

Improvement of Estrogen Deficiency Symptoms by the Intake of Long-Term Fermented Soybeans (Doenjang) Rich in Bacillus Species through Modulating Gut Microbiota in Estrogen-Deficient Rats
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. TMD Production Process and Sample Collection
2.2. Ovariectomy Procedure
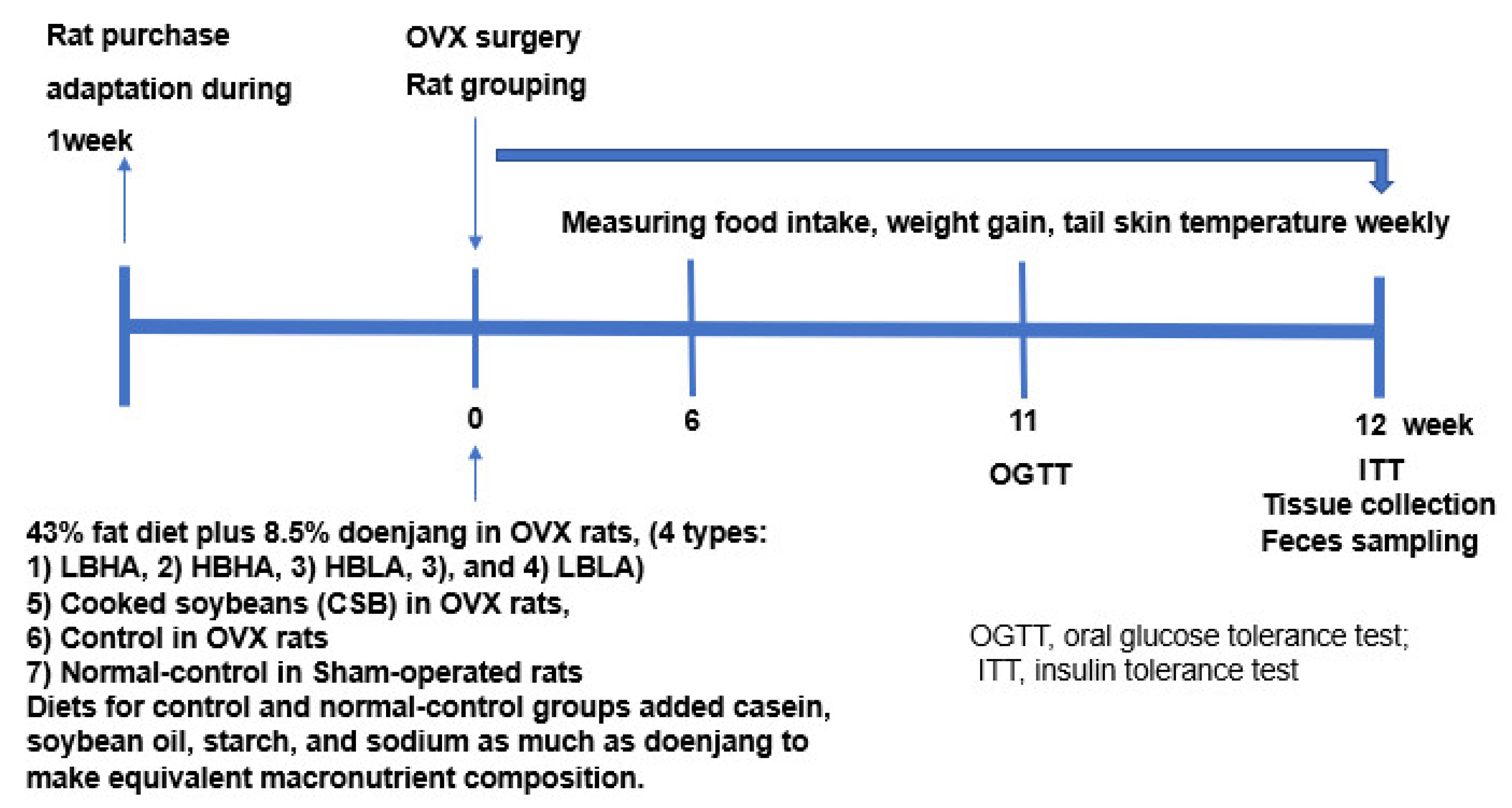
2.3. Experimental Design
2.4. Diet Preparation
2.5. Tail Skin Temperature
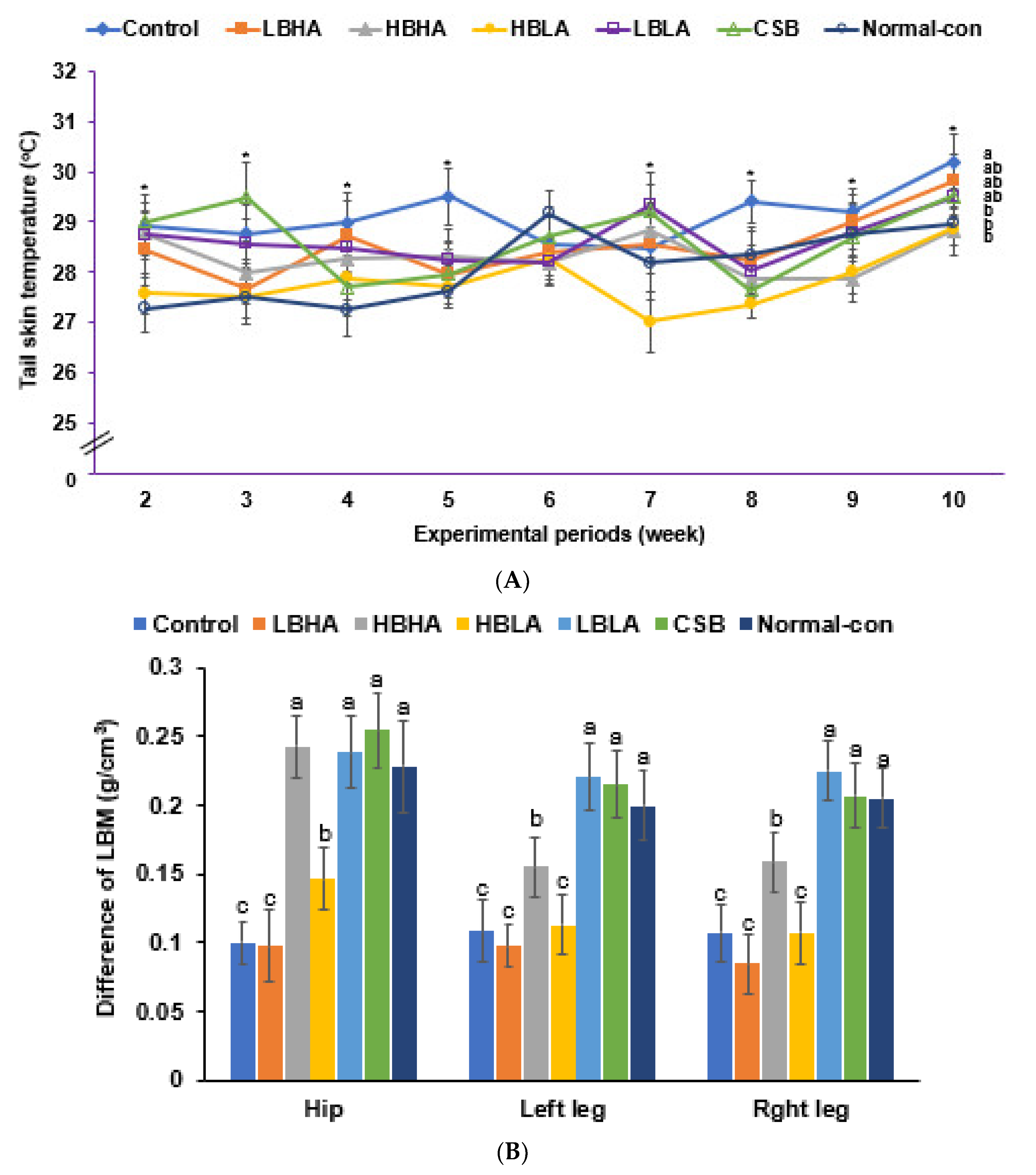
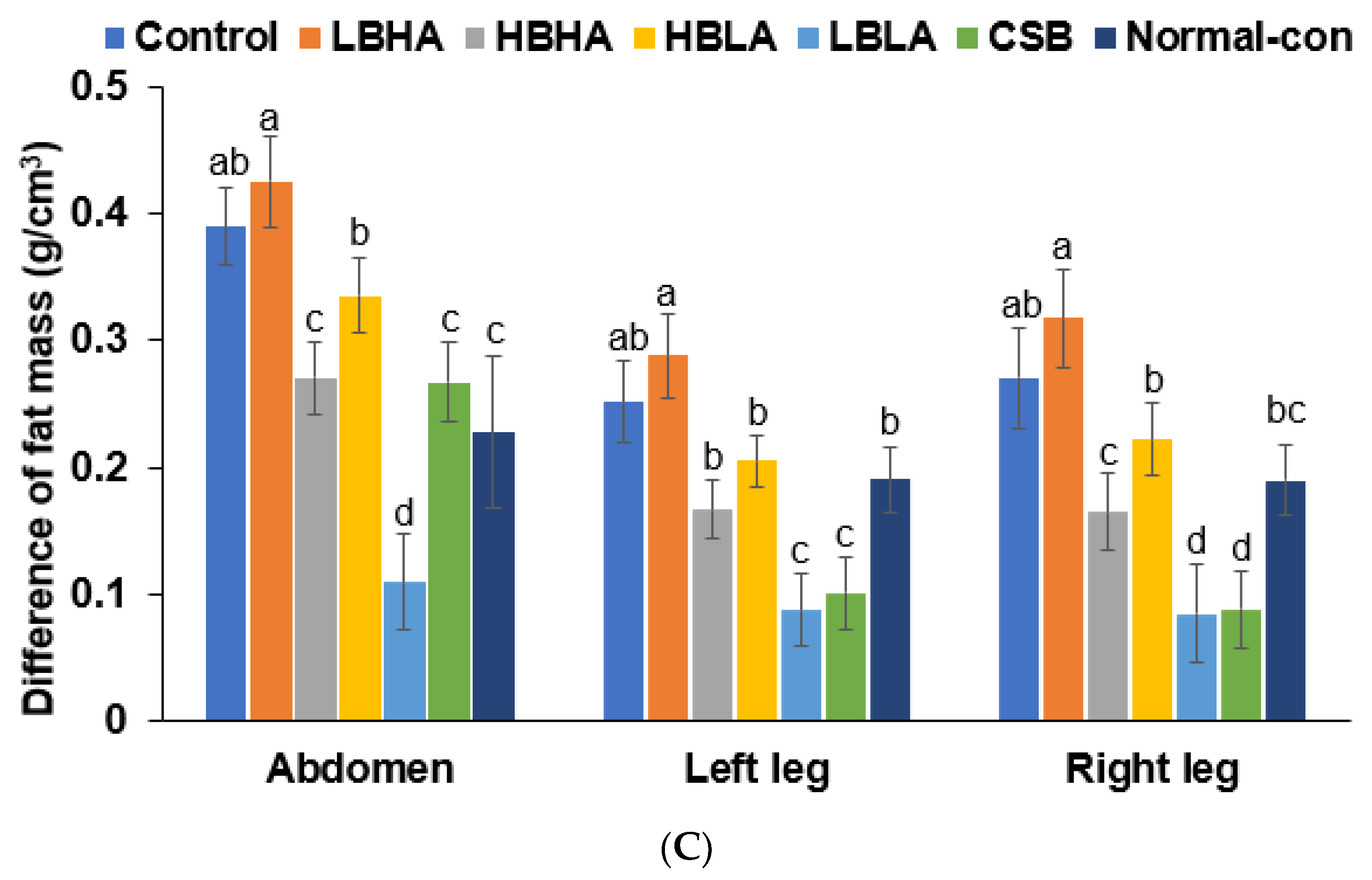
2.6. Fat and Skeletal Muscle Composition
2.7. Insulin Resistance and Lipid Profiles
2.8. Gene Expression by the Real-Time PCR Method
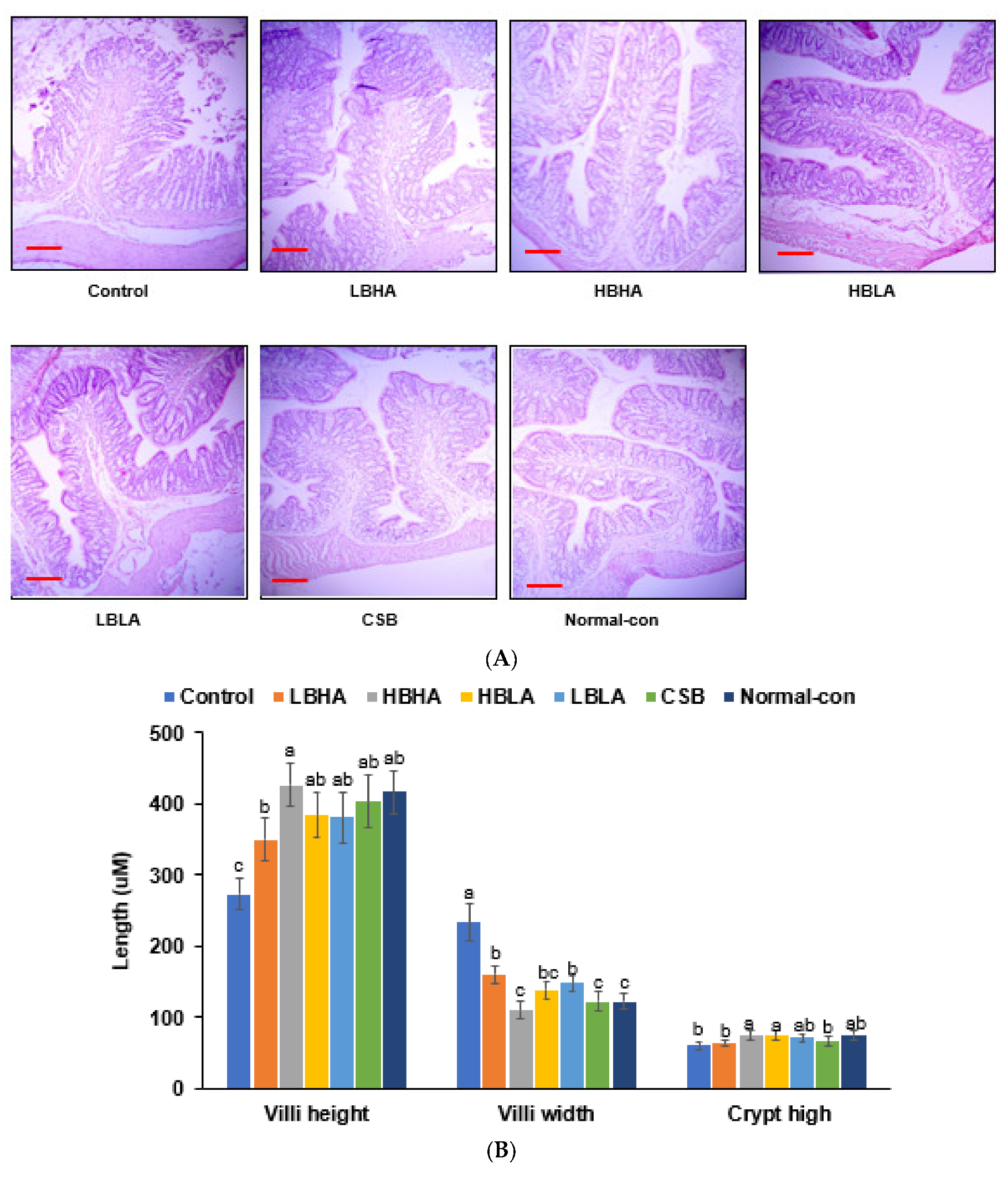
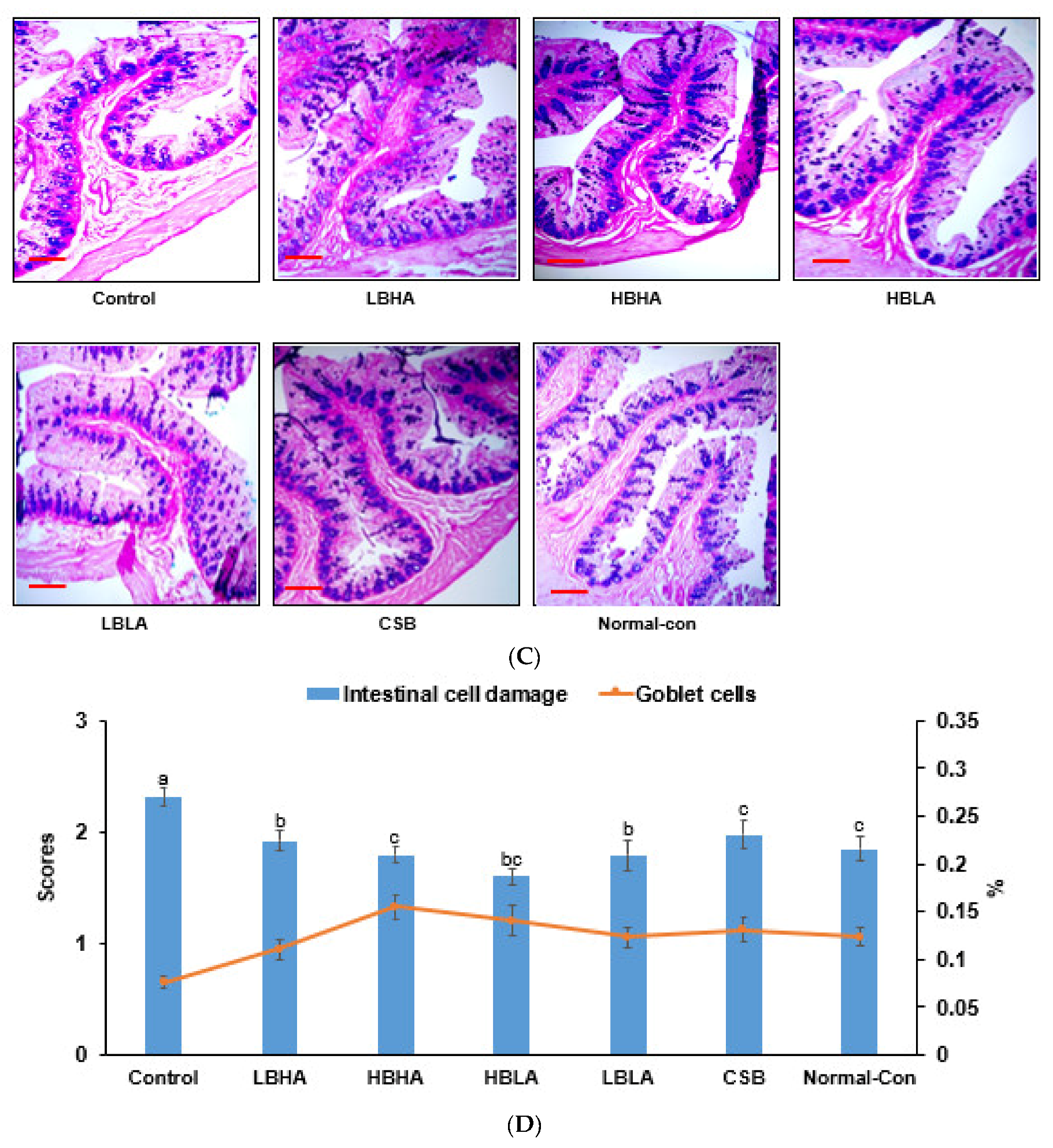
2.9. Histology of the Large Intestines
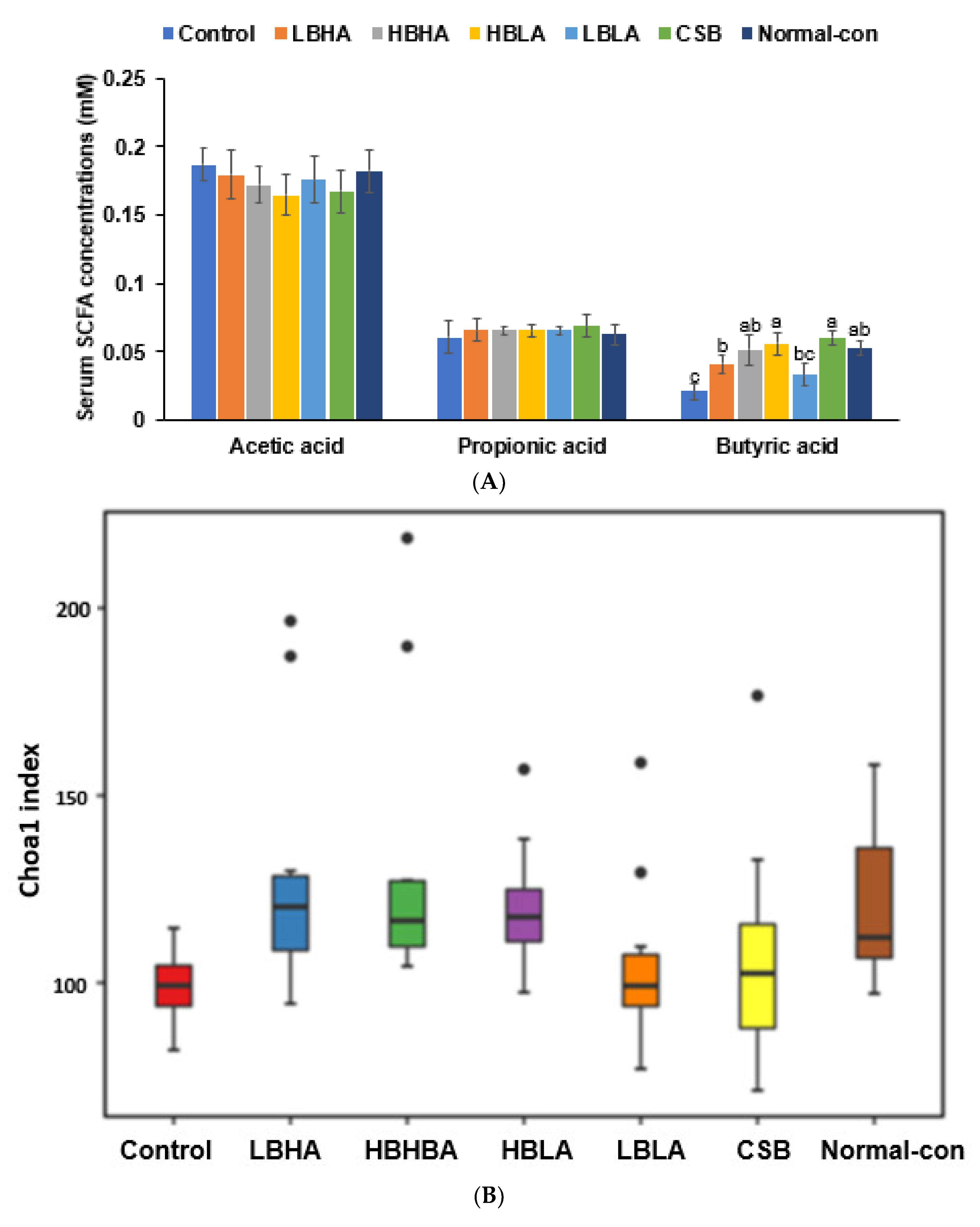
2.10. Serum Short-Chain Fatty Acids (SCFA) Concentrations and Gut Microbiome
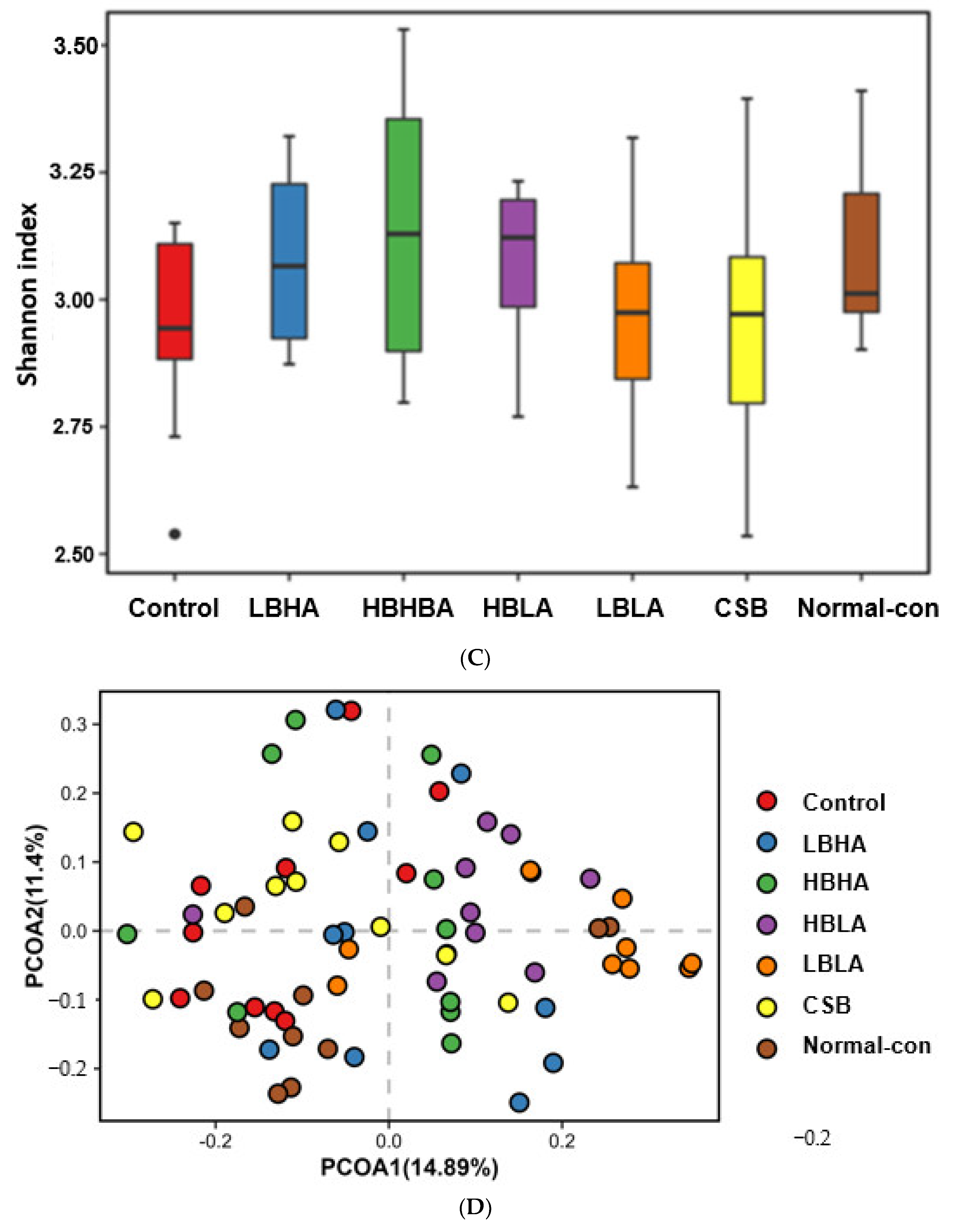
2.11. Metabolic Functions of the Gut Microbiomes by PICRUSt2 Pipeline Analysis
2.12. Statistical Analysis
3. Results
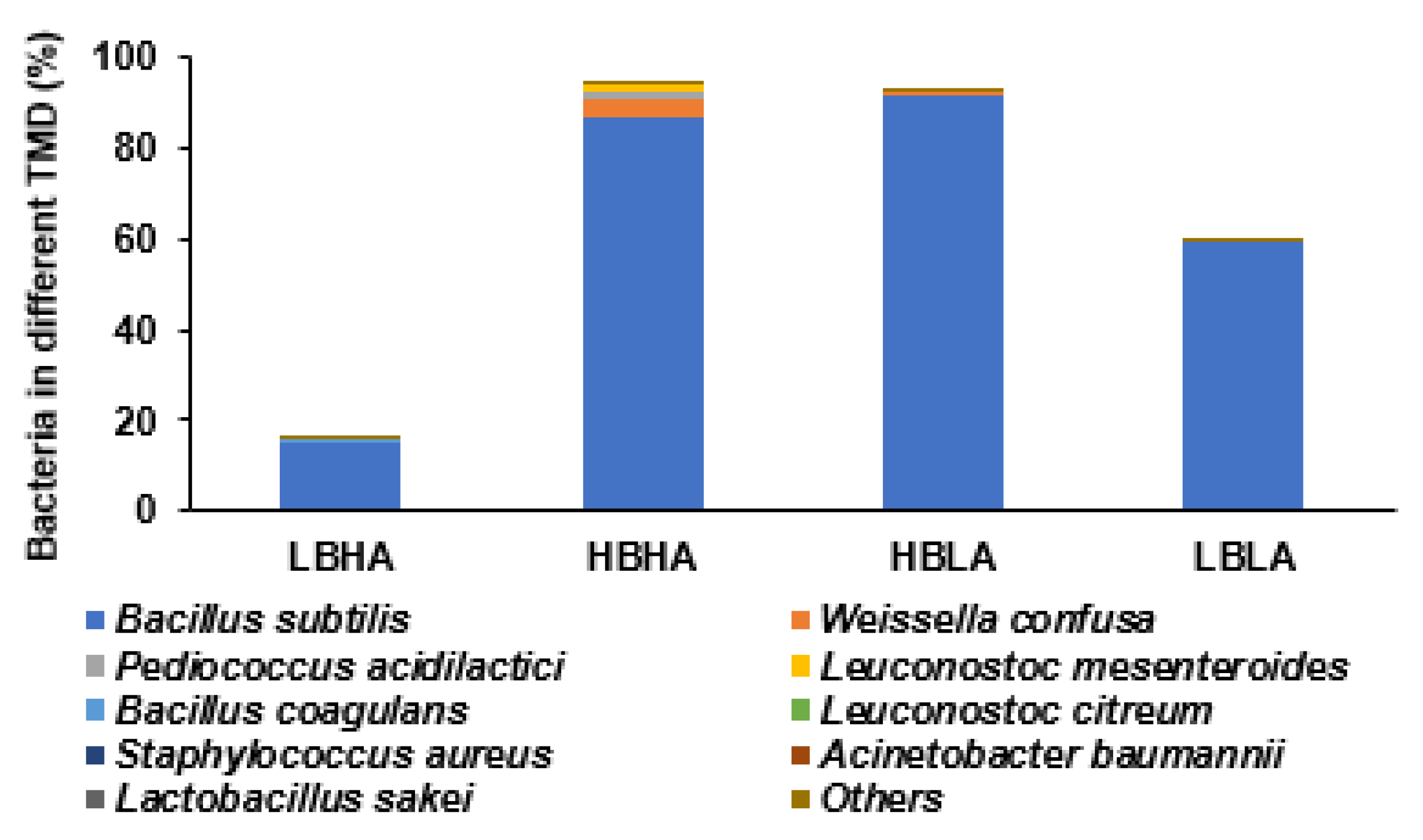
3.1. Characteristics of TMD according to Bacillus spp. and Biogenic Amine Concentrations
3.2. Isoflavonoid Contents
3.3. Uterine Weight, Serum 17β-Estradiol Levels, and Tail Skin Temperature
3.4. Insulin Resistance and Lipid Profiles
3.5. Lipid Metabolism in the Liver
3.6. Histology of the Large Intestines
3.7. SCFA in the Portal Vein and Gut Microbiota
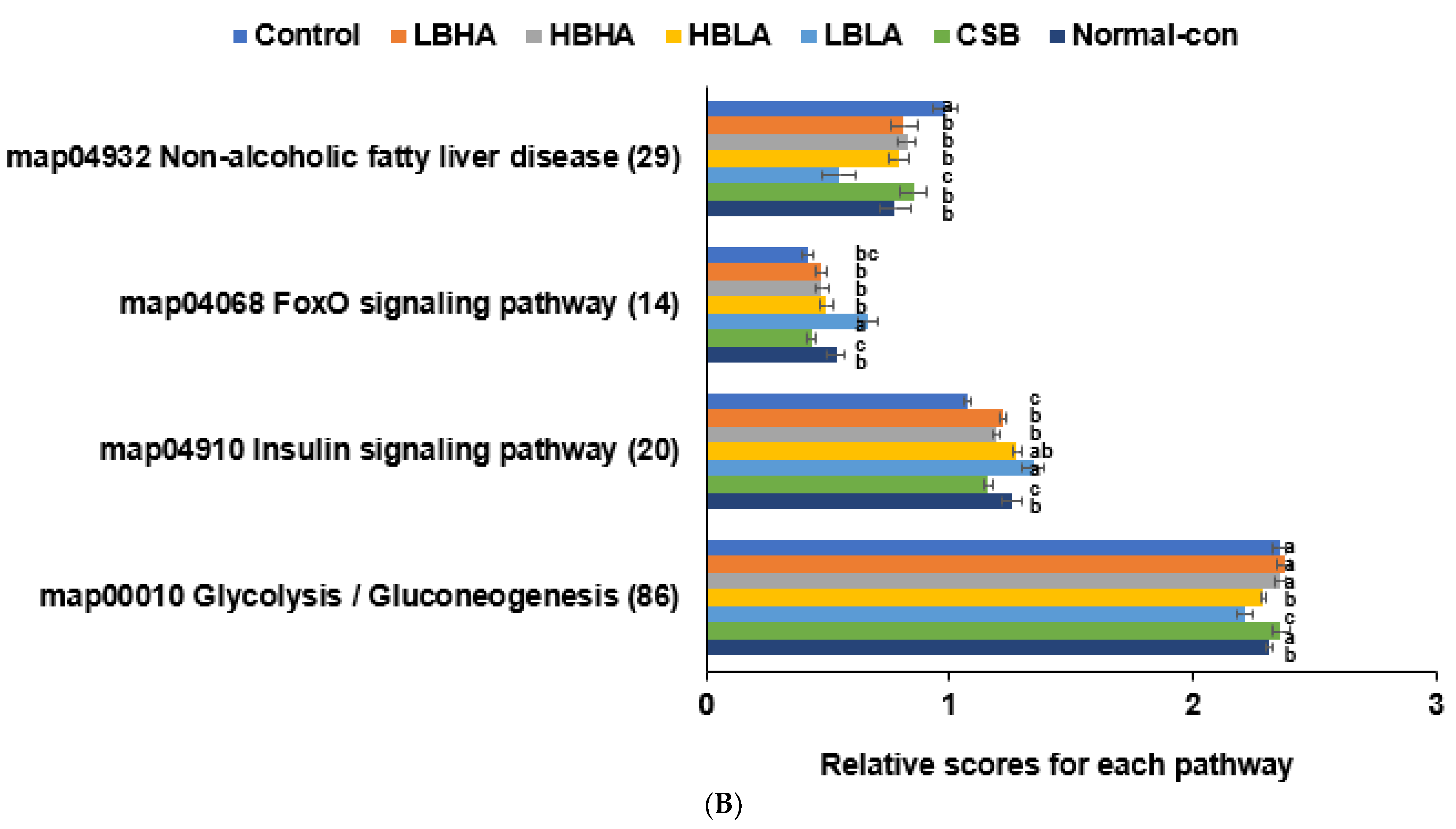
3.8. Metagenome Analysis of Fecal Bacteria
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kontis, V.; Bennett, J.E.; Mathers, C.D.; Li, G.; Foreman, K.; Ezzati, M. Future life expectancy in 35 industrialised countries: Projections with a Bayesian model ensemble. Lancet 2017, 389, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, R.A.; Gompel, A. Management of menopause: A view towards prevention. Lancet. Diabetes Endocrinol. 2022, 10, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Daily, J.W.; Ko, B.S.; Ryuk, J.; Liu, M.; Zhang, W.; Park, S. Equol Decreases Hot Flashes in Postmenopausal Women: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. J. Med. Food 2019, 22, 127–139. [Google Scholar] [CrossRef]
- Lobo, R.A. Hormone-replacement therapy: Current thinking. Nat. Rev. Endocrinol. 2017, 13, 220–231. [Google Scholar] [CrossRef]
