

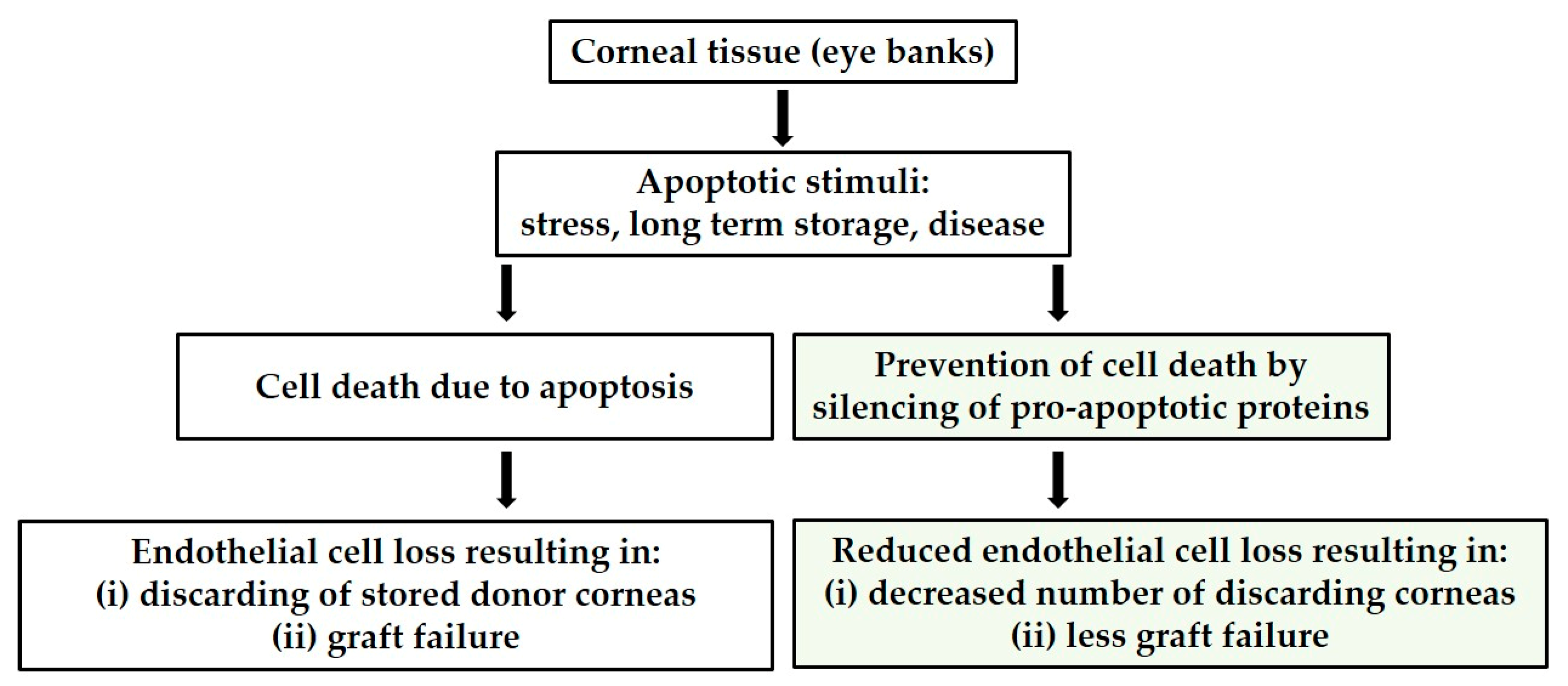
Suppressing Pro-Apoptotic Proteins by siRNA in Corneal Endothelial Cells Protects against Cell Death

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Models
2.1.1. In Vitro—Corneal Endothelial Cell Line
2.1.2. Ex Vivo—Donor Cornea
2.2. Inhibition of Pro-Apoptotic Proteins by Small Interfering RNA (siRNA)
2.3. Cell Viability of HCECs
2.4. Apoptosis Induction in HCECs
2.5. Morphology of HCECs
2.6. Apoptosis Assays
2.6.1. Caspase-3 Activity
2.6.2. TUNEL Assay
2.6.3. Annexin V-FITC/PI
2.7. Statistical Analysis
3. Results
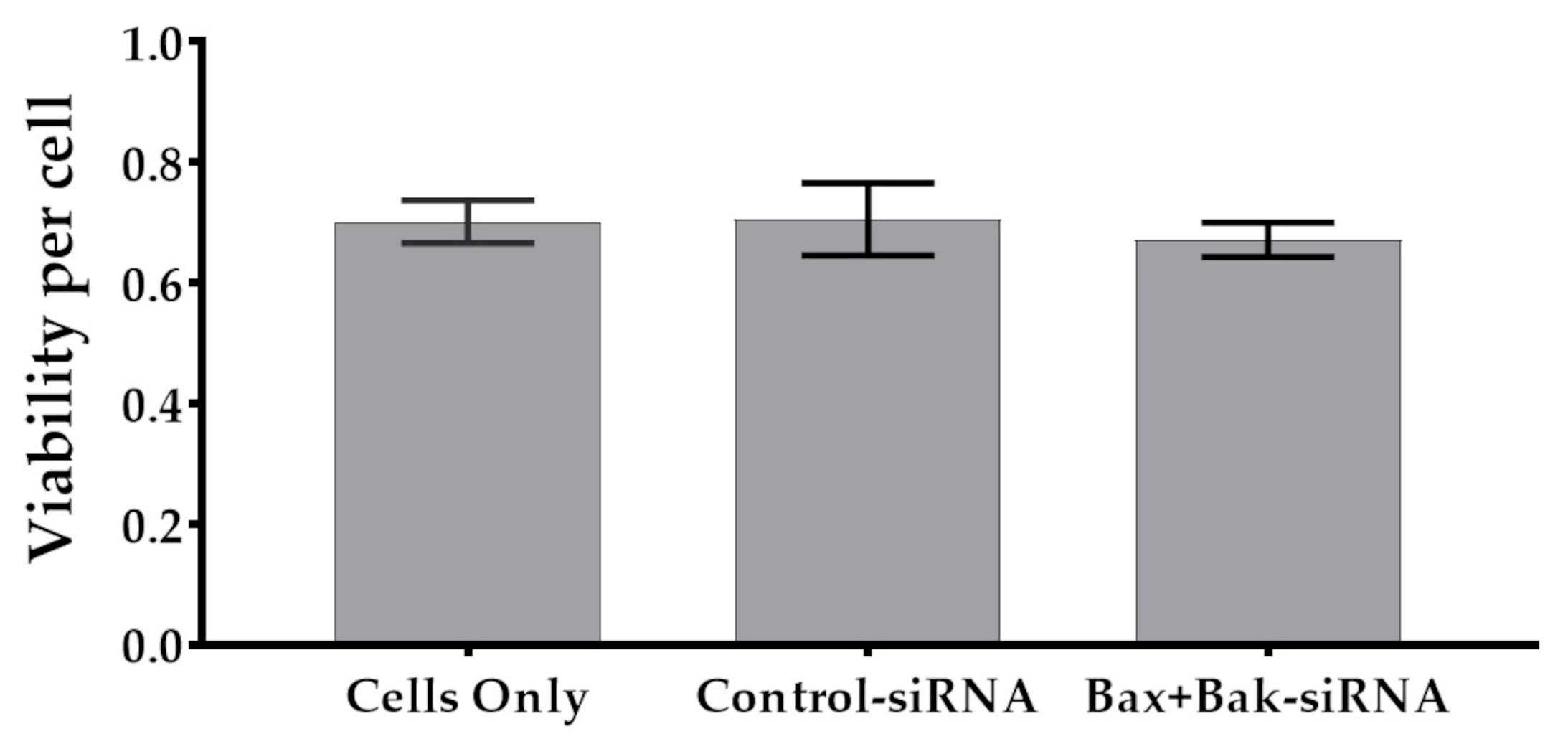
3.1. Unchanged Cell Viability after Accell siRNA Transfection
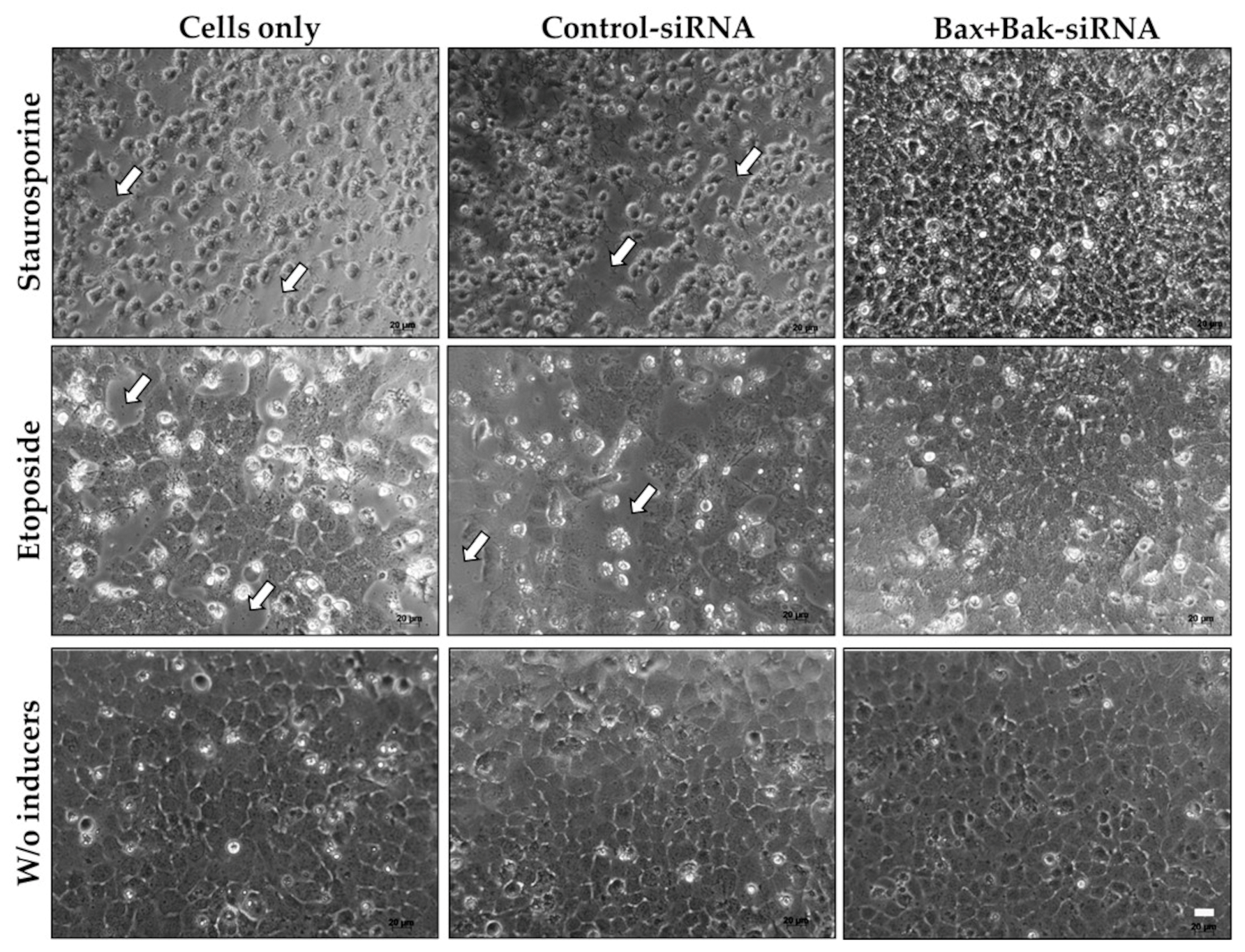
3.2. Evaluation of HCEC Morphology after Apoptosis Induction
3.3. Bax+Bak-siRNA Transfection Reduced Caspase-3 Activity in HCECs
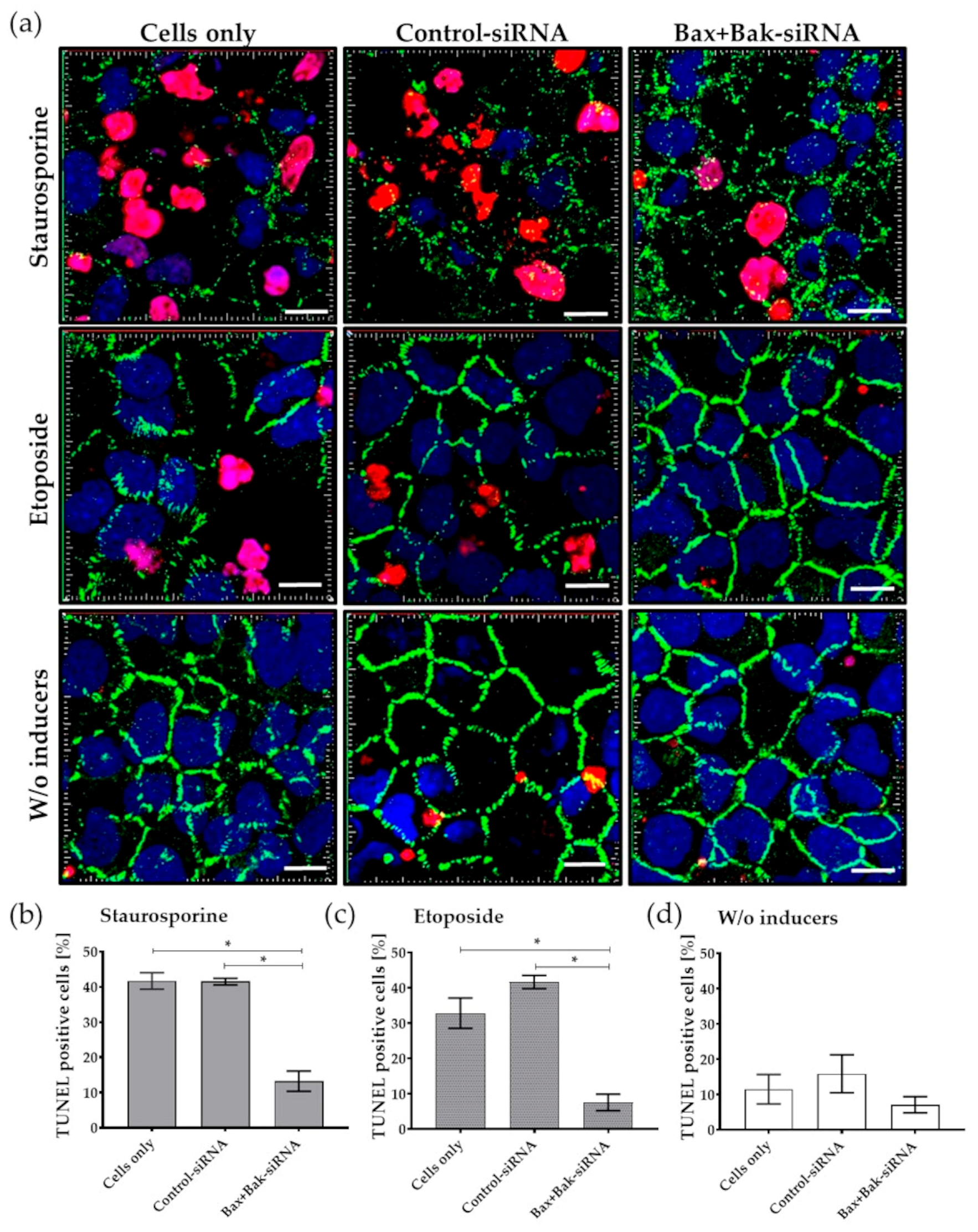
3.4. Inhibition of Pro-Apoptotic Proteins Results in Reduced Apoptosis Rates
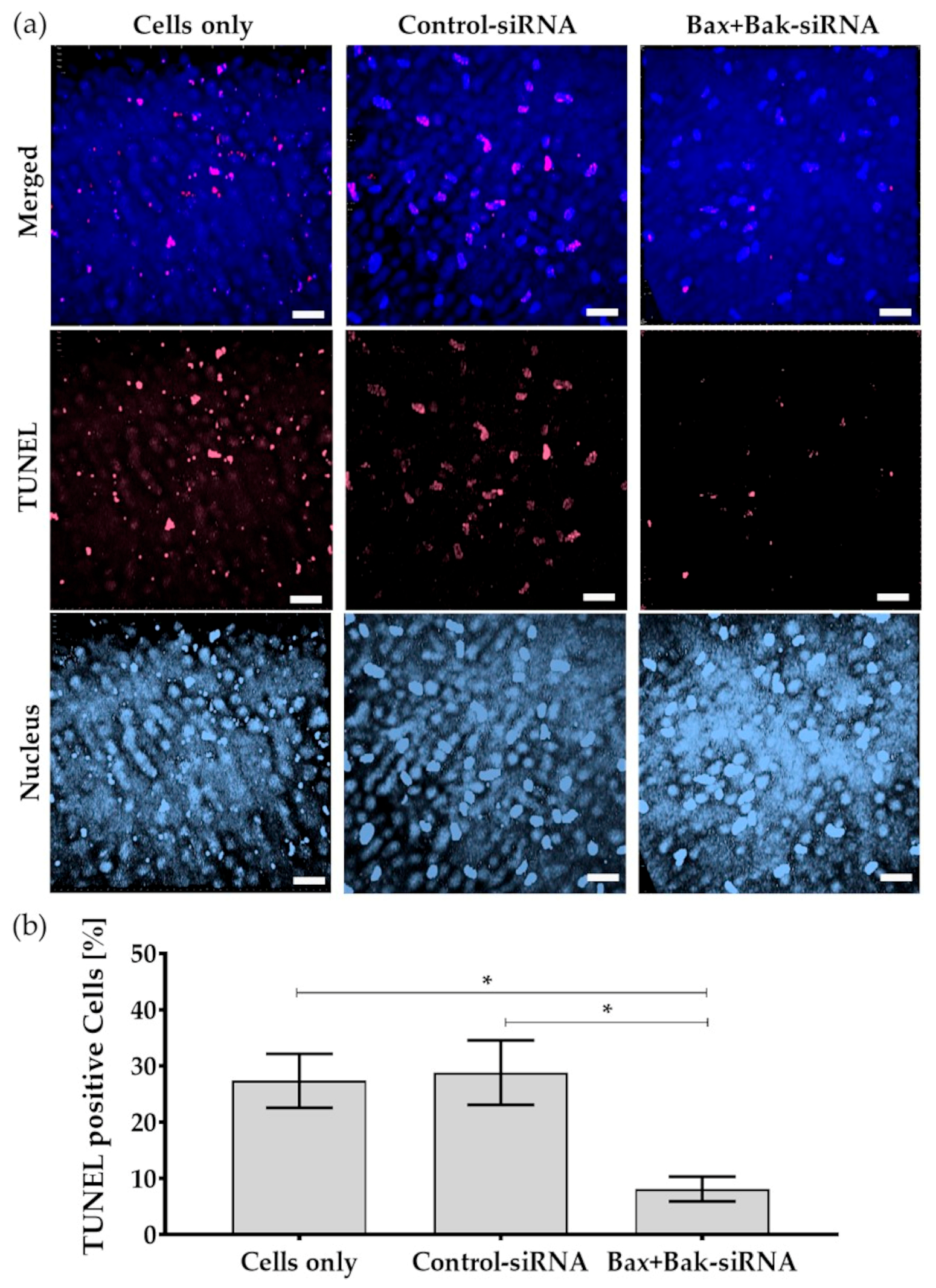
3.5. Bax+BaksiRNA Transfection Results in Reduced Development of Apoptosis
3.6. Reduced Apoptosis Rate in Human Donor Corneal Endothelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kreutziger, G.O. Lateral membrane morphology and gap junction structure in rabbit corneal endothelium. Exp. Eye Res. 1976, 23, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Waring, G.O., 3rd; Bourne, W.M.; Edelhauser, H.F.; Kenyon, K.R. The corneal endothelium. Normal and pathologic structure and function. Ophthalmology 1982, 89, 531–590. [Google Scholar] [CrossRef] [PubMed]
- Capella, J.A. Regeneration of endothelium in diseased and injured cornea. Am. J. Ophthalmol. 1972, 74, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Joyce, N.C. Proliferative response of corneal endothelial cells from young and older donors. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, F.; Ortiz-Morales, G.; O’Connor-Cordova, M.A.; Sancén-Herrera, J.P.; Zavala, J.; Valdez-Garcia, J.E. Fuchs endothelial corneal dystrophy: An updated review. Int. Ophthalmol. 2024, 44, 61. [Google Scholar] [CrossRef] [PubMed]
- Price, M.O.; Mehta, J.S.; Jurkunas, U.V.; Price, F.W., Jr. Corneal endothelial dysfunction: Evolving understanding and treatment options. Prog. Retin. Eye Res. 2021, 82, 100904. [Google Scholar] [CrossRef]
- Ong Tone, S.; Kocaba, V.; Böhm, M.; Wylegala, A.; White, T.L.; Jurkunas, U.V. Fuchs endothelial corneal dystrophy: The vicious cycle of Fuchs pathogenesis. Prog. Retin. Eye Res. 2021, 80, 100863. [Google Scholar] [CrossRef]
