Scutellaria baicalensis Flavones as Potent Drugs against Acute Respiratory Injury during SARS-CoV-2 Infection: Structural Biology Approaches




Abstract
1. Introduction
2. Materials and Methods
2.1. Ligand Preparation
2.2. Assessment of Compounds’ Drug- and Lead-Likeness Features
2.3. Compounds’ Predicted Molecular Features and Pharmacophore Fingerprint Search
2.4. Compounds’ Computational Pharmacodynamic Profiles
2.5. Computational ADME-Tox Profiles of Natural Compounds
3. Results and Discussion
3.1. Drug-Likeness, Lead-Likeness, and ADME-Tox Features
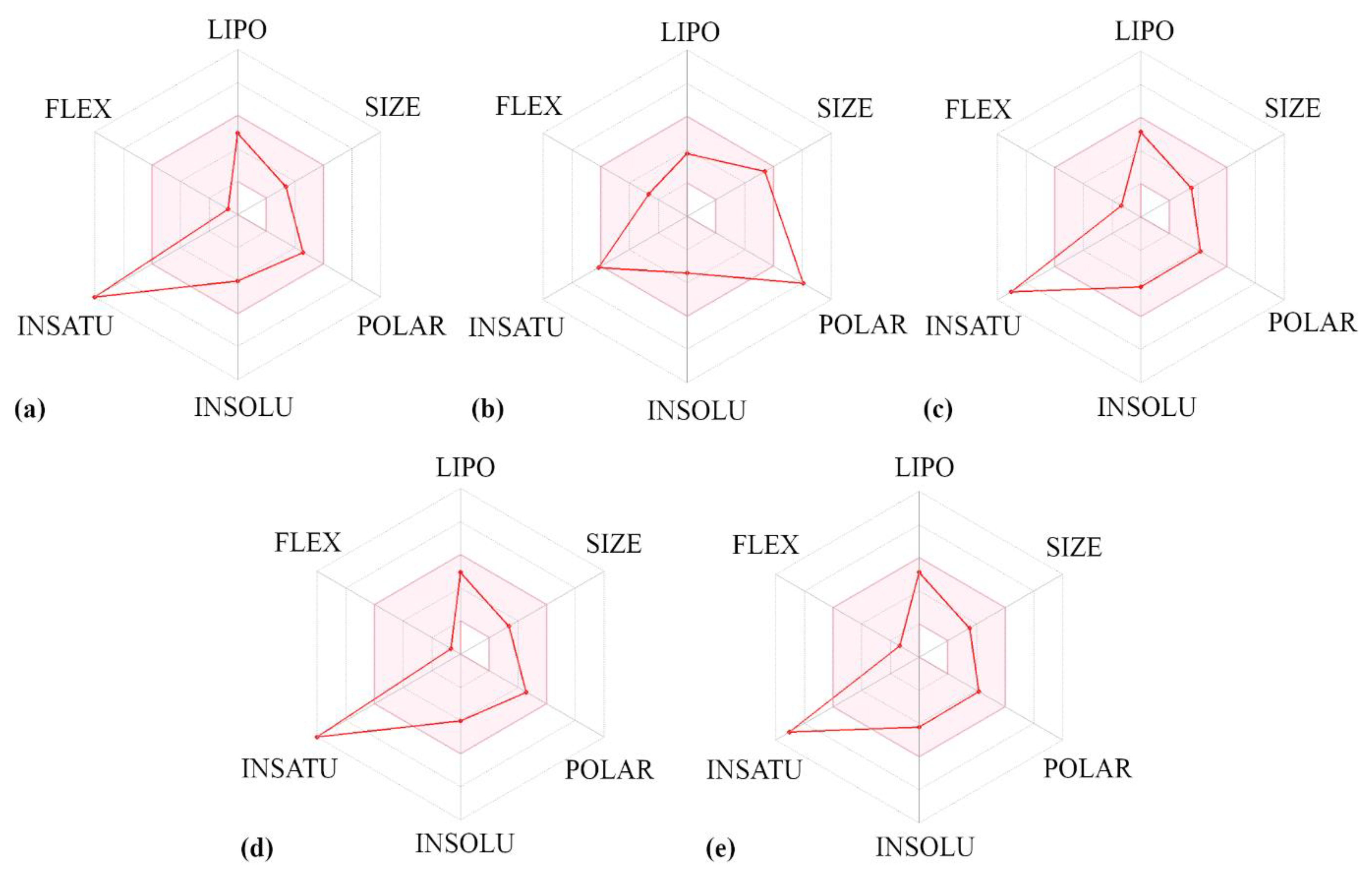
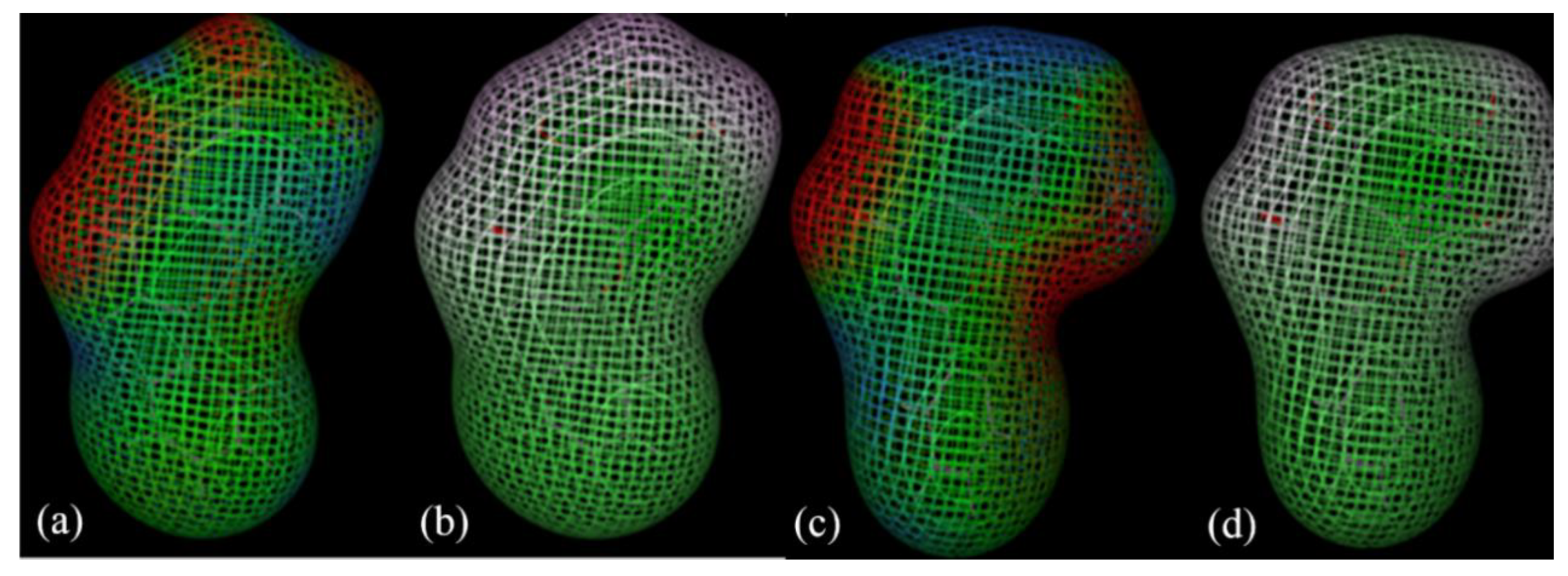
3.2. Predicted Molecular Features of Flavones and Their Pharmacophore Fingerprint Search
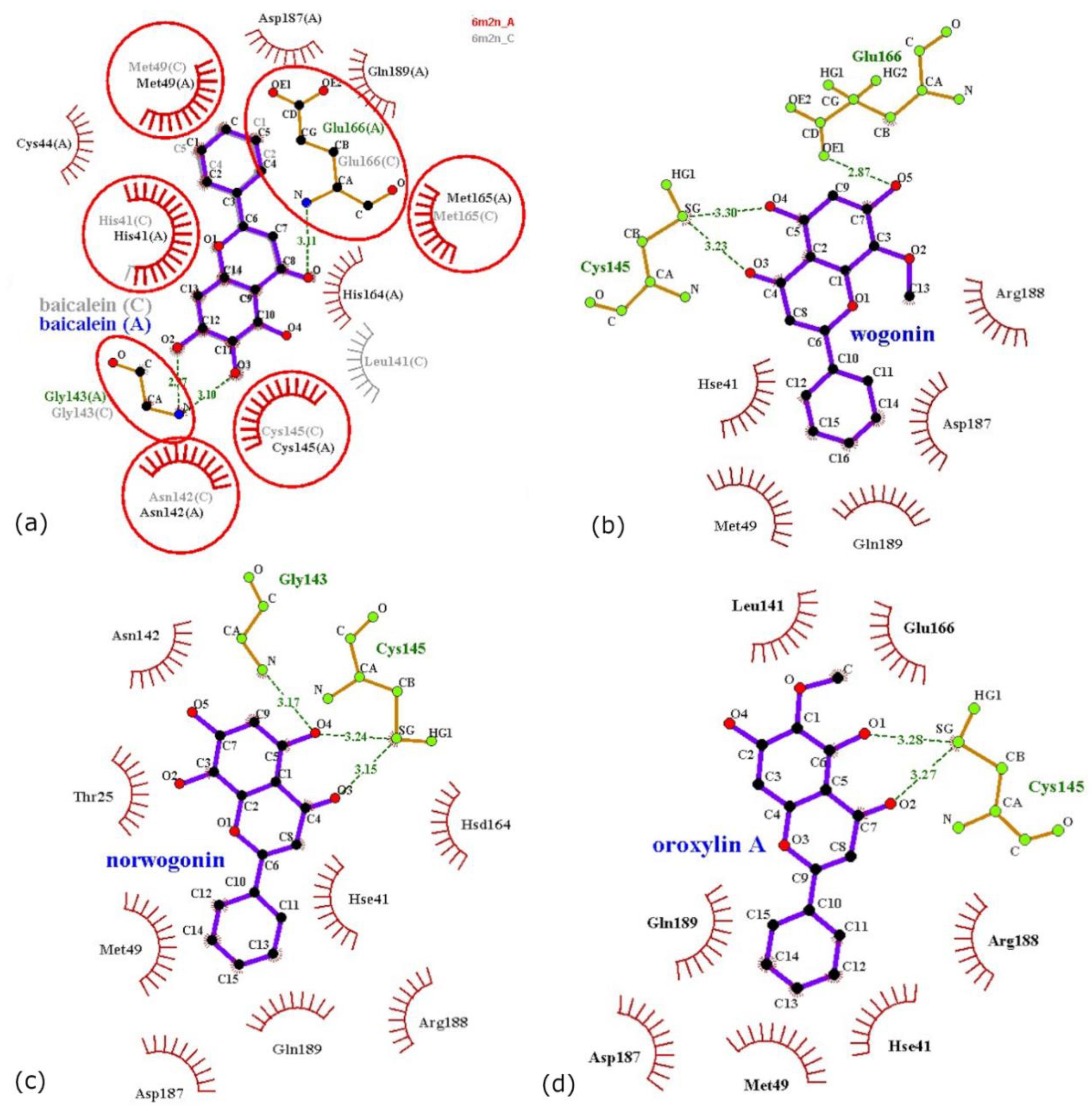
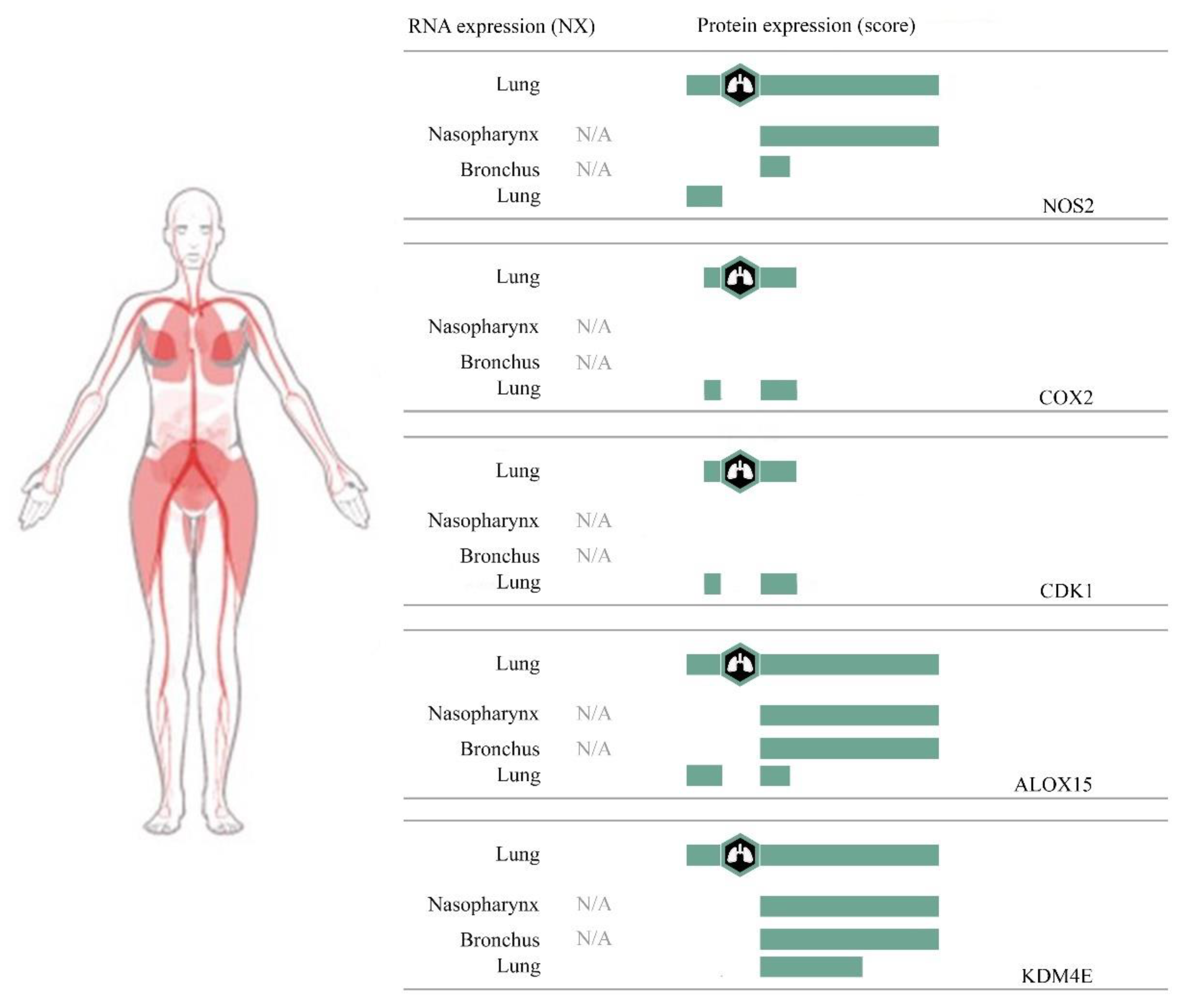
3.3. Computational Pharmacodynamic Profiles of Baicalein, Wogonin, Norwogonin, and Oroxylin A
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Marini, J.J.; Gattinoni, L. Management of COVID-19 Respiratory Distress. JAMA 2020, 323, 2329. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ji, P.; Pang, J.; Zhong, Z.; Li, H.; He, C.; Zhang, J.; Zhao, C. Clinical Characteristics of 3062 COVID-19 Patients: A Meta-analysis. J. Med. Virol. 2020, 92, 1902–1914. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-L.; Peng, M.-L.; Lee, S.-S.; Chen, C.-J.; Chen, W.-Y.; Yang, M.-L.; Kuan, Y.-H. Endotoxin-Induced Acute Lung Injury in Mice Is Protected by 5,7-Dihydroxy-8-Methoxyflavone via Inhibition of Oxidative Stress and HIF-1α: Endotoxin induced acute lung injury in mice. Environ. Toxicol. 2016, 31, 1700–1709. [Google Scholar] [CrossRef] [PubMed]
- Manjili, R.H.; Zarei, M.; Habibi, M.; Manjili, M.H. COVID-19 as an Acute Inflammatory Disease. J. Immunol. 2020, ji2000413. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological Findings of COVID-19 Associated with Acute Respiratory Distress Syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Pons, S.; Fodil, S.; Azoulay, E.; Zafrani, L. The Vascular Endothelium: The Cornerstone of Organ Dysfunction in Severe SARS-CoV-2 Infection. Crit. Care 2020, 24, 353. [Google Scholar] [CrossRef]
- McGonagle, D.; O’Donnell, J.S.; Sharif, K.; Emery, P.; Bridgewood, C. Immune Mechanisms of Pulmonary Intravascular Coagulopathy in COVID-19 Pneumonia. Lancet Rheumatol. 2020, 2, e437–e445. [Google Scholar] [CrossRef]
- Pan, X.; Li, J.; Tu, X.; Wu, C.; Liu, H.; Luo, Y.; Dong, X.; Li, X.; Pan, L.-L.; Sun, J. Lysine-Specific Demethylase-1 Regulates Fibroblast Activation in Pulmonary Fibrosis via TGF-Β1/Smad3 Pathway. Pharmacol. Res. 2020, 152, 104592. [Google Scholar] [CrossRef]
- Tavassoly, I.; Hu, Y.; Zhao, S.; Mariottini, C.; Boran, A.; Chen, Y.; Li, L.; Tolentino, R.E.; Jayaraman, G.; Goldfarb, J.; et al. Genomic Signatures Defining Responsiveness to Allopurinol and Combination Therapy for Lung Cancer Identified by Systems Therapeutics Analyses. Mol. Oncol. 2019, 13, 1725–1743. [Google Scholar] [CrossRef]