- Kim, H.J.; Suh, H.J.; Kim, J.H.; Kang, S.C.; Park, S.; Lee, C.H.; Kim, J.S. Estrogenic activity of glyceollins isolated from soybean elicited with Aspergillus sojae. J. Med. Food 2010, 13, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Cross, T.L.; Zidon, T.M.; Welly, R.J.; Park, Y.M.; Britton, S.L.; Koch, L.G.; Rottinghaus, G.E.; de Godoy, M.R.C.; Padilla, J.; Swanson, K.S.; et al. Soy Improves Cardiometabolic Health and Cecal Microbiota in Female Low-Fit Rats. Sci. Rep. 2017, 7, 9261. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zheng, T.; Hui, H.; Xie, G. Soybean isoflavones modulate gut microbiota to benefit the health weight and metabolism. Front. Cell. Infect. Microbiol. 2022, 12, 1288. [Google Scholar] [CrossRef]
- Jeong, D.Y.; Jeong, S.Y.; Zhang, T.; Wu, X.; Qiu, J.Y.; Park, S. Chungkookjang, a soy food, fermented with Bacillus amyloliquefaciens protects gerbils against ishcmeic stroke injury, and post-stroke hyperglycemia. Food Res. Int. 2020, 128, 108769. [Google Scholar] [CrossRef]
- Heng, Y.; Kim, M.J.; Yang, H.J.; Kang, S.; Park, S. Lactobacillus intestinalis efficiently produces equol from daidzein and chungkookjang, short-term fermented soybeans. Arch. Microbiol. 2019, 201, 1009–1017. [Google Scholar] [CrossRef]
- Wuttke, M.; Li, Y.; Li, M.; Sieber, K.B.; Feitosa, M.F.; Gorski, M.; Tin, A.; Wang, L.; Chu, A.Y.; Hoppmann, A.; et al. A catalog of genetic loci associated with kidney function from analyses of a million individuals. Nat. Genet. 2019, 51, 957–972. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Zhang, T.; Yue, Y.; Jeong, S.J.; Ryu, M.S.; Wu, X.; Yang, H.J.; Jeong, D.Y. Alleviation of Metabolic Disturbance by Substituting Kanjang High in Bacillus for Salt through Modulation of Gut Microbiota in Estrogen-Deficient Rats. Foods 2022, 11, 1951. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Shin, M.; Jeong, S.; Yang, H.; Jeong, D. Characteristic analysis and production of short-ripened Korean traditional soy sauce added with rice bran. J. Korean Soc. Food Sci. Nutr. 2014, 43, 550–556. [Google Scholar] [CrossRef] [Green Version]
- Linares, D.M.; Ross, P.; Stanton, C. Beneficial Microbes: The pharmacy in the gut. Bioengineered 2016, 7, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.J.; Ryu, M.S.; Yang, H.J.; Wu, X.H.; Jeong, D.Y.; Park, S.M. Bacterial Distribution, Biogenic Amine Contents, and Functionalities of Traditionally Made Doenjang, a Long-Term Fermented Soybean Food, from Different Areas of Korea. Microorganisms 2021, 9, 1348. [Google Scholar] [CrossRef]
- Lee, Y.J.; Park, S.Y.; Lee, S.J.; Boo, Y.C.; Choi, J.Y.; Kim, J.E. Ucma, a direct transcriptional target of Runx2 and Osterix, promotes osteoblast differentiation and nodule formation. Osteoarthr. Cartil. 2015, 23, 1421–1431. [Google Scholar] [CrossRef] [Green Version]
