
Simultaneous Improvement of Final Product-Tolerance and Thermostability of GH39 Xylosidase for Prebiotic Production by Directed Evolution

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Biochemical Reagents
2.2. Construction of Mutant Library
2.3. High-Throughput Screening of Mutant Library
2.4. Expression, Purification and Tolerance Assays of Wild Type and Variants
2.5. Construction and Characteristics of Single Site Variants
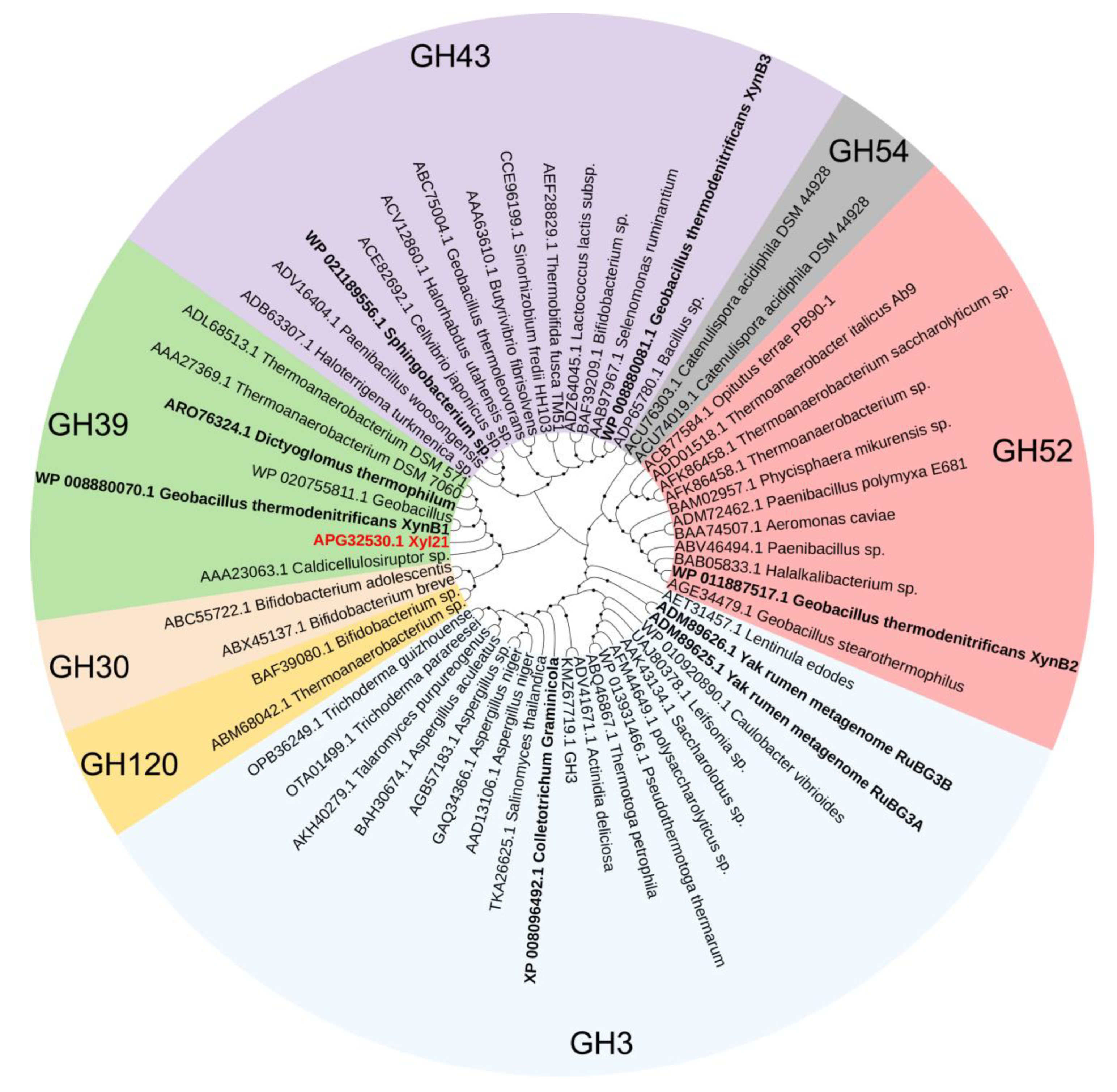
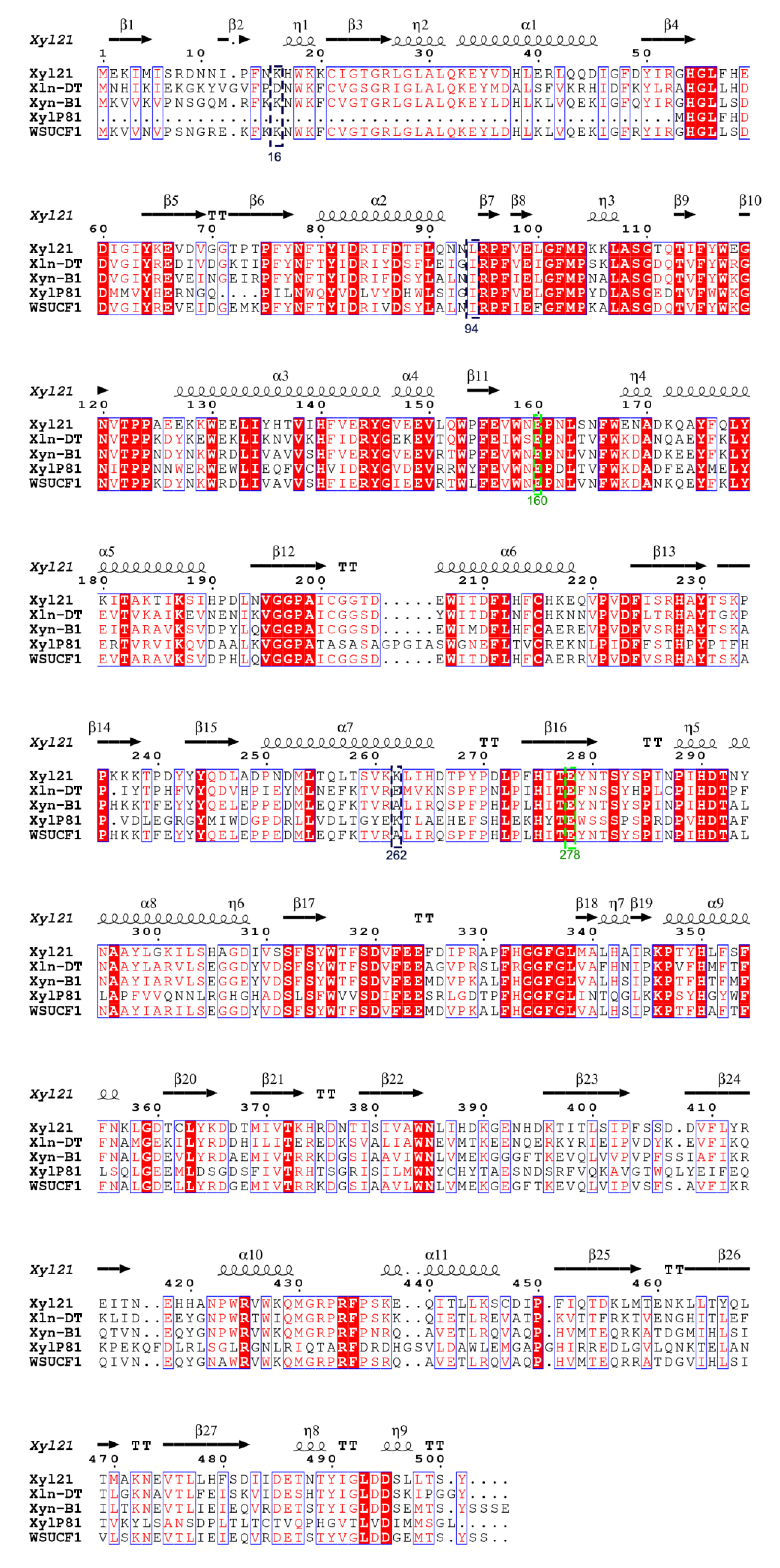
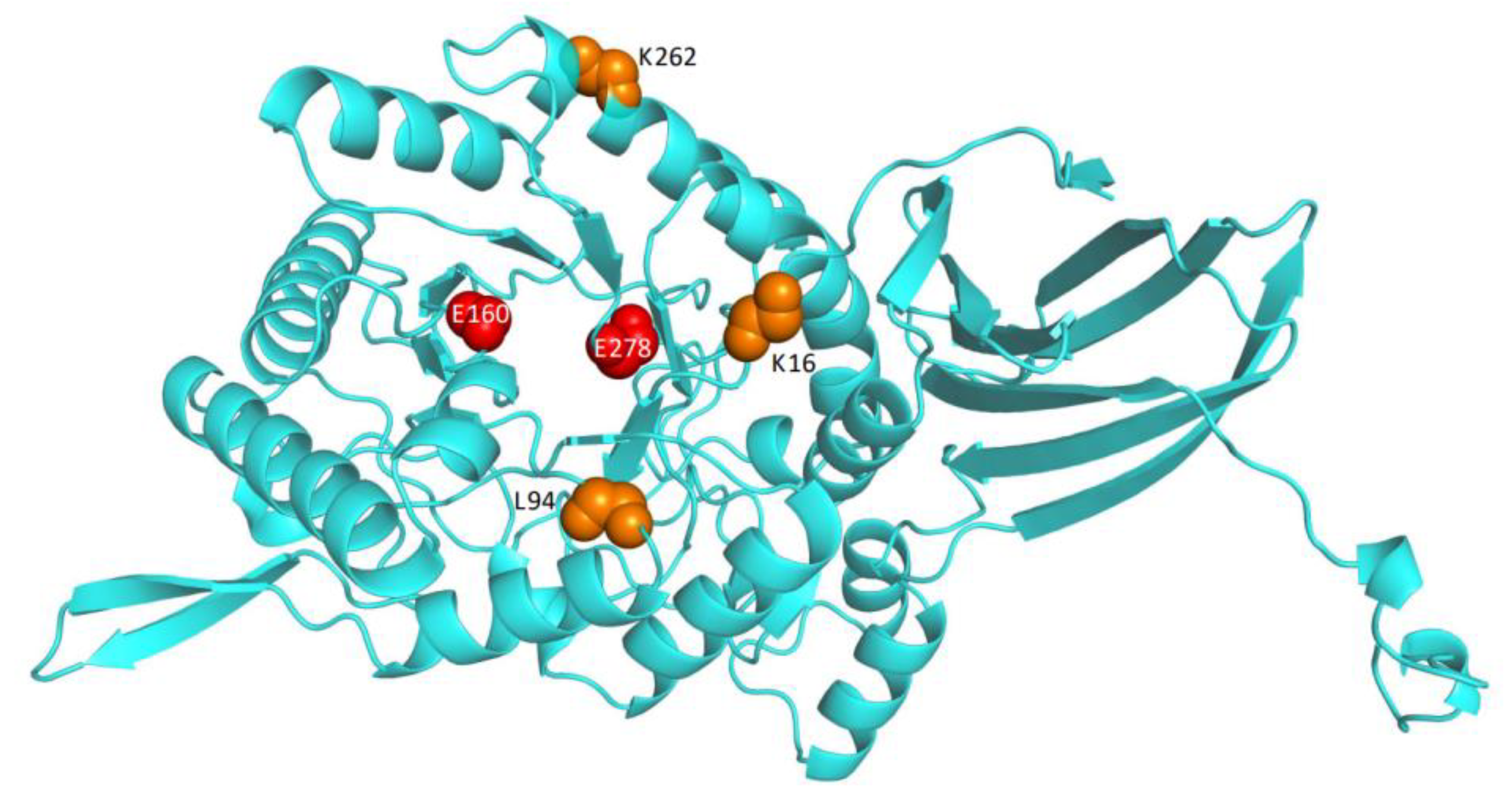
2.6. Sequence Alignment, Homology Modeling, and Structural Analysis
2.7. Statistical Analysis
3. Results and Discussion
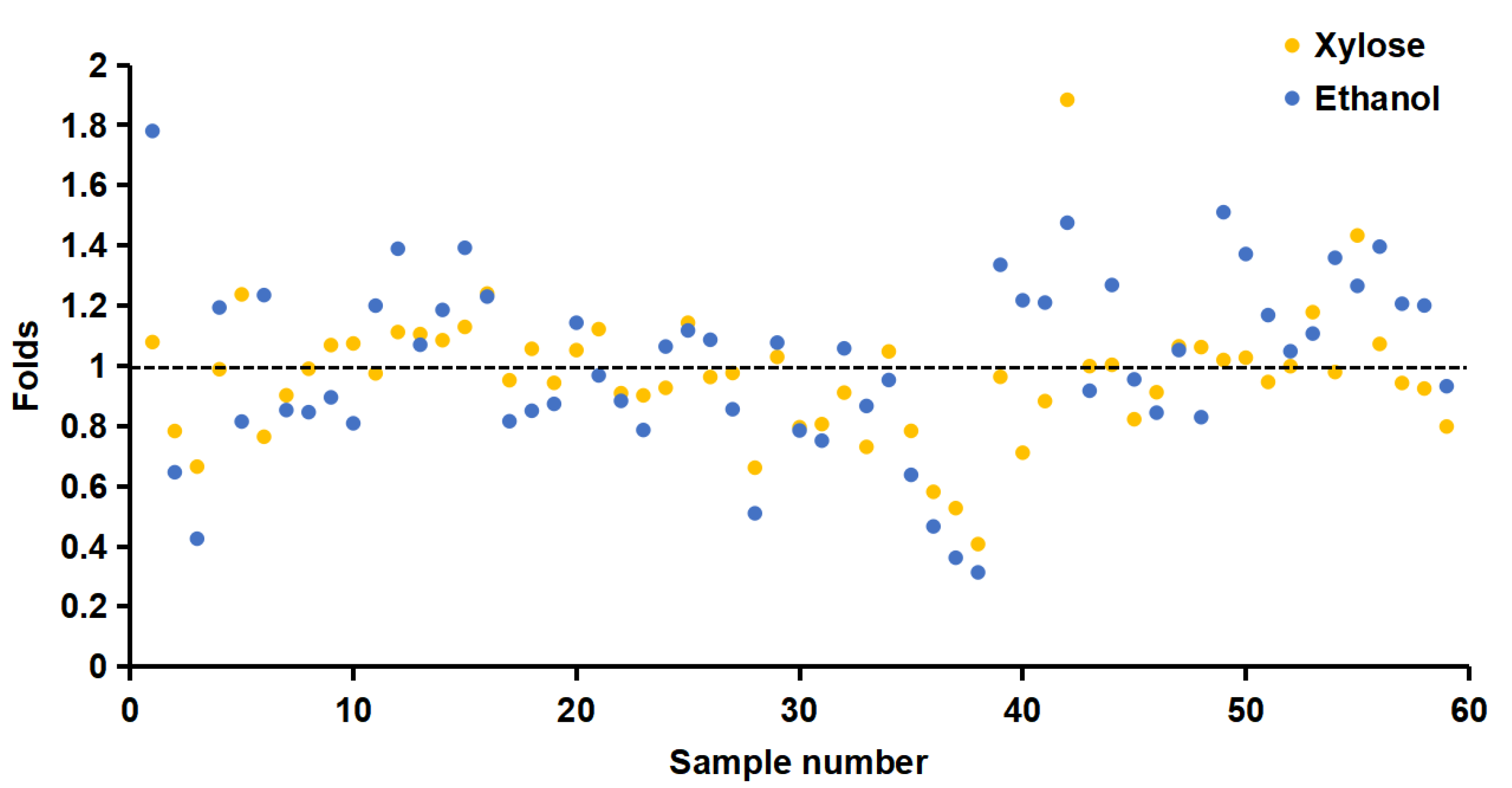
3.1. Construction of the Random Mutagenesis Library by Error-Prone PCR
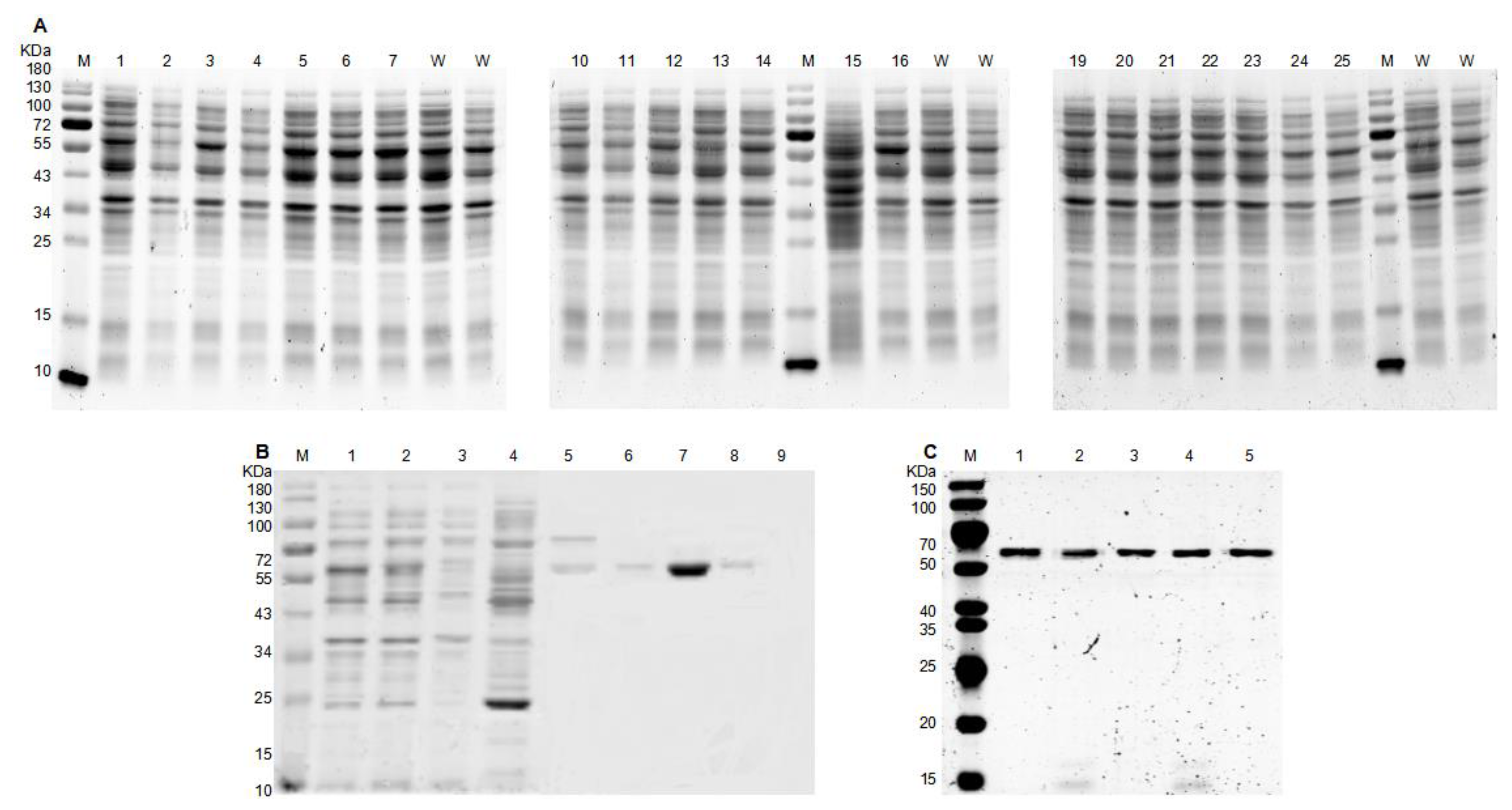
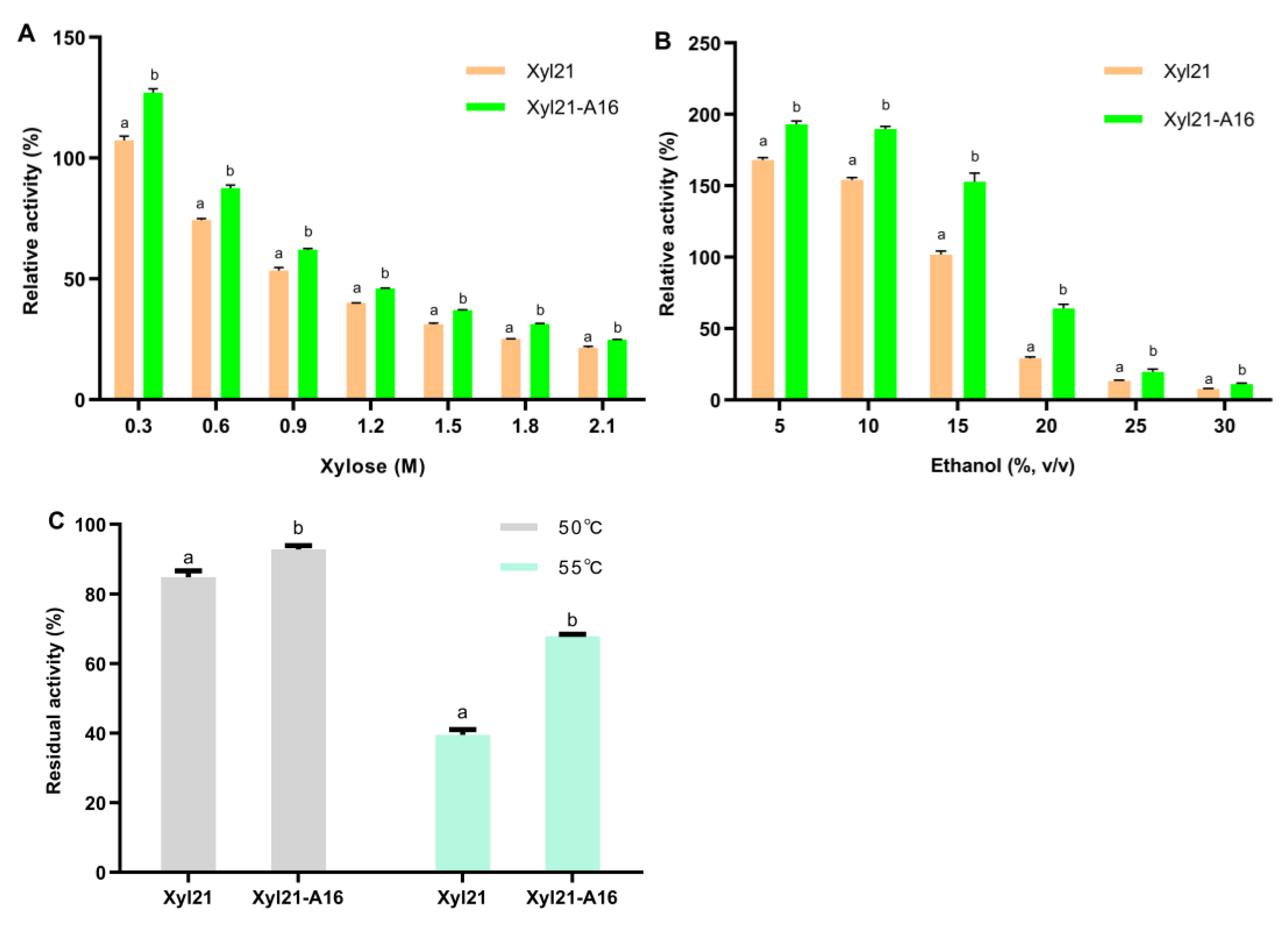
3.2. Protein Expression, Purification, and Characterization of Mutants
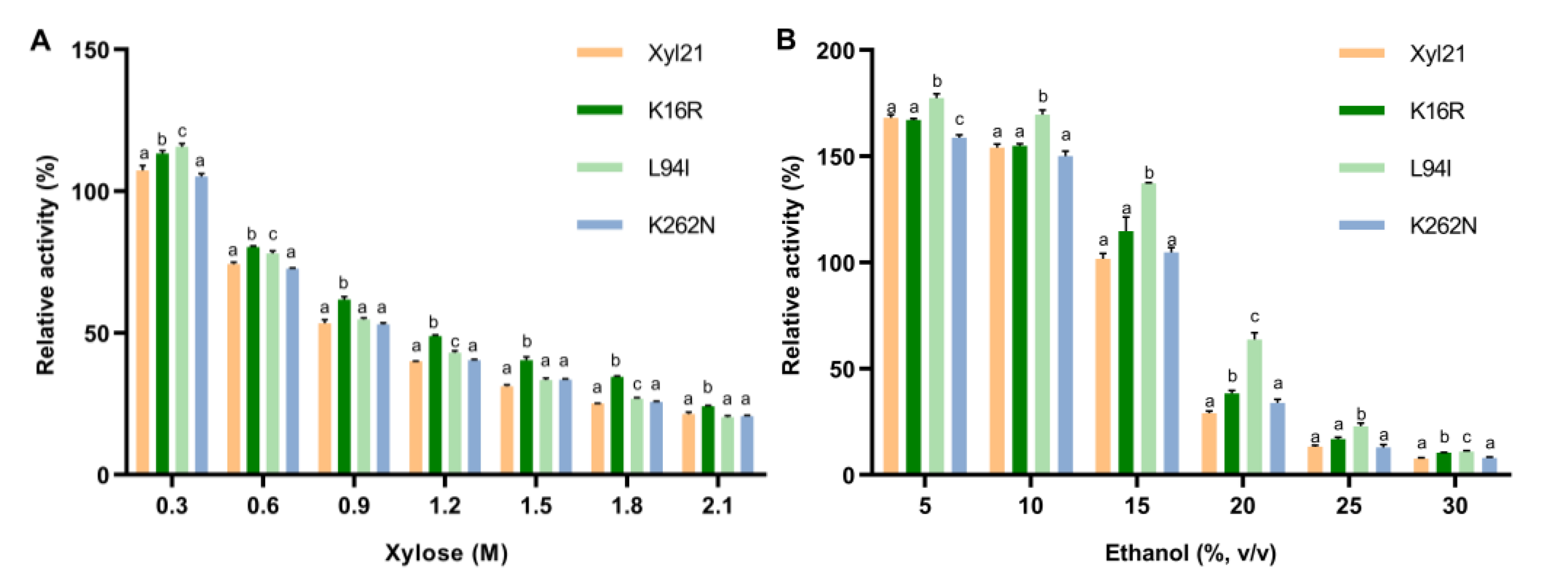
3.3. Xylose and Ethanol Tolerance Analysis of Single Site Mutants K16R, L94I and K262N
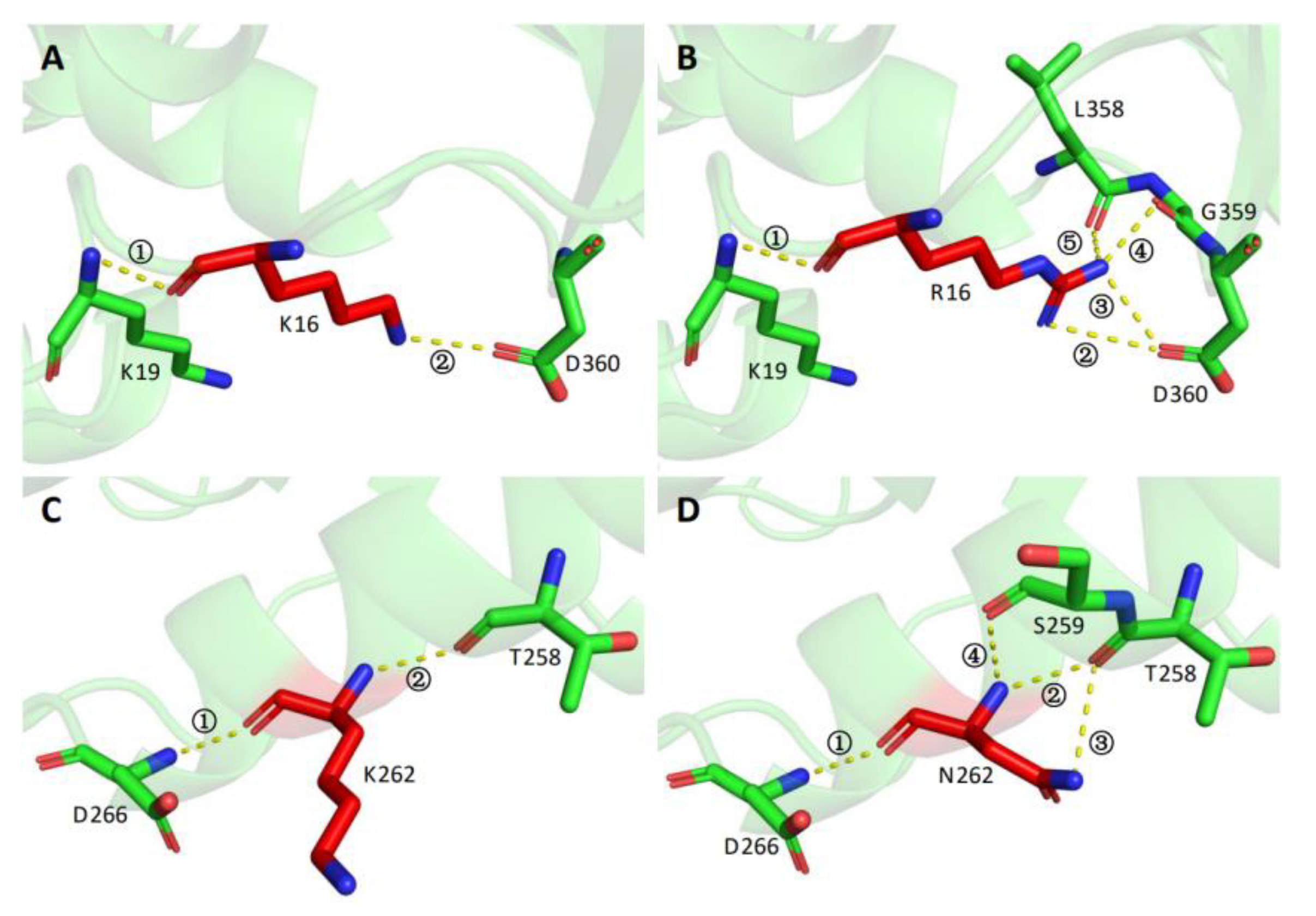
3.4. Protein Structure Modeling and Analysis of Intramolecular Interactions in the Wild Type and the Variants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.; Dilokpimol, A.; Kabel, M.A.; de Vries, R.P. Fungal xylanolytic enzymes: Diversity and applications. Bioresour. Technol. 2022, 344, 126290. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, C.; Terrone, C.C.; Masarin, F.; Carmona, E.C.; Brienzo, M. In vitro study of the effect of xylooligosaccharides obtained from banana pseudostem xylan by enzymatic hydrolysis on probiotic bacteria. Biocatal. Agric. Biotechnol. 2021, 33, 101973. [Google Scholar] [CrossRef]
