||||||||||||||||||
| Pennisetum aff peruvianum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - | - |
| Ageratina tambillensis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 17 | 20 | - | - | - |
| Solanum mite(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - | - |
| Centropogon altus(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - | - |
| Cecropia angustifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | - | 67 | 17 | - | - | - | - |
| Cecropia strigose(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - | - |
| Clusia aff pallida(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - | - |
| Cavendishia punctate(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Pilea aff bassleriana(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Pilea pavonii(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Palicourea sp nov?sat2(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Acinodendron aff ferrugineum(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Thelypteris euchlora(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Adianthum raddianum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Coriaria myrtifolia(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Solanum asperolanatum(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | 33 | - | - | - | - | - |
| Miconia sanguinea(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 17 | - | - | - | - |
| Canna paniculata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 17 | - | - | - | - |
| Chromolaena leptocephala(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | 20 | - | - | - |
| Miconia sp. 08_sat(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 17 | - | - | - | - |
| Meriania tomentosa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 20 | - | - | - | - |
| Psammisia coarctata(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Anthurium grande(H) | - | - | - | 29 | - | 17 | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Cedrelo fissilis-Ficetum maximae ass. nov. | ||||||||||||||||||||||||
| Ficus maxima(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | - |
| Dieffenbachia sp. 02_sat(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 83 | - | - | - | - |
| Dioscorea sp. 01_sat(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - |
| Inga adenophylla(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - |
| Serjania communis(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - |
| Begonia parviflora(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | - | - | 67 | 20 | - | - | - |
| Heliconia subulata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | 67 | - | - | - | - |
| Lastreopsis effuse(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - |
| Ocotea obovata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - |
| Centropogon granulosus(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - |
| Philodendron aff killipii(Ep) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - |
| Selaginella trisulcata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - |
| Croton lechheri(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - |
| Campyloneurum ophicaulon(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - |
| Prestoea carderi(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | 20 | - | - | - |
| Vernonanthura patens(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | 50 | - | - | - | - |
| Urera baccifera(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | - | - | 50 | - | - | 25 | - |
| Alsophila mostellaria(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Thelypteris jamesonii(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Melastomataceae 02_sat(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Thelypteris salzmannii(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Costus sp. 02_sat(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Urera verrucosa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Columnea inaquilatera(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Serpocaulon loriceum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | 25 | - | 33 | - | - | - | - |
| Pteris altissima(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Nephrolepis pectinata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - |
| Bignoniaceae(C) | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | 33 | - | - | - | - |
| Cecropio albicantis-Weinmannietum glomeratae ass. nov. | ||||||||||||||||||||||||
| Cecropia albicans(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 | - | - | - |
| Weinmannia latifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 13 | - | - | - | - | - | 60 | - | - | - |
| Solanum aff sycophanta(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 | - | - | - |
| Tibouchina saxosa(SS) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 | - | - | - |
| Sticherus revolutus(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 | - | - | - |
| Cavendishia nolbilis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 | - | - | - |
| Piper augustum(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | 25 | - |
| Oreopanax aff cheirophyllus(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Vaccinium crenatum(SS) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Miconia galactantha(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Centropogon reflexus(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Dioscorea mitoensis(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Weinmannia auriculata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Centropogon hirtus(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Ageratina rhytidodes(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Thelypteris pachyrhachys(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Bomarea pseudopurpurea(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Elleanthus aurantiacus(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Weinmannia microphylla(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Tibouchina lepidota(SS) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Disterigma acuminatum(SS) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Clusia trochiformis-Cecropia utcubambana community | ||||||||||||||||||||||||
| Fuchsia macrophylla(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | 100 | - | - |
| Serpocaulon adnatum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | 100 | 25 | - |
| Barnadesia corymbosa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | - | - | - | - | - | 100 | - | - |
| Ipomoea sp._tin_3(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Anthurium sp._tin_3(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Piper semicordulatum(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Canna bangii(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Margaritopsis boliviana(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Schefflera pentandra(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Dryopteris patula(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 25 | - |
| Palicourea angustifolia(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Chelyocarpo ulei-Acacietum loretensis ass. nov. | ||||||||||||||||||||||||
| Acacia loretensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - |
| Chelyocarpus ulei(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - |
| Miconia aff cremophylla(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - |
| Selaginella haematodes(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 75 | - |
| Acalypha stricta(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 75 | - |
| Banara guianensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20 | - | 50 | - |
| Heliconia rostrate(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Corytoplectus speciosus(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Clitoria pozuzoensis(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Sloanea ptariana(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Thelypteris patens(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Perebea angustifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Miconia amplexicaulis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Bauhinia tarapotensis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Polybotrya caudata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Piper arboreum(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Paullinia serjaniifolia(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Boehmeria pavonii(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Tectaria antioquoiana(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Pachiro brevipedis-Euterpetum catingae | ||||||||||||||||||||||||
| Mauritiella aculeata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 |
| Caraipa utilis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 |
| Euterpe catinga(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 |
| Pachira brevipes(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 80 |
| Doliocarpus major(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 70 |
| Ocotea olivacea(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 |
| Clusia amazonica(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | 60 |
| Amaioua corymbosa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 60 |
| Cathedra acuminata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 |
| Chrysophyllum bombycinum(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 |
| Mauritia carana(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 |
| Lindsaea divaricata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 |
