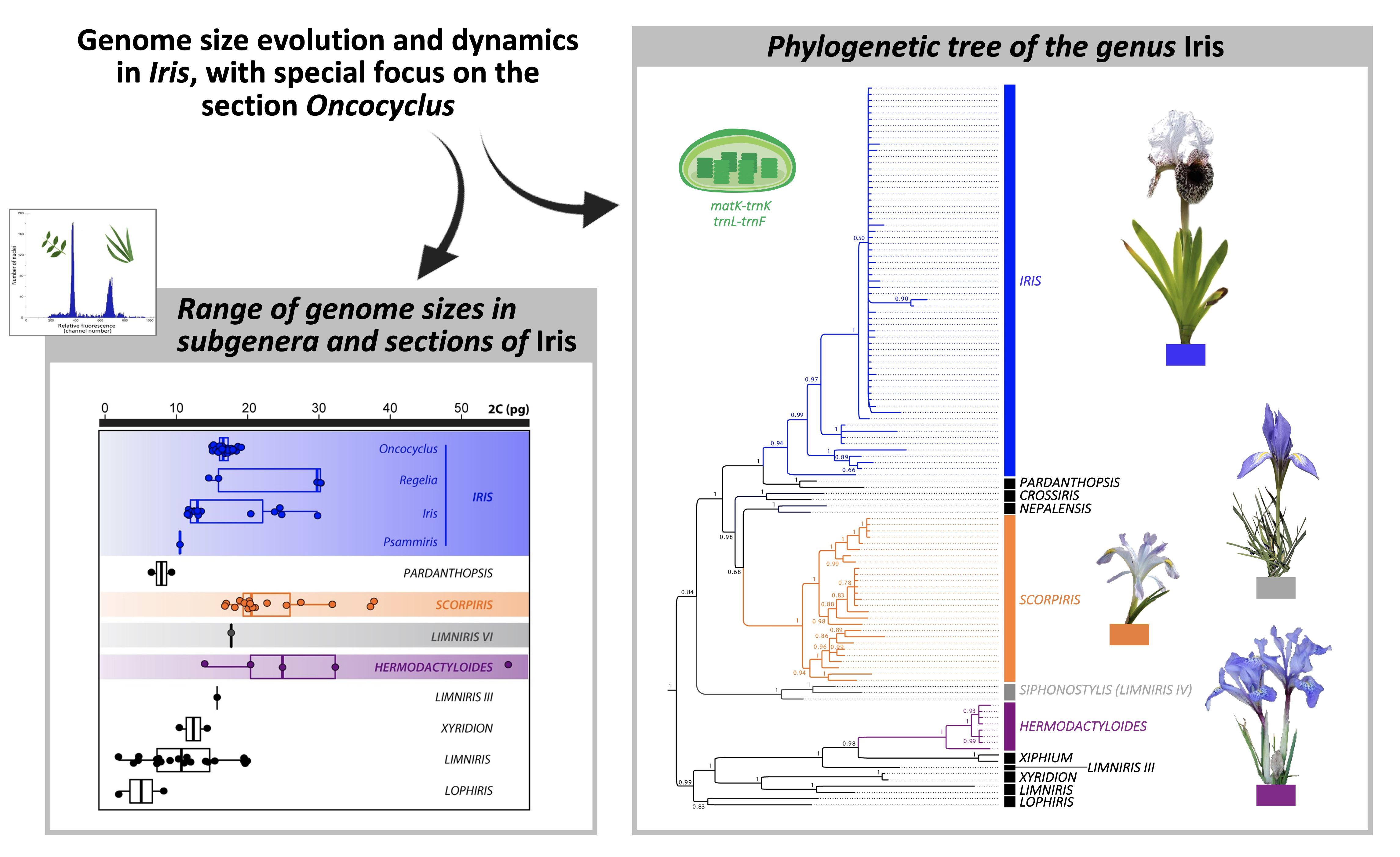
Genome Size Evolution and Dynamics in Iris, with Special Focus on the Section Oncocyclus



 ,
,  ,
,  and
and
Abstract

1. Introduction
2. Results
2.1. Iris Phylogenetic Reconstruction
2.2. Genome Size Diversity across Iris Subgenera and Sections
2.3. Genome Size Evolution in Oncocyclus Section
3. Discussion
3.1. Iris Displays a Great Diversity of C-Values but They Are Unequally Distributed across Subgenera and Sections
3.2. Genome Size Dynamics Could Have Contributed to the Radiation of Oncocyclus Irises
3.3. Lebanese Populations Attributed to Iris persica (Subgenus Scorpiris) Should Be Segregated into a Distinct Species
4. Materials and Methods
4.1. Material
4.2. Molecular Phylogenetic Reconstruction
4.3. DNA Content Assessment
4.4. Tempo and Mode of Character Evolution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rix, M. Section Oncocyclus. In A Guide to Species Irises; The Species Group of the British Iris Society; Cambridge Press: London, UK, 1997. [Google Scholar]
- Sapir, Y.; Shmida, A.; Ne’eman, G. Pollination of Oncocyclus irises (Iris: Iridaceeae) by night-sheltering male bees. Plant Biol. 2005, 7, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Sapir, Y.; Shmida, A.; Ne’eman, G. Morning floral heat as a reward to the pollinators of the Oncocyclus irises. Oecologia 2006, 147, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Vereecken, N.J.; Wilson, C.A.; Hötling, S.; Schulz, S.; Banketov, S.A.; Mardulyn, P. Pre-adaptations and the evolution of pollination by sexual deception: Cope’s rule of specialization revisited. Proc. R. Soc. B Biol. Sci. 2012, 279, 4786–4794. [Google Scholar] [CrossRef] [PubMed]
- Lavi, R.; Sapir, Y. Are pollinators the agents of selection for the extreme large size and dark color in Oncocyclus irises? New Phytol. 2015, 205, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Vereecken, N.J.; Dorchin, A.; Dafni, A. Reply to Lavi & Sapir (2015): Floral colour and pollinator-mediated selection in Oncocyclus irises (Iridaceae). New Phytol. 2015, 207, 948–949. [Google Scholar] [PubMed]
