1. Introduction
Plants can be a large source of biocidal compounds potentially suitable to formulate new pesticides for sustainable management of plant pathogens and pests, also including phytoparasitic nematodes [
1].
Presence of a variety of nematicidal phytochemicals in many botanical families has increasingly focused the attention of scientists and farmers to plant-derived nematicidal products [
2,
3]. Previous studies of our research group documented a strong activity on phytoparasitic nematodes for chemical constituents of extracts from Asteraceae and Brassicaceae plants, such as phenolics and glucosinolates [
4,
5], as well as for the essential oils from many aromatic and medicinal plants [
6,
7].
Saponins represent a wide group of specialized phytochemicals, consisting of a triterpene or steroid aglycone to which one or more sugar chains are attached, present in many plant families but particularly abundant in the Fabaceae plants [
8]. Saponins were acknowledged for a wide range of biological activities, including a cytotoxic, antibiotic, anti-inflammatory and molluscicidal activity [
8,
9]. In addition, a high anthelmintic activity against gastrointestinal nematodes from donkey [
10] and goats [
11] was also proved for alfalfa (
Medicago sativa L.) saponin mixtures.
The activity of saponins was also demonstrated on phytoparasitic nematodes, as affecting juvenile (
J2) motility or egg and
J2 viability of the root-knot nematodes
Meloidogyne incognita Kofoid et White (Chitw) [
12] and
M. javanica Treub [
13]. Moreover, saponin-rich extracts from
Quillaja saponaria Molina were found to have significant nematicidal effects on other economically relevant phytonematode species, such as the dagger nematode
Xiphinema index Thorne et Allen and the root lesion nematode
Pratylenchus thornei Sher et Allen [
14], as well as significantly reducing
M. incognita infestation on field tomato (
Solanum lycopersicum L.) or melon (
Cucumis melo L.) crops [
15].
The
Medicago genus (Fabaceae family) contains 83 different herbaceous or shrub plant species, mainly distributed around the Mediterranean basin but also adapted to a large range of environmental conditions, which produce several specialized metabolites such as coumarins, flavonoids, naphtoquinones, alkaloids and also saponins [
8,
16,
17,
18,
19].
Saponins from
Medicago spp. are formed by complex mixtures of high molecular weight triterpene glycosides with medicagenic acid, hederagenin, zahnic acid, bayogenin and soyasapogenols A and B as the dominant aglycones [
20,
21].
Medicago saponins have been reported to possess a large spectrum of biological and pharmacological effects, such as cytotoxic, antitumor, fungicidal, molluscicidal, antibacterial and antiviral activities [
8,
22,
23,
24].
Previous studies of our research group assessed also the nematotoxic potential of active saponins obtained from various
Medicago species. Argentieri et al. (2008) [
25] described the nematicidal activity of saponins and derived prosapogenins and sapogenins from
M. arborea,
M. arabica and
M. sativa against
X. index, as well as in vitro bioassays demonstrated the activity of saponin mixtures from
M. sativa against
X. index,
M. incognita and the potato cyst nematode
Globodera rostochiensis Wollenweber [
26]. The species
M. heyniana Greuter,
M. hybrida (Pourr.) Trautv.,
M. lupulina L.,
M. murex Willd. and
M. truncatula Gaertn. are also widespread as fodder plants throughout the Mediterranean basin but, adversely to
M. sativa, their biological activities were scarcely documented. The saponin content of these species suggests that they may also possess nematicidal properties exploitable for the formulation of new nematicidal products Therefore, a study was carried out to investigate the nematicidal activity of saponin-rich extracts from these five
