
Effect of Granzyme K, FasL and Interferon-γ Expression in Placentas with Preeclampsia
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Data
2.2. Tissue Processing
2.3. Double Immunofluorescence
2.4. RNA Isolation and RT-qPCR
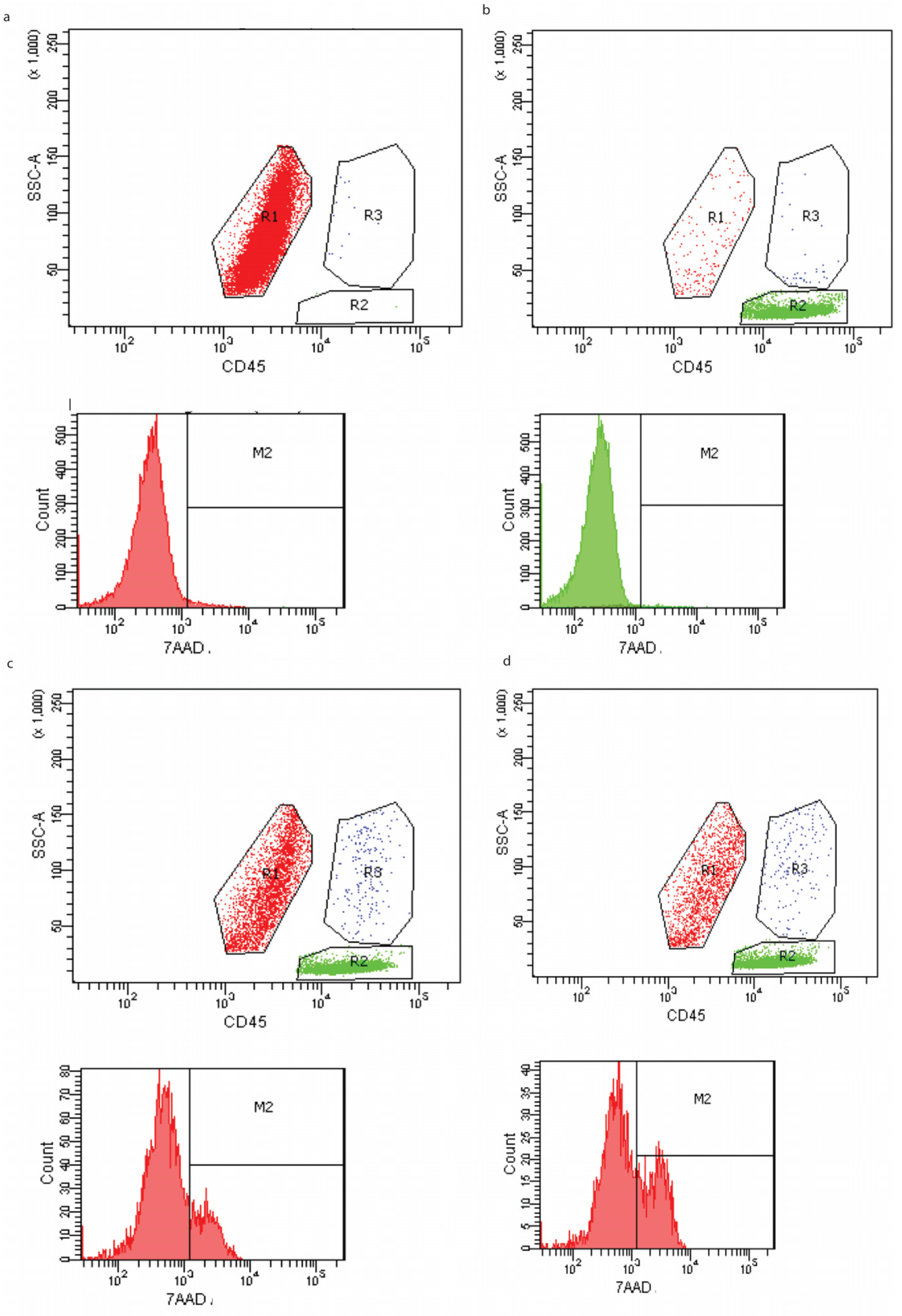
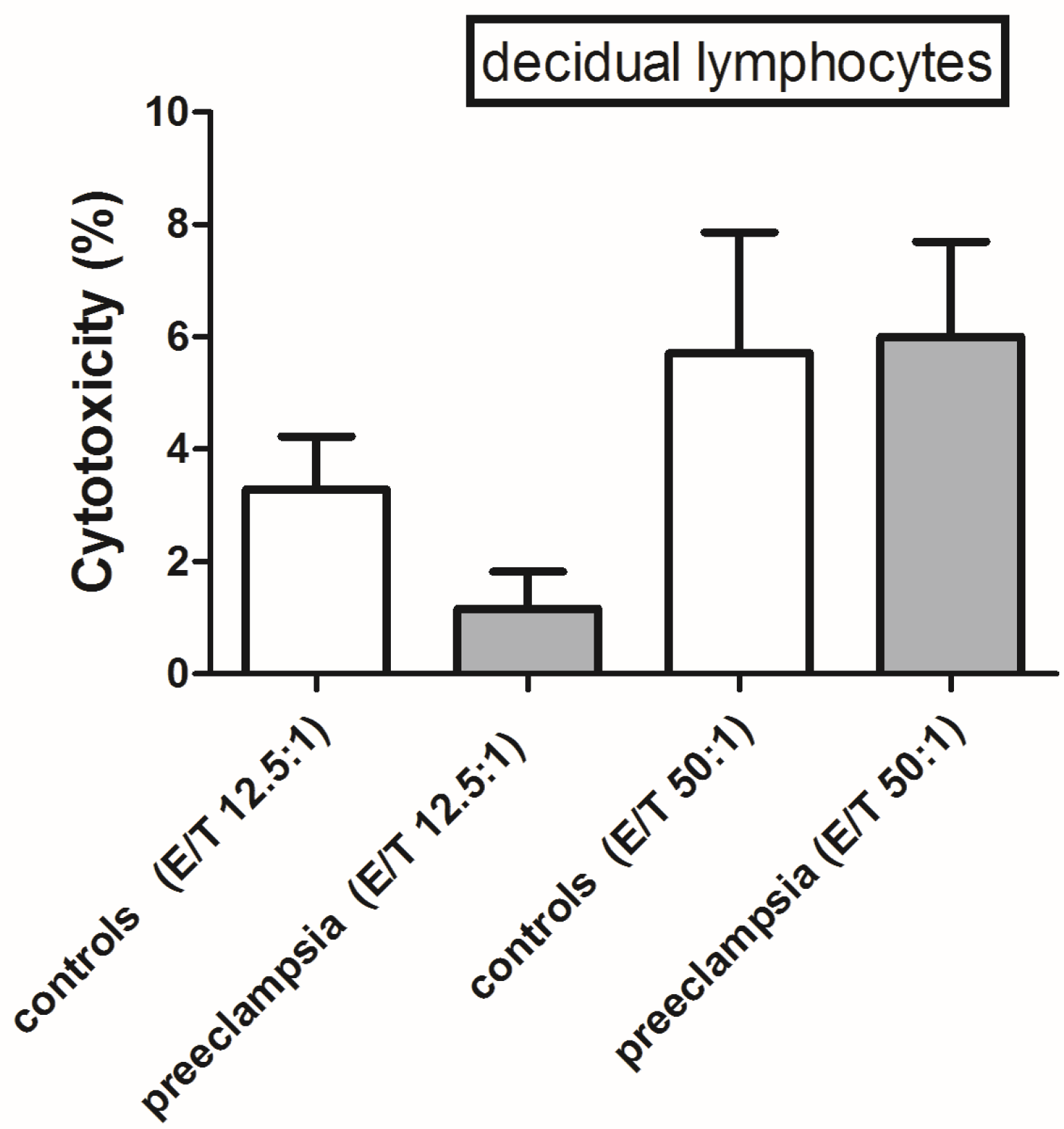
2.5. Cytotoxic Assay
2.6. Statistical Analysis
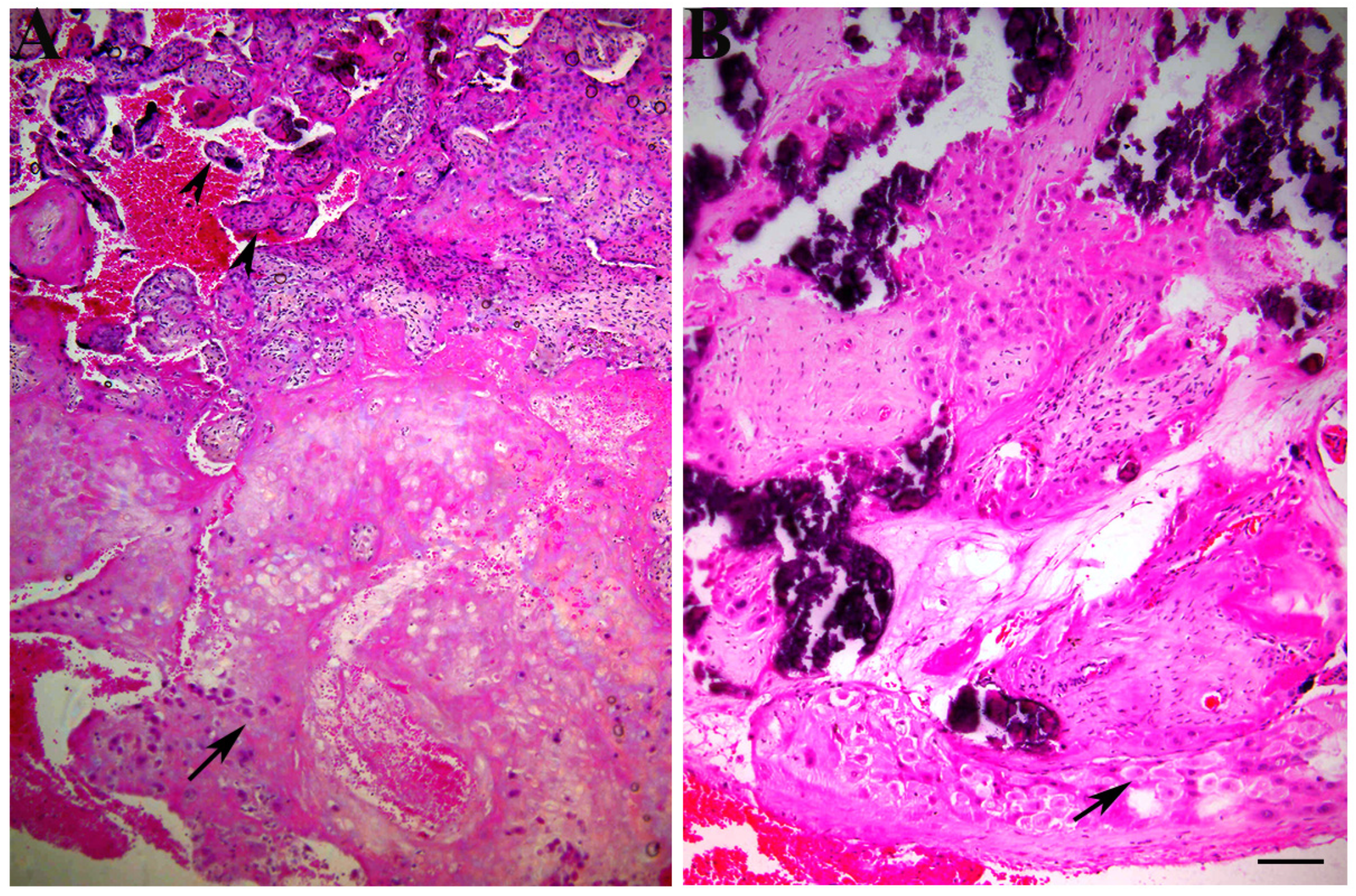
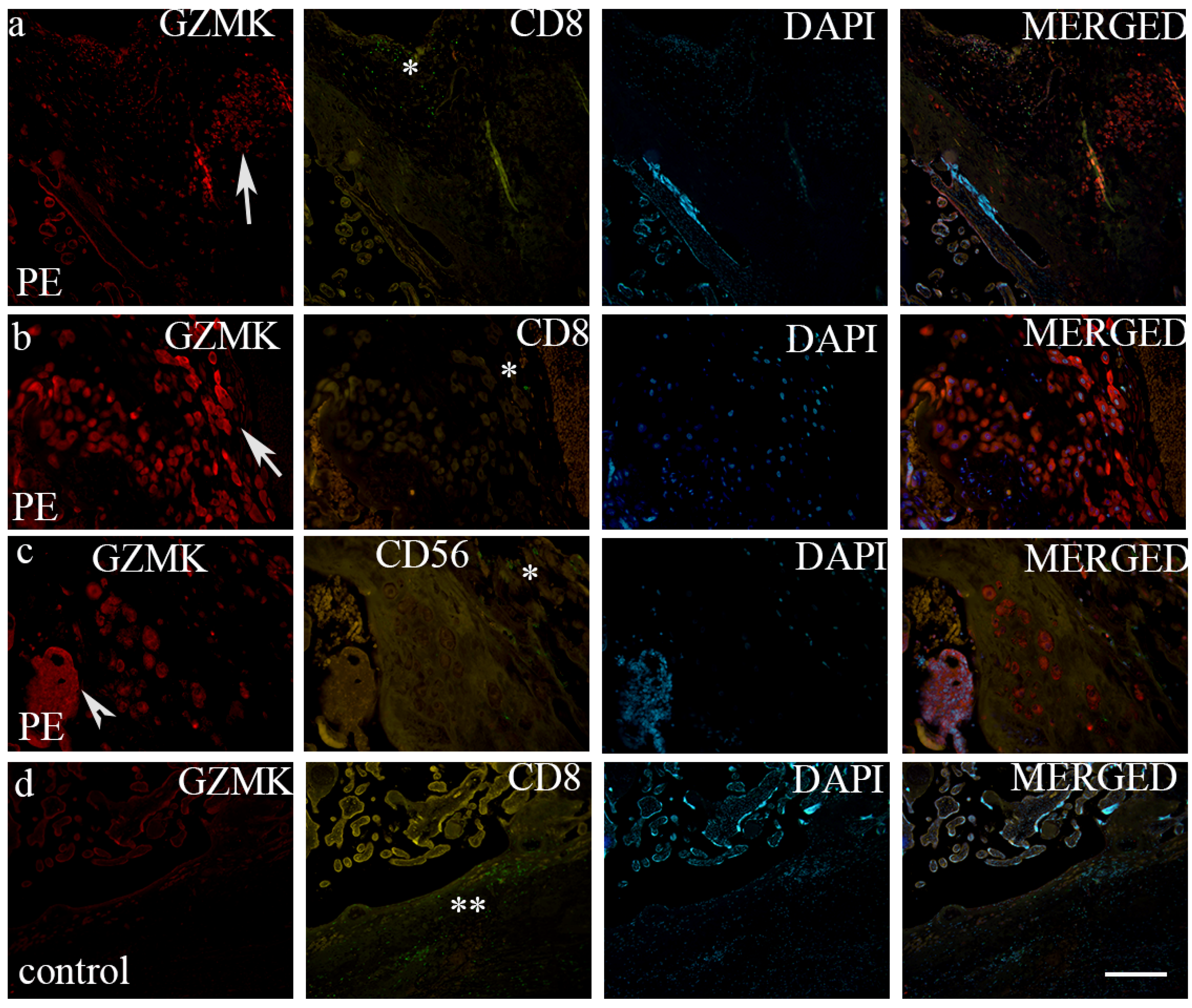
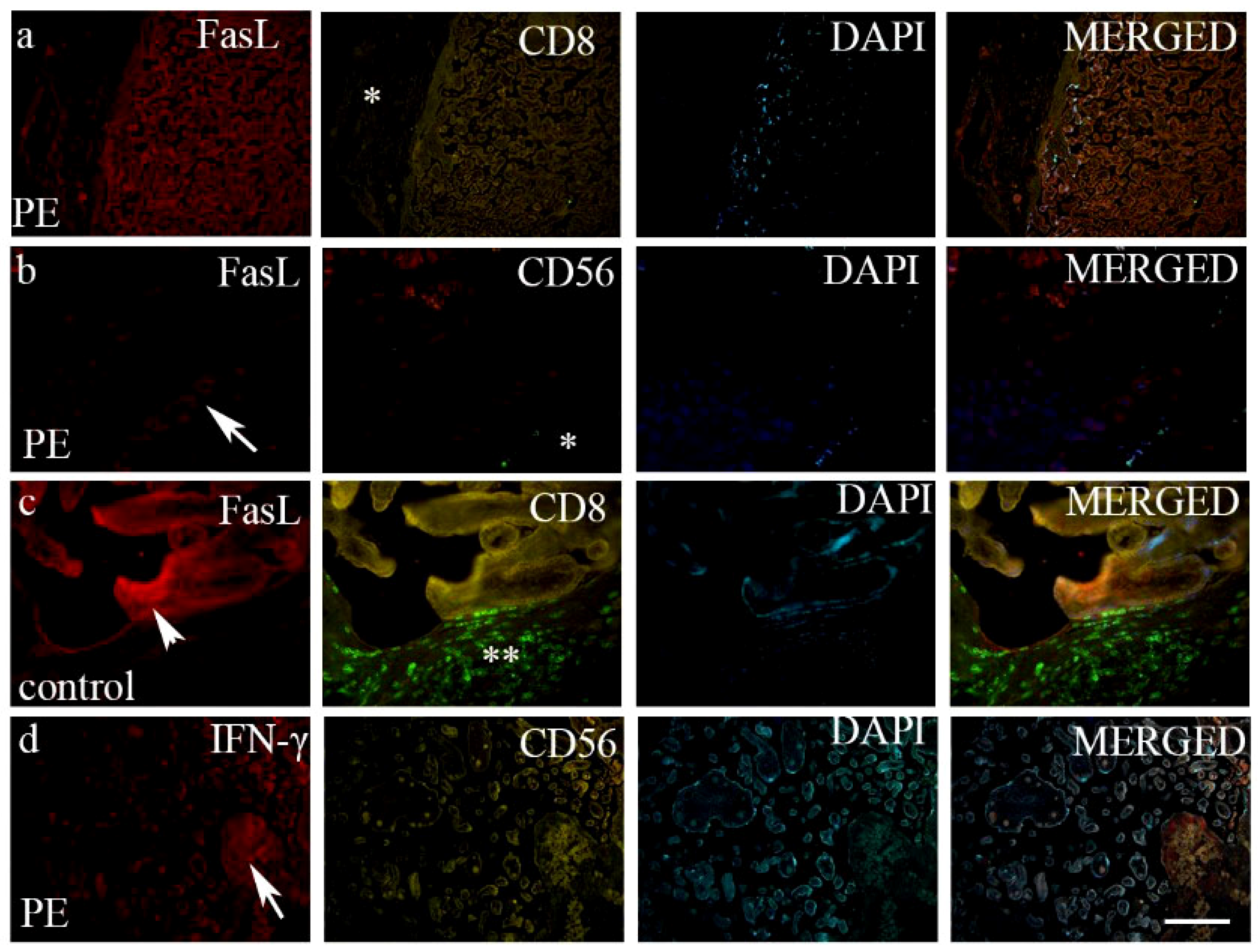
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farah, O.; Nguyen, C.; Tekkatte, C.; Parast, M.M. Trophoblast lineage-specific differentiation and associated alterations in preeclampsia and fetal growth restriction. Placenta 2020, 102, 4–9. [Google Scholar] [CrossRef]
- Pollheimer, J.; Knöfler, M. The role of the invasive, placental trophoblast in human pregnancy. Wien. Med. Wochenschr. 2012, 162, 187–190. [Google Scholar] [CrossRef]
- Wang, T.; Lian, Y. The relationship between Fas and Fas ligand gene polymorphism and preeclampsia risk. Biosci. Rep. 2019, 39, BSR20181901. [Google Scholar] [CrossRef]
- Soljic, V.; Barbaric, M.; Vukoja, M.; Curlin, M.; Orlovic Vlaho, M.; Cerni Obrdalj, E.; Lasic Arapovic, L.; Bevanda Glibo, D.; Vukojevic, K. Decreased Expression of Cytotoxic Proteins in Decidual CD8(+) T Cells in Preeclampsia. Biology 2021, 10, 1037. [Google Scholar] [CrossRef]
- Kohan-Ghadr, H.-R.; Kadam, L.; Jain, C.; Armant, D.R.; Drewlo, S. Potential role of epigenetic mechanisms in regulation of trophoblast differentiation, migration, and invasion in the human placenta. Cell Adhes. Migr. 2016, 10, 126–135. [Google Scholar] [CrossRef]
- Nelissen, E.C.; van Montfoort, A.P.; Dumoulin, J.C.; Evers, J.L. Epigenetics and the placenta. Hum. Reprod. Update 2011, 17, 397–417. [Google Scholar] [CrossRef]
