Leaf Gas Exchange of Tomato Depends on Abscisic Acid and Jasmonic Acid in Response to Neighboring Plants under Different Soil Nitrogen Regimes



 and
and 
Abstract
:1. Introduction
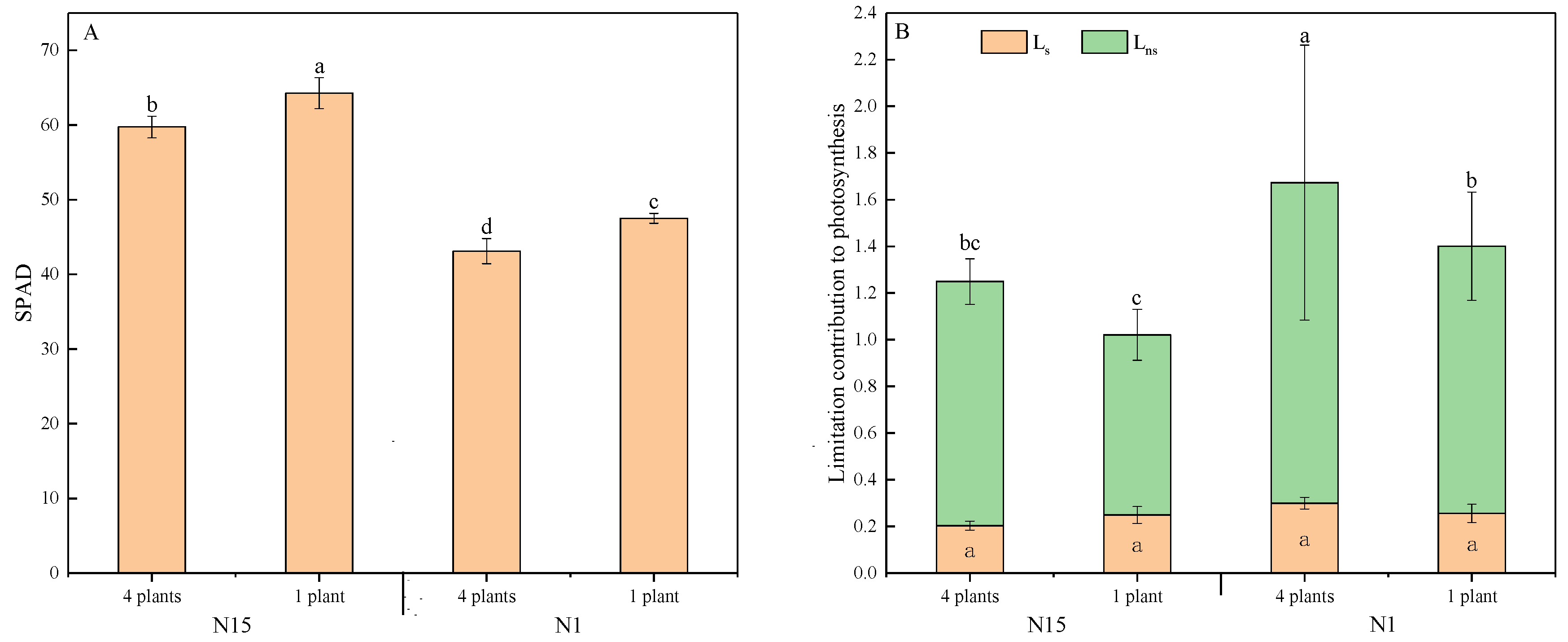
2. Results
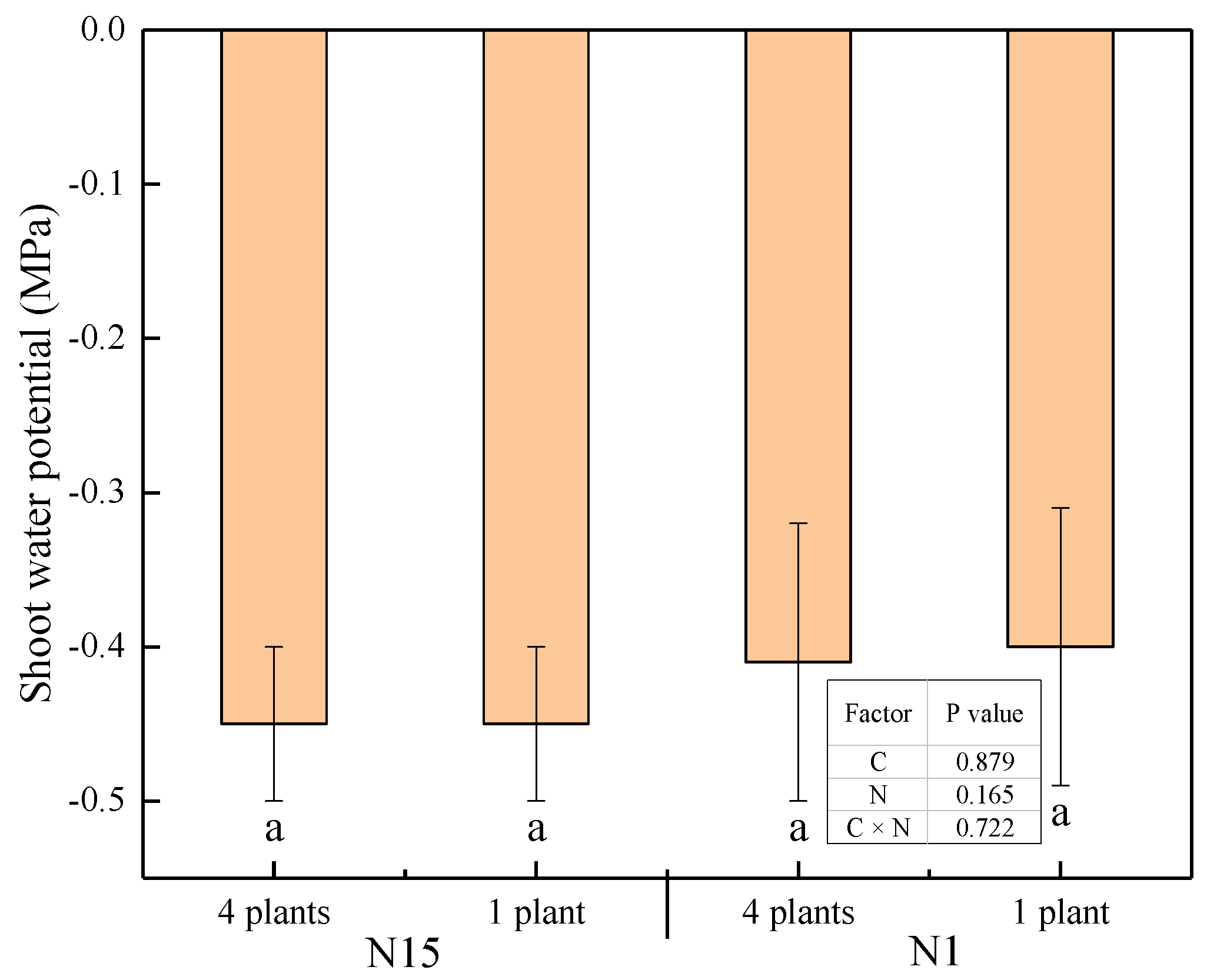
2.1. Leaf Water Status
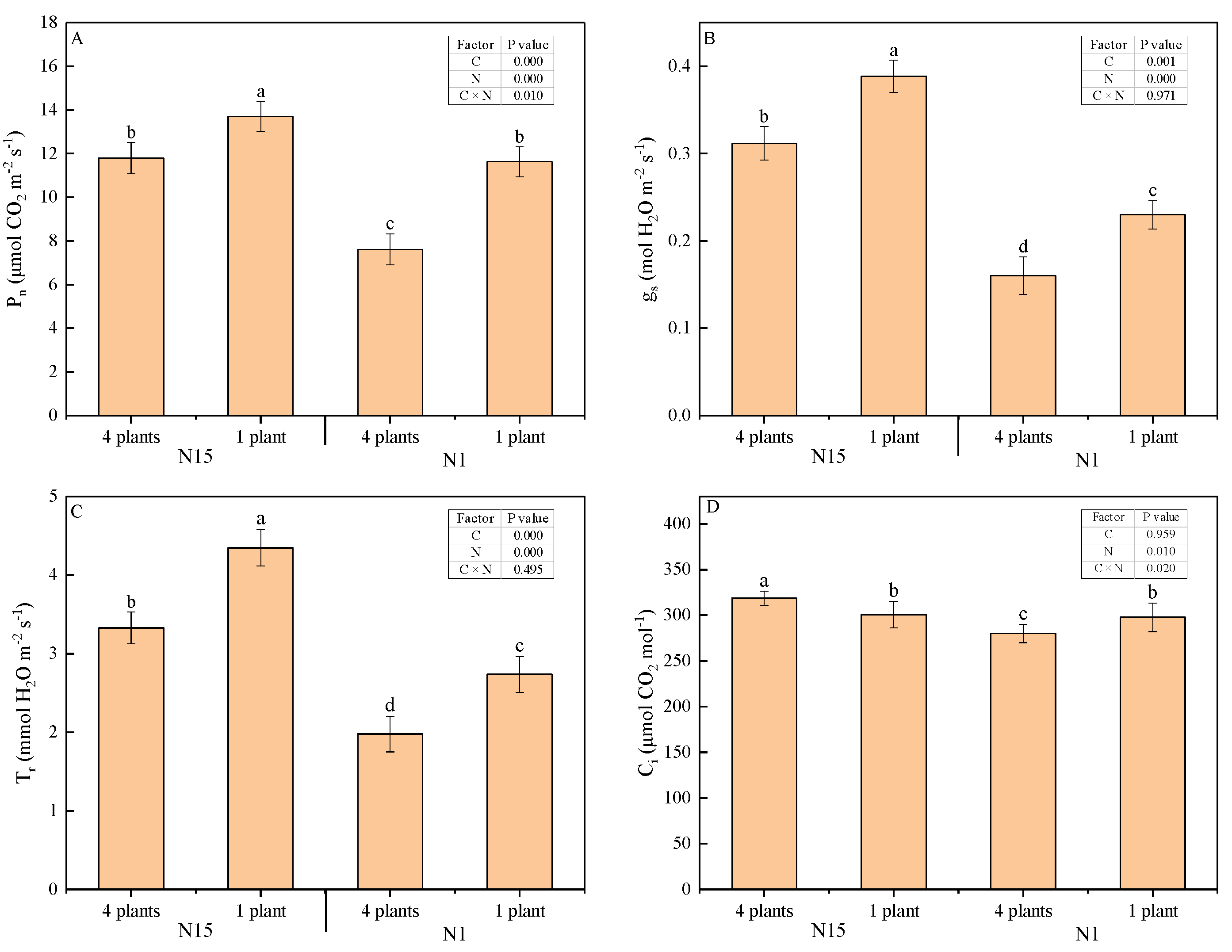
2.2. Plant Growth and Leaf Gas Exchange
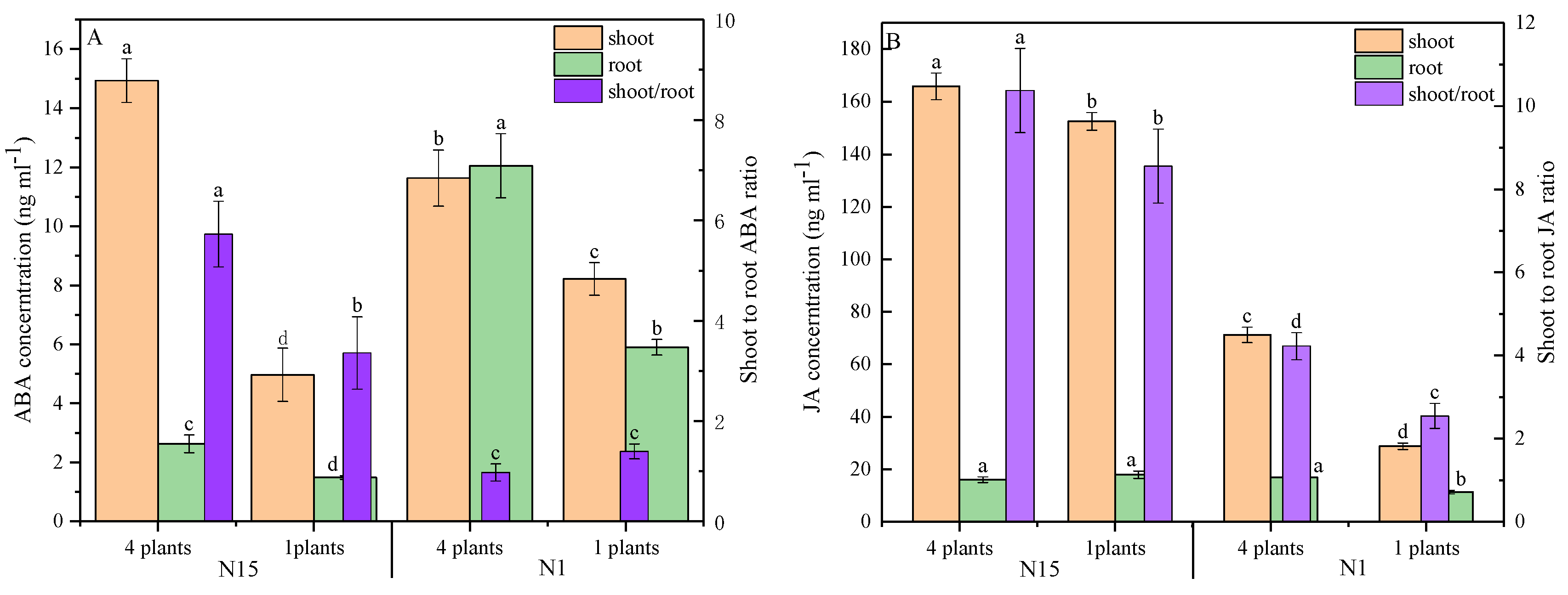
2.3. Hormonal Response
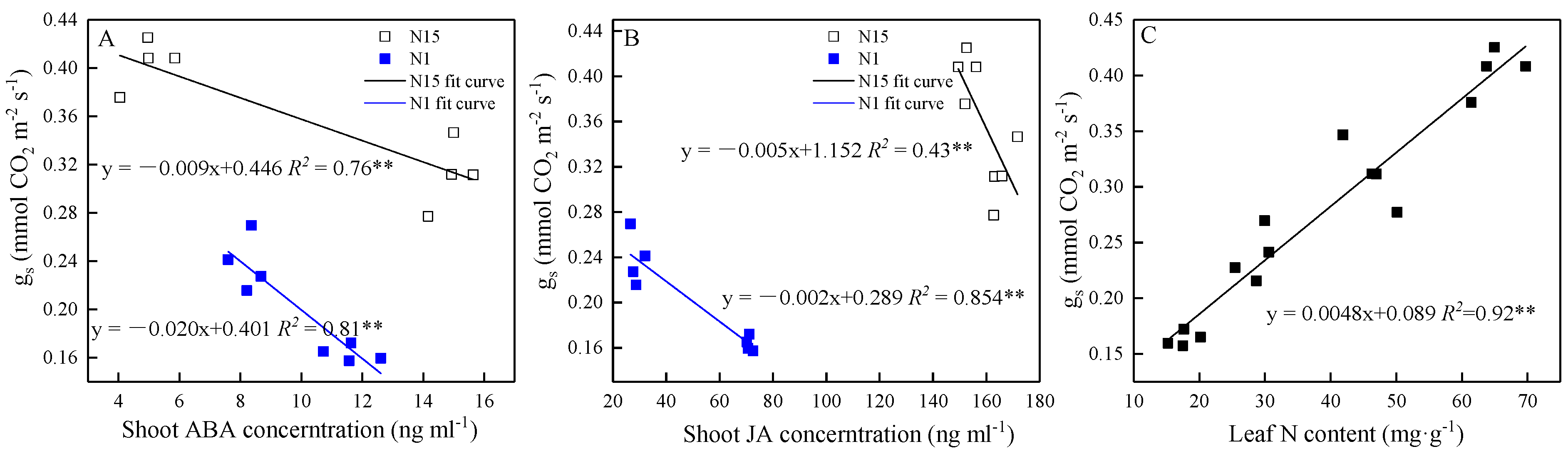
2.4. Correlations of gs, Pn, Hormones, and Leaf N Content
2.5. Correlations of Chlorophyll Index and Leaf nitrogen Content
3. Discussion
3.1. Effects of Neighbor Competing and Nitrogen Availability on Leaf Gas Exchange and Abscisic Acid (ABA)
3.2. Effects of Neighbor Competing and Nitrogen Availability on Leaf Gas Exchange and Jasmonic Acid (JA)
3.3. Effects of Neighbor Competing and Nitrogen Availability on Pn
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Measurement of Plant Development, SPAD and Leaf N Content
4.3. Determination of Leaf Gas Exchange Parameters
4.4. Root and Shoot Sap Collection
4.5. ABA and JA Measurement
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Simon, J.; Dannenmann, M.; Gasche, R.; Holst, J.; Mayer, H.; Papen, H.; Rennenberg, H. Competition for nitrogen between adult European beech and its offspring is reduced by avoidance strategy. For. Ecol. Manag. 2011, 262, 105–114. [Google Scholar] [CrossRef]
- Wu, P.; Wang, G.; Farooq, T.H.; Li, Q.; Zou, X.; Ma, X. Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? J. Soils Sediments 2017, 17, 2775–2785. [Google Scholar] [CrossRef]
- Cai, H.; Lu, Y.; Xie, W.; Zhu, T.; Lian, X. Transcriptome response to nitrogen starvation in rice. J. Biosci. 2012, 37, 731–747. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Mommer, L.; Voesenek, L.A.C.J. Molecular mechanisms of plant competition: Neighbour detection and response strategies. Funct. Ecol. 2013, 27, 841–853. [Google Scholar] [CrossRef]
