
Anti-Biofilm Effects of Z102-E of Lactiplantibacillus plantarum against Listeria monocytogenes and the Mechanism Revealed by Transcriptomic Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preliminary Preparation
2.2. Analysis of Growth Curve and MIC
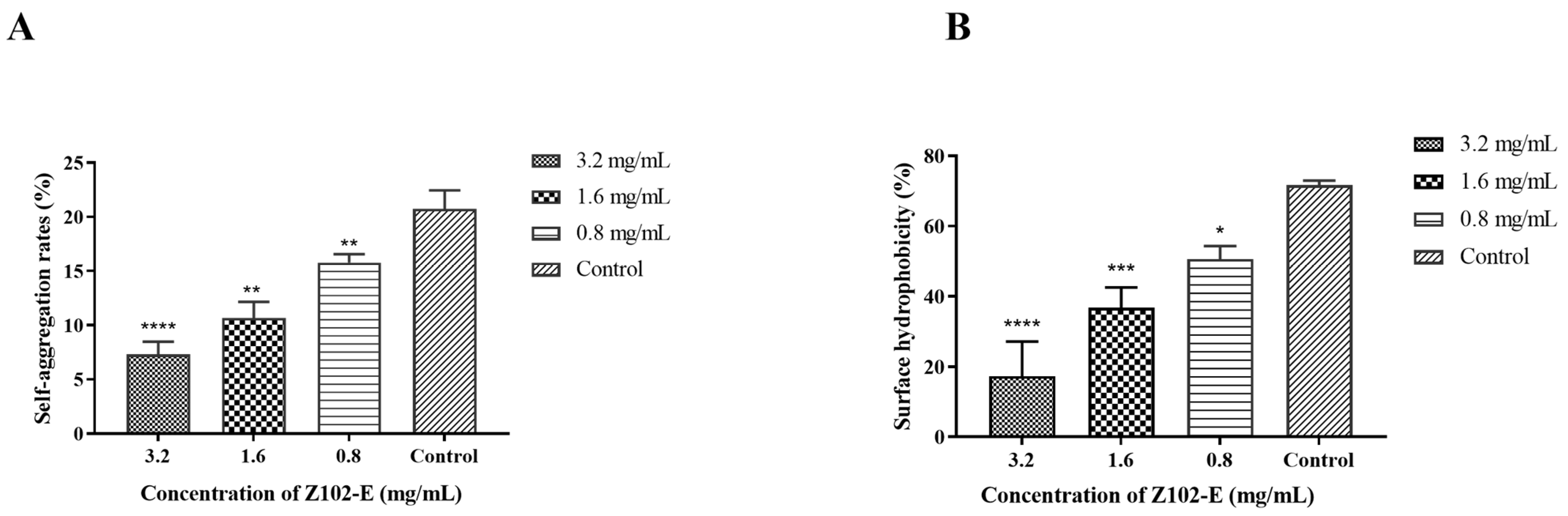
2.3. Effects of Z102-E on Auto-Aggregation and Surface Hydrophobicity of L. monocytogenes
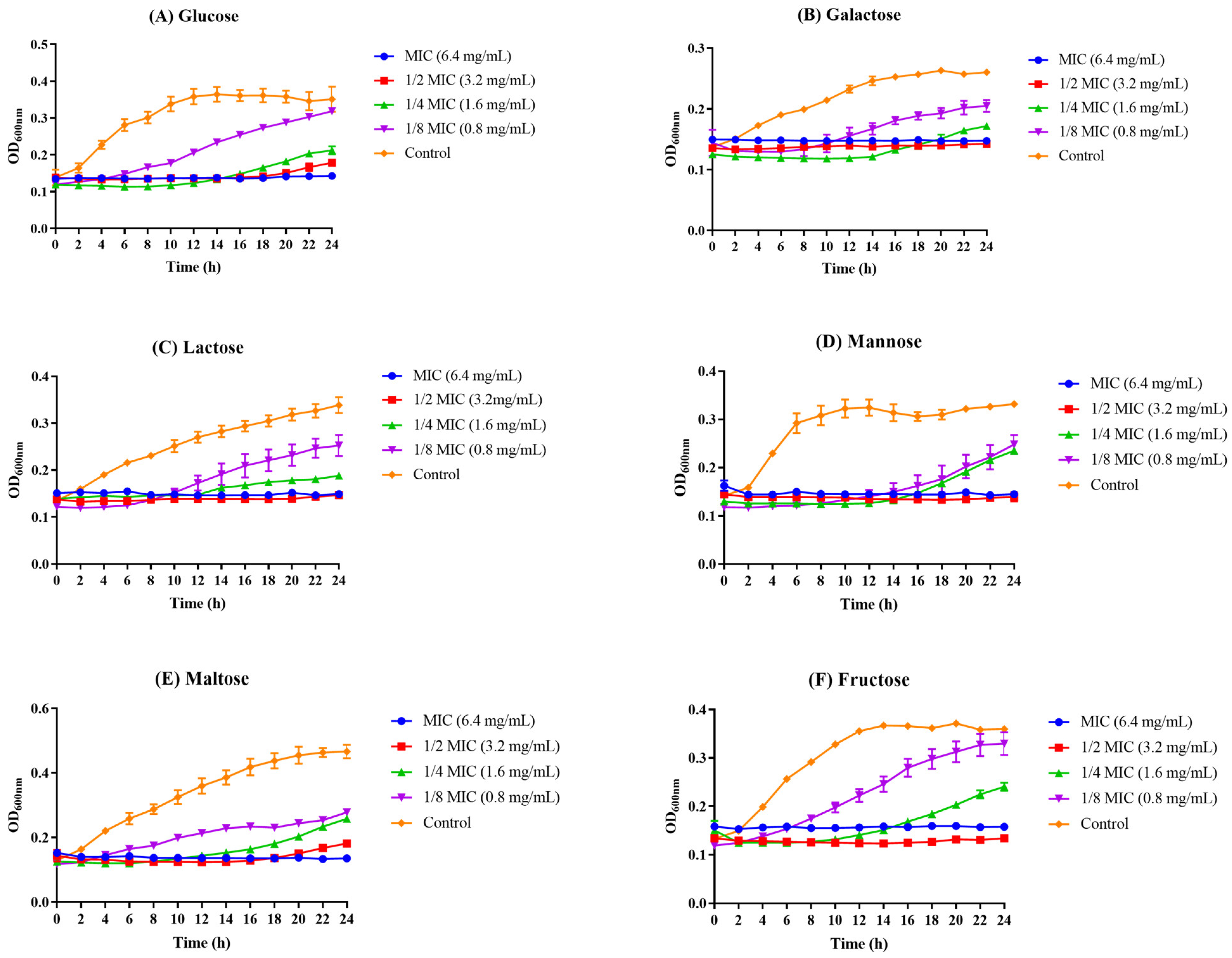
2.4. The Effect of Z102-E on Carbohydrate Utilization of L. monocytogenes
2.5. Motility Assay
2.6. Effect of Z102-E on AI-2 Signal Molecules of L. monocytogenes
2.7. Effect of Z102-E on Biofilm Formation of L. monocytogenes
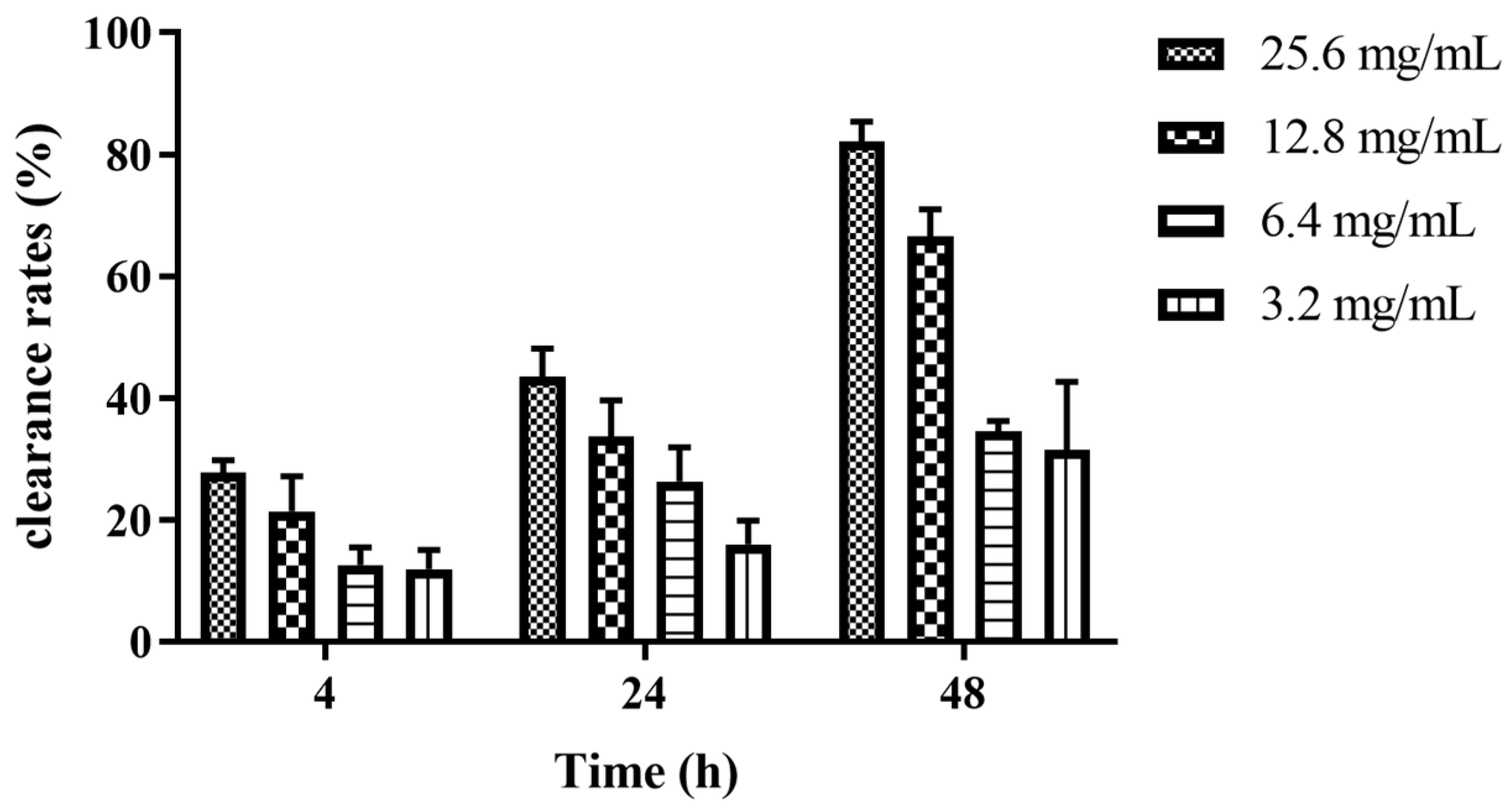
2.8. Removal Effects of Z102-E on the Mature Biofilm of L. monocytogenes
2.9. Quantification of Extracellular Polysaccharides in Biofilm
2.10. Effect of Z102-E on Extracellular Proteins in L. monocytogenes Biofilm
2.11. Observation of Biofilm by Inverted Fluorescence Microscopy
2.12. Transcriptomic Analysis
2.13. Data Analysis
3. Results
3.1. The Antibacterial Effect of Z102-E against L. monocytogenes
3.2. Effects of Z102-E on the Self-Aggregation and Surface Hydrophobicity of L. monocytogenes
3.3. The Influence of Z102-E on Carbohydrate Utilization of L. monocytogenes
3.4. The Inhibition on Motility
3.5. Influence of Z102-E on Biofilm Formation of L. monocytogenes
3.6. Elimination Effects of Z102-E on the Mature Biofilm of L. monocytogenes
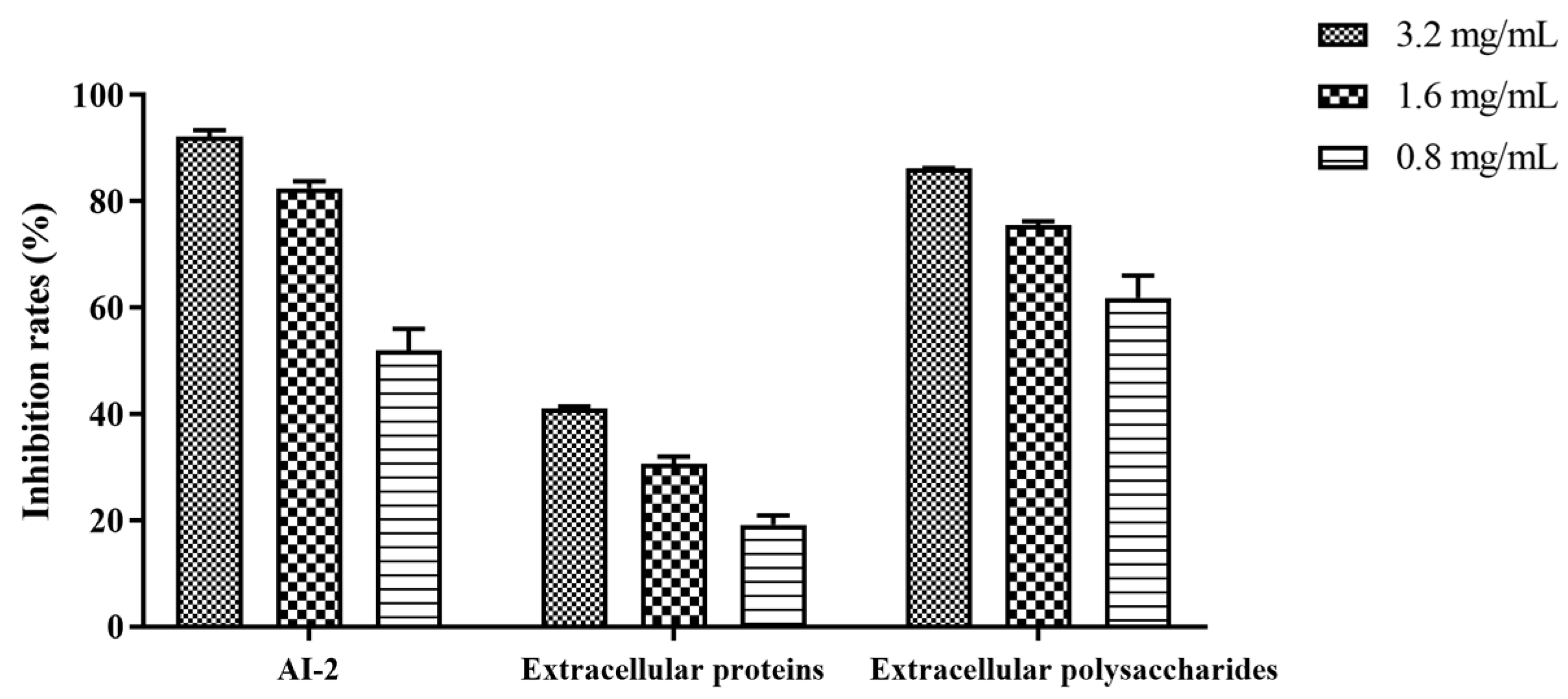
3.7. Effect of Z102-E on AI-2 Signal Molecule, Extracellular Proteins, and Extracellular Polysaccharides of L. monocytogenes Biofilm
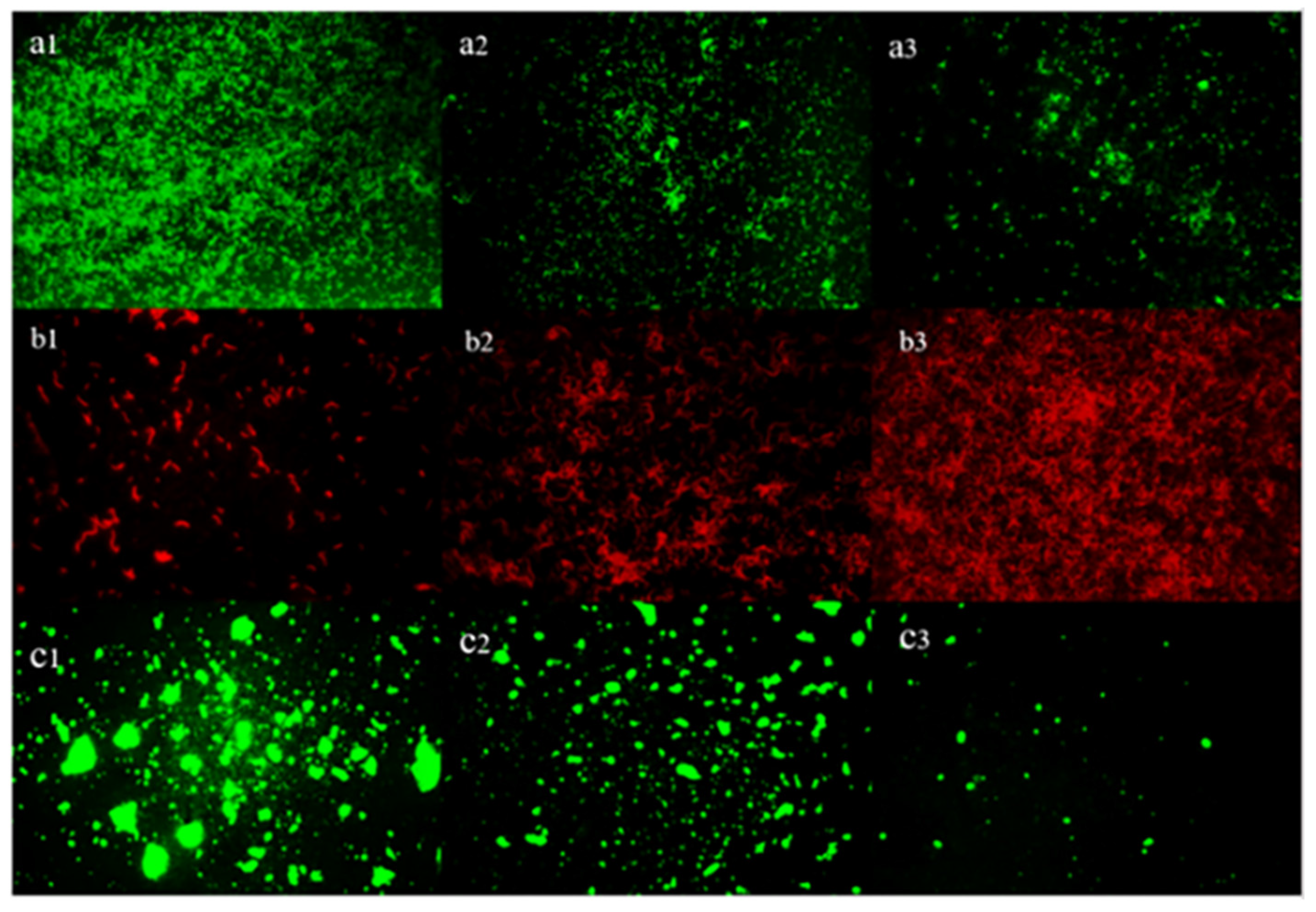
3.8. Inverted Fluorescence Microscopy Observation of the Cells and EPS in Biofilm
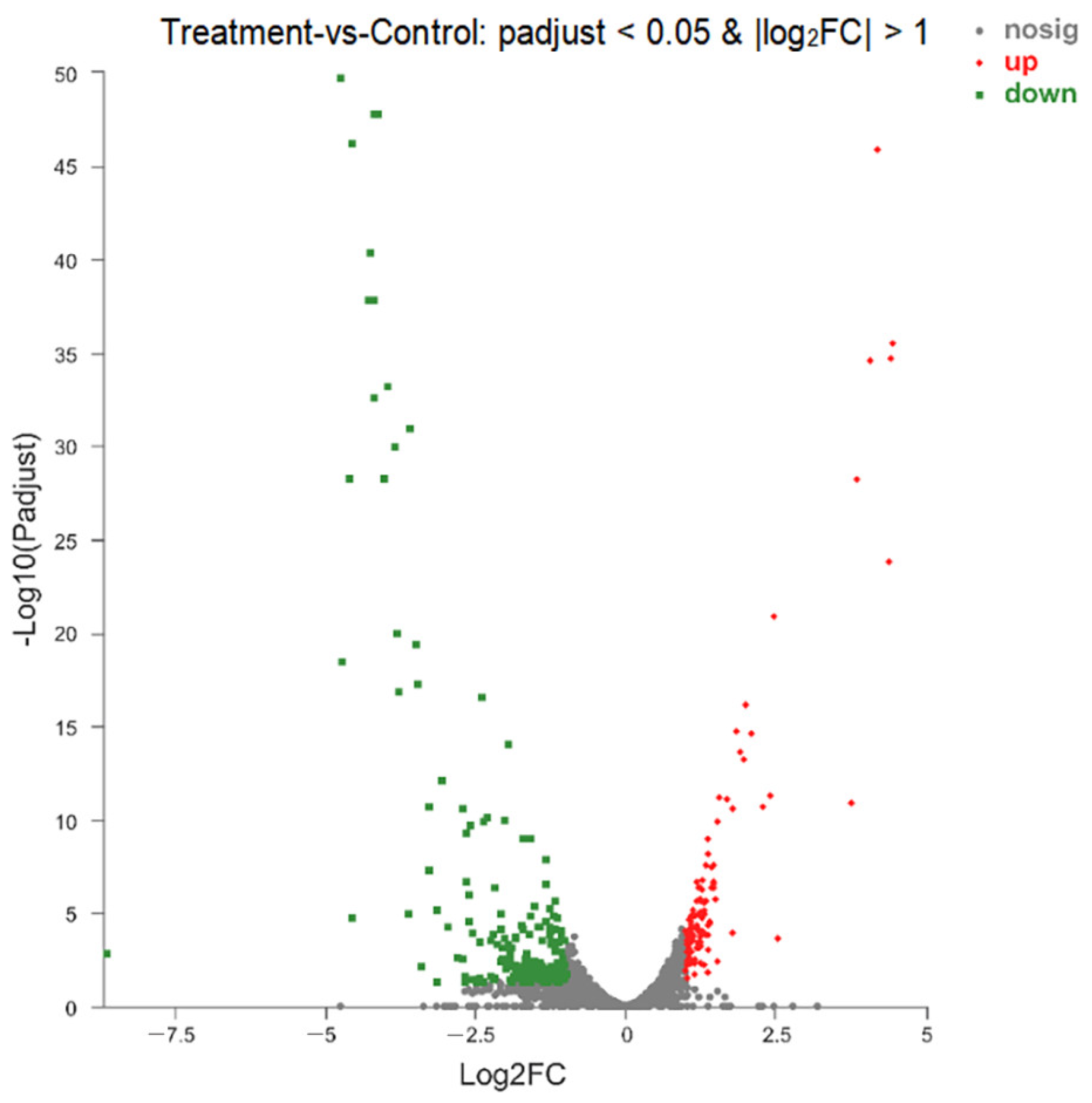
3.9. Analysis of DEGs
3.10. Cluster Analysis
3.11. Gene Ontology (GO) Functional Analysis
3.12. GO Enrichment Analysis
3.13. KEGG Function Analysis
3.14. KEGG Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, M.L.; Ricke, S.C.; Donaldson, J.R. Establishment of Listeria monocytogenes in the gastrointestinal tract. Microorganisms 2019, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Auad, J.P. Overview of Listeria monocytogenes infection. Rev. Chil. Infectol. 2018, 35, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Clementi, F. The occurrence of Listeria monocytogenes in mass catering: An overview in the European Union. Int. J. Hosp. Manag. 2016, 57, 9–17. [Google Scholar] [CrossRef]
- Pennone, V.; Barron, U.G.; Hunt, K.; Cadavez, V.; McAuliffe, O.; Butler, F. Omnibus modeling of Listeria monocytogenes growth rates at low temperatures. Foods 2021, 10, 1099. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M.M.; Brouwer, M.C.; Vazquez-Boland, J.A.; van de Beek, D. Human listeriosis. Clin. Microbiol. Rev. 2023, 36. [Google Scholar] [CrossRef] [PubMed]