- Nurzadeh, M.; Ghalandarpoor-Attar, S.M.; Ghalandarpoor-Attar, S.N.; Rabiei, M. The Role of Interferon (IFN)-γ in Extravillous Trophoblast Cell (EVT) Invasion and Preeclampsia Progression. Reprod. Sci. 2023, 30, 1462–1469. [Google Scholar] [CrossRef]
- Liu, H.; Wang, W.; Liu, C. Increased expression of IFN-γ in preeclampsia impairs human trophoblast invasion via a SOCS1/JAK/STAT1 feedback loop. Exp. Ther. Med. 2021, 21, 112. [Google Scholar] [CrossRef]
- Yang, Y.; Su, X.; Xu, W.; Zhou, R. Interleukin-18 and interferon gamma levels in preeclampsia: A systematic review and meta-analysis. Am. J. Reprod. Immunol. 2014, 72, 504–514. [Google Scholar] [CrossRef]
- Paul, S.; Lal, G. The Molecular Mechanism of Natural Killer Cells Function and Its Importance in Cancer Immunotherapy. Front. Immunol. 2017, 8, 1124. [Google Scholar] [CrossRef]
- Zhuang, B.; Shang, J.; Yao, Y. HLA-G: An Important Mediator of Maternal-Fetal Immune-Tolerance. Front. Immunol. 2021, 12, 744324. [Google Scholar] [CrossRef]
- Wei, X.; Yang, X. The central role of natural killer cells in preeclampsia. Front. Immunol. 2023, 14, 1009867. [Google Scholar] [CrossRef]
- Kieffer, T.E.C.; Laskewitz, A.; Vledder, A.; Scherjon, S.A.; Faas, M.M.; Prins, J.R. Decidual memory T-cell subsets and memory T-cell stimulatory cytokines in early- and late-onset preeclampsia. Am. J. Reprod. Immunol. 2020, 84, e13293. [Google Scholar] [CrossRef]
- Addison, E.G.; North, J.; Bakhsh, I.; Marden, C.; Haq, S.; Al-Sarraj, S.; Malayeri, R.; Wickremasinghe, R.G.; Davies, J.K.; Lowdell, M.W. Ligation of CD8alpha on human natural killer cells prevents activation-induced apoptosis and enhances cytolytic activity. Immunology 2005, 116, 354–361. [Google Scholar] [CrossRef]
- Barakonyi, A.; Miko, E.; Szereday, L.; Polgar, P.D.; Nemeth, T.; Szekeres-Bartho, J.; Engels, G.L. Cell death mechanisms and potentially cytotoxic natural immune cells in human pregnancies complicated by preeclampsia. Reprod. Sci. 2014, 21, 155–166. [Google Scholar] [CrossRef]