- Sapir, Y.; Shmida, A. Species concepts and ecogeographical divergence of Oncocyclus irises. Isr. J. Plant Sci. 2002, 50, 119–127. [Google Scholar] [CrossRef]
- Wilson, C.A.; Padiernos, J.; Sapir, Y. The royal irises (Iris subg. Iris sect. Oncocyclus): Plastid and low-copy nuclear data contribute to an understanding of their phylogenetic relationships. Taxon 2016, 65, 35–46. [Google Scholar] [CrossRef]
- Cohen, O.; Avishai, M. The irises still exist: The conservation status of species Iris section Oncocyclus in Israel, a century after their description. Ann. Bot. 2000, 58, 145–160. [Google Scholar] [CrossRef]
- Saad, L.; Talhouk, S.N.; Mahy, G. Decline of endemic Oncocyclus irises (Iridaceae) of Lebanon: Survey and conservation needs. Oryx 2009, 43, 91–96. [Google Scholar] [CrossRef]
- Goldblatt, P.; Takei, M. Chromosome cytology of Iridaceae-patterns of variation, determination of ancestral base numbers, and mode of karyotype change. Ann. Mo. Bot. Gard. 1997, 84, 285–304. [Google Scholar] [CrossRef]
- Avishai, M.; Zohary, D. Chromosome in the Oncocyclus irises. Bot. Gaz. 1977, 138, 502–511. [Google Scholar] [CrossRef]
- Abdel Samad, N.; Bou Dagher-Kharrat, M.; Hidalgo, O.; El Zein, R.; Douaihy, B.; Siljak-Yakovlev, S. Unlocking the karyological and cytogenetic diversity of Iris from Lebanon: Oncocyclus section shows a distinctive profile and relative stasis during its continental radiation. PLoS ONE 2016, 11, e0160816. [Google Scholar] [CrossRef] [PubMed]
- Bou Dagher-Kharrat, M.; Abdel Samad, N.; Douaihy, B.; Bourge, M.; Fridlender, A.; Siljak-Yakovlev, S.; Brown, S.C. Nuclear DNA C-values for biodiversity screening: Case of the Lebanese flora. Plant Biosyst. 2013, 147, 1228–1237. [Google Scholar] [CrossRef]
- Leitch, I.J.; Johnston, E.; Pellicer, J.; Hidalgo, O.; Bennett, M.D. Plant DNA C-values Database, Release 7.1. 2019. Available online: https://cvalues.science.kew.org (accessed on 9 September 2020).