- Allerberger, F.; Wagner, M. Listeriosis: A resurgent foodborne infection. Clin. Microbiol. Infect. 2010, 16, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in fresh produce: Outbreaks, prevalence and contamination levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Jordan, K.; McAuliffe, O. Listeria monocytogenes in Foods. Adv. Food Nutr. Res. 2018, 86, 181–213. [Google Scholar] [CrossRef]
- Poulsen, K.P.; Czuprynski, C.J. Pathogenesis of listeriosis during pregnancy. Anim. Health Res. Rev. 2013, 14, 30–39. [Google Scholar] [CrossRef]
- Craig, A.M.; Dotters-Katz, S.; Kuller, J.A.; Thompson, J.L. Listeriosis in pregnancy: A review. Obstet. Gynecol. Surv. 2019, 74, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Halbedel, S.; Wilking, H.; Holzer, A.; Kleta, S.; Fischer, M.A.; Luth, S.; Pietzka, A.; Huhulescu, S.; Lachmann, R.; Krings, A.; et al. Large nationwide outbreak of invasive listeriosis associated with blood sausage, Germany, 2018-2019. Emerg. Infect. Dis. 2020, 26, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Tau, N.P.; Smouse, S.L.; Allam, M.; Ismail, A.; Ramalwa, N.R.; Disenyeng, B.; Ngomane, M.; Thomas, J. Outbreak of Listeria monocytogenes in South Africa, 2017-2018: Laboratory activities and experiences associated with whole-genome sequencing analysis of isolates. Foodborne Pathog. Dis. 2019, 16, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, F.; Rezaee, M.A.; Ebrahimzadeh, S.; Yousefi, L.; Nouri, R.; Kafil, H.S.; Gholizadeh, P. Novel strategies to combat bacterial biofilms. Mol. Biotechnol. 2021, 63, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yao, H.; Zhao, X.; Ge, C. Biofilm formation and control of foodborne pathogenic bacteria. Molecules 2023, 28, 2432. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Claessen, D.; Rozen, D.E.; Kuipers, O.P.; Sogaard-Andersen, L.; van Wezel, G.P. Bacterial solutions to multicellularity: A tale of biofilms, filaments and fruiting bodies. Nat. Rev. Microbiol. 2014, 12, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Galie, S.; Garcia-Gutierrez, C.; Miguelez, E.M.; Villar, C.J.; Lombo, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Sharan, M.; Vijay, D.; Dhaka, P.; Bedi, J.S.; Gill, J.P.S. Biofilms as a microbial hazard in the food industry: A scoping review. J. Appl. Microbiol. 2022, 133, 2210–2234. [Google Scholar] [CrossRef]
- Gemmell, C.T.; Parreira, V.R.; Farber, J.M. Controlling Listeria monocytogenes growth and biofilm formation using flavonoids. J. Food Prot. 2022, 85, 639–646. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Chen, X.; Qu, C. Review controlling Listeria monocytogenes in ready-to-eat meat and poultry products: An overview of outbreaks, current legislations, challenges, and future prospects. Trends Food Sci. Technol. 2021, 116, 24–35. [Google Scholar] [CrossRef]