- Capetti, C.C.M.; Vacilotto, M.M.; Dabul, A.N.G.; Sepulchro, A.G.V.; Pellegrini, V.O.A.; Polikarpov, I. Recent advances in the enzymatic production and applications of xylooligosaccharides. World J. Microbiol. Biotechnol. 2021, 37, 169. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Domínguez, M.; de Eugenio, L.I.; Peñalver, P.; Belmonte-Reche, E.; Morales, J.C.; Poveda, A.; Jiménez-Barbero, J.; Prieto, A.; Plou, F.J.; Martínez, M.J. Enzymatic synthesis of a novel neuroprotective hydroxytyrosyl glycoside. J. Agric. Food Chem. 2017, 65, 10526–10533. [Google Scholar] [CrossRef]
- Nieto-Domínguez, M.; de Eugenio, L.I.; Barriuso, J.; Prieto, A.; Fernández de Toro, B.; Canales-Mayordomo, Á.; Martínez, M.J. Novel pH-stable glycoside hydrolase family 3 β-xylosidase from Talaromyces amestolkiae: An enzyme displaying regioselective transxylosylation. Appl. Environ. Microbiol. 2015, 81, 6380–6392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, N.; Xu, S.; Han, X.; Li, C.; Wei, X.; Liu, Y.; Tu, T.; Tang, X.; Zhou, J.; et al. Glycoside hydrolase family 39 β-xylosidases exhibit β-1,2-xylosidase activity for transformation of notoginsenosides: A new EC subsubclass. J. Agric. Food Chem. 2019, 67, 3220–3228. [Google Scholar] [CrossRef]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng. Res. 2017, 41, 435–443. [Google Scholar] [CrossRef]
- Liu, W.C.; Gong, T.; Zhu, P. Advances in exploring alternative Taxol sources. RSC. Adv. 2016, 6, 48800–48809. [Google Scholar] [CrossRef]