- Pellicer, J.; Leitch, I.J. The Plant DNA C-values database (release 7.1): An updated online repository of plant genome size data for comparative studies. New Phytol. 2019, 226, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Pellicer, J.; Hidalgo, O.; Dodsworth, S.; Leitch, I.J. Genome size diversity and its impact on the evolution of land plants. Genes 2018, 9, 88. [Google Scholar] [CrossRef]
- Kapralov, M.V.; Stift, M.; Filatov, D.A. Evolution of genome size in Hawaiian endemic genus Schiedea (Caryophyllaceae). Trop. Plant Biol. 2009, 2, 77–83. [Google Scholar] [CrossRef]
- Garnatje, T.; Hidalgo, O.; Vitales, D.; Pellicer, J.; Vallès, J.; Robin, O.; Garcia, S.; Siljak-Yakovlev, S. Swarm of terminal 35S in Cheirolophus (Asteraceae, Centaureinae). Genome 2012, 55, 529–535. [Google Scholar] [CrossRef]
- Hidalgo, O.; Vitales, D.; Vallès, J.; Garnatje, T.; Siljak-Yakovlev, S.; Leitch, I.J.; Pellicer, J. Cytogenetic insights into an oceanic island radiation: The dramatic evolution of pre-existing traits in Cheirolophus (Asteraceae, Cardueae-Centaureinae). Taxon 2017, 66, 146–157. [Google Scholar] [CrossRef]
- Naciri, Y.; Linder, H.P. The genetics of evolutionary radiations. Biol. Rev. 2020, 95, 1055–1072. [Google Scholar] [CrossRef]
- Guo, J.; Wilson, C.A. Molecular phylogeny of crested Iris based on five plastid markers (Iridaceae). Syst. Bot. 2013, 38, 987–995. [Google Scholar] [CrossRef]
- Ikinci, N.; Hall, T.; Lledo, M.D.; Clarkson, J.J.; Tillie, N.; Seisums, A.; Saito, T.; Harley, M.; Chase, M.W. Molecular phylogenetics of the juno irises, Iris subgenus Scorpiris (Iridaceae), based on six plastid markers. Bot. J. Linn. Soc. 2011, 167, 281–300. [Google Scholar] [CrossRef]
- Wilson, C.A. 2011. Subgeneric classification in Iris re-examined using chloroplast sequence data. Taxon 2011, 60, 27–35. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef]
- Garcia, S.; Garnatje, T.; Twibell, J.D.; Vallès, J. Genome size variation in the Artemisia arborescens complex (Asteraceae, Anthemideae) and its cultivars. Genome 2006, 49, 244–253. [Google Scholar] [CrossRef]
- Marie, D.; Brown, S.C. A cytometric exercise in plant DNA histograms, with 2C values for 70 species. Biol. Cell 1993, 78, 41–51. [Google Scholar] [CrossRef]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Two new nuclear isolation buffers for plant DNA flow cytometry: A test with 37 species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometry analysis of the cell cycle in intact plant tissues. Science 1993, 220, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)-a community resource of plant chromosome numbers. New Phytol. 2015, 206, 19–26. [Google Scholar] [CrossRef]
- Hall, T.; Seisums, A. 793. Iris wallisiae: Iridaceae. Curtis’s Bot. Mag. 2014, 31, 238–248. [Google Scholar] [CrossRef]
- Jozghasemi, S.; Rabiei, V.; Soleymani, A.; Khalighi, A. Karyotype analysis of seven Iris species native to Iran. Caryologia 2016, 69, 351–361. [Google Scholar] [CrossRef]
- Bai, C.; Alverson, W.S.; Follansbee, A.; Waller, D.M. New reports of nuclear DNA content for 407 vascular plant taxa from the United States. Ann. Bot. 2012, 110, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, P.; Walbot, V.; Zimmer, E.A. Estimation of genome size (C-value) in Iridaceae by cytophotometry. Ann. Mo. Bot. Gard. 1984, 71, 176–180. [Google Scholar] [CrossRef]