- Ren, W.G.; Kong, L.B.; Mi, H.M.; Zhao, S.X.; Zhang, Y.G.; Wang, R.Q.; Nan, Y.M. Activation of Fas/FasL and its downstream signaling pathway promotes development of alcoholic steatohepatitis and liver fibrosis in mice. Zhonghua Gan Zang Bing Za Zhi = Zhonghua Ganzangbing Zazhi = Chin. J. Hepatol. 2013, 21, 129–133. [Google Scholar] [CrossRef]
- Dong, Y.; Shen, X.; He, M.; Wu, Z.; Zheng, Q.; Wang, Y.; Chen, Y.; Wu, S.; Cui, J.; Zeng, Z. Activation of the JNK-c-Jun pathway in response to irradiation facilitates Fas ligand secretion in hepatoma cells and increases hepatocyte injury. J. Exp. Clin. Cancer Res. CR 2016, 35, 114. [Google Scholar] [CrossRef]
- Heidari, Z.; Mahmoudzadeh-Sagheb, H.; Sheibak, N.; Nourzaei, N. Quantitative changes of extravillous trophoblast cells in placentas of systemic lupus erythematosus patients. J. Obstet. Gynaecol. 2017, 37, 746–751. [Google Scholar] [CrossRef]
- Heidari, Z.; Mahmoudzadeh-Sagheb, H.; Sheibak, N. Quantitative changes of extravillous trophoblast cells in heavy smoker mothers compared with healthy controls. Reprod. Fertil. Dev. 2018, 30, 409–414. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Introduction: The Clans and Families of Cysteine Peptidases. In Handbook of Proteolytic Enzymes; Academic Press: Cambridge, MA, USA, 2013; pp. 1743–1773. [Google Scholar] [CrossRef]
- Chowdhury, D.; Lieberman, J. Death by a thousand cuts: Granzyme pathways of programmed cell death. Annu. Rev. Immunol. 2008, 26, 389–420. [Google Scholar] [CrossRef]
- Xu, Y.; Plazyo, O.; Romero, R.; Hassan, S.S.; Gomez-Lopez, N. Isolation of Leukocytes from the Human Maternal-fetal Interface. J. Vis. Exp. JoVE 2015, 99, e52863. [Google Scholar] [CrossRef]