- Lemon, K.P.; Higgins, D.E.; Kolter, R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J. Bacteriol. 2007, 189, 4418–4424. [Google Scholar] [CrossRef]
- Guttenplan, S.B.; Kearns, D.B. Regulation of flagellar motility during biofilm formation. FEMS Microbiol. Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef] [PubMed]
- Colagiorgi, A.; Di Ciccio, P.; Zanardi, E.; Ghidini, S.; Ianieri, A. A look inside the Listeria monocytogenes biofilms extracellular matrix. Microorganisms 2016, 4, 22. [Google Scholar] [CrossRef]
- Low, K.E.; Howell, P.L. Gram-negative synthase-dependent exopolysaccharide biosynthetic machines. Curr. Opin. Struct. Biol. 2018, 53, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Steiger, E.L.; Muelli, J.R.; Braissant, O.; Waltimo, T.; Astasov-Frauenhoffer, M. Effect of divalent ions on cariogenic biofilm formation. BMC Microbiol. 2020, 20, 287. [Google Scholar] [CrossRef]
- Devaraj, A.; Novotny, L.A.; Robledo-Avila, F.H.; Buzzo, J.R.; Mashburn-Warren, L.; Jurcisek, J.A.; Tjokro, N.O.; Partida-Sanchez, S.; Bakaletz, L.O.; Goodman, S.D. The extracellular innate-immune effector HMGB1 limits pathogenic bacterial biofilm proliferation. J. Clin. Investig. 2021, 131, e140527. [Google Scholar] [CrossRef]
- Valderrama, W.B.; Cutter, C.N. An ecological perspective of Listeria monocytogenes biofilms in food processing facilities. Crit. Rev. Food Sci. Nutr. 2013, 53, 801–817. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Racimo, F.; Schluter, J.; Levy, S.B.; Foster, K.R. Importance of positioning for microbial evolution. Proc. Natl. Acad. Sci. USA 2014, 111, E1639–E1647. [Google Scholar] [CrossRef]
- More, T.T.; Yadav, J.S.S.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Extracellular polymeric substances of bacteria and their potential environmental applications. J. Environ. Manag. 2014, 144, 1–25. [Google Scholar] [CrossRef]
- Perez-Velazquez, J.; Golgeli, M.; Garcia-Contreras, R. Mathematical modelling of bacterial quorum sensing: A review. Bull. Math. Biol. 2016, 78, 1585–1639. [Google Scholar] [CrossRef] [PubMed]
- Remy, B.; Mion, S.; Plener, L.; Elias, M.; Chabriere, E.; Daude, D. Interference in bacterial quorum sensing: A biopharmaceutical perspective. Front. Pharmacol. 2018, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Lyu, F.; Liu, S.; Zhang, J.; Ding, Y.; Chen, W.; Zhou, X. Involvement of bacterial quorum sensing signals in spoilage potential of Aeromonas veronii bv. veronii isolated from fermented surimi. J. Food Biochem. 2018, 42, e12487. [Google Scholar] [CrossRef]
