Climate Change Enhanced Carotenoid Pro-Vitamin A Levels of Selected Plantain Cultivars

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Carotenoid Analysis
4. Results and Discussion
Carotenoid Contents Depending on Maturation and Seasonality
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Shetty, P. Addressing Micronutrient Malnutrition to Achieve Nutrition Security. In Combating Micronutrient Deficiencies: Food-based Approaches; Thompson, B., Amoroso, L., Eds.; CAB International: Wallingford, UK; FAO: Rome, Italy, 2011; pp. 28–40. [Google Scholar]
- Grieger, J.A.; Clifton, V.L. A review of the impact of dietary intakes in human pregnancy on infant birthweight. Nutrients 2014, 7, 153–178. [Google Scholar] [CrossRef] [Green Version]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; De, O.M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 427–451. [Google Scholar] [CrossRef]
- Ritchie, H.; Roser, M. Micronutrient Deficiency. Our World in Data. 2017. Available online: https://ourworldindata.org/micronutrient-deficiency#citation (accessed on 17 December 2019).
- Ghana Micronutrient Survey Final Report. 2017. Available online: https://osf.io/j7bp9/ (accessed on 19 March 2020).
- Melendez-Martinez, A.J.; Vicario, I.M.; Heredia, F.J. Nutritional importance of carotenoid pigments. Arch. Latinoam. Nutr. 2004, 54, 149–154. [Google Scholar]
- IFPRI (International Food Policy Research Institute). Global Nutrition Report: From Promise to Impact: Ending Malnutrition by 2030. 2016. Available online: www.data.unicef.org/wp-content/uploads/2016/06/130565-1.pdf (accessed on 17 December 2019).
- WHO (World Health Organization). 2009. Available online: http://www.who.int/nutrition/topics/vad/en/ (accessed on 28 January 2009).
- HarvestPlus. 2009. Available online: http://www.harvestplus.org/vita.html (accessed on 28 January 2018).
- Welch, R.M.; Graham, R.D. Agriculture: The real nexus for enhancing bioavailable micronutrients in food crops. J. Trace Elem. Med. Biol. 2005, 18, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genc, Y.; Humphries, J.M.; Lyons, G.H.; Graham, R.D. Exploiting genotypic variation in plant nutrient accumulation to alleviate micronutrient deficiency in populations. J. Trace Elem. Med. Biol. 2005, 18, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, S.; White, A.; Buttriss, J.L. Biofortified crops for tackling micronutrient deficiencies—What impact are these having in developing countries and could they be of relevance within Europe? Nutr. Bull. 2018, 319–357. [Google Scholar] [CrossRef]
- Gibson, R.S. Strategies for Preventing Multi-micro nutrient Deficiencies: A Review of Experiences with Food-based Approaches in Developing Countries. In Combating Micronutrient Deficiencies: Food-based Approaches; Thompson, B., Amoroso, L., Eds.; CAB International: Wallingford, UK; FAO: Rome, Italy, 2011; pp. 7–27. [Google Scholar]
- Gibson, R.S.; Anderson, V.P. A review of interventions based on dietary diversification/modification strategies with the potential to enhance intakes of total and absorbable zinc. Food Nutr. Bull. 2009, 30, S108–S143. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Safety and Nutritional Adequacy of Irradiated Food; WHO: Geneva, Switzerland, 1994. [Google Scholar]
- Lu, Q.Y.; Zhang, Y.; Wang, Y.; Wang, D.; Lee, R.P.; Gao, K.; Byrns, R.; Heber, D. California Hass avocado: Profiling of carotenoids, tocopherol, fatty acid, and fat content during maturation and from different growing areas. J. Agric. Food Chem. 2009, 57, 10408–10413. [Google Scholar] [CrossRef] [Green Version]
- Dhuique-Mayer, C.; Fanciullino, A.L.; Dubois, C.; Ollitrault, P. Effect of genotype and environment on citrus juice carotenoid content. J. Agric. Food Chem. 2009, 57, 9160–9168. [Google Scholar] [CrossRef]
- González-Molina, E.; Moreno, D.A.; García-Viguera, C. Genotype and harvest time influence the phytochemical quality of Fino lemon juice (Citrus limon (L.) Burm. F.) for industrial use. J. Agric. Food Chem. 2008, 56, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- FAO (Food and Agriculture Organization). 2009. Available online: http://faostat.fao.org/site/567/default.aspx#ancor (accessed on 28 January 2009).