- Kentner, E.K.; Arnold, M.L.; Wessler, S.R. Characterization of high-copy-number retrotransposons from the large genomes of the Louisiana iris species and their use as molecular markers. Genetics 2003, 164, 685–697. [Google Scholar] [PubMed]
- Olszewska, M.J.; Osiecka, R. The relationship between 2C DNA content, life cycle type, systematic position, and the level of DNA endoreplication in nuclei of parenchyma cells during growth and differentiation of roots in some monocotyledonous species. Biochem. Physiol. Pflanz. 1982, 177, 319–336. [Google Scholar] [CrossRef]
- Pustahija, F.; Brown, S.C.; Bogunić, F.; Bašić, N.; Muratović, E.; Ollier, S.; Hidalgo, O.; Bourge, M.; Stevanović, V.; Siljak-Yakovlev, S. Small genomes dominate in plants growing on serpentine soils in West Balkans, an exhaustive study of 8 habitats covering 308 taxa. Plant Soil 2013, 373, 427–453. [Google Scholar] [CrossRef]
- Siljak-Yakovlev, S.; Pustahija, F.; Šolić, E.M.; Bogunić, F.; Muratović, E.; Bašić, N.; Catrice, O.; Brown, C.S. Towards a database of genome size and chromosome number of Balkan flora: C-values in 343 taxa with novel values for 252. Adv. Sci. Lett. 2010, 3, 190–213. [Google Scholar] [CrossRef]
- Šmarda, P.; Bureš, P.; Horová, L.; Leitch, I.J.; Mucina, L.; Pacini, E.; Tichý, L.; Grulich, V.; Rotreklová, O. Ecological and evolutionary significance of genomic GC content diversity in monocots. Proc. Natl. Acad. Sci. USA 2014, 111, E4096–E4102. [Google Scholar] [CrossRef]
- Veselý, P.; Bureš, P.; Šmarda, P.; Pavlíček, T. Genome size and DNA base composition of geophytes: The mirror of phenology and ecology? Ann. Bot. 2012, 109, 65–75. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, B.; Bai, C. New reports of nuclear DNA content for 66 traditional Chinese medicinal plant taxa in China. Caryologia 2013, 66, 375–383. [Google Scholar] [CrossRef]
- Zonneveld, B.J.; Leitch, I.J.; Bennett, M.D. First nuclear DNA amounts in more than 300 angiosperms. Ann. Bot. 2005, 96, 229–244. [Google Scholar] [CrossRef]
- Choi, B.; Weiss-Schneeweiss, H.; Temsch, E.M.; So, S.; Myeong, H.H.; Jang, T.S. Genome size and chromosome number evolution in Korean Iris L. species (Iridaceae Juss.). Plants 2020, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Jakše, J.; Sladonja, B.; Hruševar, D.; Landeka, N.; Brana, S.; Bohanec, B.; Milović, M.; Vladović, D.; Mitić, B.; et al. Molecular study of selected taxonomically critical taxa of the genus Iris L. from the broader Alpine-Dinaric area. Plants 2020, 9, 1229. [Google Scholar] [CrossRef] [PubMed]
- Puttick, M.N.; Clark, J.; Donoghue, P.C. Size is not everything: Rates of genome size evolution, not C-value, correlate with speciation in angiosperms. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152289. [Google Scholar] [CrossRef] [PubMed]
- Igea, J.; Miller, E.F.; Papadopulos, A.S.; Tanentzap, A.J. Seed size and its rate of evolution correlate with species diversification across angiosperms. PLoS Biol. 2017, 15, e2002792. [Google Scholar] [CrossRef]