- Litwak, S.A.; Wilson, J.L.; Chen, W.; Garcia-Rudaz, C.; Khaksari, M.; Cowley, M.A.; Enriori, P.J. Estradiol prevents fat accumulation and overcomes leptin resistance in female high-fat diet mice. Endocrinology 2014, 155, 4447–4460. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kang, S.; Jeong, D.Y.; Jeong, S.Y.; Park, J.J.; Yun, H.S. Cyanidin and malvidin in aqueous extracts of black carrots fermented with Aspergillus oryzae prevent the impairment of energy, lipid and glucose metabolism in estrogen-deficient rats by AMPK activation. Genes Nutr. 2015, 10, 455. [Google Scholar] [CrossRef]
- Yang, H.J.; Ko, B.S.; Kwon, D.Y.; Lee, H.W.; Kim, M.J.; Ryuk, J.; Kang, S.; Kim, D.S.; Park, S. Asian Elm tree inner bark prevents articular cartilage deterioration in ovariectomized obese rats with monoiodoacetate-induced osteoarthritis. Menopause 2016, 23, 197–208. [Google Scholar] [CrossRef]
- Park, S.; Kim, D.S.; Kang, S.; Moon, B.R. Fermented soybeans, Chungkookjang, prevent hippocampal cell death and beta-cell apoptosis by decreasing pro-inflammatory cytokines in gerbils with transient artery occlusion. Exp. Biol. Med. 2015, 241, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Hong, S.M.; Lee, J.E.; Sung, S.R. Exercise improves glucose homeostasis that has been impaired by a high-fat diet by potentiating pancreatic beta-cell function and mass through IRS2 in diabetic rats. J. Appl. Physiol. 2007, 103, 1764–1771. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Yuan, H.; Zhang, T.; Wu, X.; Huang, S.K.; Cho, S.M. Long-term silk peptide intake promotes skeletal muscle mass, reduces inflammation, and modulates gut microbiota in middle-aged female rats. Biomed. Pharmacother. 2021, 137, 111415. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Kim, M.J.; Yang, H.J.; Park, S. Chitosan alleviated menopausal symptoms and modulated the gut microbiota in estrogen-deficient rats. Eur. J. Nutr. 2020, 60, 1907–1919. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Zhang, T.; Wu, X.G.; Kim, M.J.; Kim, Y.H.; Yang, E.S.; Yoon, Y.S.; Park, S. Aqueous Blackcurrant Extract Improves Insulin Sensitivity and Secretion and Modulates the Gut Microbiome in Non-Obese Type 2 Diabetic Rats. Antioxidants 2021, 10, 756. [Google Scholar] [CrossRef]