- Dunker, S.L.; Armitage, W.J.; Armitage, M.; Brocato, L.; Figueiredo, F.C.; Heemskerk, M.B.A.; Hjortdal, J.; Jones, G.L.A.; Konijn, C.; Nuijts, R.M.M.A.; et al. Practice patterns of corneal transplantation in Europe: First report by the European Cornea and Cell Transplantation Registry. J. Cataract. Refract. Surg. 2021, 47, 865–869. [Google Scholar] [CrossRef]
- Tidke, S.C.; Tidake, P.A. Review of Corneal Blindness: Causes and Management. Cureus 2022, 14, e30097. [Google Scholar] [CrossRef]
- Burton, M.J. Corneal Blindness: Prevention, treatment and rehabilitation. Community Eye Health 2009, 22, 33–35. [Google Scholar]
- Li, S.; Kim, E.; Bonanno, J.A. Fluid transport by the cornea endothelium is dependent on buffering lactic acid efflux. Am. J. Physiol. Cell Physiol. 2016, 311, C116–C126. [Google Scholar] [CrossRef]
- Ruusuvaara, P.; Setala, K. Long-term follow-up of cryopreserved corneal endothelium: A specular microscopic study. Acta Ophthalmol. 1988, 66, 687–691. [Google Scholar] [CrossRef]
- Anitha, V.; Tandon, R.; Shah, S.G.; Radhakrishnan, N.; Singh, S.; Murugesan, V.; Patwardhan, V.; Ravindran, M. Corneal blindness and eye banking: Current strategies and best practices. Indian. J. Ophthalmol. 2023, 71, 3142–3148. [Google Scholar] [CrossRef] [PubMed]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Reinshagen, H.; Maier, P.C.; Böhringer, D. Aktivitäten der Sektion Gewebetransplantation und Biotechnologie der Deutschen Ophthalmologischen Gesellschaft: Leistungsbericht 2018 [The 2018 Activity Report of the Tissue Transplantation and Biotechnology Section of the German Ophthalmological Society]. Klin. Monbl. Augenheilkd. 2021, 238, 186–190. [Google Scholar] [CrossRef]
- Reinshagen, H.; Böhringer, D.; Schroeter, J.; Maier, P.C. Aktivitäten der Sektion Gewebetransplantation und Biotechnologie der Deutschen Ophthalmologischen Gesellschaft: Leistungsbericht 2021 [The 2021 Activity Report of the Tissue Transplantation and Biotechnology Section of the German Ophthalmological Society]. Klin. Monbl. Augenheilkd. 2023; ahead of print. [Google Scholar] [CrossRef]
- Fuchsluger, T.A. Protektion von Hornhautendothel vor Zelluntergang durch Gen- und Zelltherapie [Protection of corneal endothelium from apoptosis by gene and cell therapy]. Ophthalmologe 2016, 113, 524–528. [Google Scholar] [CrossRef]
- Bonato, P.; Bagno, A. Replace or Regenerate? Diverse Approaches to Biomaterials for Treating Corneal Lesions. Biomimetics 2024, 9, 202. [Google Scholar] [CrossRef]
- Williams, K.A.; Lowe, M.; Bartlett, C.; Kelly, T.L.; Coster, D.J. Risk factors for human corneal graft failure within the Australian corneal graft registry. Transplantation 2008, 86, 1720–1724. [Google Scholar] [CrossRef]