- Rohman, A.; Dijkstra, B.W.; Puspaningsih, N.N.T. β-Xylosidases: Structural diversity, catalytic mechanism, and inhibition by monosaccharides. Int. J. Mol. Sci. 2019, 20, 5534. [Google Scholar] [CrossRef]
- Smułek, W.; Kaczorek, E.; Hricovíniová, Z. Alkyl Xylosides: Physico-chemical properties and influence on environmental bacteria cells. J. Surfactants Deterg. 2017, 20, 1269–1279. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Chen, S.; Wang, Y.; Liu, Z.; Xia, Z.; Zhang, M.; Luo, X.; Song, Y.; Zhao, J.; Zhang, T. Biochemical characterization of a novel halo/organic-solvents/final-products tolerant GH39 xylosidase from saline soil and its synergic action with xylanase. Int. J. Biol. Macromol. 2020, 164, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Shukla, P. Current trends in protein engineering: Updates and progress. Curr. Protein Pept. Sci. 2019, 20, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Kan, S.B.; Lewis, R.D.; Chen, K.; Arnold, F.H. Directed evolution of cytochrome c for carbon-silicon bond formation: Bringing silicon to life. Science 2016, 354, 1048–1051. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Fu, C.; Huang, W. Improving the activity of the endoglucanase, Cel8M from Escherichia. coli by error-prone PCR. Enzyme Microb. Technol. 2016, 86, 52–58. [Google Scholar] [CrossRef]
- Huang, L.; Zheng, D.; Zhao, Y.; Ma, J.; Li, Y.; Xu, Z.; Shan, M.; Shao, S.; Guo, Q.; Zhang, J.; et al. Improvement of the alkali stability of Penicillium cyclopium lipase by error-prone PCR. Electron. J. Biotechnol. 2019, 39, 91–97. [Google Scholar] [CrossRef]
- Li, Q.; Wu, T.; Qi, Z.; Zhao, L.; Pei, J.; Tang, F. Characterization of a novel thermostable and xylose-tolerant GH39 β-xylosidase from Dictyoglomus thermophilum. BMC. Biotechnol. 2018, 18, 29. [Google Scholar] [CrossRef]