Medicago species against the phytoparasitic nematodes
M. incognita,
X. index and
G. rostochiensis, as well as to assess the suppressiveness of soil amendments with their plant material to
M. incognita on tomato.
3. Discussion
The saponin extract of the five
Medicago species was demonstrated as strongly active on
X. index and, except for
M. hybrida, on
M. incognita at concentrations above 250 µg mL
−1, as confirming the almost complete mortality previously reported for a 16 or 24 h exposure of
M incognita J2 and
X. index females to a 500 μg mL
−1 solution of
M. sativa saponins [
26]. As in this cited study,
G. rostochiensis J2 sensitivity to the five saponin mixtures was higher than that of
M. incognita and
X. index specimens, but lower than that previously showed to
M. sativa saponins, which caused 40%–54% mortality even after a 4 and 8 h exposure, respectively, to a 125 μg mL
−1 concentration [
26]. Literature data on the activity of
Medicago saponins on phytoparasitic nematodes are almost exclusively limited to the previous studies of our research group, which proved a biocidal activity of saponins, prosapogenins and sapogenins from
M. arborea,
M. arabica and
M. sativa on
X. index [
25], as well as the activity of
M. sativa saponin mixtures against the same nematode species tested in this study [
26]. In addition, soil treatments with a crude extract of
M. sativa saponins resulted in a significant reduction of
M. incognita infestation on tomato [
33]. More generally, only few data have been reported on the nematicidal effects of saponins from other plants. A significant effect on the motility of
M. incognita J2 was reported for the saponins from
Asparagus spp. [
34], and saponin solutions were found to significantly reduce the number of
M.
javanica eggs and viable
J2 both in vitro and in soil [
13]. Furthermore, the total saponin fraction of
Portulaca oleracea L. and
Lantana camara L. strongly reduced the in vitro motility of
Meloidogyne spp.
J2, as well as almost completely inhibited gall formation on eggplant roots under greenhouse conditions [
35]. Adversely, the single saponin fraction of a
Q. saponaria extract showed a poor nematicidal effect on a range of phytoparasitic nematodes, including
X. index, the northern root-knot nematode
M. hapla Chitwood,
P. thornei,
Tylenchorhynchus sp. and
Helicotylenchus sp. [
14].
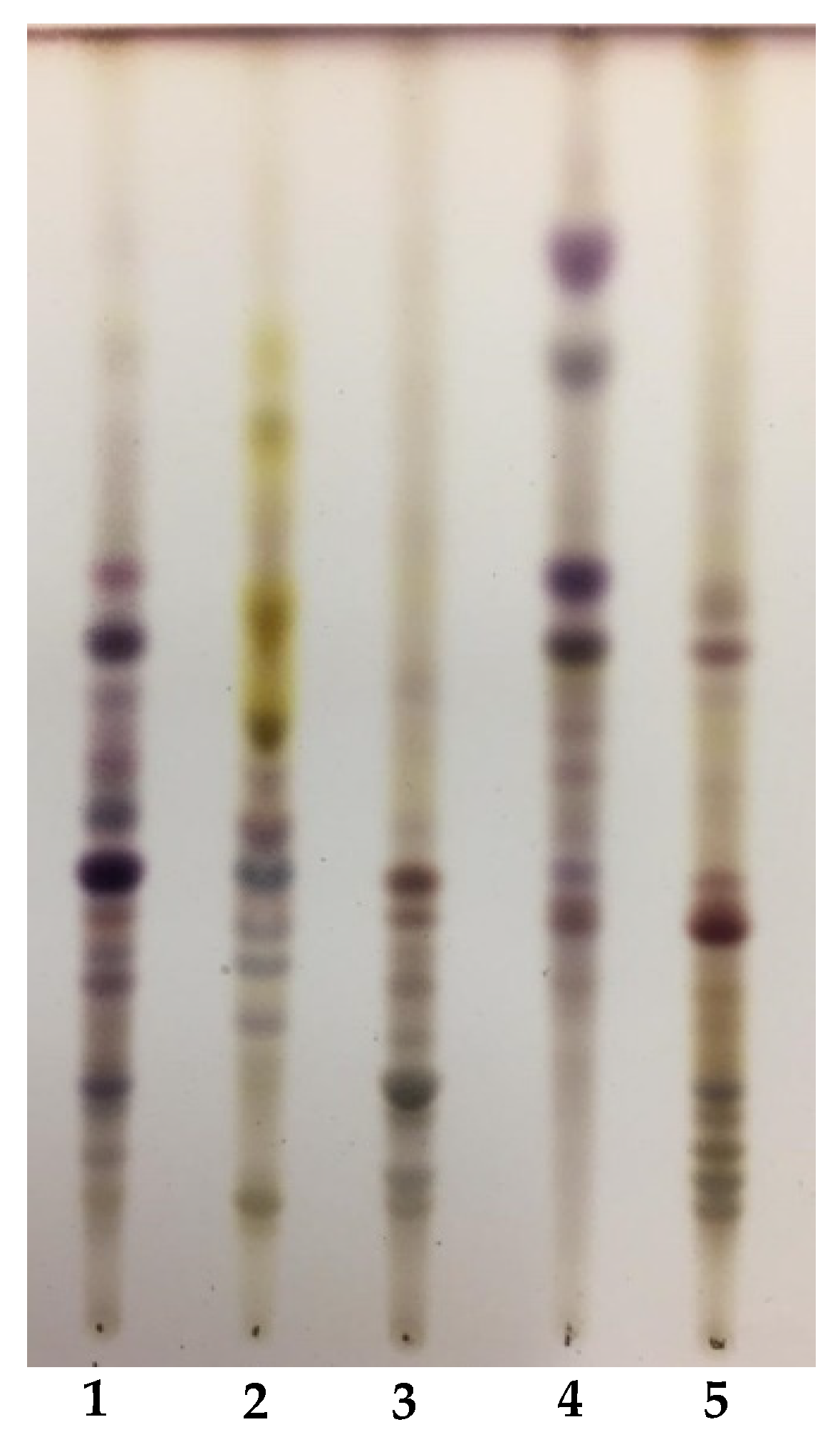
Crude saponin content of the five
Medicago species under investigation, as ranging from 0.60% of
M. truncatula to 1.62% of