- Kayyali, U.S.; Donaldson, C.; Huang, H.; Abdelnour, R.; Hassoun, P.M. Phosphorylation of Xanthine Dehydrogenase/Oxidase in Hypoxia. J. Biol. Chem. 2001, 276, 14359–14365. [Google Scholar] [CrossRef]
- Abrial, C.; Grassin-Delyle, S.; Salvator, H.; Brollo, M.; Naline, E.; Devillier, P. 15-Lipoxygenases Regulate the Production of Chemokines in Human Lung Macrophages: 15-LOXs Affect Production of Chemokines in Lung Macrophages. Br. J. Pharmacol. 2015, 172, 4319–4330. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, G.; Kaliamurthi, S.; Kaushik, A.C.; Khan, A.; Wei, Y.-K.; Cho, W.C.; Gu, K.; Wei, D.-Q. Identification of Target Gene and Prognostic Evaluation for Lung Adenocarcinoma Using Gene Expression Meta-Analysis, Network Analysis and Neural Network Algorithms. J. Biomed. Inform. 2018, 86, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Liao, H.; Shi, F.; Chen, X.; Hu, H.; Tian, M.; Wang, L.; Ying, S. Inhibition of Cyclooxygenase‑2 Sensitizes Lung Cancer Cells to Radiation‑induced Apoptosis. Oncol. Lett. 2017, 14, 5959–5965. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Park, G.Y.; Christman, J.W. Involvement of Cyclooxygenase-2 and Prostaglandins in the Molecular Pathogenesis of Inflammatory Lung Diseases. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L797–L805. [Google Scholar] [CrossRef]
- Wang, X.; Gray, Z.; Willette-Brown, J.; Zhu, F.; Shi, G.; Jiang, Q.; Song, N.-Y.; Dong, L.; Hu, Y. Macrophage Inducible Nitric Oxide Synthase Circulates Inflammation and Promotes Lung Carcinogenesis. Cell Death Discov. 2018, 4, 46. [Google Scholar] [CrossRef]
- Betakova, T.; Kostrabova, A.; Lachova, V.; Turianova, L. Cytokines Induced During Influenza Virus Infection. Curr. Pharm. Des. 2017, 23, 1. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and Immunological Features of Severe and Moderate Coronavirus Disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Rossaint, J.; Nadler, J.L.; Ley, K.; Zarbock, A. Eliminating or Blocking 12/15-Lipoxygenase Reduces Neutrophil Recruitment in Mouse Models of Acute Lung Injury. Crit. Care 2012, 16, R166. [Google Scholar] [CrossRef]
- Danilov, A.V.; Hu, S.; Orr, B.; Godek, K.; Mustachio, L.M.; Sekula, D.; Liu, X.; Kawakami, M.; Johnson, F.M.; Compton, D.A.; et al. Dinaciclib Induces Anaphase Catastrophe in Lung Cancer Cells via Inhibition of Cyclin-Dependent Kinases 1 and 2. Mol. Cancer Ther. 2016, 15, 2758–2766. [Google Scholar] [CrossRef]
- Babu, B.R.; Griffith, O.W. Design of Isoform-Selective Inhibitors of Nitric Oxide Synthase. Curr. Opin. Chem. Biol. 1998, 2, 491–500. [Google Scholar] [CrossRef]