- Fernández-Mazuecos, M.; Blanco-Pastor, J.L.; Juan, A.; Carnicero, P.; Forrest, A.; Alarcón, M.; Vargas, P.; Glover, B.J. Macroevolutionary dynamics of nectar spurs, a key evolutionary innovation. New Phytol. 2019, 222, 1123–1138. [Google Scholar] [CrossRef]
- Hidalgo, O.; Pellicer, J.; Christenhusz, M.; Schneider, H.; Leitch, A.R.; Leitch, I.J. Is there an upper limit to genome size? Trends Plant Sci. 2017, 22, 567–573. [Google Scholar] [CrossRef]
- Wang, H.; Talavera, M.; Min, Y.; Flaven, E.; Imbert, E. Neutral processes contribute to patterns of spatial variation for flower colour in the Mediterranean Iris lutescens (Iridaceae). Ann. Bot. 2016, 117, 995–1007. [Google Scholar] [CrossRef]
- Sung, C.J.; Bell, K.L.; Nice, C.C.; Martin, N.H. Integrating Bayesian genomic cline analyses and association mapping of morphological and ecological traits to dissect reproductive isolation and introgression in a Louisiana Iris hybrid zone. Mol. Ecol. 2018, 27, 959–978. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Genomic plant DNA preparation from fresh tissue-CTAB method. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Gravendeel, B.; Chase, M.W.; de Vogel, E.F.; Roos, M.C.; Mes, T.H.; Bachmann, K. Molecular phylogeny of Coelogyne (Epidendroideae; Orchidaceae) based on plastid RFLPs, matK, and nuclear ribosomal ITS sequences: Evidence for polyphyly. Am. J. Bot. 2001, 88, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Soltis, D.E. matK DNA sequences and phylogenetic reconstruction in Saxifragaceae s. str. Syst. Bot. 1994, 19, 143–156. [Google Scholar] [CrossRef]
- Penn, O.; Privman, E.; Ashkenazy, H.; Landan, G.; Graur, D.; Pupko, T. GUIDANCE: A web server for assessing alignment confidence scores. Nucleic Acids Res. 2010, 38, W23–W28. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit Software, Version 5.0.9; North Carolina State University: Raleigh, NC, USA, 1999. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer v1.4. 2007. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 1 October 2014).
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233. [Google Scholar] [CrossRef]
- Clark, J.; Hidalgo, O.; Pellicer, J.; Liu, H.; Marquardt, J.; Robert, Y.; Christenhusz, M.; Zhang, S.; Gibby, M.; Leitch, I.J.; et al. Genome evolution of ferns: Evidence for relative stasis of genome size across the fern phylogeny. New Phytol. 2016, 210, 1072–1082. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Meade, A. BayesTrees v. 1.3. School of Biological Science; University of Reading: Reading, UK, 2011. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: An R package for analyses of phylogenetics and evolution. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Swenson, N.G. Comparative methods and phylogenetic signal. In Functional and Phylogenetic Ecology in R; Springer: New York, NY, USA, 2014. [Google Scholar]
- Lee, C.; Blay, S.; Mooers, A.Ø.; Singh, A.; Oakley, T.H. CoMET: A Mesquite package for comparing models of continuous character evolution on phylogenies. Evol. Bioinform. 2006, 2, 183–186. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis; 2011; Available online: http://mesquiteproject.org (accessed on 13 October 2013).