- Pierik, R.; Whitelam, G.C.; Voesenek, L.A.C.J.; de Kroon, H.; Visser, E.J.W. Canopy studies on ethylene-insensitive tobacco identify ethylene as a novel element in blue light and plant-plant signalling. Plant J. 2004, 38, 310–319. [Google Scholar] [CrossRef]
- Vysotskaya, L.; Wilkinson, S.; Davies, W.J.; Arkhipova, T.; Kudoyarova, G. The effect of competition from neighbours on stomatal conductance in lettuce and tomato plants. Plant Cell Environ. 2011, 34, 729–737. [Google Scholar] [CrossRef]
- Masclaux, F.G.; Bruessow, F.; Schweizer, F.; Gouhier-Darimont, C.; Keller, L.; Reymond, P. Transcriptome analysis of intraspecific competition in Arabidopsis thaliana reveals organ-specific signatures related to nutrient acquisition and general stress response pathways. BMC Plant Biol. 2012, 12, 227. [Google Scholar] [CrossRef] [Green Version]
- Pierik, R.; de Wit, M. Shade avoidance: Phytochrome signalling and other aboveground neighbour detection cues. J. Exp. Bot. 2014, 65, 2815–2824. [Google Scholar] [CrossRef] [Green Version]
- Ballaré, C.L.; Pierik, R. The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Keuskamp, D.H.; Sasidharan, R.; Pierik, R. Physiological regulation and functional significance of shade avoidance responses to neighbors. Plant Signal Behav. 2010, 5, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Pierik, R.; Sasidharan, R.; Voesenek, L.A.C.J. Growth Control by Ethylene: Adjusting Phenotypes to the Environment. J. Plant Growth Regul. 2007, 26, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Vysotskaya, L.B.; Veselov, S.Y.; Kudoyarova, G.R. Effect of Competition and Treatment with Inhibitor of Ethylene Perception on Growth and Hormone Content of Lettuce Plants. J. Plant Growth Regul. 2017, 36, 450–459. [Google Scholar] [CrossRef]
- He, J.; Liang, Y.-K. Stomata. In eLS; JohnWiley & Sons, Ltd: Chichester, UK, 2018. [Google Scholar] [CrossRef]