- Huang, D.; Liu, J.; Qi, Y.; Yang, K.; Xu, Y.; Feng, L. Synergistic hydrolysis of xylan using novel xylanases, β-xylosidases, and an α-L-arabinofuranosidase from Geobacillus thermodenitrificans NG80-2. Appl. Microbiol. Biotechnol. 2017, 101, 6023–6037. [Google Scholar] [CrossRef]
- Maruthamuthu, M.; Jiménez, D.J.; van Elsas, J.D. Characterization of a furan aldehyde-tolerant β-xylosidase/α-arabinosidase obtained through a synthetic metagenomics approach. J. Appl. Microbiol. 2017, 123, 145–158. [Google Scholar] [CrossRef]
- Bao, L.; Huang, Q.; Chang, L.; Sun, Q.; Zhou, J.; Lu, H. Cloning and characterization of two β-glucosidase/xylosidase enzymes from yak rumen metagenome. Appl. Biochem. Biotechnol. 2012, 166, 72–86. [Google Scholar] [CrossRef]
- Carvalho, D.R.; Carli, S.; Meleiro, L.P.; Rosa, J.C.; Oliveira, A.H.C.; Jorge, J.A.; Furriel, R.P.M. A halotolerant bifunctional β-xylosidase/α-l-arabinofuranosidase from Colletotrichum graminicola: Purification and biochemical characterization. Int. J. Biol. Macromol. 2018, 114, 741–750. [Google Scholar] [CrossRef]
- Romero, A.M.; Mateo, J.J.; Maicas, S. Characterization of an ethanol-tolerant 1,4-β-xylosidase produced by Pichia membranifaciens. Lett. Appl. Microbiol. 2012, 55, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Zhu, C.; Wang, Z.; Luo, H.; Fu, R.; Zhao, X.; Zhao, X.; Jiang, L. Purification and characterization of a glucose-tolerant β-glucosidase from black plum seed and its structural changes in ionic liquids. Food Chem. 2019, 274, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Bankeeree, W.; Akada, R.; Lotrakul, P.; Punnapayak, H.; Prasongsuk, S. Enzymatic hydrolysis of black liquor xylan by a novel xylose-tolerant, thermostable β-xylosidase from a tropical strain of Aureobasidium pullulans CBS 135684. Appl. Biochem. Biotechnol. 