- Morse, J.S.; Lalonde, T.; Xu, S.; Liu, W.R. Learning from the Past: Possible Urgent Prevention and Treatment Options for Severe Acute Respiratory Infections Caused by 2019-nCoV. ChemBioChem 2020, 21, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.T.; Serrano, M.L.; Pujol, F.H.; Rangel, H.R. Unrevealing Sequence and Structural Features of Novel Coronavirus Using in Silico Approaches: The Main Protease as Molecular Target. EXCLI J. 2020, 19, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of Therapeutic Targets for SARS-CoV-2 and Discovery of Potential Drugs by Computational Methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef] [PubMed]
- Avram, S.; Milac, A.-L.; Borcan, L.-C.; Mihailescu, D.; Borcan, F.; Castanho, M. Designing of Artificial Peptides for an Improved Antiviral Activity. Curr. Proteom. 2018, 15, 258–266. [Google Scholar] [CrossRef]
- Avram, S.; Puia, A.; Udrea, A.M.; Mihailescu, D.; Mernea, M.; Dinischiotu, A.; Oancea, F.; Stiens, J. Natural Compounds Therapeutic Features in Brain Disorders by Experimental, Bioinformatics and Cheminformatics Methods. Curr. Med. Chem. 2020, 27, 78–98. [Google Scholar] [CrossRef]
- Zhi, H.-J.; Zhu, H.-Y.; Zhang, Y.-Y.; Lu, Y.; Li, H.; Chen, D.-F. In Vivo Effect of Quantified Flavonoids-Enriched Extract of Scutellaria Aicalensis Root on Acute Lung Injury Induced by Influenza A Virus. Phytomedicine 2019, 57, 105–116. [Google Scholar] [CrossRef]
- Udrea, A.-M.; Avram, S.; Nistorescu, S.; Pascu, M.-L.; Romanitan, M.O. Laser Irradiated Phenothiazines: New Potential Treatment for COVID-19 Explored by Molecular Docking. J. Photochem. Photobiol. B Biol. 2020, 211, 111997. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, X.-Y.; Martin, C. Scutellaria Baicalensis, the Golden Herb from the Garden of Chinese Medicinal Plants. Sci. Bull. 2016, 61, 1391–1398. [Google Scholar] [CrossRef]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A Review of Its Traditional Uses, Botany, Phytochemistry, Pharmacology and Toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, S.; DasSharma, S.; Banik, R.; Chakraborty, A.; Dinda, M. Therapeutic Potentials of Baicalin and Its Aglycone, Baicalein against Inflammatory Disorders. Eur. J. Med. Chem. 2017, 131, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Dong, Y.; Gao, Y.; Du, Z.; Wang, Y.; Cheng, P.; Chen, A.; Huang, H. The Fascinating Effects of Baicalein on Cancer: A Review. Int. J. Mol. Sci. 2016, 17, 1681. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.M.; Haseeb, A.; Ansari, M.Y.; Devarapalli, P.; Haynie, S.; Haqqi, T.M. Wogonin, a Plant Derived Small Molecule, Exerts Potent Anti-Inflammatory and Chondroprotective Effects through the Activation of ROS/ERK/Nrf2 Signaling Pathways in Human Osteoarthritis Chondrocytes. Free. Radic. Biol. Med. 2017, 106, 288–301. [Google Scholar] [CrossRef]
- Shimizu, T.; Shibuya, N.; Narukawa, Y.; Oshima, N.; Hada, N.; Kiuchi, F. Synergistic Effect of Baicalein, Wogonin and Oroxylin A Mixture: Multistep Inhibition of the NF-ΚB Signalling Pathway Contributes to an Anti-Inflammatory Effect of Scutellaria Root Flavonoids. J. Nat. Med. 2018, 72, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Song, H.-H.; Lee, J.-S.; Ko, H.-J.; Song, J.-H. Inhibitory Effects of Norwogonin, Oroxylin A, and Mosloflavone on Enterovirus 71. Biomol. Ther. 2016, 24, 552–558. [Google Scholar] [CrossRef]