- Global Trade. Africa’s Plantain Market to Reach Over 30M Tonnes by 2025. IndexBox AI Platform. 2019. Available online: https://www.globaltrademag.com/africas-plantain-market-to-reach-over-30m-tonnes-by-2025 (accessed on 12 January 2020).
- Englberger, L.; Aalbersberg, W.; Ravi, P.; Bonnin, E.; Marks, G.C.; Fitzgerald, M.H.; Elymore, J. Further analyses on Micronesian banana, taro, breadfruit and other foods for provitamin A carotenoids and minerals. J. Food Compos. Anal. 2003, 16, 219–236. [Google Scholar] [CrossRef]
- Englberger, L.; Schierle, J.; Marks, G.C.; Fitzgerald, M.H. Micronesian banana, taro, and other foods: Newly recognized sources of provitamin A and other carotenoids. J. Food Compos. Anal. 2003, 16, 3–19. [Google Scholar] [CrossRef]
- Englberger, L.; Schierle, J.; Aalbersberg, W.; Hofmann, P.; Humphries, J.; Huang, A.; Lorens, A.; Levendusky, A.; Daniells, J.; Marks, G.C.; et al. Carotenoid and vitamin content of Karat and other Micronesian banana cultivars. Int. J. Food Sci. Nutr. 2006, 57, 399–418. [Google Scholar] [CrossRef] [PubMed]
- Englberger, L.; Wills, R.B.H.; Blades, B.; Dufficy, L.; Daniells, J.W.; Coyne, T. Carotenoid content and flesh color of selected banana cultivars growing in Australia. Food Nutr. Bull. 2006, 27, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.W.; Garming, H.; Ekesa, B.; Roux, N.; Van den Bergh, I. Exploiting banana biodiversity to reduce vitamin A deficiency-related illness: A fast and cost-effective strategy. In Proceedings of the Tropical Fruits in Human Health and Nutrition Conference, Brisbane, Australia, 5–8 November 2008. in press. [Google Scholar]
- Davey, M.W.; Van den Bergh, I.; Markham, R.; Swennen, R.; Keulemans, J. Genetic variability in Musa fruit provitamin A carotenoids and mineral micronutrient contents. Food Chem. 2009, in press. [Google Scholar] [CrossRef]
- Davey, M.W.; Saeys, W.; Hof, E.; Ramon, H.; Swennen, R.; Keulemans, J. Application of near- infrared reflectance spectroscopy (NIRS) to the evaluation of carotenoids in Musa fruit pulp. J. Agric. Food Chem. 2009, 57, 1742–1751. [Google Scholar] [CrossRef]
- Garming, H.; Ekesa, B. An Ex-Ante Assessment of the Impact of Musa Cultivars with High Levels of Beta-Carotenes on the Burden of Vitamin A Deficiency Related Illness in Three Sub-Saharan Africa Countries; Bioversity International, Montpellier 2008.