2018, 184, 919–934. [Google Scholar] [PubMed]
- Li, Q.; Tong, X.; Jiang, Y.; Li, D.; Zhao, L. Improvements in xylose stability and thermalstability of GH39 β-xylosidase from Dictyoglomus thermophilum by site-directed mutagenesis and insights into its xylose tolerance mechanism. Enzyme Microb. Technol. 2021, 151, 109921. [Google Scholar] [CrossRef]
- Ndata, K.; Nevondo, W.; Cekuse, B.; van Zyl, L.J.; Trindade, M. Characterization of a highly xylose tolerant β-xylosidase isolated from high temperature horse manure compost. BMC Biotechnol. 2021, 21, 61. [Google Scholar] [CrossRef]
- Kaur, J.; Sharma, R. Directed evolution: An approach to engineer enzymes. Crit. Rev. Biotechnol. 2006, 26, 165–199. [Google Scholar] [CrossRef]
- Ruan, Y.; Zhang, R.; Xu, Y. Directed evolution of maltogenic amylase from Bacillus licheniformis R-53: Enhancing activity and thermostability improves bread quality and extends shelf life. Food Chem. 2022, 381, 132222. [Google Scholar] [CrossRef]
- Álvarez-Machancoses, Ó.; De Andrés-Galiana, E.J.; Fernández-Martínez, J.L.; Kloczkowski, A. Robust prediction of single and multiple point protein mutations stability changes. Biomolecules 2019, 10, 67. [Google Scholar] [CrossRef]
- Isorna, P.; Polaina, J.; Latorre-García, L.; Cañada, F.J.; González, B.; Sanz-Aparicio, J. Crystal structures of Paenibacillus polymyxa beta-glucosidase B complexes reveal the molecular basis of substrate specificity and give new insights into the catalytic machinery of family I glycosidases. J. Mol. Biol. 2007, 371, 1204–1218. [Google Scholar] [CrossRef]