- Mun, E.G.; Park, J.E.; Cha, Y.S. Effects of Doenjang, a Traditional Korean Soybean Paste, with High-Salt Diet on Blood Pressure in Sprague-Dawley Rats. Nutrients 2019, 11, 2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, Y.-S.; Yang, J.-A.; Back, H.-I.; Kim, S.-R.; Kim, M.-G.; Jung, S.-J.; Song, W.O.; Chae, S.-W. Visceral fat and body weight are reduced in overweight adults by the supplementation of Doenjang, a fermented soybean paste. Nutr. Res. Pract. 2012, 6, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Park, N.Y.; Rico, C.W.; Lee, S.C.; Kang, M.Y. Comparative effects of doenjang prepared from soybean and brown rice on the body weight and lipid metabolism in high fat-fed mice. J. Clin. Biochem. Nutr. 2012, 51, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Mea, H.J.; Yong, P.V.C.; Wong, E.H. An overview of Acinetobacter baumannii pathogenesis: Motility, adherence and biofilm formation. Microbiol. Res. 2021, 247, 126722. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, S.; Serry, F.; Abdellatif, H.; Abbas, H. Elimination of multidrug-resistant Proteus mirabilis biofilms using bacteriophages. Arch. Virol. 2019, 164, 2265–2275. [Google Scholar] [CrossRef]
- Healy, B.; Cooney, S.; O’Brien, S.; Iversen, C.; Whyte, P.; Nally, J.; Callanan, J.J.; Fanning, S. Cronobacter (Enterobacter sakazakii): An opportunistic foodborne pathogen. Foodborne Pathog. Dis. 2010, 7, 339–350. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Jeong, D.Y.; Kim, D.S.; Park, S. Chungkookjang with High Contents of Poly-γ-Glutamic Acid Improves Insulin Sensitizing Activity in Adipocytes and Neuronal Cells. Nutrients 2018, 10, 1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosravi, A.; Razavi, S.H. Therapeutic effects of polyphenols in fermented soybean and black soybean products. J. Funct. Foods 2021, 81, 104467. [Google Scholar] [CrossRef]
- Shukla, S.; Park, H.-K.; Kim, J.-K.; Kim, M. Determination of biogenic amines in Korean traditional fermented soybean paste (Doenjang). Food Chem. Toxicol. 2010, 48, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Kim, M.-J.; Moon, B. Various biogenic amines in Doenjang and changes in concentration depending on boiling and roasting. Appl. Biol. Chem. 2017, 60, 273–279. [Google Scholar] [CrossRef]
- Su-Yeon, K.; Hyeong-Eun, K.; Yong-Suk, K. The potentials of Bacillus licheniformis strains for inhibition of B. cereus growth and reduction of biogenic amines in cheonggukjang (Korean fermented unsalted soybean paste). Food Control 2017, 79, 87–93. [Google Scholar] [CrossRef]