- Godoy-Ramirez, K.; Franck, K.; Gaines, H. A novel method for the simultaneous assessment of natural killer cell conjugate formation and cytotoxicity at the single-cell level by multi-parameter flow cytometry. J. Immunol. Methods 2000, 239, 35–44. [Google Scholar] [CrossRef]
- Takahashi, H.; Yamamoto, T.; Yamazaki, M.; Murase, T.; Matsuno, T.; Chishima, F. Natural Cytotoxicity Receptors in Decidua Natural Killer Cells of Term Normal Pregnancy. J. Pregnancy 2018, 2018, 4382084. [Google Scholar] [CrossRef] [PubMed]
- Anft, M.; Netter, P.; Urlaub, D.; Prager, I.; Schaffner, S.; Watzl, C. NK cell detachment from target cells is regulated by successful cytotoxicity and influences cytokine production. Cell. Mol. Immunol. 2020, 17, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Toldi, G.; Rigo, J., Jr.; Stenczer, B.; Vasarhelyi, B.; Molvarec, A. Increased prevalence of IL-17-producing peripheral blood lymphocytes in pre-eclampsia. Am. J. Reprod. Immunol. 2011, 66, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Travis, O.K.; White, D.; Pierce, W.A.; Ge, Y.; Stubbs, C.Y.; Spradley, F.T.; Williams, J.M.; Cornelius, D.C. Chronic infusion of interleukin-17 promotes hypertension, activation of cytolytic natural killer cells, and vascular dysfunction in pregnant rats. Physiol. Rep. 2019, 7, e14038. [Google Scholar] [CrossRef] [PubMed]
- Warren, B.B.; Moyer, G.C.; Manco-Johnson, M.J. Hemostasis in the Pregnant Woman, the Placenta, the Fetus, and the Newborn Infant. Semin. Thromb. Hemost. 2023, 49, 319–329. [Google Scholar] [CrossRef]
- Karthikeyan, V.J.; Lip, G.Y.; Baghdadi, S.; Lane, D.A.; Beevers, D.G.; Blann, A.D. Soluble Fas and Fas ligand in pregnancy: Influence of hypertension. Angiology 2012, 63, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Strand, S.; Strand, D.; Seufert, R.; Mann, A.; Lotz, J.; Blessing, M.; Lahn, M.; Wunsch, A.; Broering, D.C.; Hahn, U.; et al. Placenta-derived CD95 ligand causes liver damage in hemolysis, elevated liver enzymes, and low platelet count syndrome. Gastroenterology 2004, 126, 849–858. [Google Scholar] [CrossRef]