- McCarey, B.E.; Kaufman, H.E. Improved corneal storage. Investig. Ophthalmol. 1974, 13, 165–173. [Google Scholar]
- Naguib, Y.W.; Saha, S.; Skeie, J.M.; Acri, T.; Ebeid, K.; Abdel-Rahman, S.; Kesh, S.; Schmidt, G.A.; Nishimura, D.Y.; Banas, J.A.; et al. Solubilized ubiquinol for preserving corneal function. Biomaterials 2021, 275, 120842. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.A.; Brereton, H.M.; Coster, D.J. Prospects for genetic modulation of corneal graft survival. Eye 2009, 23, 1904–1909. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Chaurasia, S.S.; Medeiros, F.W. Apoptosis in the initiation, modulation and termination of the corneal wound healing response. Exp. Eye Res. 2007, 85, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Fuchsluger, T.A.; Jurkunas, U.; Kazlauskas, A.; Dana, R. Anti-apoptotic gene therapy prolongs survival of corneal endothelial cells during storage. Gene Ther. 2011, 18, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Panikker, P.; D’Souza, S.; Shetty, R.; Mohan, R.R.; Ghosh, A. Corneal regeneration using gene therapy approaches. Cells 2023, 12, 1280. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sinha, N.R.; Mohan, R.R. Corneal gene therapy: Structural and mechanistic understanding. Ocul. Surf. 2023, 29, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Amador, C.; Shah, R.; Ghiam, S.; Kramerov, A.A.; Ljubimov, A.V. Gene Therapy in the Anterior Eye Segment. Curr. Gene Ther. 2022, 22, 104–131. [Google Scholar] [CrossRef]
- Mohan, R.R.; Martin, L.M.; Sinha, N.R. Novel insights into gene therapy in the cornea. Exp. Eye Res. 2021, 202, 108361. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Thieme, D.; Czugala, M.; Kruse, F.E.; Fuchsluger, T.A. Lamin Cleavage: A reliable marker for studying staurosporine-induced apoptosis in corneal tissue. Investig. Ophth Vis. Sci. 2017, 58, 5794–5801. [Google Scholar] [CrossRef]
- Morris, D.L.; Johnson, S.; Bleck, C.K.E.; Lee, D.; Tjandra, N. Humanin selectively prevents the activation of pro-apoptotic protein BID by sequestering it into fibers. J. Biol. Chem. 2020, 295, 18226–18238. [Google Scholar] [CrossRef]
- O’Brien, M.A.; Kirby, R. Apoptosis: A review of pro-apoptotic and anti-apoptotic pathways and dysregulation in disease. J. Vet. Emerg. Crit. Care 2008, 18, 572–585. [Google Scholar] [CrossRef]
- Barcia, R.N.; Dana, M.R.; Kazlauskas, A. Corneal graft rejection is accompanied by apoptosis of the endothelium and is prevented by gene therapy with bcl-xL. Am. J. Transplant. 2007, 7, 2082–2089. [Google Scholar] [CrossRef]