M. heyniana, is in good agreement with published data, which reported a crude saponin content of about 0.5%–1.5% dry matter in medics [
27,
36,
37,
38]. The high molecular weight compounds detected at higher amounts in
M. hybrida,
M. lupulina and
M. truncatula extracts, in particular saponins containing glycosides of medicagenic and zanhic acids, were previously identified in the aerial parts of
Medicago species [
8], including
M. arborea [
28],
M. sativa [
31],
M. truncatula [
37] and
M. marina [
32]. Given that zanhic acid is synthesized in the green parts of the plants [
39], its glycosides have not previously been found in
M. lupulina and
M. hybrida [
8,
38] when roots were investigated.
The biological activity of saponins is dependent on the number of side sugar chains attached to the sapogenins as well as to the nature of the sapogenin itself [
8]. Thus, previous in vitro investigations allowed to relate the nematoxic effect of
M. sativa saponins to the high amount of medicagenic acid. When pure aglycones have been used in in vitro bioassays, hederagenin was shown to be even more toxic than medicagenic acid and bayogenin against
X. index, while soyasaponin I, containing soyasapogenol B as a glycone, was the less-active saponin [
25]. This finding seems to be confirmed also in the present study by the nematotoxic properties against
X. index displayed by the three species
M. heyniana,
M. hybrida and
M. murex, which produce saponins with high amounts of the two aglycones hederagenin and bayogenin. In addition,
M. lupulina with a high amount of medicagenic acid was particularly active against
M. incognita.
The mechanisms of the nematicidal activity of saponins are still not clearly elucidated, as different hypotheses were suggested by literature studies. Changes in cell permeability following the specific interaction of saponins with cell membranes were generally hypothesized as a cause of biological effects of these compounds [
8,
25]. More recently, Ibrahim and Srour [
33] observed a decrease of cholesterol in eggs of root-knot nematodes, according to a concentration effect relationship, following soil treatments with a
M. sativa saponin extract.
As suggested in our previous studies [
25], a specific interaction of saponins with the nematode cuticle ultrastructure may be also involved in their mechanism of activity. Based on this hypothesis, differences in the cuticle chemical components of the nematode species could account for the different response of the three nematode species tested in this study to the five saponin mixtures [
26].