- Eom, J.S.; Seo, B.Y.; Choi, H.S. Biogenic Amine Degradation by Bacillus Species Isolated from Traditional Fermented Soybean Food and Detection of Decarboxylase-Related Genes. J. Microbiol. Biotechnol. 2015, 25, 1519–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, R.R. Menopausal hot flashes: Mechanisms, endocrinology, treatment. J. Steroid Biochem. Mol. Biol. 2014, 142, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Bansal, R.; Aggarwal, N. Menopausal Hot Flashes: A Concise Review. J. Mid-Life Health 2019, 10, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.H.; Lee, C.; Kim, H.Y.; Lee, J.Y. What to do with troublesome hot flush? Korean Med. J. 2018, 33, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Koo, S.; Ahn, Y.; Lim, J.Y.; Cho, J.; Park, H.Y. Obesity associates with vasomotor symptoms in postmenopause but with physical symptoms in perimenopause: A cross-sectional study. BMC Women’s Health 2017, 17, 126. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kang, S.; Kim, D.S. Severe calcium deficiency increased visceral fat accumulation, down-regulating genes associated with fat oxidation, and increased insulin resistance while elevating serum parathyroid hormone in estrogen-deficient rats. Nutr. Res. 2020, 73, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Giustina, A.; Berardelli, R.; Gazzaruso, C.; Mazziotti, G. Insulin and GH-IGF-I axis: Endocrine pacer or endocrine disruptor? Acta Diabetol. 2015, 52, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Nagata, C.; Takatsuka, N.; Kawakami, N.; Shimizu, H. Soy Product Intake and Hot Flashes in Japanese Women: Results from a Community-based Prospective Study. Am. J. Epidemiol. 2001, 153, 790–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martiniakova, M.; Babikova, M.; Omelka, R. Pharmacological agents and natural compounds: Available treatments for osteoporosis. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2020, 71, 1–4. [Google Scholar] [CrossRef]
- Ko, S.H.; Jung, Y. Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women. Nutrients 2021, 13, 4556. [Google Scholar] [CrossRef]
- Hodson, L.; Harnden, K.; Banerjee, R.; Real, B.; Marinou, K.; Karpe, F.; Fielding, B.A. Lower resting and total energy expenditure in postmenopausal compared with premenopausal women matched for abdominal obesity. J. Nutr. Sci. 2014, 3, e3. [Google Scholar] [CrossRef] [Green Version]