- Khawas, P.; Das, A.J.; Sit, N.; Badwaik, L.S.; Deka, S.C. Nutritional Composition of Culinary Musa ABB at Different Stages of Development. Am. J. Food Sci. Technol. 2014, 2, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Adepoju, O.T.; Sunday, B.E.; Folaranmi, O.A. Nutrient composition and contribution of plantain (Musa paradisiacea) products to dietary diversity of Nigerian consumers. Afr. J. Biotechnol. 2012, 11, 13601–13605. [Google Scholar] [CrossRef]
- Egbebi, A.O.; Bademosi, T.A. Chemical compositions of ripe and unripe banana and plantain. Int. J. Trop. Med. Public Health 2011, 1, 1–5. [Google Scholar]
- Davey, M.V.; Stals, E.; Ngoh-Newilah, G.; Tomekpe, K.; Lusty, C.; Markham, R.; Swennen, R.; Keulemans, J. Sampling strategies and variability in fruit pulp micronutrient contents of West and Central African bananas and plantains (Musa species). J. Agric. Food Chem. 2007, 55, 2633–2644. [Google Scholar] [CrossRef] [PubMed]
- Ebi, K.l.; Ziska, L.H. Increases in atmospheric carbon dioxide: Anticipated negative effects on food quality. PLoS Med. 2018, 15, e1002600. [Google Scholar] [CrossRef] [PubMed]
- Pilbeam, D.J. Breeding crops for improved mineral nutrition under climate change conditions. J. Exp. Bot. 2015, 66, 3511–3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P.; Pintado, M.M.; Vasconcelos, M.W. Preserving the nutritional quality of crop plants under a changing climate: Importance and strategies. Plant Soil 2019. [Google Scholar] [CrossRef] [Green Version]
- Björn, L.O.; Widell, S.; Wang, T. Evolution of UV-B regulation and protection in plants. Adv. Space Res. 2002, 30, 1557–1562. [Google Scholar] [CrossRef]
- Sullivan, J.H.; Teramura, A.H. Field study of the interaction between solar ultraviolet-B radiation and drought on photosynthesis and growth of soybean. Plant Physiol. 1990, 92, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Gwynn-Jones, D. Short-term impacts of enhanced UV-B radiation on photoassimilate allocation and metabolism: A possible interpretation for time-dependent inhibition of growth. Plant Ecol. 2001, 154, 67–73. [Google Scholar] [CrossRef]
- Kyparissis, A.; Drilias, P.; Petropoulou, Y.; Grammatikopoulos, G.; Manetas, Y. Effects of UV-B radiation and additional irrigation on the Mediterranean evergreen sclerophyll Ceratonia siliqua L. under field conditions. Plant Ecol. 2001, 154, 189–193. [Google Scholar] [CrossRef]