- Song, J.-W.; Long, J.-Y.; Xie, L.; Zhang, L.-L.; Xie, Q.-X.; Chen, H.-J.; Deng, M.; Li, X.-F. Applications, Phytochemistry, Pharmacological Effects, Pharmacokinetics, Toxicity of Scutellaria Baicalensis Georgi. and Its Probably Potential Therapeutic Effects on COVID-19: A Review. Chin. Med. 2020, 15, 102. [Google Scholar] [CrossRef]
- Song, J.; Zhang, L.; Xu, Y.; Yang, D.; Zhang, L.; Yang, S.; Zhang, W.; Wang, J.; Tian, S.; Yang, S.; et al. The Comprehensive Study on the Therapeutic Effects of Baicalein for the Treatment of COVID-19 In Vivo and In Vitro. Biochem. Pharmacol. 2021, 183, 114302. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of Flavonoids against Coronavirus Infection. Chemico-Biol. Interact. 2020, 328, 109211. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, W.; Ni, Y.; Li, M.; Chen, J.; Liu, X.; Tan, X.; Li, J. Therapeutic Mechanism of Toujie Quwen Granules in COVID-19 Based on Network Pharmacology. BioData Min. 2020, 13, 15. [Google Scholar] [CrossRef]
- Su, H.; Yao, S.; Zhao, W.; Li, M.; Liu, J.; Shang, W.; Xie, H.; Ke, C.; Hu, H.; Gao, M.; et al. Anti-SARS-CoV-2 Activities in Vitro of Shuanghuanglian Preparations and Bioactive Ingredients. Acta Pharmacol. Sin. 2020, 41, 1167–1177. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Bai, C.; He, F.; Xie, Y.; Zhou, H. Review on the Potential Action Mechanisms of Chinese Medicines in Treating Coronavirus Disease 2019 (COVID-19). Pharmacol. Res. 2020, 158, 104939. [Google Scholar] [CrossRef] [PubMed]
- Vilar, S.; Cozza, G.; Moro, S. Medicinal Chemistry and the Molecular Operating Environment (MOE): Application of QSAR and Molecular Docking to Drug Discovery. Curr. Top. Med. Chem. 2008, 8, 1555–1572. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings 1PII of Original Article: S0169-409X9600423-1. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A Knowledge-Based Approach in Designing Combinatorial or Medicinal Chemistry Libraries for Drug Discovery. 1. A Qualitative and Quantitative Characterization of Known Drug Databases. J. Comb. Chem. 1999, 1, 55–68. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular Properties That Influence the Oral Bioavailability of Drug Candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Egan, W.J.; Merz, K.M.; Baldwin, J.J. Prediction of Drug Absorption Using Multivariate Statistics. J. Med. Chem. 2000, 43, 3867–3877. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Udrea, A.-M. Computational Approaches of New Perspectives in the Treatment of Depression during Pregnancy. Farmacia 2018, 66, 680–687. [Google Scholar] [CrossRef]