Nevertheless, we have already noticed in a previous study [
5] that the nematicidal activity of plant extracts and/or pure metabolites may vary according to the nematode species and the life stage of the same nematode species. Thus, for example, we have already observed that the potato cyst nematode
G. rostiochiensis was more susceptible than
M. incognita to the toxic effect of
A. annua and its active compound, artemisinin [
5]. Consistently,
G. rostochiensis J2 was more sensitive to the five
Medicago extracts than
M. incognita and
X. index also in the present study.
The effect of the five saponin extracts on the hatchability of M. incognita and G. rostochiensis eggs was the exact opposite, as the percentage hatch of M. incognita eggs was significantly reduced and did not affect or even stimulate the hatchability of G. rostochiensis eggs. Effects of saponins on nematode egg hatchability were scarcely documented, as literature studies mainly described their effects on J2 motility and viability or on nematode infestation on host plants. The toxicity of the five saponin solutions to M. incognita eggs agrees with their effects on infective J2, whereas the stimulation of G. rostochiensis egg hatchability can be considered an unexpected result. An increased permeability of the cyst wall by the interaction with saponins may be hypothesized, though the exact biochemical mechanisms need to be specifically investigated.
The infestation of
M. incognita on tomato plants was significantly reduced by soil amendments with the dry biomass of the five
Medicago species, in full agreement with previous reports of a reduced infestation of
M. incognita both on potted and field tomato or of the carrot cyst nematode
Heterodera carotae Jones on field carrot following soil treatments with
M. sativa biomass [
40]. Moreover, previous studies also documented a significant suppression of phytoparasitic species
M. javanica,
Paratrichodorus sp. and
Criconemella xenoplax joined to an increase of beneficial free-living nematodes, in a soil amended with
M. sativa pelleted biomass [
41], as well as an activity against a wide range of soilborne fungal strains of soil amendments with
M. truncatula aerial parts [
37].
Results from the in vitro experiments indicated that saponins from plant tissues of the five
Medicago species are surely involved in the strong root-knot nematode suppression by soil incorporation of these plant materials. However, as already remarked in our previous studies [
25,
26], further contributory mechanisms should be also hypothesized, such as the nematoxicity of other bioactive metabolites from
Medicago plants [
42], an ammonia release by the decomposition of
Medicago biomasses in soil [
43,
44] or an increase of phytonematode-suppressive microorganisms on the favorable substrate represented by
Medicago tissues [
45].
In particular it is known that
Medicago species also synthesize polyphenolics such as glycosyl derivatives of apigenin, luteolin, chrysoeriol and tricin in
M. truncatula [
19,
46] and in
M. sativa [
47,
48]. Several biological functions related with plant structural protection, regulation of plant environmental communication and control of plant physiological events have been ascribed to plant polyphenolics. It has also been shown that they have a role as plant defense compounds against a range of microorganisms and that they are involved in plant–nematode interactions acting as defense compounds [
3,
49,
50]. Thus, the presence of this type of metabolite in the biomass can reasonably be involved in the regulation of plant–nematode interactions when administered in soil to infested tomato plants.
The improved tomato growth in soil amended with the five
Medicago species plant material also agrees with our previous reports of a biostimulating effect of soil incorporation with
M. sativa biomass [
42]. As previously noted for
M. sativa, the growth effect of
Medicago amendments may be related not only to the reduced nematode infestation but also to the physiological role of other specialized
Medicago metabolites as well as to a general improvement of soil physical, chemical and microbiological properties such as an increased nitrogen content [
51].

 ,
, 



{kind=link}