- Uchiyama, T.; Miyazaki, K.; Yaoi, K. Characterization of a novel β-glucosidase from a compost microbial metagenome with strong transglycosylation activity. J. Biol. Chem. 2013, 288, 18325–18334. [Google Scholar]
- Li, N.; Han, X.; Xu, S.; Li, C.; Wei, X.; Liu, Y.; Zhang, R.; Tang, X.; Zhou, J.; Huang, Z. Glycoside hydrolase family 39 β-xylosidase of Sphingomonas showing salt/ethanol/trypsin tolerance, low-pH/low-temperature activity, and transxylosylation activity. J. Agric. Food. Chem. 2018, 66, 9465–9472. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Poljak, A.; Guilhaus, M.; De Francisci, D.; Curmi, P.M.; Feller, G.; D’Amico, S.; Gerday, C.; Uversky, V.N.; Cavicchioli, R. Role of lysine versus arginine in enzyme cold-adaptation: Modifying lysine to homo-arginine stabilizes the cold-adapted alpha-amylase from Pseudoalteramonas haloplanktis. Proteins 2006, 64, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Mukhia, S.; Kumar, R. Industrial applications of cold-adapted enzymes: Challenges, innovations and future perspective. 3 Biotech. 2021, 11, 426. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Zheng, J.; Wang, X.; Wang, X.; Huang, H.; Yang, H.; Tu, T.; Wang, Y.; Bai, Y.; Yao, B.; et al. Improvement of thermostability and catalytic efficiency of glucoamylase from Talaromyces leycettanus JCM12802 via site-directed mutagenesis to enhance industrial saccharification applications. Biotechnol. Biofuels 2021, 14, 202. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Family | Xylose Tolerance | Ethanol Tolerance | Optimum Temperature/°C | Optimum pH | Thermostability/1 min | Reference |