- Wood, D.J.; De Vlieg, J.; Wagener, M.; Ritschel, T. Pharmacophore Fingerprint-Based Approach to Binding Site Subpocket Similarity and its Application to Bioisostere Replacement. J. Chem. Inf. Model. 2012, 52, 2031–2043. [Google Scholar] [CrossRef]
- Rácz, A.; Bajusz, D.; Héberger, K. Life beyond the Tanimoto Coefficient: Similarity Measures for Interaction Fingerprints. J. Cheminform. 2018, 10, 48. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated Data and New Features for Efficient Prediction of Protein Targets of Small Molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Bank, R.P.D. RCSB PDB—6M2N: SARS-CoV-2 3CL Protease (3CL pro) in Complex with a Novel Inhibitor. Available online: https://www.rcsb.org/structure/6M2N (accessed on 13 October 2020).
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a Protein-Small Molecule Docking Web Service Based on EADock DSS. Nucleic Acids Res. 2011, 39 (Suppl. 2), W270–W277. [Google Scholar] [CrossRef] [PubMed]
- Grosdidier, A.; Zoete, V.; Michielin, O. Fast Docking Using the CHARMM Force Field with EADock DSS. J. Comput. Chem. 2011, 32, 2149–2159. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Hastings, J.; Owen, G.; Dekker, A.; Ennis, M.; Kale, N.; Muthukrishnan, V.; Turner, S.; Swainston, N.; Mendes, P.; Steinbeck, C. ChEBI in 2016: Improved Services and an Expanding Collection of Metabolites. Nucleic Acids Res. 2016, 44, D1214–D1219. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand-Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. PkCSM: Predicting Small-Molecule Pharmacokinetic and Toxicity Properties Using Graph-Based Signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Drwal, M.N.; Banerjee, P.; Dunkel, M.; Wettig, M.R.; Preissner, R. ProTox: A Web Server for the in Silico Prediction of Rodent Oral Toxicity. Nucleic Acids Res. 2014, 42, W53–W58. [Google Scholar] [CrossRef]
- Lovering, F.; Bikker, J.; Humblet, C. Escape from Flatland: Increasing Saturation as an Approach to Improving Clinical Success. J. Med. Chem. 2009, 52, 6752–6756. [Google Scholar] [CrossRef]
- Chen, H.; Guan, B.; Chen, X.; Chen, X.; Li, C.; Qiu, J.; Yang, D.; Liu, K.J.; Qi, S.; Shen, J. Baicalin Attenuates Blood-Brain Barrier Disruption and Hemorrhagic Transformation and Improves Neurological Outcome in Ischemic Stroke Rats with Delayed t-PA Treatment: Involvement of ONOO-MMP-9 Pathway. Transl. Stroke Res. 2018, 9, 515–529. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kang, H.-C.; Shim, J.; Sohn, N.-W. Scutellaria Baicalensis Attenuates Blood-Brain Barrier Disruption after Intracerebral Hemorrhage in Rats. Am. J. Chin. Med. 2012, 40, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, N.; Guo, Q.; Ji, H.; Zhao, D.; Xie, S.; Li, X.; Qiu, Z.; Han, D.; Chen, X.; et al. Inhibitory Effects of Wogonin on Catalytic Activity of Cytochrome P450 Enzyme in Human Liver Microsomes. Eur. J. Drug Metab. Pharmacokinet. 