- Shi, L.; Gong, H.M.; Ru, M.Y. Expressions and significances of Fas/FasL and Bcl-2 in placentae of cases with preeclampsia. Matern. Child Health Care China 2011, 26, 3291–3294. [Google Scholar]
- Resic Karara, J.; Zekic Tomas, S.; Marusic, J.; Roje, D.; Kuzmic Prusac, I. Fas and FasL expression in placentas complicated with intrauterine growth retardation with and without preeclampsia. J. Matern. Fetal Neonatal Med. 2016, 29, 1154–1159. [Google Scholar] [CrossRef]
- Banerjee, S.; Smallwood, A.; Moorhead, J.; Chambers, A.E.; Papageorghiou, A.; Campbell, S.; Nicolaides, K. Placental Expression of Interferon-γ (IFN-γ) and Its Receptor IFN-γR2 Fail to Switch from Early Hypoxic to Late Normotensive Development in Preeclampsia. J. Clin. Endocrinol. Metab. 2005, 90, 944–952. [Google Scholar] [CrossRef]
- Sheibak, N.; Mahmoudzadeh-Sagheb, H.; Moudi, B.; Heidari, Z. Elevated immunoexpression of interferon-gamma in placenta tissue samples from pregnancies complicated with preeclampsia compared to the placenta previa. Pregnancy Hypertens. 2020, 22, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Liao, A.-H. Quantitative abnormalities of fetal trophoblast cells in maternal circulation in preeclampsia. Prenat. Diagn. 2008, 28, 1160–1166. [Google Scholar] [CrossRef]
- Cemgil Arikan, D.; Aral, M.; Coskun, A.; Ozer, A. Plasma IL-4, IL-8, IL-12, interferon-gamma and CRP levels in pregnant women with preeclampsia, and their relation with severity of disease and fetal birth weight. J. Matern. Fetal Neonatal Med. 2012, 25, 1569–1573. [Google Scholar] [CrossRef]