- Arkhipova, T.N.; Vysotskaya, L.B.; Martinenko, E.V.; Ivanov, I.I.; Kudoyarova, G.R. Participation of cytokinins in plant response to competitors. Russ. J. Plant Physiol. 2015, 62, 524–533. [Google Scholar] [CrossRef]
- De Ollas, C.; Dodd, I.C. Physiological impacts of ABA-JA interactions under water-limitation. Plant Mol. Biol. 2016, 91, 641–650. [Google Scholar] [CrossRef] [PubMed]
- de Ollas, C.; Hernando, B.; Arbona, V.; Gómez-Cadenas, A. Jasmonic acid transient accumulation is needed for abscisic acid increase in citrus roots under drought stress conditions. Physiol. Plant. 2013, 147, 296–306. [Google Scholar] [CrossRef]
- Mahouachi, J.; Arbona, V.; Gómez-Cadenas, A. Hormonal changes in papaya seedlings subjected to progressive water stress and re-watering. Plant Growth Regul. 2007, 53, 43–51. [Google Scholar] [CrossRef]
- Pedranzani, H.; Sierra-de-Grado, R.; Vigliocco, A.; Miersch, O.; Abdala, G. Cold and water stresses produce changes in endogenous jasmonates in two populations of Pinus pinaster Ait. Plant Growth Regul. 2007, 52, 111–116. [Google Scholar] [CrossRef]
- Ali, M.; Baek, K.-H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Herde, O.; Pena-Cortes, H.; Willmitzer, L.; Fisahn, J. Stomatal responses to jasmonic acid, linolenic acid and abscisic acid in wild-type and ABA-deficient tomato plants. Plant Cell Environ. 1997, 20, 136–141. [Google Scholar] [CrossRef]
- de Ollas, C.; Arbona, V.; Gómez-Cadenas, A.; Dodd, I.C. Attenuated accumulation of jasmonates modifies stomatal responses to water deficit. J. Exp. Bot. 2018, 69, 2103–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kegge, W.; Weldegergis, B.T.; Soler, R.; Vergeer-Van Eijk, M.; Dicke, M.; Voesenek, L.A.C.J.; Pierik, R. Canopy light cues affect emission of constitutive and methyl jasmonate-induced volatile organic compounds in Arabidopsis thaliana. New Phytol. 2013, 200, 861–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossa, R.; López-Carbonell, M.; Jubany-Marí, T.; Alegre, L. Interplay Between Abscisic Acid and Jasmonic Acid and its Role in Water-oxidative Stress in Wild-type, ABA-deficient, JA-deficient, and Ascorbate-deficient Arabidopsis Plants. J. Plant Growth Regul. 2011, 30, 322–333. [Google Scholar] [CrossRef]
- Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of endogenous abscisic acid in methyl jasmonate-induced stomatal closure in Arabidopsis. Plant Physiol. 2011, 156, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janusauskaite, D.; Feiziene, D.; Feiza, V. Nitrogen-induced variations in leaf gas exchange of spring triticale under field conditions. Acta Physiol. Plant. 2017, 39, 1–12. [Google Scholar] [CrossRef]
- Li, D.; Tian, M.; Cai, J.; Jiang, D.; Cao, W.; Dai, T. Effects of low nitrogen supply on relationships between photosynthesis and nitrogen status at different leaf position in wheat seedlings. Plant Growth Regul. 2013, 70, 257–263. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Gong, S.; Xu, D.; Sui, J. Nitrogen fertigation effect on photosynthesis, grain yield and water use efficiency of winter wheat. Agric. Water Manag. 2017, 179, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Davies, W.J.; Kudoyarova, G.; Hartung, W. Long-distance ABA Signaling and Its Relation to Other Signaling Pathways in the Detection of Soil Drying and the Mediation of the Plant’s Response to Drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; de Ollas, C.; Dodd, I.C. Long-distance ABA transport can mediate distal tissue responses by affecting local ABA concentrations. J. Integr. Plant Biol. 2018, 60, 16–33. [Google Scholar] [CrossRef] [Green Version]
- Vysotskaya, L.B.; Arkhipova, T.N.; Kudoyarova, G.R.; Veselov, S.Y. Dependence of growth inhibiting action of increased planting density on capacity of lettuce plants to synthesize ABA. J. Plant Physiol. 2018, 220, 69–73. [Google Scholar] [CrossRef]
- Davies, W.J.; Zhang, J. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Broadley, M.R.; Escobar-Gutiérrez, A.J.; Burns, A.; Burns, I.G. Nitrogen-limited growth of lettuce is associated with lower stomatal conductance. New Phytol. 2001, 152, 97–106. [Google Scholar] [CrossRef]