- Fuchsluger, T.A.; Jurkunas, U.; Kazlauskas, A.; Dana, R. Corneal endothelial cells are protected from apoptosis by gene therapy. Hum. Gene Ther. 2011, 22, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Orzáez, M.; Sancho, M.; Marchán, S.; Mondragón, L.; Montava, R.; Valero, J.G.; Landeta, O.; Basañez, G.; Carbajo, R.J.; Pineda-Lucena, A.; et al. Apaf-1 inhibitors protect from unwanted cell death in in vivo models of kidney ischemia and chemotherapy induced ototoxicity. PLoS ONE 2014, 9, e110979. [Google Scholar] [CrossRef]
- Gemayel, M.C.; Bhatwadekar, A.D.; Ciulla, T. RNA therapeutics for retinal diseases. Expert. Opin. Biol. Ther. 2021, 21, 603–613. [Google Scholar] [CrossRef]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the delivery of RNA therapeutics: From concept to clinical reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.S.; Ding, H.; Kennington, L.; Moore, J.T.; Schelter, J.; Burchard, J.; Linsley, P.S.; Aronin, N.; Xu, Z.; Zamore, P.D. Designing siRNA that distinguish between genes that differ by a single nucleotide. PLoS Genet. 2006, 2, e140. [Google Scholar] [CrossRef]
- Schiroli, D.; Gómara, M.J.; Maurizi, E.; Atkinson, S.D.; Mairs, L.; Christie, K.A.; Cobice, D.F.; McCrudden, C.M.; Nesbit, M.A.; Haro, I.; et al. Effective in vivo topical delivery of sirna and gene silencing in intact corneal epithelium using a modified cell-penetrating peptide. Mol. Ther. Nucleic Acids 2019, 17, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://horizondiscovery.com/en/gene-modulation/knockdown/sirna/products/accell-sirna-reagents (accessed on 16 May 2024).
- Bednarz, J.; Teifel, M.; Friedl, P.; Engelmann, K. Immortalization of human corneal endothelial cells using electroporation protocol optimized for human corneal endothelial and human retinal pigment epithelial cells. Acta Ophthalmol. Scand. 2000, 78, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Valtink, M.; Gruschwitz, R.; Funk, R.H.; Engelmann, K. Two clonal cell lines of immortalized human corneal endothelial cells show either differentiated or precursor cell characteristics. Cells Tissues Organs 2008, 187, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Leake, D.; Boese, Q.; Scaringe, S.; Marshall, W.S.; Khvorova, A. Rational siRNA design for RNA interference. Nat. Biotechnol. 2004, 22, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Van Kooten, T.G.; Klein, C.L.; Wagner, M.; Kirkpatrick, C.J. Focal adhesions and assessment of cytotoxicity. J. Biomed. Mater. Res. 1999, 46, 33–43. [Google Scholar] [CrossRef]