| Vantanea parviflora(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 30 |
| SUPRATROPICAL RAINFORESTS FROM SOUTHERN PERU WITH POLYLEPIS (POLYLEPIDETEA TARAPACANO-BESSERI) | ||||||||||||||||||||||||
| Bomarea ovata(C) | 100 | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Berberis peruviana(S) | 100 | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Peperomia hartwegiana(H) | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| MESO- SUPRA TROPICAL RAINFOREST WITH OREOPANAX (PRUNO RIGIDAE-OREOPANACETEA FLORIBUNDI), AND SHRUB FORMATIONS WITH BACCHARIS (CLEMATIDO PERUVIANAE-BACCHARITETEA LATIFOLIAE) | ||||||||||||||||||||||||
| Oreopanax eriocephalus(T) | - | 60 | - | - | 100 | - | - | 20 | 67 | 100 | 50 | 14 | - | - | - | 20 | - | - | - | - | - | - | - | - |
| Clematis haenkeana(C) | - | 40 | - | - | 14 | - | - | - | - | 80 | 25 | 43 | 50 | - | - | 20 | - | - | - | - | - | - | - | - |
| Hesperomeles ferruginea(T) | - | - | - | - | 100 | - | - | - | - | 60 | 50 | - | - | - | 13 | - | - | - | - | - | - | - | - | - |
| Myrcianthes myrsinoides(T) | - | - | - | - | - | - | - | 20 | 33 | 100 | 75 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Mauria heterophylla(S) | - | - | - | - | - | - | - | - | 33 | 80 | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Adiantum concinnum(H) | - | - | - | - | - | - | - | 20 | 67 | 60 | - | - | - | - | - | 20 | - | - | - | - | - | - | - | - |
| Oreocallis grandiflora(S) | - | - | - | - | - | - | - | 40 | - | 60 | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Vallea stipularis(S) | - | - | - | - | - | - | - | 20 | 33 | - | - | 29 | - | - | - | - | - | - | - | - | 20 | - | - | - |
| Baccharis latifolia(S) | - | - | - | - | - | - | - | - | - | - | 25 | 100 | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Berberis beauverdiana(S) | - | - | - | - | - | - | - | - | - | 100 | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Delostoma integrifolium(T) | - | - | - | - | - | - | - | 40 | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Solanum oblongifolium(S) | - | - | - | - | - | - | - | 40 | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Oreopanax aff cuspidatus(T) | - | - | - | - | - | - | - | 20 | 67 | 40 | 25 | - | - | - | - | - | - | 25 | - | - | - | - | - | - |
| Oxalis peduncularis(C) | - | - | - | - | - | - | - | - | 33 | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Salvia punctate(SS) | - | - | - | - | - | - | - | - | 33 | - | 75 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Mutisia acuminata(C) | - | - | - | - | - | - | - | - | 33 | - | 25 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lupinus mutabilis(S) | - | - | - | - | - | - | - | - | 67 | - | 50 | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Citharexylum flexuosum(S) | - | - | - | - | - | - | - | 60 | - | 80 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Monactis flaverioides(S) | - | - | - | - | - | - | - | - | 33 | 40 | - | 100 | 100 | - | 13 | - | - | - | - | - | - | - | - | - |
| Prunus rigida(T) | - | - | - | - | - | - | - | - | - | 80 | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Clusia pseudomangle(S) | - | - | - | - | - | - | - | 40 | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Fuchsia ayavacensis(S) | - | - | - | - | - | - | - | - | 40 | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Monnina pilosa(S) | - | - | - | - | - | - | - | 20 | 33 | 100 | 50 | - | - | - | - | 20 | - | - | - | - | - | - | - | - |
| Lepechinia mollis(S) | - | - | - | - | - | - | - | - | - | 40 | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Otholobium munyense(S) | - | 20 | - | - | - | - | - | - | - | - | 25 | 43 | 100 | - | - | - | - | - | - | - | - | - | - | - |
| Hedyosmum scabrum(T) | - | - | - | - | - | - | - | 40 | 33 | - | - | - | - | - | 13 | 20 | - | 25 | - | - | - | - | - | - |
| Miconia denticulate(S) | - | - | - | - | - | - | - | 100 | 67 | 80 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Ilex uniflora(T) | - | - | - | - | - | - | - | 60 | 67 | 80 | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| Aphelandra acanthifolia(S) | - | - | - | - | - | - | - | 40 | 33 | 60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cronquistianthus marrubiifolius(S) | - | - | - | - | - | - | - | 40 | 33 | 80 | 75 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Passiflora sagasteguii(C) | - | - | - | - | - | - | - | 60 | 33 | 80 | 75 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Clusia multiflora(T) | - | - | - | - | - | - | - | 40 | 67 | 80 | - | - | - | - | 25 | - | - | - | 33 | - | - | - | - | - |
| Myrsine latifolia(T) | - | - | - | - | - | - | - | 40 | 33 | 100 | - | - | - | - | - | 20 | 100 | - | - | - | - | - | - | - |
| Croton abutiloides(S) | - | - | - | - | - | - | - | 20 | - | 60 | - | - | - | - | - | 60 | - | - | - | - | - | - | - | - |
| Frangula sphaerosperma(S) | - | - | - | - | - | - | - | 20 | - | 40 | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - |
| THERMO-MESOTROPICAL FORESTS WITH ALNUS (ALNETEA ACUMINATAE) | ||||||||||||||||||||||||
| Alnus acuminata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | 20 | 100 | 25 | - | - | 20 | - | - | - |
| Pteris deflexa(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | 40 | - | - | - | - | - | - | - | - |
| Escallonia pendula(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 50 | 25 | - | - | - | - | - | - |
| Thelypteris pennata(H) | - | - | - | - | - | 17 | - | - | - | - | - | - | - | - | - | 60 | - | 50 | - | - | - | - | - | - |
| Pentacalia reflexa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 75 | - | - | - | - | - | - |
| Bia alienata(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 50 | - | - | - | - | - | - |
| THERMOTROPICAL RAINFOREST FROM SOUTHERN PERU (SERPOCAULO DASYPLEURONIS-ALCHORNEION LATIFOLIAE all. nov.) | ||||||||||||||||||||||||
| Lasiacis ligulata(H) | - | - | 71 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 25 | - |
| Alchornea latifolia(S) | - | - | 57 | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Psychotria poeppigiana(S) | - | - | 43 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Serpocaulon dasypleuron(H) | - | - | 80 | 71 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lindsaea arcuata(H) | - | 40 | - | 29 | - | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cyathea delgadii(T) | - | - | 86 | - | - | - | - | - | - | - | - | - | - | - | 13 | - | - | - | 67 | - | - | - | - | - |
| Olyra latifolia(H) | - | - | 86 | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Graffenrieda cucullata(T) | - | - | 86 | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | - | 67 | 100 | 40 | - | - | - |
| Trema micrantha(T) | - | - | - | 57 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| THERMO- INFRATROPICAL RAINFORESTS FROM NORTHERN PERU (PTERIDI CRETICAE-CYATHEION CARACASANAE all. nov.) | ||||||||||||||||||||||||
| Pteris cretica(H) | - | - | - | 14 | - | 67 | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Pennisetum peruvianum(H) | - | - | - | - | - | 67 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - |
| Cyrtocymura scorpioides(S) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Fuchsia mathewsii(S) | - | - | - | - | - | 33 | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Cecropia utcubambana(T) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | 17 | - | 100 | 50 | - |
| Seemannia sylvatica(H) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | 20 | - | - | - | 33 | - | - | - | - |
| Miconia adinantha(S) | - | - | - | - | - | 33 | - | 40 | - | - | - | - | - | - | - | - | - | 50 | - | - | - | - | - | - |
| Begonia peruviana(C) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | 13 | - | - | - | 67 | 67 | - | - | - | - |
| Piper lineatum(S) | - | - | 14 | - | - | 33 | - | - | - | - | - | - | - | - | - | 100 | - | 25 | - | - | 20 | - | - | - |
| MESO- TO INFRATROPICAL RAINFORESTS FROM CENTRAL PERU (SANCHEZIO OBLONGAE-HEDYOSMION RACEMOSAE all. nov.) | ||||||||||||||||||||||||
| Piper chanchamayanum(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 33 | - | - | - | - |
| Zeugites americanus(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | 17 | - | 100 | - | - |
| Miconia affinis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 83 | - | - | - | - |
| Sanchezia oblonga(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 83 | - | 25 | - | - |
| Geonoma stricta(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 50 | - | - | - | - |
| Dennstaedtia auriculata(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 50 | - | - | - | - |
| Gurania lobate(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 50 | - | - | - | - |
| Anthurium aff amoenum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 50 | - | - | - | - |
| Liabum nudicaule(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 50 | - | - | 25 | - |
| Blepharodon salicinus(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 33 | - | - | - | - |
| Weinmannia glomerata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | 80 | - | - | - |
| Baccharis decussata subsp. jelskii(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | 60 | - | - | - |
| Pilea haenkei(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 17 | - | 100 | - | - |
| Piper malifolium(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 17 | - | 100 | - | - |
| Passiflora rubra(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | 20 | - | 50 | - |
| Pteris podophylla(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 83 | - | - | 25 | - |
| Fuchsia ovalis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | 33 | - | - | - | - |
| Hedyosmum racemosum(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | 83 | - | - | - | - |