- Geraci, A.; Calvani, R.; Ferri, E.; Marzetti, E.; Arosio, B.; Cesari, M. Sarcopenia and Menopause: The Role of Estradiol. Front. Endocrinol. 2021, 12, 682012. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Du, Y.; Oh, C.; No, J. Effects of Soy Foods in Postmenopausal Women: A Focus on Osteosarcopenia and Obesity. J. Obes. Metab. Syndr. 2020, 29, 180–187. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of Estrogens in the Regulation of Liver Lipid Metabolism. Adv. Exp. Med. Biol. 2017, 1043, 227–256. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Kim, S.; Kim, H.S. Hepatoprotective Effects of Soybean Embryo by Enhancing Adiponectin-Mediated AMP-Activated Protein Kinase α Pathway in High-Fat and High-Cholesterol Diet-Induced Nonalcoholic Fatty Liver Disease. J. Med. Food 2016, 19, 549–559. [Google Scholar] [CrossRef]
- Torre-Villalvazo, I.; Tovar, A.R.; Ramos-Barragán, V.E.; Cerbón-Cervantes, M.A.; Torres, N. Soy protein ameliorates metabolic abnormalities in liver and adipose tissue of rats fed a high fat diet. J. Nutr. 2008, 138, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Kwak, C.S.; Park, S.C.; Song, K.Y. Doenjang, a fermented soybean paste, decreased visceral fat accumulation and adipocyte size in rats fed with high fat diet more effectively than nonfermented soybeans. J. Med. Food 2012, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Santoro, N.; Kaplan, R.C.; Qi, Q. Spotlight on the Gut Microbiome in Menopause: Current Insights. Int. J. Women’s Health 2022, 14, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Krishnan, H.B.; Pham, Q.; Yu, L.L.; Wang, T.T. Soy and Gut Microbiota: Interaction and Implication for Human Health. J. Agric. Food Chem. 2016, 64, 8695–8709. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.E.; Kim, K.A.; Han, M.J.; Kim, D.H. Doenjang, a fermented Korean soybean paste, inhibits lipopolysaccharide production of gut microbiota in mice. J. Med. Food 2014, 17, 67–75. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | LBHA | HBHA | HBLA | LBLA |
|---|---|---|---|---|
| Total isoflavonoid aglycones (mg/kg) | 0.54 ± 0.06 b | 0.50 ± 0.02 b | 0.67 ± 0.00 a | 0.28 ± 0.05 c |
| Daidzein (mg/kg) | 0.21 ± 0.03 b | 0.22 ± 0.01 b | 0.26 ± 0.00 a | 0.09 ± 0.02 c |
| Genistein (mg/kg) | 0.28 ± 0.03 b | 0.25 ± 0.01 b | 0.36 ± 0.00 a | 0.12 ± 0.03 c |
| Glycitein (mg/kg) | 0.05 ± 0.01 a | 0.04 ± 0.00 a | 0.04 ± 0.00 a | 0.01 ± 0.00 b |
| Daidzin (mg/kg) | - | - | - | 0.02 ± 0.01 |
| Genistin (mg/kg) | - | - | - | 0.03 ± 0.01 |
| Glycitin (mg/kg) | - | - | - | - |
| Histamine (mg/kg) | 501 ± 0.78 b | 628 ± 0.34 a | 13.1 ± 0.10 d | 33.2 ± 0.20 c |
| Tyramine (mg/kg) | 1655 ± 0.81 a | 1088 ± 0.80 b | 20.8 ± 0.73 d | 155 ± 0.83 c |
| Sodium (wt %) | 5.14 ± 0.04 a | 4.37 ± 0.10 b | 4.87 ± 0.04 a | 4.52 ± 0.08 ab |