2011, 36, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Yang, Y.; Wang, X.; Wei, M.; Guo, Q.; Zhao, L. Oroxyloside Ameliorates Acetaminophen-Induced Hepatotoxicity by Inhibiting JNK Related Apoptosis and Necroptosis. J. Ethnopharmacol. 2020, 258, 112917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal Structure of SARS-CoV-2 Main Protease Provides a Basis for Design of Improved α-Ketoamide Inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Labbé, R.M.; Holowatyj, A.; Yang, Z.-Q. Histone Lysine Demethylase (KDM) Subfamily 4: Structures, Functions and Therapeutic Potential. Am. J. Transl. Res. 2013, 6, 1–15. [Google Scholar]
- Saul, D.; Weber, M.; Zimmermann, M.H.; Kosinsky, R.L.; Hoffmann, D.B.; Menger, B.; Taudien, S.; Lehmann, W.; Komrakova, M.; Sehmisch, S. Effect of the Lipoxygenase Inhibitor Baicalein on Bone Tissue and Bone Healing in Ovariectomized Rats. Nutr. Metab. Lond 2019, 16, 4. [Google Scholar] [CrossRef]
- Hsu, S.-L.; Hsieh, Y.-C.; Hsieh, W.-C.; Chou, C.-J. Baicalein Induces a Dual Growth Arrest by Modulating Multiple Cell Cycle Regulatory Molecules. Eur. J. Pharmacol. 2001, 425, 165–171. [Google Scholar] [CrossRef]
- Avram, S.; Bologa, C.G.; Holmes, J.; Bocci, G.; Wilson, T.B.; Nguyen, D.-T.; Curpan, R.; Halip, L.; Bora, A.; Yang, J.J.; et al. DrugCentral 2021 supports drug discovery and repositioning. Nucleic Acids Res. 2020, gkaa997. [Google Scholar] [CrossRef]
- Ni, T.; He, Z.; Dai, Y.; Yao, J.; Guo, Q.; Wei, L. Oroxylin A Suppresses the Development and Growth of Colorectal Cancer through Reprogram of HIF1α-Modulated Fatty Acid Metabolism. Cell Death Dis. 2017, 8, e2865. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Shen, S.-C.; Chen, L.-G.; Lee, T.J.-F.; Yang, L.-L. Wogonin, Baicalin, and Baicalein Inhibition of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Gene Expressions Induced by Nitric Oxide Synthase Inhibitors and Lipopolysaccharide11. Biochem. Pharmacol. 2001, 61, 1417–1427. [Google Scholar] [CrossRef]
- Kim, H.P.; Son, K.H.; Chang, H.W.; Kang, S.S. Anti-Inflammatory Plant Flavonoids and Cellular Action Mechanisms. J. Pharmacol. Sci. 2004, 96, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Yang, C.; Yu, Z.; Li, X.; Mu, Q.; Liao, G.; Yu, B. Natural Products as LSD1 Inhibitors for Cancer Therapy. Acta Pharm. Sin. B 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-C.; Shen, D.-D.; Ren, M.; Liu, X.-Q.; Wang, Z.-R.; Liu, Y.; Zhang, Q.-N.; Zhao, L.-J.; Zhao, L.-J.; Ma, J.-L.; et al. Baicalin, a Natural LSD1 Inhibitor. Bioorganic Chem. 2016, 69, 129–131. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IUPAC Name | XLogP3 | Molecular Weight g/mol | H-Bond Donor | H-Bond Acceptor |
|---|---|---|---|---|---|
| baicalein | 5,6,7-trihydroxy-2-phenylchromen-4-one | 1.7 | 270.24 | 3 | 5 |
| baicalin | (2S,3S,4S,5R,6S)-6-(5,6-dihydroxy-4-oxo-2-phenylchromen-7-yl)oxy-3,4,5-trihydroxyoxane-2-carboxylic acid | 1.1 | 446.4 | 6 | 11 |