|---|---|---|---|---|---|---|---|
| Metagenomic DNA of soil (Xyl21) | 39 | 111% a (0.3 M xylose), 74% (0.6 M xylose), 53% (0.9 M xylose), 20% (2.1 M xylose) | 176% (5%, v/v), 105% (15%, v/v), 20% (20%, v/v) | 45 | 5.5 | 50 °C, 84%, 55 °C, 39% | This study |
| Metagenomic DNA of soil (Xyl21-A16) | 39 | 130% (0.3 M xylose), 89% (0.6 M xylose), 62% (0.9 M xylose), 20% (2.1 M xylose) | 190% (5%, v/v), 156% (15%, v/v), 63% (20%, v/v) | 45 | 5.5 | 50 °C, 93%, 55 °C, 68% | This study |
| Dictyoglomus thermophilum | 39 | 60% (3 M xylose) | 100% (10%, v/v) | 75 | 6.0 | 85 °C, 90% | [16] |
| Geobacillus thermodenitrificans NG80-2 | 39 | 50% (400 mM xylose) | 85% (5%, v/v) | 60 | 5.5 | 75 °C, 100% | [17] |
| Sphingobacterium sp. | 43 | 100%, 54%, 249%, 91% (20 mM glucose, xylose, arabinose, galactose) | 26% (10%, v/v) | 20 | 8.0 | 30 min, 30 °C, 90% | [18] |
| Geobacillus thermodenitrificans NG80-2 | 43 | 50% (600 mM xylose) | 79% (5%, v/v) | 65 | 6.0 | 75 °C, 90% | [17] |
| Geobacillus thermodenitrificans NG80-2 | 52 | 50% (300 mM xylose) | 81% (5%, v/v) | 65 | 6.0 | 75 °C, 100% | [17] |
| Yak rumen metagenome 3A | 3 | 4% (5 mM glucose), 82% (5 mM xylose) | 112.7% (5%, v/v) | 40 | 7.0 | 50 °C, 75%, 55 °C, 10% | [19] |
| Yak rumen metagenome 3B | 3 | 54% (5 mM glucose), 97% (5 mM xylose) | 317% (5%, v/v) | 40 | 7.0 | 50 °C, 1% | [19] |
| Colletotrichum graminicola | 3 | 94%, 29%, 95%, 84%, 85% (50 mM glucose, xylose, galactose, mannose, arabinose) | 55% (5%, v/v) | 65 | 4.5 | 70 °C, 78%, 75 °C, 5% | [20] |