| Oreopanax polycephalus(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | 50 | - | - | - | - |
| Pteris quadriaurita(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | 20 | - | - | 33 | - | 40 | - | - | - |
| Anthurium breviscapum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | 100 | 20 | - | - | - |
| Monstera obliqua(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 17 | - | 100 | 25 | - |
| Clusia trochiformis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 10 | 40 | 100 | - | - |
| Miconia cyanocarpa(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 80 | 100 | - | - |
| Monnina marginata(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | 50 | - |
| Niphidium albopunctatissimum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | 100 | - | - |
| Dryopteris wallichiana(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | - | - | 20 | 100 | - | - |
| Cedrela fissilis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | 25 | - |
| Palicourea guianensis(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | - | - | 50 | - |
| Iresine diffusa(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 33 | 20 | - | 50 | - |
| Diplazium tungurahuae(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 33 | - | - | 25 | - |
| Inga semialata(T) | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25 | 60 |
| Piper aduncum(S) | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | - |
| Inga edulis(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 75 | - |
| MESO- TO INFRATROPICAL MOUNTAIN RAINFORESTS FROM PERU (MORELLO PUBESCENTIS-MYRSINETEA CORIACEAE cl. nov.) | ||||||||||||||||||||||||
| Niphidium crassifolium(H) | - | 60 | 29 | 14 | - | - | - | 20 | - | 20 | - | - | 50 | - | 50 | 20 | 100 | 50 | 67 | 33 | 40 | - | - | - |
| Blechnum occidentale(H) | - | - | - | - | - | - | - | 100 | 67 | 60 | - | - | - | - | - | 40 | 50 | 75 | 100 | 50 | 80 | 100 | 50 | - |
| Pteridium arachnoideum(H) | - | - | - | 14 | 33 | 83 | - | 60 | - | 80 | - | - | - | - | 13 | 40 | - | 25 | - | - | 20 | - | - | - |
| Baccharis inamoena(S) | - | 80 | - | - | 100 | 67 | 100 | - | - | - | - | - | - | - | 13 | 20 | 100 | 100 | - | - | - | - | - | |
| Ageratina sternbergiana(S) | - | 40 | - | - | - | - | - | 40 | 33 | 60 | 100 | 43 | - | - | 25 | - | 50 | - | - | - | - | - | - | |
| Myriocarpa stipitata(T) | - | 20 | - | 29 | - | 17 | 100 | - | - | - | - | - | - | - | - | 80 | - | - | 67 | - | - | - | 50 | - |
| Tradescantia cymbispatha(H) | - | 40 | - | 43 | - | 17 | - | 20 | - | - | - | - | - | - | 13 | 40 | - | 50 | - | - | - | - | - | - |
| Condaminea corymbosa(S) | - | - | - | 14 | - | 50 | - | - | - | - | - | - | - | - | 50 | - | - | - | 33 | 20 | 100 | 75 | - | - |
| Ctenitis sloanei(H) | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - | 63 | 20 | - | - | 100 | 17 | 80 | - | 50 | - |
| Solanum maturecalvans(S) | - | 60 | - | 14 | 67 | - | - | 40 | 33 | - | - | - | - | - | - | - | - | - | 33 | - | - | - | - | - |
| Saurauia peruviana(T) | - | - | - | 29 | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | 33 | 50 | 20 | 100 | - | - |
| Cavendishia bracteata(S) | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - | 25 | - | - | - | 33 | 17 | 20 | - | - | - |
| Serpocaulon caceresii(H) | - | 80 | 57 | 14 | - | 17 | - | - | - | - | - | - | - | - | - | - | - | - | - | 17 | - | - | - | - |
| Siparuna aspera(S) | - | 20 | 29 | - | - | 83 | - | - | - | - | - | - | - | - | - | - | - | 75 | - | - | - | - | 25 | - |
| Conostegia inusitata(T) | - | 20 | - | 43 | - | - | - | - | - | - | - | - | - | - | 38 | 40 | - | 75 | - | - | - | - | - | - |
| Alchornea glandulosa(T) | - | - | 43 | - | - | 17 | - | - | - | - | - | - | - | - | - | 50 | 20 | - | - | - | 17 | - | - | - |
| Palicourea amethystine(S) | - | - | - | - | - | - | - | 60 | - | - | - | - | - | - | 13 | - | - | - | 67 | 50 | 40 | - | - | - |
| Myrsine coriacea(T) | - | - | - | - | - | - | - | - | - | 60 | - | 14 | - | - | - | - | - | 75 | 33 | 83 | - | - | - | - |
| Morella pubescens(T) | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | 33 | 33 | 60 | - | - | - |
| Cyathea caracasana(T) | - | - | - | - | - | 33 | - | - | - | - | - | - | - | - | - | - | - | - | 33 | 83 | 20 | - | 25 | - |
| Munnozia hastifolia(S) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 50 | 20 | - | - | 33 | 17 | - | - | 25 | - |
| Nephrolepis cordifolia(H) | - | - | 29 | - | - | 33 | - | - | - | - | - | - | - | - | - | 60 | - | - | - | - | - | - | 50 | - |
| Heliocarpus americanus(T) | - | - | - | - | - | 67 | - | - | - | - | - | - | - | - | 13 | - | - | - | - | 100 | - | 100 | 25 | - |
| Cinchona pubescens(T) | - | - | - | - | - | 67 | - | - | - | - | - | - | - | - | 50 | - | - | - | - | 17 | - | 100 | - | - |
| Ipomoea sp.(C) | - | 20 | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | 25 | 67 | - | - | - | - | - |
| Vismia tomentosa(T) | - | - | 71 | 14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 67 | - | - | - | - |
| Begonia cyathophora(H) | - | - | - | - | 100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | 25 | - |
| Smilax domingensis(C) | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | 50 | - | - | - | 100 | - | - |
| Blechnum cordatum(H) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 13 | - | - | - | 33 | - | 40 | - | - | - |
| Disterigma alaternoides(SS) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 13 | - | - | - | 33 | - | 20 | - | - | - |
| Schefflera acuminata(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 38 | - | - | - | 33 | 50 | 20 | - | - | - |
| Ochroma pyramidale(T) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - | - | - | 100 | 100 | - |
| Festuca sp.(H) | - | - | - | - | 67 | - | - | - | - | - | 50 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Dioscorea altissima(C) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 80 | - | - | - | - | - | - | - | 60 |
| Polystichum montevidense(H) | - | 40 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 40 | - | - | - |
| Association Number in Text | 2 | 3 | 4 | 5 | 6 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Vegetation canopy (m) | 10 | 25 | 20 | 8 | 20 | 28 | 20 | 15 | 34 | 20 | 25 | 10 | 26 |
| Slope (%) | 60 | 0 | 70 | 0 | 40 | 20 | 20 | 20 | 0 | 30 | 10 | 50 | 40 |
| Altitude (m) | 2509 | 791 | 1230 | 2774 | 1689 | 2664 | 1783 | 2337 | 2378 | 2358 | 1351 | 2533 | 894 |
| Orientation | SE | - | E | - | NW | S | SW | S | S | S | E | W | NE |
| Area (m2) | 200 | 200 | 100 | 100 | 200 | 500 | 500 | 100 | 500 | 200 | 1000 | 400 | 500 |
| Smallantho parvicipitis-Oreopanacetum eriocephali ass. nov. | |||||||||||||
| Ageratina sternbergiana(S) | + | - | - | - | - | - | - | 1 | - | - | - | - | - |
| Oreopanax eriocephalus(T) | 1 | - | - | 1 | - | - | - | - | - | - | - | - | - |
| Tradescantia cymbispatha(H) | 2 | - | - | - | - | - | 1 | - | 2 | - | - | - | - |
| Serpocaulon caceresii(H) | 2 | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Solanum maturecalvans(S) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Miconia aff adinantha(S) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Serpocaulon dasypleuron(H) | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Begonia bracteosa(H) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Smallanthus parviceps(T) | 4 | - | - | - | - | - | - | - | - | - | - | - | - |
| Liabum solidagineum(S) | 2 | - | - | - | - | - | - | - | - | - | - | - | - |
| Fuchsia boliviana(S) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Melica scabra(H) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Rubus boliviensis(S) | + | - | - | - | - | - | - | - | - | - | - | - | - |
| Solanum aphyodendron(T) | 3 | - | - | - | - | - | - | - | - | - | - | - | - |
| Phenax angustifolius(T) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Croton regelianus(S) | + | - | - | - | - | - | - | - | - | - | - | - | - |
| Pityrogramma calomelanos(H) | + | - | - | - | - | - | - | - | - | - | - | - | - |
| Polystichum montevidense(H) | 2 | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Pennisetum latifolium(H) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Lozanella enantiophylla(T) | 3 | - | - | - | - | - | - | - | - | - | - | - | - |
| Equisetum sp._sg1(H) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Melastomataceae 01_sg1(S) | 3 | - | - | - | - | - | - | - | - | - | - | - | - |
| Galium hypocarpium(H) | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Iriartello setigerae-Cinchonetum micranthae ass. nov. | |||||||||||||
| Siparuna aspera(S) | - | 1 | - | - | 1 | - | - | - | 3 | - | - | - | 1 |
| Olyra latifolia(H) | - | 2 | - | - | 1 | - | - | - | - | - | - | - | - |
| Alchornea glandulosa(T) | - | 2 | - | - | - | - | 1 | - | - | - | - | - | - |
| Graffenrieda cucullata(T) | - | 3 | - | - | - | - | - | - | - | 4 | 3 | 2 | - |
| Cyathea delgadii(T) | - | 3 | - | - | - | - | - | - | - | 2 | - | - | - |
| Alchornea latifolia(S) | - | 2 | - | - | - | - | - | - | - | - | - | - | - |
| Cinchona micrantha(T) | - | 4 | - | - | - | - | - | - | - | - | - | - | - |
| Iriartella setigera(S) | - | 2 | - | - | - | - | - | - | - | - | - | - | - |
| Heliconia hirsute(H) | - | + | - | - | - | - | - | - | - | - | - | - | - |
| Vismia tomentosa(T) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Cyathea subincisa(T) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Philodendron rudgeanum(Ep) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Dieffenbachia humilis(H) | - | + | - | - | - | - | - | - | - | - | - | - | - |