| wogonin | 5,7-dihydroxy-8-methoxy-2-phenylchromen-4-one | 3 | 284.26 | 2 | 5 |
| norwogonin | 5,7,8-trihydroxy-2-phenylchromen-4-one | 2.7 | 270.24 | 3 | 5 |
| oroxylin A | 5,7-dihydroxy-6-methoxy-2-phenylchromen-4-one | 2.1 | 284.26 | 2 | 5 |
| Chemical Name | PubChem ID | SMILES | Chemical-Disease Associations |
|---|---|---|---|
| baicalein | 5281605 | C1=CC=C(C=C1)C2=CC(=O)C3=C(O2)C=C(C(=C3O)O)O | liver injury, fibrosis, glioblastoma, hyperalgesia |
| baicalin | 64982 | C1=CC=C(C=C1)C2=CC(=O)C3=C(C(=C(C=C3O2)OC4C(C(C(C(O4)C(=O)O)O)O)O)O)O | kidney injury, brain ischemia, fibrosis, inflammation, liver cirrhosis |
| wogonin | 5281703 | COC1=C(C=C(C2=C1OC(=CC2=O)C3=CC=CC=C3)O)O | kidney injury, acute lung injury edema, gastrointestinal, neoplasms, glioma |
| norwogonin | 5281674 | C1=CC=C(C=C1)C2=CC(=O)C3=C(O2)C(=C(C=C3O)O)O | HIV-1 antiviral |
| oroxylin A | 5320315 | COC1=C(C2=C(C=C1O)OC(=CC2=O)C3=CC=CC=C3)O | bacterial infections, bradycardia, hypotension, inflammation |
| Compounds | Lipinski | Ghose | Veber | Egan | Lead-Likeness |
|---|---|---|---|---|---|
| baicalein | yes | yes | yes | yes | yes |
| baicalin | no | yes | no | no | no |
| wogonin | yes | yes | yes | yes | yes |
| norwogonin | yes | yes | yes | yes | yes |
| oroxylin A | yes | yes | yes | yes | yes |
| ADME | Baicalein | Baicalin | Wogonin | Norwogonin | Oroxylin A |
|---|---|---|---|---|---|
| ABSORBTION | |||||
| Caco2 permeability numeric (log Papp in 10−6 cm) | 1.11 | −0.67 | 0.96 | 1.10 | 1.02 |
| Intestinal absorption (human) numeric (% absorbed) | 94.26 | 26.22 | 92.68 | 94.48 | 94.34 |
| P-glycoprotein substrate | yes | yes | yes | yes | yes |
| DISTRIBUTION | |||||
| Fraction unbound (human) (Fu) | 0.15 | 0.29 | 0.10 | 0.14 | 0.082 |
| BBB permeability (log BBB) | −1.06 | −1.33 | −2.23 | −0.96 | −0.11 |
| CNS permeability (Log PS) | −2.21 | −3.81 | −2.18 | −2.15 | −2.21 |
| METABOLISM | |||||
| CYP1A2 inhibitor | yes | no | yes | yes | yes |
| CYP2D6 substrate/inhibitors | no | no | no | no | no |
| CYP3A4 substrate/inhibitors | no | no | yes | no | yes |
| EXCRETION | |||||
| Total clearance (log mL/min/kg) | 0.25 | 0.04 | 0.29 | 0.25 | 0.31 |
| Renal OCT2 substrate | no | no | no | no | no |
| TOXICITY | |||||
| hepatotoxicity | inactive (0.69) | inactive (0.75) | inactive (0.72) | inactive (0.69) | inactive (0.72) |
| carcinogenicity | active (0.68) | active (0.50) | active (0.68) | active (0.68) | inactive (0.68) |
| immunotoxicity | inactive (0.99) | inactive (0.92) | inactive (0.80) | inactive (0.96) | inactive (0.81) |
| mutagenicity | active (0.51) | inactive (0.68) | inactive (0.94) | active (0.51) | inactive (0.94) |
| cytotoxicity | inactive (0.99) | inactive (0.91) | inactive (0.95) | inactive (0.99) | inactive (0.95) |
| hERG I/II inhibitors | no | no | no | no | no |
| activity on aryl hydrogen receptor | active (0.91) | inactive (0.60) | inactive (0.60) | active (0.91) | active (0.97) |