- Rajaona, A.M.; Brueck, H.; Asch, F. Leaf Gas Exchange Characteristics of Jatropha as Affected by Nitrogen Supply, Leaf Age and Atmospheric Vapour Pressure Deficit. J. Agron. Crop Sci. 2013, 199, 144–153. [Google Scholar] [CrossRef]
- Oka, M.; Shimoda, Y.; Sato, N.; Inoue, J.; Yamazaki, T.; Shimomura, N.; Fujiyama, H. Abscisic acid substantially inhibits senescence of cucumber plants (Cucumis sativus) grown under low nitrogen conditions. J. Plant Physiol. 2012, 169, 789–796. [Google Scholar] [CrossRef]
- Kang, G.; Wu, Y.; Li, G.; Wang, P.; Han, Q.; Wang, Y.; Xie, Y.; Feng, W.; Ma, D.; Wang, C.; et al. Proteomics combined with BSMV-VIGS methods identified some N deficiency-responsive protein species and ABA role in wheat seedling. Plant Soil 2019, 444, 177–191. [Google Scholar] [CrossRef]
- Dodd, I.C.; Tan, L.P.; He, J. Do increases in xylem sap pH and/or ABA concentration mediate stomatal closure following nitrate deprivation? J. Exp. Bot. 2003, 54, 1281–1288. [Google Scholar] [CrossRef]
- Olszewski, J.; Makowska, M.; Pszczółkowska, A.; Okorski, A.; Bieniaszewski, T. The effect of nitrogen fertilization on flag leaf and ear photosynthesis and grain yield of spring wheat. Plant Soil Environ. 2014, 60, 531–536. [Google Scholar] [CrossRef]
- Zhu, K.; Wang, A.; Wu, J.; Yuan, F.; Guan, D.; Jin, C.; Zhang, Y.; Gong, C. Effects of nitrogen additions on mesophyll and stomatal conductance in Manchurian ash and Mongolian oak. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Andrade, A.; Escalante, M.; Vigliocco, A.; Tordable, M.d.C.; Alemano, S. Involvement of jasmonates in responses of sunflower (Helianthus annuus) seedlings to moderate water stress. Plant Growth Regul. 2017, 83, 501–511. [Google Scholar] [CrossRef]