- Eichhorn, M.; Dohler, M.; Austen, H. Impact of UV-B radiation on photosynthetic electron transport of Wolffa arrhiza (L) Wimm. Photosynthetica 1993, 39, 613–618. [Google Scholar]
- Reed, J.; Van de Staaij, J.; Bjorn, L.O.; Caldwell, M.M. UV-B as an environmental factor in plant life: Stress and regulation. Trends Ecol. Evol. 1992, 12, 22–28. [Google Scholar]
- Barnes, P.W.; Maggard, S.; Holman, S.R.; Vergara, B.S. Interspecific variation in sensitivity to UV-B radiation in rice. Crop Sci. 1993, 33, 1041–1046. [Google Scholar] [CrossRef]
- Correia, C.M.; Areal, E.L.V.; Torres-Pereira, M.S.; Torres-Pereira, J.M.G. Interspecific variation in sensitivity to UV-B radiation in maize grown under field conditions. I growth and morphological aspects. Field Crops Res. 1998, 59, 81–89. [Google Scholar]
- Wald, J.P.; Nohr, D.; Biesalski, H.K. Rapid and easy carotenoid quantification in Ghanaian starchy staples using RP-HPLC-PDA. J. Food Compos. Anal. 2018, 67, 119–127. [Google Scholar] [CrossRef]
- Shen, J.; Jiang, C.Q.; Yan, Y.F.; Liu, B.R.; Zu, C. Effect of increased UV-B radiation on carotenoid accumulation and total antioxidant capacity in tobacco (Nicotiana tabacum L.) leaves. Genet Mol. Res. 2017, 16, 1–11. [Google Scholar] [CrossRef]
- Mark, U.; Tevini, M. Effects of solar ultraviolet -B radiation, temperature and carbon dioxide on growth and physiology of sunflower and maize seedlings. Plant Ecol. 1997, 128, 224–234. [Google Scholar] [CrossRef]
- Pérez, C.P.; Ulrichs, C.; Huyskens-Keil, S.; Schreiner, M.; Krumbein, A.; Schwarz, D.; Kläring, H.-P. Composition of Carotenoids in Tomato Fruits as Affected by Moderate UV-B Radiation before Harvest. Acta Hort. 2009, 821, 217–222. [Google Scholar] [CrossRef]
- Hu, L.; Yang, C.; Zhang, L.; Feng, J.; Xi, W. Irradiation on the Soluble Sugar, Organic Acid, and Carotenoid Content of Postharvest Sweet Oranges (Citrus sinensis (L.) Osbeck). Molecules 2019, 24, 3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedina, I.; Hidema, J.; Velitchkova, M.; Georgieva, K.; Nedeva, D. UV-B induced stress responses in three rice cultivars. Biol. Plant 2010, 54, 571–574. [Google Scholar] [CrossRef]
- Mpoloka, S.W. Effects of prolonged UV-B exposure in plants. Afr. J. Biotechnol. 2008, 7, 4874–4883. [Google Scholar]