| Control | LBHA | HBHA | HBLA | LBLA | CSB | Normal Con | |
|---|---|---|---|---|---|---|---|
| Uterine weight (g) | 0.15 ± 0.02 c | 0.14 ± 0.02 c | 0.21 ± 0.03 b | 0.13 ± 0.02 c | 0.12 ± 0.02 c | 0.15 ± 0.03 c | 0.65 ± 0.03 a |
| Serum 17β-estradiol (pg/mL) | 1.44 ± 0.11 b | 1.38 ± 0.13 b | 1.59 ± 0.22 b | 1.37 ± 0.21 b | 1.35 ± 0.26 b | 1.49 ± 0.18 b | 6.36 ± 0.85 a |
| Final weight (g) | 351 ± 26.7 a | 324 ± 17.9 b | 330 ± 18.6 b | 321 ± 24.9 b | 319 ± 28.9 b | 326 ± 31.5 b | 277 ± 28.6 c |
| Weight gain (g) | 172 ± 12.7 a | 148 ± 12.8 b | 159 ± 10.3 b | 157 ± 16.5 b | 164 ± 17.9 ab | 164 ± 18.6 ab | 113 ± 21.9 c |
| Food intake (g/day) | 13.4 ± 1.7 | 11.8 ± 1.3 | 12.3 ± 0.7 | 12.3 ± 1.3 | 13.6 ± 1.3 | 12.7 ± 1.2 | 11.4 ± 1.3 |
| Food efficiency | 13.4 ± 1.6 a | 12.6 ± 1.3 a | 13.1 ± 0.7 a | 12.7 ± 1.3 a | 12.2 ± 1.1 a | 12.9 ± 1.4 a | 9.8 ± 1.4 b |
| Uterine fat (g) | 10.3 ± 0.92 a | 10.4 ± 1.14 a | 7.86 ± 0.97 b | 7.58 ± 0.87 b | 5.73 ± 0.9 c | 7.08 ± 0.87 b | 6.04 ± 0.97 c |
| Retroperitoneal fat (g) | 5.02 ± 0.63 a | 4.46 ± 0.53 a | 3.65 ± 0.37 b | 3.55 ± 0.44 b | 2.55 ± 0.37 c | 2.88 ± 0.32 c | 3.94 ± 0.98 b |
| Visceral fat (% of bw) | 4.72 ± 0.35 a | 4.98 ± 0.44 a | 4.10 ± 0.42 b | 4.09 ± 0.42 b | 3.03 ± 0.41 c | 3.76 ± 0.38 b | 3.93 ± 0.67 b |
| Control | LBHA | HBHA | HBLA | LBLA | CSB | Normal Con | |
|---|---|---|---|---|---|---|---|
| Fasting serum glucose (mg/dL) | 116 ± 4.15 a | 108 ± 6.81 ab | 106 ± 5.95 b | 105 ± 8.02 b | 111 ± 7.06 ab | 111 ± 8.68 ab | 98.2 ± 7.08 c |
| 2 h post-prandial serum glucose (mg/dL) | 148 ± 7.84 a | 138 ± 7.52 ab | 129 ± 7.1 b | 129 ± 9.53 b | 143 ± 7.91 a | 136 ± 6.38 ab | 129 ± 6.79 b |
| Fasting serum insulin (ng/mL) | 1.52 ± 0.17 a | 1.47 ± 0.17 a | 1.07 ± 0.11 b | 0.80 ± 0.17 c | 1.34 ± 0.17 ab | 0.89 ± 0.17 c | 0.89 ± 0.09 c |
| HOMA-IR | 6.27 ± 0.09 a | 6.39 ± 0.05 a | 4.26 ± 0.06 b | 3.26 ± 0.09 c | 4.85 ± 0.06 b | 3.05 ± 0.1 c | 3.94 ± 0.98 bc |
| Serum total cholesterol (mg/dL) | 209 ± 23.4 a | 183 ± 18.3 ab | 154 ± 14.0 b | 154 ± 11.3 b | 164 ± 17.0 b | 155 ± 23.3 b | 200 ± 21.9 a |
| Serum HDL (mg/dL) | 43.7 ± 3.71 c | 53.4 ± 4.95 b | 55.6 ± 4.08 ab | 61 ± 3.17 a | 47 ± 4.8 c | 54.1 ± 4.12 ab | 59.6 ± 1.92 a |
| Serum LDL (mg/dL) | 142 ± 14.6 a | 116 ± 10.3 b | 83.8 ± 7.89 d | 80.9 ± 5.36 d | 99.3 ± 10.3 c | 88.8 ± 11.8 d | 127 ± 9.92 b |
| Serum TG (mg/dL) | 115 ± 9.59 a | 71.8 ± 12.8 c | 74.4 ± 4.61 c | 57.9 ± 6.04 d | 87.8 ± 9.9 b | 57.9 ± 6.05 d | 66.6 ± 9.92 cd |
| Control | LBHA | HBHA | HBLA | LBLA | CSB | Normal Con | |