| Miconia lourteigiana(T) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Aphelandra aurantiaca(S) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Blakea sawadae(C) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Palicourea fastigiata(S) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Liabum acuminatum(S) | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Acalypho macrostachyae-Cecropietum polystachyae ass. nov. | |||||||||||||
| Saurauia peruviana(T) | - | - | 1 | - | - | - | - | - | - | - | 2 | 1 | - |
| Cecropia polystachya(T) | - | - | 4 | - | - | - | - | - | - | - | - | - | + |
| Chusquea scandens(S) | - | - | 1 | - | - | 4 | 1 | - | 1 | - | - | 4 | - |
| Pteris cretica(H) | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| Morella pubescens(T) | - | - | 2 | - | - | - | - | - | - | - | + | - | - |
| Trema micrantha(T) | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| Anthurium grande(H) | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| Acalypha macrostachya(T) | - | - | 4 | - | - | - | - | - | - | - | - | - | - |
| Urera caracasana(T) | - | - | 2 | - | - | - | - | - | - | - | - | - | - |
| Lycianthes inaequilatera(S) | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| Bignoniaceae(C) | - | - | + | - | - | - | + | - | - | - | 1 | - | - |
| Psychotria trichostoma(S) | - | - | 3 | - | - | - | - | - | - | - | - | - | - |
| Philodendron divaricatum(C) | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| Peperomia pseudorhynchophoros(H) | - | - | 2 | - | - | - | - | - | - | - | - | - | - |
| Thelypteris brausei(H) | - | - | + | - | - | - | - | - | - | - | - | - | - |
| Crotono churumayensis-Hesperomeletum ferrugineae ass. nov. | |||||||||||||
| Rubus praecox(S) | - | - | - | 1 | - | - | - | - | 1 | - | - | - | - |
| Baccharis inamoena(S) | - | - | - | 4 | 2 | + | - | 3 | 2 | - | - | - | - |
| Hesperomeles ferruginea(T) | - | - | - | 2 | - | 1 | - | - | - | - | - | - | - |
| Festuca sp.(H) | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| Cuphea cordata(S) | - | - | - | 3 | - | - | - | - | - | - | - | - | - |
| Croton churumayensis(T) | - | - | - | 3 | - | - | - | - | - | - | - | - | - |
| Begonia cyathophora(H) | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| Senna birostris(S) | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| Ditassa sp._sg(Ep) | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| Peperomia sp._sg158(H) | - | - | - | 1 | - | - | - | - | - | - | - | - | - |
| Viburno reticulati-Weinmannietum spruceanae ass. nov. | |||||||||||||
| Pteridium arachnoideum(H) | - | - | - | - | 2 | - | - | - | - | - | - | - | - |
| Lindsaea arcuata(H) | - | - | - | - | 2 | - | - | - | - | - | - | - | - |
| Condaminea corymbosa(S) | - | - | - | - | 1 | - | - | - | - | - | - | - | 3 |
| Nephrolepis cordifolia(H) | - | - | - | - | 1 | - | 1 | - | - | - | - | - | 2 |
| Viburnum reticulatum(T) | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| Weinmannia spruceana(T) | - | - | - | - | 4 | - | - | - | - | - | - | - | - |
| Fuchsia mathewsii(S) | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| Seemannia sylvatica(H) | - | - | - | - | + | - | - | - | - | - | 1 | - | - |
| Pouzolzia poeppigiana(T) | - | - | - | - | + | - | - | - | - | - | - | - | - |
| Clusia crenata(T) | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| Pitcairnia paniculate(H) | - | - | - | - | + | - | - | - | - | - | - | - | - |
| Anthurium hamiltonii(H) | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| Elaphoglossum aff andicola(H) | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| Myrsine manglilla(S) | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| Hypolepido parallelogrammae-Alnetum acuminatae ass. nov. | |||||||||||||
| Niphidium crassifolium(H) | - | - | - | - | - | 1 | - | + | + | + | + | 1 | - |
| Munnozia hastifolia(S) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Alnus acuminata(T) | - | - | - | - | - | 3 | - | 4 | + | - | - | 1 | - |
| Ctenitis sloanei(H) | - | - | - | - | - | 1 | - | - | - | 3 | - | 2 | 2 |
| Hypolepis parallelogramma(H) | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Solanum sp. 03(S) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Hypolepis obtusata(H) | - | - | - | - | - | 2 | - | - | - | - | - | - | - |
| Cyathea lechleri(T) | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Aulonemia longiaristata(H) | - | - | - | - | - | 2 | - | - | - | - | - | - | - |
| Viburnum incarum(S) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Cronquistianthus sp. 01_cut(S) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Escallonia myrtilloides(S) | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Weinmannia cymbifolia(S) | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Ageratina neblinense(S) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Clusia ducu(T) | - | - | - | - | - | 2 | - | - | - | - | - | - | - |
| Psidium sp._cut7(T) | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Rubiaceae sp. 01_cut7(S) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Tree shiny lanceolate leaves_cut7(T) | - | - | - | - | - | + | - | - | - | - | - | - | - |
| Desfontainia spinosa(S) | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Hedyosmum translucidum(T) | - | - | - | - | - | 3 | - | - | - | - | - | - | - |
| Pleurothallis cordata(Ep) | - | - | - | - | - | 3 | - | - | - | - | - | - | - |
| Lueheo divaricatae-Ceroxylonetum peruviani ass. nov. | |||||||||||||
| Dioscorea altissima(C) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Piper lineatum(S) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Adiantum concinnum(H) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Escallonia pendula(T) | - | - | - | - | - | - | 4 | 1 | - | - | - | - | - |
| Pteris deflexa(H) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Luehea divaricata(T) | - | - | - | - | - | - | 3 | - | - | - | - | - | - |
| Serpocaulon fraxinifolium(H) | - | - | - | - | - | - | + | - | - | - | - | - | - |
| Styloceras laurifolium(S) | - | - | - | - | - | - | 3 | - | - | - | - | - | - |
| Pteris quadriaurita(H) | - | - | - | - | - | - | 1 | - | - | 1 | - | 2 | - |
| Asplenium cuneatum(H) | - | - | - | - | - | - | + | - | - | - | - | - | - |
| Asplenium alatum(H) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Lycianthes sp._got5(S) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Miconia sp. 01_got5(S) | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Sclerophyllous tree broad leaves_got5(T) | - | - | - | - | - | - | 2 | - | - | - | - | - | - |
| Ceroxylon peruvianum(T) | - | - | - | - | - | - | 3 | - | - | - | - | - | - |
| Mutisio wurdackii-Alnetum acuminatae ass. nov. | |||||||||||||
| Blechnum occidentale(H) | - | - | - | - | - | - | - | 2 | 2 | 1 | - | 1 | - |
| Myrsine latifolia(T) | - | - | - | - | - | - | - | 2 | - | - | - | - | - |
| Pentacalia reflexa(S) | - | - | - | - | - | - | - | + | - | - | - | - | - |
| Bia alienate(C) | - | - | - | - | - | - | - | 1 | - | - | - | - | - |
| Mutisia wurdackii(C) | - | - | - | - | - | - | - | 2 | 1 | - | - | - | - |
| Weinmannia costulata(S) | - | - | - | - | - | - | - | 1 | - | - | - | - | - |
| Cortaderia jubata(H) | - | - | - | - | - | - | - | + | - | - | - | - | - |
| Axinaeo tomentosae-Ceroxylonetum quindiuensis ass. nov. | |||||||||||||
| Conostegia inusitata(T) | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Myrsine coriacea(T) | - | - | - | - | - | - | - | - | 1 | - | 2 | - | - |
| Cedrela odorata(T) | - | - | - | - | - | - | - | - | 3 | - | - | - | - |
| Miconia adinantha(S) | - | - | - | - | - | - | - | - | 2 | - | - | - | - |
| Cronquistianthus chachapoyensis(S) | - | - | - | - | - | - | - | - | + | - | - | - | - |
| Ceroxylon quindiuense(T) | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Solanum barbulatum(S) | - | - | - | - | - | - | - | - | + | - | - | - | - |
| Axinaea tomentosa(S) | - | - | - | - | - | - | - | - | + | - | - | - | - |
| Vismia pozuzoensis(T) | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Hedyosmum lechleri(T) | - | - | - | - | - | - | - | - | 4 | - | - | - | - |
| Palicourea stipularis(S) | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Begonia sp._ocol13(H) | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Dryopteris wallichiana(H) | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Hydrangea jelskii(S) | - | - | - | - | - | - | - | - | 2 | - | - | - | - |
| Piperi chanchamayani-Cecropietum strigosae ass. nov. | |||||||||||||
| Cecropia angustifolia(T) | - | - | - | - | - | - | - | - | - | 1 | - | - | - |
| Cyathea caracasana(T) | - | - | - | - | - | - | - | - | - | 1 | 1 | - | 1 |
| Anthurium breviscapum(H) | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - |
| Diplazium tungurahuae(H) | - | - | - | - | - | - | - | - | - | 1 | - | - | 1 |
| Iresine diffusa(C) | - | - | - | - | - | - | - | - | - | 1 | - | 1 | + |
| Cecropia strigose(T) | - | - | - | - | - | - | - | - | - | 3 | - | - | - |
| Pteris podophylla(H) | - | - | - | - | - | - | - | - | - | 1 | 2 | - | - |
| Centropogon altus(S) | - | - | - | - | - | - | - | - | - | 2 | - | - | - |
| Pennisetum aff peruvianum(H) | - | - | - | - | - | - | - | - | - | 2 | - | - | - |
| Piper chanchamayanum(S) | - | - | - | - | - | - | - | - | - | 1 | - | - | - |
| Clusia aff pallida(T) | - | - | - | - | - | - | - | - | - | 1 | - | - | - |
| Ageratina tambillensis(S) | - | - | - | - | - | - | - | - | - | + | - | - | - |
| Dennstaedtia auriculata(H) | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - |
| Liabum nudicaule(S) | - | - | - | - | - | - | - | - | - | 1 | - | - | + |
| Miconia sp. 08_sat(S) | - | - | - | - | - | - | - | - | - | + | - | - | - |
| Pilea haenkei(S) | - | - | - | - | - | - | - | - | - | 1 | + | - | - |
| Piper malifolium(T) | - | - | - | - | - | - | - | - | - | 1 | - | - | - |