- Kronborg, C.S.; Gjedsted, J.; Vittinghus, E.; Hansen, T.K.; Allen, J.; Knudsen, U.B. Longitudinal measurement of cytokines in pre-eclamptic and normotensive pregnancies. Acta Obs. Gynecol. Scand. 2011, 90, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Vafaei, H.; Faraji, S.; Ahmadi, M.; Tabei, S.M.B.; Fereidoni, S.; Shiravani, Z.; Hosseini, S.N.; Asadi, N.; Kasraeian, M.; Faraji, A.; et al. Alteration in IFN-gamma and CCL2 serum levels at first trimester of pregnancy contribute to development of preeclampsia and fetal growth restriction. Taiwan J. Obs. Gynecol. 2023, 62, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, M.; Lee, C.L.; Wong, V.H.H.; Wang, X.; Bai, K.; Wu, J.; Koistinen, H.; Seppala, M.; Lee, K.F.; Yeung, W.S.B.; et al. Decidual glycodelin-A polarizes human monocytes into a decidual macrophage-like phenotype through Siglec-7. J. Cell Sci. 2020, 133, jcs244400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, H. Role of Decidual Natural Killer Cells in Human Pregnancy and Related Pregnancy Complications. Front. Immunol. 2021, 12, 728291. [Google Scholar] [CrossRef]
- Theresine, M.; Patil, N.D.; Zimmer, J. Airway Natural Killer Cells and Bacteria in Health and Disease. Front. Immunol. 2020, 11, 585048. [Google Scholar] [CrossRef]
- Fauriat, C.; Long, E.O.; Ljunggren, H.G.; Bryceson, Y.T. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood 2010, 115, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghari, M.A.; Arefnezhad, R.; Parhizkar, F.; Hejazi, M.S.; Motavalli Khiavi, F.; Mahmoodpoor, A.; Yousefi, M. T lymphocytes and preeclampsia: The potential role of T-cell subsets and related MicroRNAs in the pathogenesis of preeclampsia. Am. J. Reprod. Immunol. 2021, 86, e13475. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preeclampsia (n = 15) | Control (n = 15) | p Value | |
|---|---|---|---|
| Maternal age (years), mean ± SD | 28.8 ± 3.8 | 30.7 ± 5.1 | 0.3130 |
| Gestational age (weeks), mean ± SD | 34.8 ± 1.7 | 34.5 ± 2.1 | 0.4910 |