| activity on mitochondrial membrane potential | active (1.00) | inactive (0.63) | active (0.92) | active (1.00) | active (0.92) |
| activity on phosphoprotein tumor suppressor (p53) | inactive (0.97) | inactive (0.89) | inactive (0.86) | inactive (0.97) | inactive (0.86) |
| Max. tolerated dose (human) (log mg/kg/day) | 0.49 | 0.65 | 0.15 | 0.48 | −0.08 |
| Compounds | AM1-LUMO eV | AM1-HOMO eV | vsurf_S Å2 | CASA_POS Å2 | CASA_NEG Å2 |
|---|---|---|---|---|---|
| baicalein | −0.71 | −8.93 | 417.42 | 577.84 | 337.03 |
| baicalin | −0.77 | −8.83 | 592.19 | 1655.53 | 945.98 |
| wogonin | −0.70 | −9.03 | 445.75 | 671.13 | 287.35 |
| norwogonin | −0.67 | −8.78 | 418.22 | 579.01 | 327.94 |
| oroxylin A | −0.73 | −9.12 | 443.53 | 689.63 | 282.60 |
| Enzymes | Baicalein Binding Probability | Wogonin Binding Probability | Norwogonin Binding Probability | Oroxylin A Binding Probability |
|---|---|---|---|---|
| KDM4D | 1 | - | 0.60 | 0.18 |
| XDH | 1 | 0.10 | 0.60 | 0.18 |
| ALOX15 | 1 | 0.13 | 0.62 | 0.19 |
| CDK1 | 1 | 0.10 | 0.62 | 0.18 |
| COX-2 | 0.30 | 1 | 0.17 | 0.30 |
| iNOS | 0.30 | 1 | 0.16 | 0.60 |
| Enzymes | Baicalein Experimental Activity | Baicalin Experimental Activity | Norwogonin Experimental Activity | Wogonin Experimental Activity | Oroxylin A Experimental Activity |
|---|---|---|---|---|---|
| XDH | - | - | - | - | - |
| ALOX15 | yes-inhibitor [68] | - | - | - | - |
| CDK1 | yes-inhibitor [69] | yes-inhibitor [69,70] | - | - | yes-inhibitor [71] |
| COX-2 | yes-inhibitor [68] not inhibited [72] | not inhibited [72] | - | yes-inhibitor [73] not inhibited [72] | - |
| iNOS | not inhibited [72] | not inhibited [72] | - | not inhibited [73] not inhibited [72] | - |
| LSD1 | yes-inhibitor [74] | yes-inhibitor [75] | - | yes-inhibitor [74] | yes-inhibitor [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Udrea, A.-M.; Mernea, M.; Buiu, C.; Avram, S. Scutellaria baicalensis Flavones as Potent Drugs against Acute Respiratory Injury during SARS-CoV-2 Infection: Structural Biology Approaches. Processes 2020, 8, 1468. https://doi.org/10.3390/pr8111468
Udrea A-M, Mernea M, Buiu C, Avram S. Scutellaria baicalensis Flavones as Potent Drugs against Acute Respiratory Injury during SARS-CoV-2 Infection: Structural Biology Approaches. Processes. 2020; 8(11):1468. https://doi.org/10.3390/pr8111468
Chicago/Turabian StyleUdrea, Ana-Maria, Maria Mernea, Cătălin Buiu, and Speranța Avram. 2020. "Scutellaria baicalensis Flavones as Potent Drugs against Acute Respiratory Injury during SARS-CoV-2 Infection: Structural Biology Approaches" Processes 8, no. 11: 1468. https://doi.org/10.3390/pr8111468
APA StyleUdrea, A.-M., Mernea, M., Buiu, C., & Avram, S. (2020). Scutellaria baicalensis Flavones as Potent Drugs against Acute Respiratory Injury during SARS-CoV-2 Infection: Structural Biology Approaches. Processes, 8(11), 1468. https://doi.org/10.3390/pr8111468