|---|---|---|---|---|---|---|---|
| Serum AST (IU/L) | 63.5 ± 4.16 a | 49.4 ± 3.71 c | 56.6 ± 2.61 b | 54.3 ± 3.57 b | 54.2 ± 3.22 b | 55.6 ± 2.48 b | 56.1 ± 2.27 b |
| Serum ALT (IU/L) | 32.8 ± 3.86 a | 28 ± 4.26 b | 16.1 ± 2.84 c | 18.3 ± 3.46 c | 29.6 ± 4.39 ab | 16.9 ± 2.44 c | 17.9 ± 1.63 c |
| Glycogen (mg/g tissue) | 24.4 ± 3.45 b | 29.1 ± 4.27 ab | 34.5 ± 3.75 a | 35.5 ± 2.79 a | 27.7 ± 3.63 b | 28.3 ± 3.94 b | 31.7 ± 4.05 a |
| TG (mg/g tissue) | 294 ± 11.8 a | 223 ± 15.2 c | 218 ± 12.9 c | 229 ± 18.6 c | 241 ± 11.2 b | 252 ± 16.4 b | 243 ± 12.1 b |
| Cholesterol (mg/g tissue) | 369 ± 10.7 a | 291 ± 13.0 c | 367 ± 18.0 a | 367 ± 16.8 a | 332 ± 13.5 b | 385 ± 9.60 a | 255 ± 14.5 d |
| PPAR-γ mRNA (AU) | 1 ± 0 c | 2.69 ± 0.47 b | 2.67 ± 0.43 b | 3.27 ± 0.33 a | 2.91 ± 0.29 b | 2.8 ± 0.42 b | 2.57 ± 0.29 b |
| SREBP-1c mRNA (AU) | 1 ± 0 a | 0.81 ± 0.06 b | 0.65 ± 0.11 bc | 0.47 ± 0.08 c | 0.94 ± 0.13 a | 0.41 ± 0.04 c | 0.42 ± 0.06 c |
| CPT-1 mRNA (AU) | 1 ± 0 c | 1.52 ± 0.16 b | 2.01 ± 0.19 a | 1.56 ± 0.17 b | 1.2 ± 0.13 c | 1.59 ± 0.19 b | 2.02 ± 0.31 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Yue, Y.; Jeong, S.-J.; Ryu, M.-S.; Wu, X.; Yang, H.-J.; Li, C.; Jeong, D.-Y.; Park, S. Improvement of Estrogen Deficiency Symptoms by the Intake of Long-Term Fermented Soybeans (Doenjang) Rich in Bacillus Species through Modulating Gut Microbiota in Estrogen-Deficient Rats. Foods 2023, 12, 1143. https://doi.org/10.3390/foods12061143
Zhang T, Yue Y, Jeong S-J, Ryu M-S, Wu X, Yang H-J, Li C, Jeong D-Y, Park S. Improvement of Estrogen Deficiency Symptoms by the Intake of Long-Term Fermented Soybeans (Doenjang) Rich in Bacillus Species through Modulating Gut Microbiota in Estrogen-Deficient Rats. Foods. 2023; 12(6):1143. https://doi.org/10.3390/foods12061143
Chicago/Turabian StyleZhang, Ting, Yu Yue, Su-Ji Jeong, Myeong-Seon Ryu, Xuangao Wu, Hee-Jong Yang, Chen Li, Do-Youn Jeong, and Sunmin Park. 2023. "Improvement of Estrogen Deficiency Symptoms by the Intake of Long-Term Fermented Soybeans (Doenjang) Rich in Bacillus Species through Modulating Gut Microbiota in Estrogen-Deficient Rats" Foods 12, no. 6: 1143. https://doi.org/10.3390/foods12061143
APA StyleZhang, T., Yue, Y., Jeong, S.-J., Ryu, M.-S., Wu, X., Yang, H.-J., Li, C., Jeong, D.-Y., & Park, S. (2023). Improvement of Estrogen Deficiency Symptoms by the Intake of Long-Term Fermented Soybeans (Doenjang) Rich in Bacillus Species through Modulating Gut Microbiota in Estrogen-Deficient Rats. Foods, 12(6), 1143. https://doi.org/10.3390/foods12061143