- Liu, S.Y.; Lu, H.Y.; Zhang, S.L.; Shi, Y.; Chen, Q.H. Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review. Pharmaceutics 2022, 14, 427. [Google Scholar] [CrossRef] [PubMed]
- Sharahi, J.Y.; Azimi, T.; Shariati, A.; Safari, H.; Tehrani, M.K.; Hashemi, A. Advanced strategies for combating bacterial biofilms. J. Cell. Physiol. 2019, 234, 14689–14708. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.C.; Yu, Y.Y.; Jin, X.; Qiu, J.Z.; Guan, X.; Huang, T.P. Beyond risk: Bacterial biofilms and their regulating approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.L.; Yu, M.; Shan, H.; Tian, X.R.; Zheng, Y.F.; Xue, C.X.; Zhang, X.H. Characterization of a novel n-acylhomoserine lactonase rmml from Ruegeria mobilis YJ3. Mar. Drugs 2018, 16, 370. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.Y.; Zuo, J.; Wang, H.K.; Grenier, D.; Yi, L.; Wang, Y. Contribution of quorum sensing to virulence and antibiotic resistance in zoonotic bacteria. Biotechnol. Adv. 2022, 59, 107965. [Google Scholar] [CrossRef] [PubMed]
- Romling, U.; Balsalobre, C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J. Intern. Med. 2012, 272, 541–561. [Google Scholar] [CrossRef]
- de la Fuente-Nunez, C.; Reffuveille, F.; Fernandez, L.; Hancock, R.E.W. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef]
- van Asselt, E.D.; Arrizabalaga-Larranaga, A.; Focker, M.; Berendsen, B.J.A.; van de Schans, M.G.M.; Van der Fels-Klerx, H.J. Chemical food safety hazards in circular food systems: A review. Crit. Rev. Food Sci. Nutr. 2022, 63, 10319–10331. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Diab, R.; Ghazvini, K.; Bazzaz, B.S.F. Breakthroughs in bacterial resistance mechanisms and the potential ways to combat them. Microb. Pathog. 2016, 95, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control biofilm-forming pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef] [PubMed]
- Lamin, A.; Kaksonen, A.H.; Cole, I.S.; Chen, X.B. Quorum sensing inhibitors applications: A new prospect for mitigation of microbiologically influenced corrosion. Bioelectrochemistry 2022, 145, 108050. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Chiou, J.; Chatterjee, P.; Otto, M. Alternative approaches to treat bacterial infections: Targeting quorum-sensing. Expert Rev. Anti-Infect. Ther. 2020, 18, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Zhu, J.; Gong, S.; Liu, H.; Yu, H. Antimicrobial characteristics of lactic acid bacteria isolated from homemade fermented foods. BioMed Res. Int. 2018, 2018, 5416725. [Google Scholar] [CrossRef] [PubMed]
- Webb, L.; Ma, L.Y.; Lu, X.N. Impact of lactic acid bacteria on the control of Listeria monocytogenes in ready-to-eat foods. Food Qual. Saf. 2022, 6, fyac045. [Google Scholar] [CrossRef]