- de Wit, M.; Spoel, S.H.; Sanchez-Perez, G.F.; Gommers, C.M.M.; Pieterse, C.M.J.; Voesenek, L.A.C.J.; Pierik, R. Perception of low red:far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. Plant J. 2013, 75, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Courbier, S.; Grevink, S.; Sluijs, E.; Bonhomme, P.O.; Kajala, K.; Van Wees, S.C.M.; Pierik, R. Far-red light promotes Botrytis cinerea disease development in tomato leaves via jasmonate-dependent modulation of soluble sugars. Plant Cell Environ. 2020, 43, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Imran, Q.M.; Yun, B.-W.; Lee, I.-J. Osmoprotective functions conferred to soybean plants via inoculation with Sphingomonas sp. LK11 and exogenous trehalose. Microbiol. Res. 2017, 205, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Munemasa, S.; Oda, K.; Watanabe-Sugimoto, M.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. The coronatine-insensitive 1 Mutation Reveals the Hormonal Signaling Interaction between Abscisic Acid and Methyl Jasmonate in Arabidopsis Guard Cells. Specific Impairment of Ion Channel Activation and Second Messenger Production. Plant Physiol. 2007, 143, 1398–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemann, M.; Dhakarey, R.; Hazman, M.; Miro, B.; Kohli, A.; Nick, P. Exploring Jasmonates in the Hormonal Network of Drought and Salinity Responses. Front. Plant Sci. 2015, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C. Action of jasmonates in plant stress responses and development—Applied aspects. Biotechnol. Adv. 2014, 32, 31–39. [Google Scholar] [CrossRef]
- Jang, S.-W.; Hamayun, M.; Sohn, E.-Y.; Shin, D.-H.; Kim, K.-U.; Lee, B.-H.; Lee, I.-J. Effect of elevated nitrogen levels on endogenous gibberellin and jasmonic acid contents of three rice (Oryza sativa L.) cultivars. J. Plant Nutr. Soil Sci. 2008, 171, 181–186. [Google Scholar] [CrossRef]
- Wang, X.; Du, T.; Huang, J.; Peng, S.; Xiong, D. Leaf hydraulic vulnerability triggers the decline in stomatal and mesophyll conductance during drought in rice. J. Exp. Bot. 2018, 69, 4033–4045. [Google Scholar] [CrossRef]