- Staehlke, S.; Barth, T.; Muench, M.; Schroeter, J.; Wendlandt, R.; Oldorf, P.; Peters, R.; Nebe, B.; Schulz, A.-P. The Impact of Ultrashort Pulse Laser Structuring of Metals on In-Vitro Cell Adhesion of Keratinocytes. J. Funct. Biomater. 2024, 15, 34. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wong, Y.L.; Walkden, A. Current Perspectives on Corneal Transplantation. Clin. Ophthalmol. 2022, 16, 631–646. [Google Scholar] [CrossRef]
- Albon, J.; Tullo, A.B.; Aktar, S.; Boulton, M.E. Apoptosis in the endothelium of human corneas for transplantation. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2887–2893. [Google Scholar]
- Thanuja, M.Y.; Tellakula, S.S.; Suryavanshi, S.V.; Keerthana, G.S.; Ranganath, S.H. Fusogenic liposome-coated nanoparticles for rapid internalization into donor corneal endothelial tissue to enable prophylaxis before transplantation. Nanoscale Adv. 2023, 5, 6410–6422. [Google Scholar] [CrossRef]
- Chatauret, N.; Thuillier, R.; Hauet, T. Preservation strategies to reduce ischemic injury in kidney transplantation: Pharmacological and genetic approaches. Curr. Opin. Organ. Transplant. 2011, 16, 180–187. [Google Scholar] [CrossRef]
- Pallet, N.; Dieudé, M.; Cailhier, J.; Hébert, M. The molecular legacy of apoptosis in transplantation. Am. J. Transplant. 2012, 12, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.A.; Klebe, S. Gene therapy for corneal dystrophies and disease, where are we? Curr. Opin. Ophthalmol. 2012, 23, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Staurenghi, F.; McClements, M.E.; Salman, A.; MacLaren, R.E. Minicircle delivery to the neural retina as a gene therapy approach. Int. J. Mol. Sci. 2022, 23, 11673. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.L.; Karatza, E.; Mody, H.; Nagaraja Shastri, P.; Khajeh Pour, S.; Yang, T.Y.; Swanson, M.; Chao, D.; Devineni, D. Clinical pharmacology perspective on development of adeno-associated virus vector-based retina gene therapy. Clin. Pharmacol. Ther. 2024, 115, 1212–1232. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, M.; Chen, F.; Goldberg, J.L.; Sabel, B.A. Nanomedicine and drug delivery to the retina: Current status and implications for gene therapy. Naunyn Schmiedebergs Arch. Pharmacol. 2022, 395, 1477–1507. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.T.; Tighe, S.; Chen, S.L.; Zhang, Y.; Chen, S.Y.; Kao, W.W.Y.; Tseng, S.C.G. Manufacturing of human corneal endothelial grafts. Ocul. Surf. 2023, 29, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Vitte, P.A.; Bombrun, A.; Rostovtseva, T.K.; Montessuit, S.; Hiver, A.; Schwarz, M.K.; Church, D.J.; Korsmeyer, S.J.; Martinou, J.C.; et al. Bax channel inhibitors prevent mitochondrion- mediated apoptosis and protect neurons in a model of global brain ischemia. J. Biol. Chem. 2005, 280, 42960–42970. [Google Scholar] [CrossRef] [PubMed]