| Adianthum raddianum(H) | - | - | - | - | - | - | - | - | - | + | - | - | - |
| Geonoma stricta(S) | - | - | - | - | - | - | - | - | - | 1 | + | - | - |
| Gurania lobata(C) | - | - | - | - | - | - | - | - | - | 1 | - | - | - |
| Palicourea amethystina(S) | - | - | - | - | - | - | - | - | - | + | - | - | - |
| Cedrelo fissilis-Ficetum maximae ass. nov. | |||||||||||||
| Heliocarpus americanus(T) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Begonia peruviana(C) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Cavendishia bracteata(S) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Serpocaulon loriceum(H) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Schefflera acuminata(T) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Begonia parviflora(C) | - | - | - | - | - | - | - | - | - | - | + | 1 | - |
| Heliconia subulate(H) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Vernonanthura patens(S) | - | - | - | - | - | - | - | - | - | - | 2 | - | - |
| Lastreopsis effuse(H) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Hedyosmum racemosum(T) | - | - | - | - | - | - | - | - | - | - | 4 | - | - |
| Cedrela fissilis(T) | - | - | - | - | - | - | - | - | - | - | 1 | - | 3 |
| Blepharodon salicinus(C) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Oreopanax polycephalus(T) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Miconia affinis(T) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Anthurium aff amoenum(H) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Sanchezia oblonga(S) | - | - | - | - | - | - | - | - | - | - | 3 | - | - |
| Ficus maxima(T) | - | - | - | - | - | - | - | - | - | - | 4 | - | - |
| Dieffenbachia sp. 02_sat(H) | - | - | - | - | - | - | - | - | - | - | 2 | - | - |
| Alsophila mostellaria(T) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Ocotea obovata(T) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Dioscorea sp. 01_sat(C) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Cyclanthus bipartitus(H) | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 |
| Croton lechheri(T) | - | - | - | - | - | - | - | - | - | - | 2 | - | - |
| Palicourea guianensis(S) | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 |
| Columnea inaquilatera(H) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Elaphoglossum latifolium(H) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Megalastrum pulverulentum(H) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Ficus cuatrecasasiana(T) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Melastomataceae 02_sat(S) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Siparuna tomentosa(S) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Sorocea sp._sat8(S) | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Clusia aff tarmensis(T) | - | - | - | - | - | - | - | - | - | - | + | - | - |
| Cecropio albicantis-Weinmannietum glomeratae ass. nov. | |||||||||||||
| Prestoea carderi(S) | - | - | - | - | - | - | - | - | - | - | 1 | 2 | - |
| Blechnum cordatum(H) | - | - | - | - | - | - | - | - | - | - | - | 2 | - |
| Weinmannia latifolia(T) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Disterigma alaternoides(SS) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Piper augustum(S) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Disterigma acuminatum(SS) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Weinmannia glomerata(T) | - | - | - | - | - | - | - | - | - | - | - | 4 | - |
| Anthurium linganii(H) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Bromelia sp._tin_1(H) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Clidemia sp._tin_1(S) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Munnozia silphioides(H) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Miconia cyanocarpa(S) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Tibouchina saxosa(SS) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Centropogon reflexus(C) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Centropogon hirtus(S) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Fuchsia macrophylla(S) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Oyedaea sp._tin_1(S) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Cecropia albicans(T) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Bocconia integrifolia(T) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Oxalis dolichopoda(C) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Clusiaceae_tin_1(S) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Weinmannia auriculata(T) | - | - | - | - | - | - | - | - | - | - | - | 4 | - |
| Miconia monzoniensis(S) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Dilleniaceae_tin_1(S) | - | - | - | - | - | - | - | - | - | - | - | + | - |
| Clusia trochiformis(S) | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Chelyocarpo ulei-Acacietum loretensis ass. nov. | |||||||||||||
| Myriocarpa stipitata(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Clusia amazonica(T) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Heliconia hirsute(H) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Smilax purhampuy(C) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Ochroma pyramidale(T) | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Ladenbergia oblongifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Urera baccifera(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Dichorisandra hexandra(C) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Casearia fasciculata(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Boehmeria pavonii(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Tectaria antioquoiana(H) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Monstera obliqua(H) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Serpocaulon adnatum(H) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Polybotrya caudata(H) | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Chelyocarpus ulei(S) | - | - | - | - | - | - | - | - | - | - | - | - | 3 |
| Selaginella haematodes(H) | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Corytoplectus speciosus(H) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Cissus sp._tin_4(C) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Anthurium sp._tin_4(H) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Acacia loretensis(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Cecropia aff obtusifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Acalypha stricta(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Sloanea ptariana(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Justicia poeppigiana(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Rubiaceae_tin_4(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Bauhinia tarapotensis(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Perebea angustifolia(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Psychotria_tin_4(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Paullinia serjaniifolia(C) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Solanum sp._tin_4(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Miconia aff abbreviate(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Liabum eriocaulon(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Gurania rhizantha(C) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Senna multijuga(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Inga chartacea(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Miconia amplexicaulis(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Monnina marginata(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Gloxinia xanthophylla(H) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Miconia aff cremophylla(S) | - | - | - | - | - | - | - | - | - | - | - | - | 3 |
| Viburnum hallii(S) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Gunnera sp._tin_4(H) | - | - | - | - | - | - | - | - | - | - | - | - | + |
| Gouania aptera(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Ficus americana(T) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Gonzalagunia bunchosioides(S) | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
References
- Sclultz, J. The Ecozones of the World. In The Ecological Divisions of the Geosphere; Springer: Berlin, Germany, 1995. [Google Scholar]
- Galán de Mera, A.; Vicente Orellana, J.A. Cronosequences of vegetation—A bioclimatic theory for interpreting the patterns of relic vegetation types. Phytocoenologia 2007, 37, 471–494. [Google Scholar] [CrossRef]
- Gentry, A.H. Patterns of diversity and floristic composition in neotropical montane forests. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; The New York Botanical Garden: New York, NY, USA, 1995; pp. 103–126. [Google Scholar]
- Distler, T.; Jørgensen, P.M.; Graham, A.; Davidse, G.; Jiménez, I. Determinants and prediction of broad-scale plant richness across the western Neotropics. Ann. Missouri Bot. Gard. 2009, 96, 470–491. [Google Scholar] [CrossRef]
- Cabrera, A.L.; Willink, A. Biogeografía de América Latina; Organización de Estados Americanos: Washington, DC, USA, 1980. [Google Scholar]
- Seibert, P. Farbatlas Südamerika. Landschaften und Vegetation; Ulmer: Stuttgart, Germany, 1996. [Google Scholar]
- Rangel, O.; Lowy, P.D.; Aguilar, M.; Garzón, A. Tipos de vegetación en Colombia. Colomb. Divers. Biótica 1997, 2, 89–381. [Google Scholar]
- Weberbauer, A. The phytogeography of the Peruvian Andes. In Flora of Peru; MacBride, J.F., Ed.; Field Museum of Natural History (Botany): Chicago, IL, USA, 1936; Volume 13, pp. 13–81. [Google Scholar]
- Weberbauer, A. El Mundo Vegetal de los Andes Peruanos; Ministerio de Agricultura: Lima, Perú, 1945.