- Hossain, M.I.; Kim, K.; Mizan, M.F.R.; Toushik, S.H.; Ashrafudoulla, M.; Roy, P.K.; Nahar, S.; Jahid, I.K.; Choi, C.; Park, S.H.; et al. Comprehensive molecular, probiotic, and quorum-sensing characterization of anti-listerial lactic acid bacteria, and application as bioprotective in a food (milk) model. J. Dairy Sci. 2021, 104, 6516–6534. [Google Scholar] [CrossRef]
- Aman, M.; Aneeqha, N.; Bristi, K.; Deeksha, J.; Afza, N.; Sindhuja, V.; Shastry, R.P. Lactic acid bacteria inhibits quorum sensing and biofilm formation of Pseudomonas aeruginosa strain JUPG01 isolated from rancid butter. Biocatal. Agric. Biotechnol. 2021, 36, 102115. [Google Scholar] [CrossRef]
- Wei, Y.H.; Wang, J.Z.; Liu, Z.; Pei, J.J.; Brennan, C.; Abd El-Aty, A.M. isolation and characterization of bacteriocin-producing Lacticaseibacillus rhamnosus XN2 from yak yoghurt and its bacteriocin. Molecules 2022, 27, 2066. [Google Scholar] [CrossRef]
- Melian, C.; Segli, F.; Gonzalez, R.; Vignolo, G.; Castellano, P. Lactocin AL705 as quorum sensing inhibitor to control Listeria monocytogenes biofilm formation. J. Appl. Microbiol. 2019, 127, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Gu, S.; Cui, X.; Shi, Y.; Wen, S.; Chen, H.; Ge, J. Antimicrobial, anti-adhesive and anti-biofilm potential of biosurfactants isolated from Pediococcus acidilactici and Lactobacillus plantarum against Staphylococcus aureus CMCC26003. Microb. Pathog. 2018, 127, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shangguan, W.; Wang, J.; Liao, Z.; Fang, X.; Zhong, Q. Transcriptomic analysis reveals the antibiofilm mechanism of Lacticaseibacillus rhamnosus MS1 against Vibrio Parahaemolyticus. LWT Food Sci. Technol. 2023, 176, 114529. [Google Scholar] [CrossRef]
- Rocchetti, M.T.; Russo, P.; Capozzi, V.; Drider, D.; Spano, G.; Fiocco, D. Bioprospecting antimicrobials from Lactiplantibacillus plantarum: Key factors underlying its probiotic action. Int. J. Mol. Sci. 2021, 22, 2076. [Google Scholar] [CrossRef] [PubMed]
- de Lira, F.M.; Tanaka, F.Y.R.; Rios, E.A.; Carrilho, S.M.; de Abreu, S.S.; Ferreira, G.F.; Gonzaga, N.; Pereira, U.d.P.; Tamanini, R.; Fagnani, R.; et al. Identification of lactic acid bacteria with anti-listeria activity. Characterization and application of a bacteriocinogenic strain in the control of Listeria monocytogenes in cheese. J. Dairy Res. 2023, 90, 318–323. [Google Scholar] [CrossRef]
- Mattila, M.; Lindström, M.; Somervuo, P.; Markkula, A.; Korkeala, H. Role of flhA and motA in growth of Listeria monocytogenes at low temperatures. Int. J. Food Microbiol. 2011, 148, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Melcon, C.; Alonso-Calleja, C.; Capita, R. Architecture and viability of the biofilms formed by nine Listeria strains on various hydrophobic and hydrophilic materials. Appl. Sci. 2019, 9, 5256. [Google Scholar] [CrossRef]