| Systolic RR (mmHg), mean ± SD | 167.6 ± 14.7 | 117 ± 11.3 | 0.0003 |
| Diastolic RR (mmHg), mean ± SD | 112 ± 11.4 | 79.5 ± 0.7 | 0.0014 |
| Birth weight (grams), mean ± SD | 2015 ± 648.3 | 2750 ± 282.8 | 0.6610 |
| Cesarean deliveries (%) | 14 (93) | 5 (33.3) | 0.0041 |
| Body mass index (BMI), mean ± SD | 22.9 ± 1.9 | 26.9 ± 4.1 | 0.0519 |
| Intrauterine growth restriction (IUGR) (%) | 11 (73) | 1 (6.6) | 0.0191 |
| Postpartum complications (%) | 6 (40) | 0 (0) | 0.1136 |
| Age of the women | 28.50 ± 5.50 | 29.47 ± 4.26 | 0.730 |
| Parity (M ± IqR) | 0.00 ±1 | 0.00 ±1 | 0.928 |
| AST | 63.00±8.70 | 13.58 ± 5.99 | 0.026 |
| ALT | 42.17 ± 6.69 | 16.68 ± 9.79 | 0.034 |
| PLT | 229-92 ± 121.00 | 278.95 ± 68.29 | 0.052 |
| Leukocyte | 10.95 ± 5.02 | 10.46 ± 2.25 | 0.921 |
| Hgb | 120.92 ± 14.16 | 115.63 ± 13.31 | 0.464 |
| Hct | 0.35 ± 0.04 | 0.31 ± 0.09 | 0.225 |
| Urea | 3.62 ± 0.74 | 4.28 ± 1.96 | 0.404 |
| Kreatinin | 74.83 ± 8.56 | 78.95 ± 13.89 | 0.402 |
| Uric acid | 336.50 ± 63.25 | 262.32 ± 70.57 | 0.004 |
| LDH | 348.08 ± 167.65 | 267.16 ± 56.41 | 0.028 |
| Antibody | Dilution | Host | Cellular Localization | Developer |
|---|---|---|---|---|
| CD8 | 1:100 | Mouse | Membrane | Dako M7103 (Dako, Glostrup, Denmark) |
| CD56 | 1:100 | Mouse | Membrane | Leica NCL-L-CD56-1B6 (Leica Biosystems Wetzlar, Germany) |
| GZMK | 1:300 | Rabbit | Cytoplasm | Sigma Aldrich HPA063181 (St. Louis, MI, USA) |
| FasL | 1:300 | Rabbit | Cytoplasm | Sigma Aldrich SAB4501532 (St. Louis, MI, USA) |
| IFN-γ | 1:100 | Rabbit | Cytoplasm | Cell signalling 8455 (Cell Signaling, Boston, MA, USA) |
| Transcript | Forward Primer | Reverse Primer | Amplicon Size | Tm F/R | TA | CG% F/R |
|---|---|---|---|---|---|---|
| GZMK | TTAAGA CCTTCT GACACC | TGGAA GACA CCTTT ACAG | 191 | 51.1/51.2 | 51.1 | 44.44/44.44 |