- Zeng, X.-P.; Zhu, K.; Lu, J.-M.; Jiang, Y.; Yang, L.-T.; Xing, Y.-X.; Li, Y.-R. Long-Term Effects of Different Nitrogen Levels on Growth, Yield, and Quality in Sugarcane. Agronomy 2020, 10, 353. [Google Scholar] [CrossRef] [Green Version]
- Wen, B.; Li, C.; Fu, X.; Li, D.; Li, L.; Chen, X.; Wu, H.; Cui, X.; Zhang, X.; Shen, H.; et al. Effects of nitrate deficiency on nitrate assimilation and chlorophyll synthesis of detached apple leaves. Plant Physiol. Biochem. 2019, 142, 363–371. [Google Scholar] [CrossRef]
- Berry, J.A.; Downton, W.J.S. Environmental Regulation of Photosynthesis. In Photosynthesis, Volume II: Development, Carbon, Metabolism, and Plant Productivity; Academic Press Inc.: New York, NY, USA; London, UK, 1982; pp. 263–343. [Google Scholar]
- You, C.; Zhu, H.; Xu, B.; Huang, W.; Wang, S.; Ding, Y.; Liu, Z.; Li, G.; Chen, L.; Ding, C.; et al. Effect of Removing Superior Spikelets on Grain Filling of Inferior Spikelets in Rice. Front. Plant Sci. 2016, 7, 1161. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot Length /(cm) | Stem Diameter /(mm) | Leaf Area /(cm2·plant−1) | Dry Biomass /(g·plant−1) | |
|---|---|---|---|---|---|
| N15P4 | 12.07 ± 0.35 a | 3.02 ± 0.09 b | 94.21 ± 7.58 b | 0.368 ± 0.016 b | |
| N15P1 | 12.24 ± 0.16 a | 3.21 ± 0.05 a | 138.23 ± 9.71 a | 0.598 ± 0.026 a | |
| N1P4 | 10.40 ± 0.45 c | 2.25 ± 0.03 d | 44.38 ± 3.70 d | 0.195 ± 0.010 d | |
| N1P1 | 11.21 ± 0.32 b | 2.61 ± 0.02 c | 69.69 ± 6.12 c | 0.261 ± 0.014 c | |
| Factors | df | p-value | |||
| Competition (C) | 1 | 0.002 ** | 0.000 ** | 0.000 ** | 0.000 ** |
| Nitrate application (N) | 1 | 0.000 ** | 0.000 ** | 0.000 ** | 0.000 ** |
| C × N | 1 | 0.029 * | 0.002 ** | 0.001 ** | 0.000 ** |
| Error | 20 | ||||
| Factors | df | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Shoot ABA | Root ABA | ABA Ratio | Shoot JA | Root JA | JA Ratio | ||
| Competition (C) | 1 | 0.000 ** | 0.000 ** | 0.010 * | 0.000 ** | 0.018 * | 0.008 ** |
| Nitrate application (N) | 1 | 0.959 ns | 0.000 ** | 0.000 ** | 0.000 ** | 0.002 ** | 0.000 ** |