- Woo, J.; Ahn, J. Probiotic-mediated. Competition, exclusion and displacement in biofilm formation by food-borne pathogens. Lett. Appl. Microbiol. 2013, 56, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ju, X.; Du, L.; Wang, L.; He, R.; Chen, Z. The Man-PTS subunit IIC is responsible for the sensitivity of Listeria monocytogenes to durancin GL. Food Sci. Nutr. 2020, 8, 150–161. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef]
- Jing, Y.; Zuo, J.; Phouthapane, V.; Chen, Z.; Han, X. An Optimized Method for Detecting AI-2 Signal Molecule by a Bioassay with Vibrio harveyi BB170. Microbiology 2021, 90, 383–391. [Google Scholar] [CrossRef]
- Govaert, M.; Smet, C.; Baka, M.; Janssens, T.; Van Impe, J. Influence of incubation conditions on the formation of model biofilms by Listeria monocytogenes and Salmonella Typhimurium on abiotic surfaces. J. Appl. Microbiol. 2018, 125, 1890–1900. [Google Scholar] [CrossRef]
- Wang, J.B.; Yu, L.Y.; Zeng, X.; Zheng, J.W.; Wang, B.; Pan, L. Screening of probiotics with efficient α-glucosidase inhibitory ability and study on the structure and function of its extracellular polysaccharide. Food Biosci. 2022, 45, 101452. [Google Scholar] [CrossRef]
- Felz, S.; Vermeulen, P.; van Loosdrecht, M.C.M.; Lin, Y.M. Chemical characterization methods for the analysis of structural extracellular polymeric substances (EPS). Water Res. 2019, 157, 201–208. [Google Scholar] [CrossRef]
- Sutrina, S.L.; McGeary, T.; Bourne, C.-A. The phosphoenolpyruvate: Sugar phosphotransferase system and biofilms in gram-positive bacteria. J. Mol. Microb. Biotechnol. 2007, 12, 269–272. [Google Scholar] [CrossRef]
- Peng, Z.; Ehrmann, M.A.; Waldhuber, A.; Niemeyer, C.; Miethke, T.; Frick, J.S.; Xiong, T.; Vogel, R.F. Phosphotransferase systems in Enterococcus faecalis OG1RF enhance anti-stress capacity in vitro and in vivo. Res. Microbiol. 2017, 168, 558–566. [Google Scholar] [CrossRef]
- Ake, F.M.D.; Joyet, P.; Deutscher, J.; Milohanic, E. Mutational analysis of glucose transport regulation and glucose-mediated virulence gene repression in Listeria monocytogenes. Mol. Microbiol. 2011, 81, 274–293. [Google Scholar] [CrossRef]
- Chen, Q.; Amster-Choder, O. BglF, the sensor of the bgl system and the beta-glucosides permease of Escherichia coli: Evidence for dimerization and intersubunit phosphotransfer. Biochemistry 1998, 37, 8714–8723. [Google Scholar] [CrossRef]
- Tran, C.T.H.; Nosworthy, N.J.; Kondyurin, A.; McKenzie, D.R.; Bilek, M.M.M. CelB and β-glucosidase immobilization for carboxymethyl cellulose hydrolysis. RSC Adv. 2013, 3, 23604–23611. [Google Scholar] [CrossRef]