- Kramer, G.F.; Norman, H.A.; Krizek, D.T.; Mirecki, R.M. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Lidon, F.C.; Ramalho, J.C. Impact of UV-B irradiation on photosynthetic performance and chloroplast membrane components in Oryza sativa L. J. Photochem. Photobiol. B Biol. 2011, 104, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.H. Effects of increasing UV-B radiation and atmospheric carbon dioxide on photosynthesis and growth implications for terrestrial ecosystems. Plant Ecol. 1997, 128, 194–206. [Google Scholar] [CrossRef]
- Hunt, J.E.; McNeil, D.L. Nitrogen status affects UV-B sensitivity of cucumber. Aust. J. Plant Physiol. 1998, 25, 79–86. [Google Scholar] [CrossRef]
- Correia, C.M.; Coutinho, J.F.; Bjorn, L.O.; Torres-Pereira, J.M.G. Ultravoilet-B radiation and nitrogen effects on growth and yield of maize under Mediterranean field conditions. Eur. J. Agron. 2000, 12, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Musil, C.F.; Chimphango, S.B.M.; Dakora, F.D. Effects of ultraviolet-B radiation on native and cultivated plants of southern Africa. Ann. Bot. 2002, 90, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Adel, A.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- De Pee, S.; West, C.E. Dietary carotenoids and their role in combating vitamin A deficiency: A review of the literature. Eur. J. Clin. Nutr. 1996, 50, S38–S53. [Google Scholar]
- Klein, B.P.; Perry, A.K. Ascorbic acid and vitamin A activity in selected vegetables from different geographical areas of the United States. J. Food Sci. 1982, 47, 941–945. [Google Scholar] [CrossRef]
- Nguyen, C.T.T.; Lim, S.; Lee, J.G.; Lee, E.J. VcBBX, VcMYB21, and VcR2R3MYB transcription factors are involved in UV-B-induced anthocyanin. J. Agric. Food Chem. 2017, 65, 2066–2073. [Google Scholar] [CrossRef]
- Wang, H.; Gui, M.; Tian, X.; Xin, X.; Wang, T.; Li, J. Effects of UVB on vitamin C, phenolics, flavonoids and their related enzyme activities in mung bean sprouts (Vigna radiata). Int. J. Food Sci. Technol. 2017, 52, 827–833. [Google Scholar] [CrossRef]
- Wu, G.; Bornman, J.F.; Bennett, S.J.; Clarke, M.W.; Fang, Z.; Johnson, S.K. Individual polyphenolic profiles and antioxidant activity in sorghum grains are influenced by very low and high solar UV radiation and genotype. J. Cereal Sci. 2017, 77, 17–23. [Google Scholar] [CrossRef]