- Hueck, K. Die Wälder Südamerikas. Ökologie, Zusammensetzung und Wirtschaftliche Bedeutung; Gustav Fischer: Sttutgart, Germany, 1966. [Google Scholar]
- Hueck, K.; Seibert, P. Vegetationskarte von Südamerika; Gustav Fischer: Stuttgart, Germany, 1981. [Google Scholar]
- Egg, B.A.; Vargas, M.C. Ecología del Perú; Bruño: Lima, Perú, 2004. [Google Scholar]
- Vázquez, R.; Rojas, R.; Monteagudo, A.; Meza, K.; Van der Werff, H.; Ortiz-Gentry, R.; Catchpole, D. Flora vascular de la selva central del Perú: Una aproximación de la composición florística de tres Áreas Naturales Protegidas. Arnaldoa 2005, 12, 112–125. [Google Scholar]
- Young, K.R.; León, B.; Jørgensen, P.M.; Ulloa, U.C. Tropical and subtropical landscapes of the andes. In The Physical Geography of South America; Veblen, T.T., Young, K.R., Orme, A.R., Eds.; Oxford University Press: New York, NY, USA, 2007; pp. 200–216. [Google Scholar]
- Malizia, A.; Blundo, C.; Carilla, J.; Osinaga, O.; Cuesta, F.; Duque, A.; Aguirre, N.; Aguirre, Z.; Ataroff, M.; Baez, S.; et al. Elevation and latitude drives structure and tree species composition in Andean forests: Results from a large-scale plot network. PLoS ONE 2020, 15, e0231553. [Google Scholar] [CrossRef]
- Dodson, C.H.; Gentry, A.H. Biological extinction in Western Ecuador. Ann. Missouri Bot. Gard. 1991, 78, 273–295. [Google Scholar] [CrossRef]
- Sánchez Vega, I.; Grados Quezada, N. Estudio florístico y ambiental del bosque relicto Mijal en la Provincia de Morropón, Piura. Arnaldoa 2007, 14, 259–268. [Google Scholar]
- Galán de Mera, A.; Sánchez Vega, I.; Montoya Quino, J.; Linares Perea, E.; Campos de la Cruz, J.; Vicente Orellana, J.A. La vegetación del Norte del Perú: De los bosques a la jalca en Cajamarca. Acta Bot. Malac. 2015, 40, 157–190. [Google Scholar] [CrossRef]
- Critical Ecosystem. Hotspot de Biodiversidad de los Andes Tropicales. Available online: andestropicales.net (accessed on 8 July 2020).
- Vásquez, P.; Tovar, A.; Young, C.; Soto, A.; Tovar, C.; Saito, J.; Regal, F.; Rivera, G.; Veliz, C.; Gushiken, E.; et al. Planificación Para la Conservación Eco-Regional de las Yungas Peruanas: Conservando la Diversidad Natural de la Selva Alta del Perú; Universidad Nacional Agraria La Molina-The Natural Conservancy: Lima, Perú, 2008. [Google Scholar]
- Hoorn, C.F.P.; Wesselingh, H.; ter Steege, M.A.; Bermudez, A.; Mora, J.; Sevink, I.; Sanmartin, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia through time: Andean uplift climate change. landscape evolution and biodiversity. Science 2010, 30, 927–931. [Google Scholar] [CrossRef]
- Rowley, D.B.; Garzione, C.N. Stable isotope-based palaeoaltimetry. Annu. Rev. Earth Planet. Sci. 2007, 35, 463–508. [Google Scholar] [CrossRef]
- Garzione, C.N.; Hoke, G.D.; Libarkin, J.C.; Withers, S.; MacFadden, B.; Eiler, J.; Ghosh, P.; Mulch, A. Rise of the Andes. Science 2008, 320, 1304–1307. [Google Scholar] [CrossRef]
- Ibaraki, M. Closing of the Central American Seaway and Neogene coastal upwelling along the Pacific coast of South America. Tectophysics 1997, 281, 99–104. [Google Scholar] [CrossRef]
- Gentry, A.H. Neotropical floristic diversity: Phytogeographical connections between Central and South America, pleistocene climatic fluctuations, or an accident of the Andean orogeny. Ann. Missouri Bot. Gard. 1982, 69, 557–593. [Google Scholar] [CrossRef]
- Walter, H. Zonas de Vegetación y Clima; Omega: Barcelona, Spain, 1997. [Google Scholar]
- Rivas-Martínez, S.; Navarro, G.; Penas, A.; Costa, M. Biogeographic map of South America. A preliminary survey. Int. J. Geobot. Res. 2011, 1, 21–40. [Google Scholar] [CrossRef]
- Takhtajan, A. Floristic Regions of the World; University of California Press: Berkeley, CA, USA, 1986. [Google Scholar]
- Morrone, J.J. Biogeografía de América Latina y el Caribe; M & T-Manuales & Tesis SEA: Zaragoza, Spain, 2001. [Google Scholar]
- Braun-Blanquet, J. Plant Sociology. The Study of Plant Communities; Koeltz Scientific Books: Koenigstein, Germany, 1932. [Google Scholar]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006.