- Tamar, E.; Koler, M.; Vaknin, A. The role of motility and chemotaxis in the bacterial colonization of protected surfaces. Sci. Rep. 2016, 6, 19616. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, X.K.; Peng, L.H.; Zhu, Y.T.; Yoshida, A.; Osatomi, K.; Yang, J.L. The Flagellar Gene Regulates Biofilm Formation and Mussel Larval Settlement and Metamorphosis. Int. J. Mol. Sci. 2020, 21, 710. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Kotlyarov, S.; Kotlyarova, A. Clinical Significance of Lipid Transport Function of ABC Transporters in the Innate Immune System. Membranes 2022, 12, 1083. [Google Scholar] [CrossRef]
- De Leon, K.B.; Zane, G.M.; Trotter, V.V.; Krantz, G.P.; Arkin, A.P.; Butland, G.P.; Walian, P.J.; Fields, M.W.; Wall, J.D. Unintended laboratory-driven evolution reveals genetic requirements for biofilm formation by Desulfovibrio vulgaris Hildenborough. Mbio 2017, 8, e01696-17. [Google Scholar] [CrossRef]
- Lee, Y.; Song, S.; Sheng, L.L.; Zhu, L.; Kim, J.S.; Wood, T.K. Substrate binding protein DppA1 of ABC transporter DppBCDF increases biofilm formation in Pseudomonas aeruginosa by inhibiting Pf5 prophage lysis. Front. Microbiol. 2018, 9, 30. [Google Scholar] [CrossRef]
- Shaw, C.; Hess, M.; Weimer, B.C. Two-component systems regulate bacterial virulence in response to the host gastrointestinal environment and metabolic cues. Virulence 2022, 13, 1666–1680. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Dou, Q.; Du, L.C.; Wang, Y. QseB/QseC: A two-component system globally regulating bacterial behaviors. Trends Microbiol. 2023, 31, 749–762. [Google Scholar] [CrossRef]
- Urban-Chmiel, R.; Marek, A.; Wieczorek, K.; Dec, M.; Stepien-Pysniak, D.; Nowaczek, A.; Osek, J. Antibiotic resistance in bacteria-A review. Antibiotics 2022, 11, 1079. [Google Scholar] [CrossRef]
- Alejandro-Navarreto, X.; Freitag, N.E. Revisiting old friends: Updates on the role of two-component signaling systems in Listeria monocytogenes survival and pathogenesis. Infect. Immun. 2024, 92, e0034523. [Google Scholar] [CrossRef]
- Guffey, A.A.; Loll, P.J. Regulation of resistance in vancomycin-resistant Enterococci: The VanRS two-component system. Microorganisms 2021, 9, 2026. [Google Scholar] [CrossRef]
- Daisley, B.A.; Koenig, D.; Engelbrecht, K.; Doney, L.; Hards, K.; Al, K.F.; Reid, G.; Burton, J.P. Emerging connections between gut microbiome bioenergetics and chronic metabolic diseases. Cell Rep. 2021, 37, 110087. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Homma, H. D-Amino acid metabolism in bacteria. J. Biochem. 2021, 170, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Neis, E.; Dejong, C.H.C.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef]
- Sharma, V.; Rodionov, D.A.; Leyn, S.A.; Tran, D.; Iablokov, S.N.; Ding, H.; Peterson, D.A.; Osterman, A.L.; Peterson, S.N. B-vitamin sharing promotes stability of gut microbial communities. Front. Microbiol. 2019, 10, 1485. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Description | FC (T/C) | Log2FC (T/C) |