- Choudhary, K.K.; Agrawal, S.B. Assessment of fatty acid profile and seed mineral nutrients of two soybean (Glycine max L.) cultivars under elevated ultraviolet-B: Role of ROS, pigments and antioxidants. Photochem. Photobiol. 2016, 92, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.R.; Patro, H.; Lokhande, S.; Bellaloui, N.; Gao, W. Ultraviolet-B radiation alters soybean growth and seed quality. Food Nutr. Sci. 2016, 7, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, R.; Agrawal, S.B. Effect of supplemental UV-B on yield, seed quality, oil content and fatty acid composition of Brassica campestris L. under natural field conditions. Qual. Assur. Saf. Crops Foods 2016, 8, 11–20. [Google Scholar] [CrossRef]
- Pfündel, E.E. Action of UV and visible radiation on chlorophyll fluorescence from dark-adapted grape leaves (Vitis vinifera L.). Photosynth. Res. 2003, 75, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.M.H.; Al Watban, A.A.; Al-Fughom, A.T. Effect of ultraviolet radiation on chlorophyll, carotenoid, protein and proline contents of some annual desert plants. Saudi J. Biol. Sci. 2011, 18, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Razungles, A.; Gunata, Z.; Pinatel, S.; Baumes, R.; Bayonove, C. Etude quantitative de composés terpéniques, norisoprénoïdes et de leurs précurseurs dans diverses variétés de raisins. Sci. Aliment. 1993, 13, 59–72. [Google Scholar]
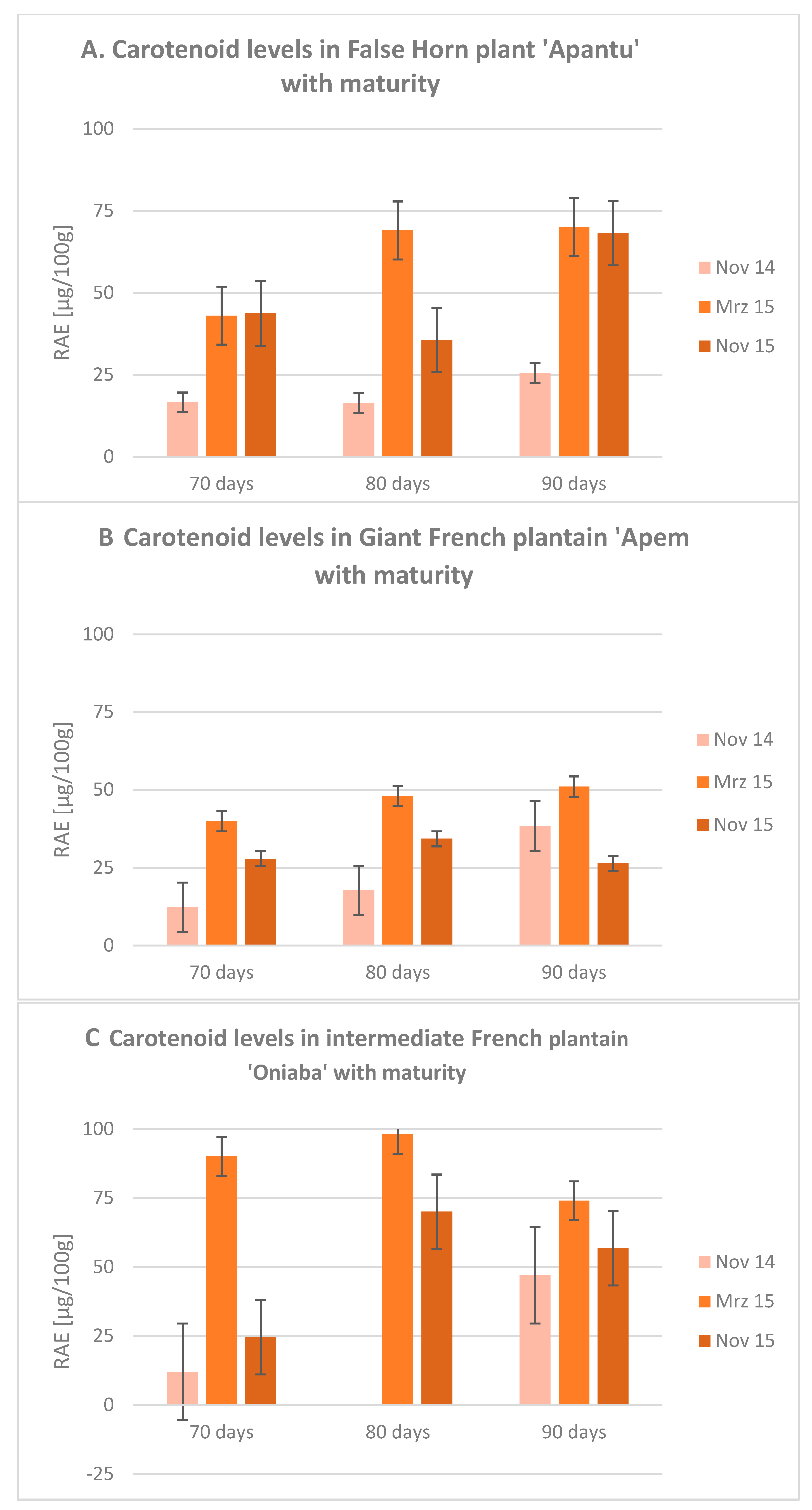

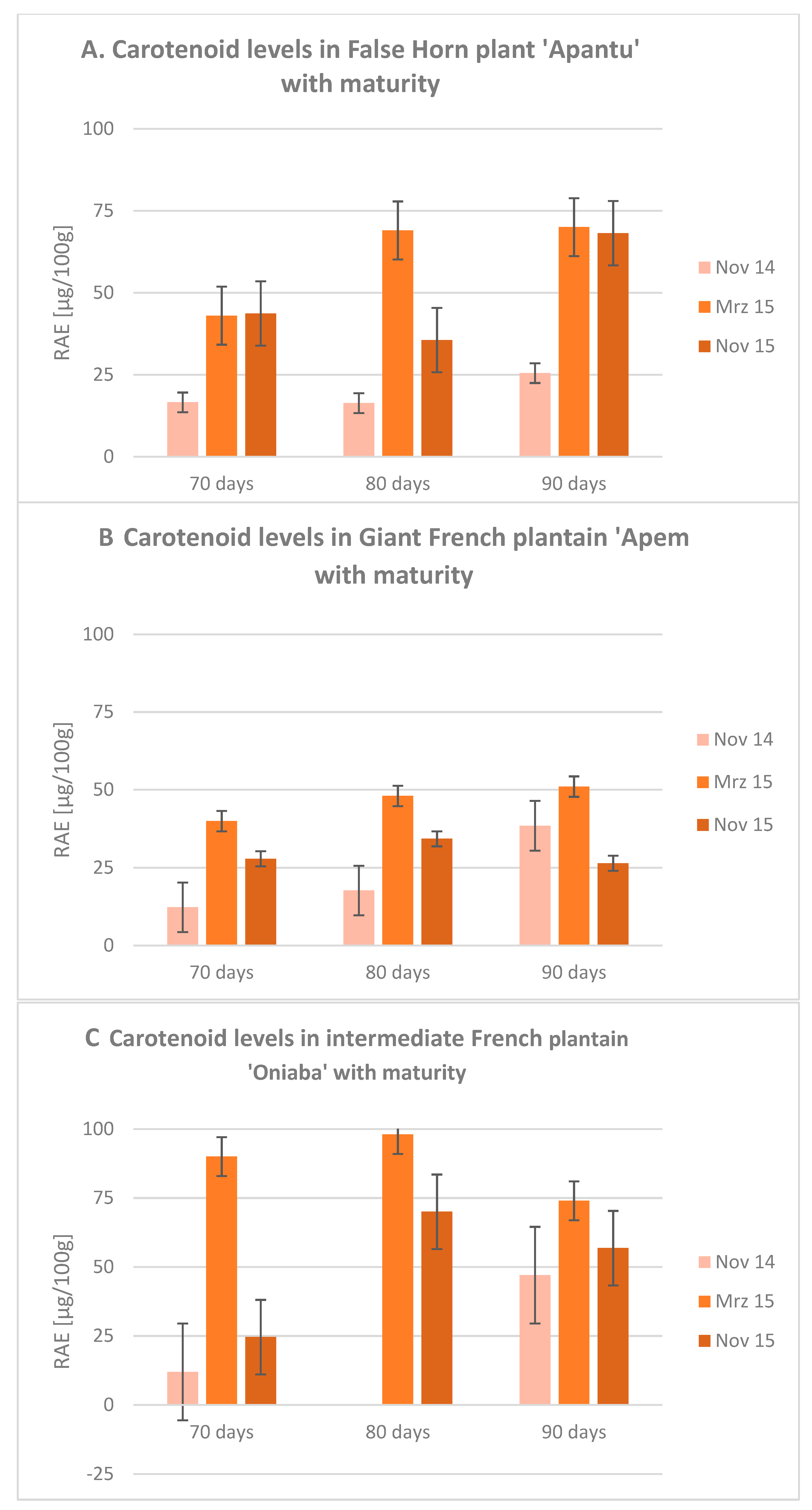{kind=link}
| Months | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sept | Oct | Nov | Dec |
| 2014 | 12 ± 0.1 | 12 ± 0.1 | 12 ± 0.01 | 12 ± 0.1 | 12 ± 0.1 | 7 ± 0.01 | 7 ± 0.01 | 8 ± 0.01 | 8 ± 0.01 | 8 ± 0.01 | 9 ± 0.01 | 12 ± 0.01 |
| 2015 | 12 ± 1.0 | 12 ± 0.1 | 12 ± 0.02 | 12 ± 0.1 | 12 ± 0.1 | 7 ± 0.01 | 7 ± 0.01 | 7 ± 0.01 | 8 ± 0.01 | 8 ± 0.01 | 9 ± 0.01 | 12 ± 0.01 |
| Cultivar | Maturity (Days) during the Rainy Season I | |||||
|---|---|---|---|---|---|---|
| Cultivar | α-carotene Levels µg/100 g Edible Pulp) at Three Maturities | β-carotene Levels µg/100 g Edible Pulp) at Three Maturities | ||||
| 70 | 80 | 90 | 70 | 80 | 90 | |
| “Apantu” | 95.7 ± 9.11a | 108 ± 13.8a | 171 ± 32.2a | 151 + 15.7a | 142 + 9.66 | 220 ± 29.2a |
| “Apem” | 78.6 ± 5.34b | 128 ± 4.31b | 291 ± 7.43b | 108 ± 9.3b | 148 + 9.25 | 386 ± 4.09b |
| “Oniaba” | 83.7 ± 12.4ab | - | 328 ± 57.7c | 102 ± 22.1b | - | 400 ± 86.4b |