- Mitchell, N.; Campbell, L.G.; Ahern, J.R.; Paine, K.C.; Giroldo, A.B.; Witney, K.D. Correlates of hybridization in plants. Evol. Lett. 2019, 3, 570–585. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Iris (Subgenus, Section, Species) | 2C in pg (SD) 1 | St 2 | Bu 3 | 2n 4 | P 5 | Origin | |
|---|---|---|---|---|---|---|---|
| Subgenus Iris | |||||||
| Section Oncocyclus | |||||||
| I. acutiloba subsp. acutiloba | 16.72 (0.17) | 2 | 2 | 20 | 1 | Azerbaijan: Jeyrankechmaz, Leg. F. Dépalle | |
| I. acutiloba subsp. lineolata | 16.73 (0.10) | 1 | 1 | 20 | 1 | Armenia (RBGK 2012-1109) | |
| I. acutiloba subsp. lineolata | 15.94 * | 2 | 2 | 20 | 1 | Armenia: Gandja, Leg. F. Dépalle | |
| I. antilibanotica | 16.83 (0.54) | 3 | 2 | 20 * | 1 | Lebanon: Kheibeh-Baalbeck, 1337 m | |
| I. antilibanotica | 17.05 (0.54) | 3 | 2 | 20 * | 1 | Lebanon: Kheibeh-Baalbeck, 1337 m | |
| I. assadiana | 17.44 (0.56) | 3 | 2 | 20 | 1 | Syria: Sadad, Leg. F. Dépalle | |
| I. atropurpurea | 18.53 (0.14) | 1 | 1 | 20 | 1 | Israel (RBGK 1998-2808) | |
| I. barnumiae x I. paradoxa f. choschab | 17.31(0.05) | 1 | 1 | 20 | 1 | Unknown Leg. Ray Drew (RBGK s.n.) | |
| I. bismarckiana | 18.24 (0.59) | 3 | 2 | 20 * | 1 | Lebanon: Sarada, 435 m | |
| I. bismarckiana | 19.24 (0.54) | 2 | 2 | 20 | 1 | Jordania: Ajloun, Leg. F. Dépalle | |
| I. camillae | 15.13 (0.49) | 3 | 2 | 20 | 1 | Azerbaijan: Tovuz, Leg. F. Dépalle | |
| I. cedretii | 16.83 (0.5) | 3 | 2 | 20 * | 1 | Lebanon: Bcharre, 1900 m | |
| I. damascene | 16.44 (0.04) | 3 | 2 | 20 | 1 | Syria: Damas, Leg. F. Dépalle | |
| I. haynei var. jordana | 16.20 (0.52) | 3 | 2 | 20 | 1 | Jordania: Umm Qais, Leg. F. Dépalle | |
| I. iberica subsp. elegantissima | 17.89 (0.18) | 1 | 1 | 20 | 1 | Turkey: 2200 m (RBGK 1999-4347) | |
| I. iberica subsp. iberica | 17.25 (0.05) | 1 | 1 | 20 | 1 | Unknown (RBGK 2002-2632) | |
| I. kirkwoodiae subsp. kirkwoodiae | 17.83 (0.06) | 1 | 1 | 20 | 1 | Turkey (RBGK 1994-2407) | |
| I. kirkwoodiae | 17.25 (0.15) | 2 | 2 | 20 | 1 | Syria: St. Simeon, Leg. F. Dépalle | |
| I. lortetii | 16.53 (0.19) | 3 | 2 | 20 * | 1 | Lebanon: Mays el Jabal, 640 m | |
| I. lortetii | 16.38 (0.29) | 2,3 | 2 | 20 | 1 | Israel: Ayelet-Hashahar, Leg. F. Dépalle | |
| I. mariae | 15.34 (0.5) | 3 | 2 | 20 | 1 | Israel: Sede Boker, Leg. F. Dépalle | |
| I. meda | 15.21 (0.49) | 3 | 2 | 20 | 1 | Iran: Baqloujeh Sardar, Leg. F. Dépalle | |
| I. mirabilis | 15.69 (0.10) | 2 | 2 | 20 | 2 | Iran: Ayerandibi, Leg. F. Dépalle | |
| I. nigricans | 16.68 * | 3 | 2 | 20 | 1 | Palestine: Bani Naim, Leg. Dr. Khalid Sawalha | |
| I. paradoxa | 17.32 (0.08) | 1 | 1 | 20 | 2 | Armenia (RBGK 1977-4470) | |
| I. petrana | 18.17 (0.11) | 1 | 1 | 20 | 1 | Unknown (RBGK 1990-3180) | |
| I. petrana | 18.01 (0.48) | 2 | 2 | 20 | 1 | Jordania: Shawbak, Leg. F. Dépalle | |
| I. samariae | 16.56 (0.04) | 3 | 2 | 20 | 1 | Israel: Majdal, Leg. F. Dépalle | |
| I. sari | 18.03 (0.11) | 1 | 1 | 20 | 1 | Unknown (RBGK 2011-1955) | |
| I. schelkovnikowii | 16.11 (0.04) | 2 | 2 | - | 1 | Azerbaijan: Mingachevir, Leg. F. Dépalle | |
| I. sofarana subsp. kasruwana | 16.68 (0.53) | 3 | 2 | 20 * | 1 | Lebanon: Ehmej, 1217 m | |