|---|---|---|---|
| lmo0344 | short chain dehydrogenase | 22.077 | 4.464464 |
| lmo0345 | sugar-phosphate isomerase | 21.196 | 4.405722 |
| lmo0343 | transaldolase | 21.032 | 4.394532 |
| lmo0342 | transketolase | 18.269 | 4.191303 |
| lmo0347 | dihydroxyacetone kinase | 16.727 | 4.064068 |
| lmo0348 | dihydroxyacetone kinase | 14.344 | 3.842421 |
| lmo1249 | hypothetical protein | 13.607 | 3.766271 |
| lmo1997 | PTS mannose transporter subunit IIA | 5.734 | 2.51942 |
| lmo0346 | triosephosphate isomerase | 5.534 | 2.468207 |
| Gene Name | Gene Description | FC (T/C) | Log2FC (T/C) |
|---|---|---|---|
| - | hypothetical protein AVP43_02707 [Geobacillus stearothermophilus] | 0.002 | −8.673013 |
| lmo1190 | hypothetical protein | 0.036 | −4.779108 |
| lmo1151 | PduA protein | 0.038 | −4.724699 |
| lmo1152 | PduB protein | 0.041 | −4.602022 |
| lmo1162 | hypothetical protein | 0.042 | −4.573928 |
| lmo1154 | diol dehydratase subunit gamma | 0.042 | −4.571224 |
| lmo1164 | ATP:cob(I)alamin adenosyltransferase PduO | 0.051 | −4.299481 |
| lmo1153 | propanediol dehydratase subunit alpha | 0.052 | −4.266549 |
| lmo1158 | PduK protein | 0.054 | −4.211739 |
| lmo1156 | diol dehydratase-reactivating factor large subunit | 0.054 | −4.200381 |
| lmo1165 | ethanolamine utilization protein EutE | 0.055 | −4.189399 |
| - | hypothetical protein LM700514_40645 [Listeria monocytogenes] | 0.057 | −4.133942 |
| lmo1161 | ethanolamine utilization protein EutJ | 0.06 | −4.054854 |
| lmo1166 | NADPH-dependent butanol dehydrogenase | 0.064 | −3.963328 |
| lmo1155 | diol dehydratase subunit gamma | 0.07 | −3.842152 |
| lmo1159 | carboxysome structural protein | 0.07 | −3.827296 |
| lmo1160 | PduL protein | 0.073 | −3.778299 |
| lmo1163 | carbon dioxide concentrating mechanism protein | 0.081 | −3.623202 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, J.; Zhang, X.; Ismael, M.; Zhong, Q. Anti-Biofilm Effects of Z102-E of Lactiplantibacillus plantarum against Listeria monocytogenes and the Mechanism Revealed by Transcriptomic Analysis. Foods 2024, 13, 2495. https://doi.org/10.3390/foods13162495
Wei J, Zhang X, Ismael M, Zhong Q. Anti-Biofilm Effects of Z102-E of Lactiplantibacillus plantarum against Listeria monocytogenes and the Mechanism Revealed by Transcriptomic Analysis. Foods. 2024; 13(16):2495. https://doi.org/10.3390/foods13162495
Chicago/Turabian StyleWei, Jinyuan, Xingguo Zhang, Mohamedelfatieh Ismael, and Qingping Zhong. 2024. "Anti-Biofilm Effects of Z102-E of Lactiplantibacillus plantarum against Listeria monocytogenes and the Mechanism Revealed by Transcriptomic Analysis" Foods 13, no. 16: 2495. https://doi.org/10.3390/foods13162495