| IFN-γ | AGCTCT GCATCG TTTTGG GTT | GTTCC ATTAT CCGCT ACAT CTGAA | 118 | 61.5/59.0 | 59.0 | 47.62/41.67 |
| FasL | TGGCCT TGTGAT CAAT GAAA | TCATC ATCTT CCCCT CCATC | 155 | 54.5/65.3 | 54.5 | 40.0/50.0 |
| GAPDH | ACCCAC TCCTCC ACCTTT GAC | TCCAC CACCC TGTTG CTGTA G | 110 | 64.5/63.9 | 63.9 | 57.14/57.14 |
| GZMK | FasL | IFN-γ | ||
|---|---|---|---|---|
| CD8 + T cells | +++/f | −/n | −/n | |
| NK + cells | +++/f | −/n | −/n | |
| Control | Villous trophoblast | +/d | +++/d | +/f |
| Extravillous trophoblast | +/d | ++/d | +/f | |
| CD8 + Tcells | +++/f | −/n | −/n | |
| Preeclampsia | NK + cells | +++/f | −/n | −/n |
| Villous trophoblast | ++/d | ++/d | +/f | |
| Extravillous trophoblast | +++/d | +/d | +/f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vukoja, M.; Ćurlin, M.; Vukojević, K.; Jelić-Knezović, N.; Kolobarić, A.; Orlović Vlaho, M.; Šoljić, V. Effect of Granzyme K, FasL and Interferon-γ Expression in Placentas with Preeclampsia. Biomedicines 2024, 12, 842. https://doi.org/10.3390/biomedicines12040842
Vukoja M, Ćurlin M, Vukojević K, Jelić-Knezović N, Kolobarić A, Orlović Vlaho M, Šoljić V. Effect of Granzyme K, FasL and Interferon-γ Expression in Placentas with Preeclampsia. Biomedicines. 2024; 12(4):842. https://doi.org/10.3390/biomedicines12040842
Chicago/Turabian StyleVukoja, Martina, Marina Ćurlin, Katarina Vukojević, Nevenka Jelić-Knezović, Anita Kolobarić, Martina Orlović Vlaho, and Violeta Šoljić. 2024. "Effect of Granzyme K, FasL and Interferon-γ Expression in Placentas with Preeclampsia" Biomedicines 12, no. 4: 842. https://doi.org/10.3390/biomedicines12040842
APA StyleVukoja, M., Ćurlin, M., Vukojević, K., Jelić-Knezović, N., Kolobarić, A., Orlović Vlaho, M., & Šoljić, V. (2024). Effect of Granzyme K, FasL and Interferon-γ Expression in Placentas with Preeclampsia. Biomedicines, 12(4), 842. https://doi.org/10.3390/biomedicines12040842