- Walker, D.A.; Kuss, P.; Epstein, H.E.; Kade, A.N.; Vonlanthen, C.M.; Raynolds, M.K.; Daniëls, F.J.A. Vegetation of zonal patterned-ground ecosystems along the North America Arctic bioclimatic gradient. Appl. Veg. Sci. 2011, 14, 440–463. [Google Scholar] [CrossRef]
- Peinado, M.; Ocaña-Peinado, F.M.; Aguirre, J.L.; Delgadillo, J.; Macías, M.A.; Díaz-Santiago, G. A phytosociological and phytogeographical survey of the coastal vegetation of western North America: Beach and dune vegetation from Baja California to Alaska. Appl. Veg. Sci. 2011, 14, 464–484. [Google Scholar] [CrossRef]
- Li, C.F.; Zelený, D.; Chytrý, M.; Chen, M.Y.; Chen, T.Y.; Chiou, C.R.; Hsia, Y.J.; Liu, H.Y.; Yang, S.Z.; Yeh, C.L.; et al. Chamaecyparis montane cloud forest in Taiwan: Ecology and vegetation classification. Ecol. Res. 2015, 30, 771–791. [Google Scholar] [CrossRef]
- Rodríguez, E.E.; Aceñolaza, P.G.; Linares Perea, E.; Galán de Mera, A. A phytosociological analysis of Butia yatay (Arecaceae) palm groves and gallery forests in Entre Rios, Argentina. Aust. J. Bot. 2017, 65, 183–202. [Google Scholar] [CrossRef]
- Ortiz, C.A.; Musarella, C.M.; Gomes, P.C.J.; Canas, Q.R.; Fuentes, P.J.C.; Cano, E. Phytosociological study, diversity and conservation status of the cloud forest in the Dominican Republic. Plants 2020, 9, 741. [Google Scholar] [CrossRef]
- Vareschi, V. Sobre las superficies de asimilación de sociedades vegetales en cordilleras tropicales y extratropicales. Bol. Soc. Venez. Ci. Nat. 1953, 79, 121–173. [Google Scholar]
- Meier, W. Flora und Vegetation des Ávila-Nationalparks (Venezuela, Küstenkordillera) unter besonderer Berücksichtigung der Nebelwaldstufe. Diss. Bot. 1998, 296, 1–485. [Google Scholar]
- Cuello, N.L.; Cleef, A.M. The forest vegetation of Ramal de Guaramacal in the Venezuelan Andes. Phytocoenologia 2009, 39, 109–156. [Google Scholar] [CrossRef]
- Cuatrecasas, J. Observaciones geobotánicas en Colombia. Trab. Mus. Nac. Ci. Nat., Ser. Bot. 1934, 27, 5–144. [Google Scholar]
- Franco, P.; Rangel, O.; Lozano, G. Estudios ecológicos en la Cordillera Oriental, II. Las comunidades vegetales de los alrededores de la laguna de Chingaza (Cundinamarca). Caldasia 1986, 15, 71–75. [Google Scholar] [CrossRef]
- Cleef, A.M.; Rangel, J.O.; Salamanca, S. The Andean rainforest of the Parque Los Nevados Transect, Cordillera Central, Colombia. Estud. Ecosistemas Tropandinos 2003, 5, 79–141. [Google Scholar]
- Rangel, O.; Cleef, A.M.; Salamanca, S.; Ariza, C. La vegetación de los bosques y selvas del Tatamá. Estud. Ecosistemas Tropandinos 2005, 6, 459–643. [Google Scholar]
- Rangel, O.; Cleef, A.M.; Arellano, H. La vegetación de los bosques y selvas del transecto Sumapaz. Estud. Ecosistemas Tropandinos 2008, 7, 695–797. [Google Scholar]
- Lauer, W.; Rafiqpoor, M.D.; Theisen, I. Physiogeographie, Vegetation und Syntaxonomie der Flora des Páramo de Papallacta (Ostkordillere Ecuador); Franz Steiner: Stuttgart, Germany, 2001. [Google Scholar]
- Bussmann, R.W. The vegetation of Reserva Biológica San Francisco, Zamora-Chinchipe, Southern Ecuador—A phytosociological synthesis. Lyonia 2003, 3, 145–254. [Google Scholar]
- Navarro, G.; Maldonado, M. Geografía Ecológica de Bolivia. Vegetación y Ambientes Acuáticos; Fundación Simón I. Patiño: Santa Cruz, Bolivia, 2002. [Google Scholar]
- Aceñolaza, P.G. Fitosociología de bosques de aliso (Alnus acuminata HBK subsp. acuminata) en la sierra de San Javier, Tucumán, Argentina. Doc. Phytosoc. 1996, 16, 505–516. [Google Scholar]
- Giusti, L.; Slanis, A.; Aceñolaza, P. Fitosociología de los bosques de aliso (Alnus acuminata H.B.K. ssp. acuminata) de Tucumán (Argentina). Lilloa 1997, 38, 93–120. [Google Scholar]
- Entrocassi, G.S.; Gavilán, R.G.; Sánchez-Mata, D. Subtropical Mountain Forests of Las Yungas: Vegetation and Bioclimate; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Galán de Mera, A.; Vicente Orellana, J.A.; Lucas García, J.A.; Probanza Lobo, A. Phytogeographical sectoring of the Peruvian coast. Glob. Ecol. Biogeogr. Lett. 1997, 6, 349–367. [Google Scholar] [CrossRef]
- Galán de Mera, A.; Linares Perea, E.; Campos de la Cruz, J.; Vicente Orellana, J.A. Interpretación fitosociológica de las lomas del desierto peruano. Rev. Biol. Trop. 2011, 59, 809–828. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Galán de Mera, A.; Sánchez Vega, I.; Linares Perea, E.; Campos, J.; Montoya, J.; Vicente Orellana, J.A. A phytosociological analysis and synopsis of the dry woodlands and succulent vegetation of the Peruvian Andes. An. Acad. Bras. Cienc. 2016, 88, 689–703. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Galán de Mera, A.; Méndez, E.; Linares Perea, E.; Campos de la Cruz, J.; Vicente Orellana, J.A. Las comunidades vegetales relacionadas con los procesos criogénicos en los Andes peruanos. Phytocoenologia 2014, 44, 121–161. [Google Scholar] [CrossRef]
- Reynel, C.; Pennington, R.T.; Särkinen, T. Cómo se Formó la Diversidad Ecológica del Perú; Fundación para el Desarrollo Agrario: Lima, Perú, 2013. [Google Scholar]
- Beaudet, G.; Gabert, P.; Philip, H.; Usselmann, P. Geomorfología de los Andes Centrales Sudamericanos (Un Corte Entre Perú Meridional y Bolivia Central); CNRS-Sociedad Geográfica de Lima: Lima, Perú, 2012. [Google Scholar]
- Izco, J. The biological reserve of San Francisco (Ecuador): Revision of the syntaxonomy and nomenclature of the vegetation. Lazaroa 2013, 34, 257–266. [Google Scholar] [CrossRef]
- Galán de Mera, A.; Rosa, M.V.; Cáceres, C. Una aproximación sintaxonómica sobre la vegetación del Perú. Clases, órdenes y alianzas. Acta Bot. Malac. 2002, 27, 75–103. [Google Scholar] [CrossRef][Green Version]
- Galán de Mera, A. Clasificación fitosociológica de la vegetación del Caribe y América del Sur. Arnaldoa 2005, 12, 86–111. [Google Scholar]
- Fernández Terrazas, E. Estudio fitosociológico de los bosques de Kewiña (Polylepis spp., Rosaceae) en la Cordillera de Cochabamba. Rev. Bol. Ecol. 1997, 2, 49–65. [Google Scholar]
- Hensen, I. Die vegetation von polylepis-wäldern der ostkordillere boliviens. Phytocoenologia 1995, 25, 235–277. [Google Scholar] [CrossRef]
- Mendoza, W.; Cano, A. El Género Polylepis en el Perú. Taxonomía, Morfología y Distribución; Editorial Académica Española: Saarbrücken, Germany, 2012. [Google Scholar]
- Seibert, P.; Menhofer, X. Die vegetation des wohngebietes der kallawaya und des hochlandes von Ulla-Ulla in den bolivianischen Anden. I. Phytocoenologia 1991, 20, 145–276. [Google Scholar] [CrossRef]
- Galán de Mera, A. Relación entre los suelos y la vegetación del Perú. Arnaldoa 1996, 4, 87–94. [Google Scholar]
- Galán de Mera, A. Una aproximación fitosociológica sobre los varillales húmedos de la Amazonía peruana. Stud. Bot. Univ. Salamanca 2001, 20, 125–133. [Google Scholar]
- Rangel, O.; Aguilar, M.; Sánchez, H.; Lowy, P.; Garzón, A.; Sánchez, L.A. Región de la Amazonía. Colomb. Divers. Biótica 1987, 1, 82–103. [Google Scholar]
- Ramia, M. Tipos de sabanas en los llanos de venezuela. Bol. Soc. Venez. Ci. Nat. 1967, 27, 264–288. [Google Scholar]
- Rangel, O.; Minorta-Cely, V. Tipos de vegetación de la Oriniquía colombiana. Colomb. Divers. Biótica 2018, 14, 533–612. [Google Scholar]
- QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: https://www.qgis.org/ (accessed on 18 July 2019).