- Hochhauser, E.; Kivity, S.; Offen, D.; Maulik, N.; Otani, H.; Barhum, Y.; Pannet, H.; Shneyvays, V.; Shainberg, A.; Goldshtaub, V.; et al. Bax ablation protects against myocardial ischemia-reperfusion injury in transgenic mice. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H2351–H2359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, Y.; Zhu, Q.; Gu, X.F.; Zhu, Y.Z. The discovery of a novel inhibitor of apoptotic protease activating factor-1 (Apaf-1) for ischemic heart: Synthesis, activity and target identification. Sci. Rep. 2016, 6, 29820. [Google Scholar] [CrossRef]
- Chancellor, D.; Barrett, D.; Nguyen-Jatkoe, L.; Millington, S.; Eckhardt, F. The state of cell and gene therapy in 2023. Mol. Ther. 2023, 31, 3376–3388. [Google Scholar] [CrossRef]
- Wilson, C.; Foster, G.H.; Bitzan, M.M. Silencing of bak ameliorates apoptosis of human proximal tubular epithelial cells by Escherichia coli-derived Shiga toxin 2. Infection 2005, 33, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.L.; Niu, Q.; Niu, P.Y.; Shi, Y.T.; Liu, C.Y.; Di Gioacchino, M.; Zhang, L.; Zhang, C.; Braga, M. Bax Gene silencing: A potential intervention in aluminum-induced neural cell death. J. Biol. Regul. Homeost. Agents 2010, 24, 7–17. [Google Scholar] [PubMed]
- Wei, M.C.; Zong, W.X.; Cheng, E.H.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacGregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.J. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.X.M.; Caballero, R.; Ali, H.; Roy, D.L.F.; Cassol, E.; Kumar, A. Transfection of hard-to-transfect primary human macrophages with Bax siRNA to reverse Resveratrol-induced apoptosis. RNA Biol. 2020, 17, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Czugala, M.; Mykhaylyk, O.; Böhler, P.; Onderka, J.; Stork, B.; Wesselborg, S.; Kruse, F.E.; Plank, C.; Singer, B.B.; Fuchsluger, T.A. Efficient and safe gene delivery to human corneal endothelium using magnetic nanoparticles. Nanomedicine 2016, 11, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Baran-Rachwalska, P.; Torabi-Pour, N.; Sutera, F.M.; Ahmed, M.; Thomas, K.; Nesbit, M.A.; Welsh, M.; Moore, C.B.T.; Saffie-Siebert, S.R. Topical siRNA delivery to the cornea and anterior eye by hybrid silicon-lipid nanoparticles. J. Control. Release 2020, 326, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Robbins, P.D.; Ghivizzani, S.C. Viral vectors for gene therapy. Pharmacol. Ther. 1998, 80, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Endo, K.-i.; Tanioka, H.; Sasaoka, M.; Tashiro, K.; Kinoshita, S.; Kageyama, M. Novel use of a chemically modified siRNA for robust and sustainable in vivo gene silencing in the retina. Sci. Rep. 2020, 10, 22343. [Google Scholar] [CrossRef]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef]