| CV | 12.4 | 10.4 | 30.6 | 10.7 | 11.8 | 30.1 |
| LSD (p < 0.05) | 12.0 | 13.2 | 20.2 | 8.9 | 10.9 | 12.3 |
| Cultivar | Maturity (Days) During Rainy Season 11 | |||||
|---|---|---|---|---|---|---|
| α-carotene Contents at Three Maturities | β-carotene Content at Three Maturities | |||||
| 70 | 80 | 90 | 70 | 80 | 90 | |
| “Apantu” | 297 ± 38.1a | 254 ± 36.3a | 474 ± 45.0a | 362 ± 20.7a | 288 ± 29.4a | 567 ± 59.7a |
| “Apem” | 310 ± 3.26b | 356 ± 5.79b | 379 ± 2.12b | 221 ± 42.6b | 279 ± 39.3a | 210 ± 44.5b |
| “Oniaba” | 160 ± 9.77c | 533 ± 60.4c | 435 ± 4.70c | 209 ± 11.8b | 558 ± 65.4b | 451 ± 3.89c |
| CV | 15.1 | 20.4 | 40.6 | 10.7 | 11.8 | 45.1 |
| LSD (p < 0.05) | 9.3 | 15.2 | 12.2 | 12.9 | 11.9 | 15.3 |
| Maturity Date (Days) during the Dry Season | ||||||
|---|---|---|---|---|---|---|
| Cultivar | α-carotene Contents at Three Maturities | β-carotene Content at Three Maturities | ||||
| 70 | 80 | 90 | 70 | 80 | 90 | |
| “Apantu” | 283 ± 5.08a | 420 ± 2.80a | 489 ± 7.57a | 358 ± 5.24a | 577 ± 4.09a | 573 ± 7.79a |
| “Apem” | 310 ± 3.26a | 356 ± 5.79b | 379 ± 2.12b | 299 ± 2.31b | 371 ± 5.09b | 386 ± 4.09b |
| “Oniaba” | 719 ± 6.93b | 793 ± 8.39c | 542 ± 0.36c | 685 ± 7.63c | 744 ± 10.3c | 577 ± 2.75a |
| CV | 18.1 | 20.4 | 40.6 | 10.7 | 31.8 | 50.1 |
| LSD (p < 0.05) | 30.3 | 15.2 | 12.2 | 12.9 | 21.9 | 16.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzomeku, B.M.; Wald, J.P.; Wünsche, J.N.; Nohr, D.; Biesalski, H.K. Climate Change Enhanced Carotenoid Pro-Vitamin A Levels of Selected Plantain Cultivars. Plants 2020, 9, 541. https://doi.org/10.3390/plants9040541
Dzomeku BM, Wald JP, Wünsche JN, Nohr D, Biesalski HK. Climate Change Enhanced Carotenoid Pro-Vitamin A Levels of Selected Plantain Cultivars. Plants. 2020; 9(4):541. https://doi.org/10.3390/plants9040541
Chicago/Turabian StyleDzomeku, Beloved Mensah, Julian P. Wald, Jens Norbert Wünsche, Donatus Nohr, and Hans K. Biesalski. 2020. "Climate Change Enhanced Carotenoid Pro-Vitamin A Levels of Selected Plantain Cultivars" Plants 9, no. 4: 541. https://doi.org/10.3390/plants9040541
APA StyleDzomeku, B. M., Wald, J. P., Wünsche, J. N., Nohr, D., & Biesalski, H. K. (2020). Climate Change Enhanced Carotenoid Pro-Vitamin A Levels of Selected Plantain Cultivars. Plants, 9(4), 541. https://doi.org/10.3390/plants9040541