- Instituto de Geología y Minería. Mapa Geológico del Perú. Escala 1: 1.000.000; Ministerio de Energía y Minas: Lima, Perú, 1975.
- Rivas-Martınez, S. Worldwide Bioclimatic Classification System. Available online: http://www.globalbioclimatics.org (accessed on 15 July 2018).
- Schwarz, T. Climate-Data.org. Available online: http://es.climate-data.org/ (accessed on 18 July 2018).
- Galán deMera, A.; Campos de la Cruz, J.; Linares-Perea, E.; Montoya-Quino, J.; Trujillo-Vera, C.; Villasante-Benavides, F.; Vicente-Orellana, J.A. Un ensayo sobre bioclimatología, vegetación y antropología en el Perú. Chloris Chil. 2017, 20, 2. [Google Scholar]
- Dengler, J.; Chitrý, M.; Ewald, J. Phytosociology. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; pp. 2767–2779. [Google Scholar]
- Phillips, O.; Miller, J.S. Global Patterns of Plant Diversity: Alwyn H. Gentry’s Forests Transect Data Set; Missouri Botanical Garden: St. Louis, MO, USA, 2002. [Google Scholar]
- Macbride, J.F. Flora of Peru; Field Museum of Natural History: Chicago, IL, USA, 1936–1995. [Google Scholar]
- Tryon, R. The ferns of Peru, Polypodiaceae (Dennstaedtieae to Oleandreae). Contrib. Gray Herb. 1964, 194, 3–253. [Google Scholar]
- Gentry, A.H. A Field Guide to the Families and Genera of Woody Plants of Northwest South America (Colombia, Ecuador, Peru) with Supplementary Notes on Herbaceous Taxa; Conservation International: Washington, DC, USA, 1993. [Google Scholar]
- Tryon, R.M.; Stolze, R.G. Pteridophyta of Peru Part I. 1. Ophioglossaceae-12. Cyatheaceae. Fieldiana Bot. 1989, 20, 1–145. [Google Scholar] [CrossRef]
- Tryon, R.M.; Stolze, R.G. Pteridophyta of Peru Part IV. 17. Dryopteridaceae. Fieldiana Bot. 1991, 27, 1–176. [Google Scholar]
- Tryon, R.M.; Stolze, R.G. Pteridophyta of Peru Part III. 16. Thelypteridaceae. Fieldiana Bot. 1992, 29, 1–80. [Google Scholar] [CrossRef]
- Tryon, R.M.; Stolze, R.G. Pteridophyta of Peru Part V. 18. Aspleniaceae-21. Polypodiaceae. Fieldiana Bot. 1993, 32, 1–190. [Google Scholar] [CrossRef]
- Tryon, R.M.; Stolze, R.G. Pteridophyta of Peru Part VI. 22. Marsileaceae-28. Isoetaceae. Fieldiana Bot. 1994, 34, 1–123. [Google Scholar] [CrossRef]
- Tovar, O. Las Gramíneas (Poaceae) del Perú. Ruizia 1993, 13, 9–480. [Google Scholar]
- Henderson, A.; Galeano, G.; Bernal, R. Field Guide to the Palms of the Americas; Princeton University Press: Princeton, NJ, USA, 1995. [Google Scholar]
- Brako, L.; Zarucchi, J.L. Catalogue of the Flowering Plants and Gymnosperms of Peru; Missouri Botanical Garden: St. Louis, MO, USA, 1993. [Google Scholar]
- Beltrán, H.; Salinas, I. Flora vascular y vegetación de los Bosques Montanos Húmedos de Carpish (Huánuco-Perú). Arnaldoa 2010, 17, 107–130. [Google Scholar]
- Monteagudo Mendoza, A.L.; Huamán Guerrero, M. Catálogo de los árboles y afines de la Selva Central del Perú. Arnaldoa 2010, 17, 203–242. [Google Scholar]
- Thiers, B.M. Index Herbariorum. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 5 December 2016).
- The Plant List. A Working List of All Plant Species, Versión 1.1. Available online: http://www.theplantlist.org/ (accessed on 19 June 2020).
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. Kongel. Danske Vidensk. Selsk. 1957, 5, 1–34. [Google Scholar]
- Tichý, L. JUICE, software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- Hammer, Ø. PAST—Paleontological Statistics Version 4.03; University of Oslo: Oslo, Norway, 2020. [Google Scholar]
- Chytrý, M.; Tichý, L.; Holt, J.; Botta-Dukat, Z. Determination of diagnostic species with statistical fidelity measures. J. Veg. Sci. 2002, 13, 79–90. [Google Scholar] [CrossRef]
- Tichý, L.; Chytrý, M. Statistical determination of diagnostic species for site groups of unequal size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Theurillat, J.P.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H. International code of phytosociological nomenclature. Appl. Veg. Sci. 2020. [CrossRef]


| VEN | COLori | COLcen | COLocc | ECU | PERU | BOL | ARG | |
|---|---|---|---|---|---|---|---|---|
| Plots; α Diversity | 14; 226 | 39; 321 | 18; 821 | 39; 550 | 103; 819 | 106; 791 | 24; 334 | 21; 89 |
| VEN | 1 | 0.04753199 | 0.04011461 | 0.05412371 | 0.04784689 | 0.02359882 | 0.01403509 | 0 |
| COLori | 0.04753199 | 1 | 0.08581436 | 0.11710677 | 0.09649123 | 0.05215827 | 0.02406015 | 0.0097561 |
| COLcen | 0.04011461 | 0.08581436 | 1 | 0.11232677 | 0.11097561 | 0.05086849 | 0.01545064 | 0.01098901 |
| COLocc | 0.05412371 | 0.11710677 | 0.11232677 | 1 | 0.08473338 | 0.05965697 | 0.01342282 | 0.00625978 |
| ECU | 0.04784689 | 0.09649123 | 0.11097561 | 0.08473338 | 1 | 0.1378882 | 0.04127257 | 0.02863436 |
| PERU | 0.02359882 | 0.05215827 | 0.05086849 | 0.05965697 | 0.1378882 | 1 | 0.06696035 | 0.025 |
| BOL | 0.01403509 | 0.02406015 | 0.01545064 | 0.01342282 | 0.04127257 | 0.06696035 | 1 | 0.07852194 |
| ARG | 0 | 0.0097561 | 0.01098901 | 0.00625978 | 0.02863436 | 0.025 | 0.07852194 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galán-de-Mera, A.; Campos-de-la-Cruz, J.; Linares-Perea, E.; Montoya-Quino, J.; Torres-Marquina, I.; Vicente-Orellana, J.A. A Phytosociological Study on Andean Rainforests of Peru, and a Comparison with the Surrounding Countries. Plants 2020, 9, 1654. https://doi.org/10.3390/plants9121654
Galán-de-Mera A, Campos-de-la-Cruz J, Linares-Perea E, Montoya-Quino J, Torres-Marquina I, Vicente-Orellana JA. A Phytosociological Study on Andean Rainforests of Peru, and a Comparison with the Surrounding Countries. Plants. 2020; 9(12):1654. https://doi.org/10.3390/plants9121654
Chicago/Turabian StyleGalán-de-Mera, Antonio, José Campos-de-la-Cruz, Eliana Linares-Perea, Juan Montoya-Quino, Iván Torres-Marquina, and José Alfredo Vicente-Orellana. 2020. "A Phytosociological Study on Andean Rainforests of Peru, and a Comparison with the Surrounding Countries" Plants 9, no. 12: 1654. https://doi.org/10.3390/plants9121654
APA StyleGalán-de-Mera, A., Campos-de-la-Cruz, J., Linares-Perea, E., Montoya-Quino, J., Torres-Marquina, I., & Vicente-Orellana, J. A. (2020). A Phytosociological Study on Andean Rainforests of Peru, and a Comparison with the Surrounding Countries. Plants, 9(12), 1654. https://doi.org/10.3390/plants9121654