- Inoue, S.; Browne, G.; Melino, G.; Cohen, G.M. Ordering of caspases in cells undergoing apoptosis by the intrinsic pathway. Cell Death Differ. 2009, 16, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Thuret, G.; Chiquet, C.; Herrag, S.; Dumollard, J.M.; Boudard, D.; Bednarz, J.; Campos, L.; Gain, P. Mechanisms of staurosporine induced apoptosis in a human corneal endothelial cell line. Br. J. Ophthalmol. 2003, 87, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Härtel, S.; Zorn-Kruppa, M.; Tykhonova, S.; Alajuuma, P.; Engelke, M.; Diehl, H.A. Staurosporine-induced apoptosis in human cornea epithelial cells in vitro. Cytometry A 2003, 55, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.F.; Muqaddas, M.; Sarwar, S. Biochemicalmechanisms of etoposide; upshot of cell death. Int. J. Pharm. Sci. Res. 2015, 6, 4920. [Google Scholar] [CrossRef]
- Petros, A.M.; Olejniczak, E.T.; Fesik, S.W. Structural biology of the Bcl-2 family of proteins. Biochim. Biophys. Acta 2004, 1644, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Chaudhry, G.E. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Díaz, C.; Ikeda, F. Roles of ubiquitin in autophagy and cell death. Semin. Cell Dev. Biol. 2019, 93, 125–135. [Google Scholar] [CrossRef]
- Logue, S.E.; Martin, S.J. Caspase activation cascades in apoptosis. Biochem. Soc. Trans. 2008, 36 Pt 1, 1–9. [Google Scholar] [CrossRef]
- Macoska, J.A.; Adsule, S.; Tantivejkul, K.; Wang, S.; Pienta, K.J.; Lee, C.T. −(−)Gossypol promotes the apoptosis of bladder cancer cells in vitro. Pharmacol. Res. 2008, 58, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Rudel, T. Caspase inhibitors in prevention of apoptosis. Herz 1999, 24, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Kressel, M.; Groscurth, P. Distinction of apoptotic and necrotic cell death by in situ labelling of fragmented DNA. Cell Tissue Res. 1994, 278, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Hoentsch, M.; von Woedtke, T.; Weltmann, K.-D.; Nebe, J.B. Time-dependent effects of low-temperature atmospheric-pressure argon plasma on epithelial cell attachment, viability and tight junction formation in vitro. J. Phys. D Appl. Phys. 2012, 45, 025206. [Google Scholar] [CrossRef]