| Pichia membranifaciens | - | 50% (200 mM glucose) | 100% (20%, v/v) | 35 | 6.0 | 50 °C, 35%, 60 °C, 3% | [21] |
| Aspergillus niger CCRC31494 | - | 100% (30 mM galactose, mannose, xylose), Ki = 543 mM (glucose) | 68% (40%, v/v) | 55 | 5.0 | 60 °C, 90% | [22] |
| Aureobasidium pullulans CBS 135684 | - | 100% (70 mM glucose, arabinose, mannose), Ki = 18.2 mM (xylose) | 84% (30%, v/v) | 70 | 6.0 | 6 h, 70 °C, 50% | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhang, Z.; Yu, Z.; Chen, S.; Zhang, M.; Zhang, T.; Luo, X.; Zhao, J.; Li, Z. Simultaneous Improvement of Final Product-Tolerance and Thermostability of GH39 Xylosidase for Prebiotic Production by Directed Evolution. Foods 2022, 11, 3039. https://doi.org/10.3390/foods11193039
Zhang Z, Zhang Z, Yu Z, Chen S, Zhang M, Zhang T, Luo X, Zhao J, Li Z. Simultaneous Improvement of Final Product-Tolerance and Thermostability of GH39 Xylosidase for Prebiotic Production by Directed Evolution. Foods. 2022; 11(19):3039. https://doi.org/10.3390/foods11193039
Chicago/Turabian StyleZhang, Zirui, Zhengjie Zhang, Zhao Yu, Shiheng Chen, Mengwei Zhang, Tongcun Zhang, Xuegang Luo, Junqi Zhao, and Zhongyuan Li. 2022. "Simultaneous Improvement of Final Product-Tolerance and Thermostability of GH39 Xylosidase for Prebiotic Production by Directed Evolution" Foods 11, no. 19: 3039. https://doi.org/10.3390/foods11193039
APA StyleZhang, Z., Zhang, Z., Yu, Z., Chen, S., Zhang, M., Zhang, T., Luo, X., Zhao, J., & Li, Z. (2022). Simultaneous Improvement of Final Product-Tolerance and Thermostability of GH39 Xylosidase for Prebiotic Production by Directed Evolution. Foods, 11(19), 3039. https://doi.org/10.3390/foods11193039