| I. sofarana subsp. sofarana | 16.22 (0.37) | 3 | 2 | 20 * | 1 | Lebanon: Dahr El-Baydar, 1640 m | |
| I. sofarana subsp. sofarana | 16.90 (0.49) | 3 | 2 | 20 * | 1 | Lebanon: Hazzerta, 1530 m | |
| I. sprengeri | 19.20 (0.16) | 1 | 1 | - | 1 | Turkey: 1000 m (RBGK 2011-1958) | |
| I. westii | 16.00 (0.75) | 3 | 2 | 20 * | 1 | Lebanon: Tawmet Jezzine, 1300 m | |
| I. yeruchamensis | 16.54 (0.05) | 3 | 2 | - | 1 | Israel: Yerocham, Leg. F. Dépalle | |
| I. yebrudii subsp. yebrudii | 16.20 (0.11) | 3 | 2 | 20 | 1 | Syria: Yebrud, Leg. F. Dépalle | |
| Section Regelia | |||||||
| I. afghanica | 14.67 (0.47) | 3 | 2 | 22 | 3 | Afghanistan, Leg. F. Dépalle | |
| I. hoogiana | 30.31 (0.13) | 1 | 1 | - | 3 | Tajikistan (RBGK 2010-2098) | |
| I. korolkovii | 16.03 (0.03) | 4 | 1 | - | 3 | Unknown (RBGK 2010-2101) | |
| I. lineata | 29.77 (0.04) | 1 | 1 | - | 3 | Tajikistan (RBGK 1993-3285) | |
| I. stolonifera | 30.24 (0.11) | 1 | 1 | - | 3 | Pamir (RBGK 1984-91) | |
| Section Iris | |||||||
| I. albicans | 24.84 (0.65) | 3 | 2 | 44–48 | 3 | Lebanon: Aley, 880 m | |
| I. scariosa | 13.09 (0.18) | 1 | 1 | - | 3 | Kazakhstan (RBGK 2014-643) | |
| Section Psammiris | |||||||
| I. bloudovii | 10.66 (0.13) | 1 | 1 | 16, 26 | 3 | Siberia (RBGK 2004-2257) | |
| Subgenus Limniris | |||||||
| I. longipetala | 15.84 (0.19) | 2 | 2 | - | 3 | Iran: Jazvanaq, Leg. F. Dépalle | |
| Subgenus Scorpiris | |||||||
| I. persica | 20.99 (0.05) | 1 | 1 | 20, 36 | 3 | Turkey: 1200 m (RBGK 2014-1875) | |
| I. persica | 18.89 (0.32) | 3 | 2 | 24 * | 3 | Lebanon: Quaa, 700 m | |
| I. persica | 19.05 * | 3 | 2 | - | 3 | Lebanon: Yammouneh | |
| I. regis-uzziae | 22.80 (0.03) | 1 | 1 | 20 | 3 | Israel: 500 m (RBGK 1987-2212) | |
| Model | ||
|---|---|---|
| Phylogenetic Signal | Tempo of Trait Change | AIC |
| Distance | 27.922 | |
| Non phylogenetic | Equal | 22.243 |
| Free | 165.507 | |
| Distance | 25.527 | |
| Pure phylogenetic | Equal | 19.454 |
| Free | 165.702 | |
| Distance | 110.326 | |
| Punctuated | Equal | 32.232 |
| Free | 142.326 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samad, N.A.; Hidalgo, O.; Saliba, E.; Siljak-Yakovlev, S.; Strange, K.; Leitch, I.J.; Dagher-Kharrat, M.B. Genome Size Evolution and Dynamics in Iris, with Special Focus on the Section Oncocyclus. Plants 2020, 9, 1687. https://doi.org/10.3390/plants9121687
Samad NA, Hidalgo O, Saliba E, Siljak-Yakovlev S, Strange K, Leitch IJ, Dagher-Kharrat MB. Genome Size Evolution and Dynamics in Iris, with Special Focus on the Section Oncocyclus. Plants. 2020; 9(12):1687. https://doi.org/10.3390/plants9121687
Chicago/Turabian StyleSamad, Nour Abdel, Oriane Hidalgo, Elie Saliba, Sonja Siljak-Yakovlev, Kit Strange, Ilia J. Leitch, and Magda Bou Dagher-Kharrat. 2020. "Genome Size Evolution and Dynamics in Iris, with Special Focus on the Section Oncocyclus" Plants 9, no. 12: 1687. https://doi.org/10.3390/plants9121687
APA StyleSamad, N. A., Hidalgo, O., Saliba, E., Siljak-Yakovlev, S., Strange, K., Leitch, I. J., & Dagher-Kharrat, M. B. (2020). Genome Size Evolution and Dynamics in Iris, with Special Focus on the Section Oncocyclus. Plants, 9(12), 1687. https://doi.org/10.3390/plants9121687