| C × N | 1 | 0.010 ** | 0.000 ** | 0.001 ** | 0.000 ** | 0.000 ** | 0.000 ** |
| Error | 8 | ||||||
| Factors | gs in N15 | gs in N1 | Pn in N15 | Pn in N1 |
|---|---|---|---|---|
| shoot ABA | ‒0.872 ** | ‒0.912 ** | ‒0.863 ** | ‒0.941 ** |
| root ABA | ‒0.936 ** | ‒0.966 ** | ‒0.821 ** | ‒0.980 ** |
| ABA ratio | ‒0.715 * | ‒0.895 ** | ‒0.834 * | ‒0.866 ** |
| shoot JA | ‒0.715 * | ‒0.992 ** | ‒0.791 * | ‒0.977 ** |
| root JA | 0.476 | ‒0.950 ** | ‒0.744 * | ‒0.967 ** |
| JA ratio | ‒0.556 | ‒0.984 ** | ‒0.757 * | ‒0.937 ** |
| Factors | df | p-Value | ||
|---|---|---|---|---|
| SPAD | Ls | Lns | ||
| Competition (C) | 1 | 0.000 ** | 0.959 ns | 0.010 ** |
| Nitrate application (N) | 1 | 0.000 ** | 0.001 ** | 0.000 ** |
| C × N | 1 | 0.907 ns | 0.002 ** | 0.258 * |
| Error | 20 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Hamani, A.K.M.; Si, Z.; Liang, Y.; Gao, Y.; Duan, A. Leaf Gas Exchange of Tomato Depends on Abscisic Acid and Jasmonic Acid in Response to Neighboring Plants under Different Soil Nitrogen Regimes. Plants 2020, 9, 1674. https://doi.org/10.3390/plants9121674
Li S, Hamani AKM, Si Z, Liang Y, Gao Y, Duan A. Leaf Gas Exchange of Tomato Depends on Abscisic Acid and Jasmonic Acid in Response to Neighboring Plants under Different Soil Nitrogen Regimes. Plants. 2020; 9(12):1674. https://doi.org/10.3390/plants9121674
Chicago/Turabian StyleLi, Shuang, Abdoul Kader Mounkaila Hamani, Zhuanyun Si, Yueping Liang, Yang Gao, and Aiwang Duan. 2020. "Leaf Gas Exchange of Tomato Depends on Abscisic Acid and Jasmonic Acid in Response to Neighboring Plants under Different Soil Nitrogen Regimes" Plants 9, no. 12: 1674. https://doi.org/10.3390/plants9121674
APA StyleLi, S., Hamani, A. K. M., Si, Z., Liang, Y., Gao, Y., & Duan, A. (2020). Leaf Gas Exchange of Tomato Depends on Abscisic Acid and Jasmonic Acid in Response to Neighboring Plants under Different Soil Nitrogen Regimes. Plants, 9(12), 1674. https://doi.org/10.3390/plants9121674