- Walker, J.A.; Quirke, P. Viewing apoptosis through a ‘TUNEL’. J. Pathol. 2001, 195, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Osthoff, K.; Walczak, H.; Dröge, W.; Krammer, P.H. Cell nucleus and DNA fragmentation are not required for apoptosis. J. Cell Biol. 1994, 127, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Groscurth, P. Morphological features of cell death. News Physiol. Sci. 2004, 19, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Bossy-Wetzel, E.; Green, D.R. Detection of apoptosis by Annexin V labeling. Methods Enzymol. 2000, 322, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. J. Vis. Exp. 2011, 50, 2597. [Google Scholar] [CrossRef]
- Thieme, D.; Reuland, L.; Lindl, T.; Kruse, F.; Fuchsluger, T. Optimized human platelet lysate as novel basis for a serum-, xeno-, and additive-free corneal endothelial cell and tissue culture. J. Tissue Eng. Regen. Med. 2018, 12, 557–564. [Google Scholar] [CrossRef]
- Wang, M.X.; Zhao, J.; Zhang, H.; Li, K.; Niu, L.Z.; Wang, Y.P.; Zheng, Y.J. Potential protective and therapeutic roles of the nrf2 pathway in ocular diseases: An update. Oxid. Med. Cell. Longev. 2020, 2020, 9410952. [Google Scholar] [CrossRef] [PubMed]
- Batliwala, S.; Xavier, C.; Liu, Y.; Wu, H.; Pang, I.H. Involvement of Nrf2 in ocular diseases. Oxid. Med. Cell. Longev. 2017, 2017, 1703810. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Donor |
|---|---|
| Average age, y * | 74 ± 7 |
| Sex, female/male | 2/2 |
| Average cell density [cells/mm2] * | 1785 ± 235 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staehlke, S.; Mahajan, S.; Thieme, D.; Trosan, P.; Fuchsluger, T.A. Suppressing Pro-Apoptotic Proteins by siRNA in Corneal Endothelial Cells Protects against Cell Death. Biomedicines 2024, 12, 1439. https://doi.org/10.3390/biomedicines12071439
Staehlke S, Mahajan S, Thieme D, Trosan P, Fuchsluger TA. Suppressing Pro-Apoptotic Proteins by siRNA in Corneal Endothelial Cells Protects against Cell Death. Biomedicines. 2024; 12(7):1439. https://doi.org/10.3390/biomedicines12071439
Chicago/Turabian StyleStaehlke, Susanne, Siddharth Mahajan, Daniel Thieme, Peter Trosan, and Thomas A. Fuchsluger. 2024. "Suppressing Pro-Apoptotic Proteins by siRNA in Corneal Endothelial Cells Protects against Cell Death" Biomedicines 12, no. 7: 1439. https://doi.org/10.3390/biomedicines12071439
APA StyleStaehlke, S., Mahajan, S., Thieme, D., Trosan, P., & Fuchsluger, T. A. (2024). Suppressing Pro-Apoptotic Proteins by siRNA in Corneal Endothelial Cells Protects against Cell Death. Biomedicines, 12(7), 1439. https://doi.org/10.3390/biomedicines12071439