
Advances in the Effects of Heat Stress on Ovarian Granulosa Cells: Unveiling Novel Ferroptosis Pathways

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Heat Stress Affects Steroid Hormone Production
2.1. On Estradiol
2.1.1. In Mammals
In Artiodactyla
In Rodents
2.1.2. In Poultry
2.2. On Progesterone
2.2.1. In Mammals
In Artiodactyla
In Rodents
2.2.2. In Poultry
2.3. On Follicle-Stimulating Hormone (FSH)
2.3.1. In Mammals
In Artiodactyla
In Rodents
2.3.2. In Poultry
3. Regulatory Mechanisms of Apoptosis in Granulosa Cells Due to Heat Stress
3.1. Cell Proliferation and Cycle Transition under Heat Stress Invasion
3.1.1. In Mammals
In Artiodactyla
In Rodents
3.1.2. In Poultry
3.2. Mitochondria Damaged by Heat Stress: Kinetic Imbalance and Autophagy
3.2.1. In Mammals
In Primates
In Artiodactyla
3.2.2. In Poultry
3.3. Heat Stress Affects the Expression of the Heat Shock Protein Family in HSPs
3.4. Heat Stress Triggers the Fas/FasL Extrinsic Apoptotic Pathway
3.5. Heat Stress Causes Imbalance in Mitochondrial Dynamics
3.6. Heat Stress Triggers ER Endoplasmic Reticulum Stress
3.7. Heat Stress Activates the Nrf2/HO-1 Pathway
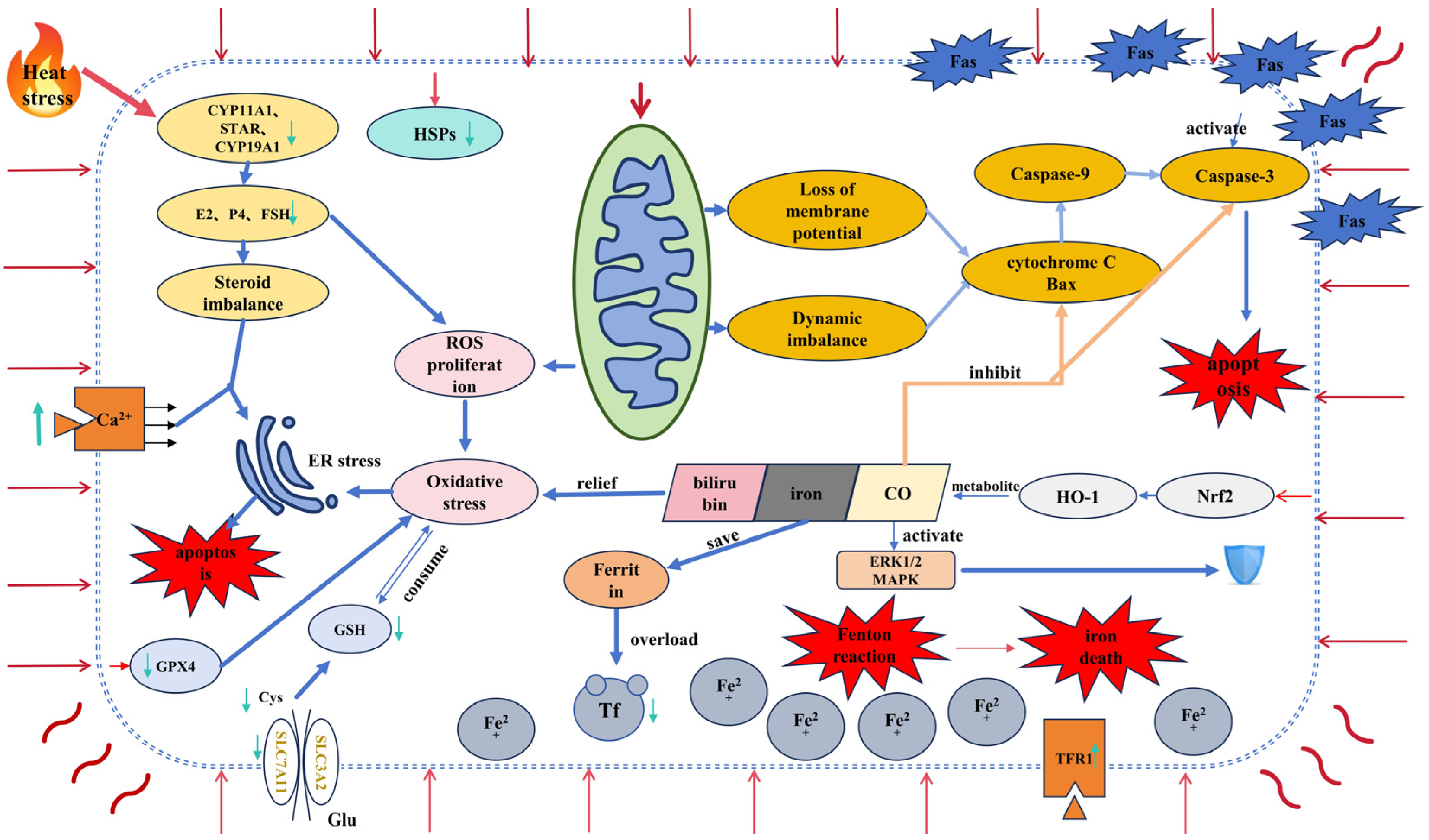
4. Interaction between Heat Stress and Ferroptosis in Granulosa Cells
4.1. Heat Stress and Ferroptosis
4.2. Ferroptosis and Granulocytes
4.3. Interaction Mechanism
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, Z.; Meidan, R.; Braw-Tal, R.; Wolfenson, D. Immediate and Delayed Effects of Heat Stress on Follicular Development and Its Association with Plasma FSH and Inhibin Concentration in Cows. J. Reprod. Fertil. 2000, 120, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Ohshima, I.; Ozawa, M.; Takahashi, S.; Tajima, A.; Shiota, M.; Miyazaki, H.; Kanai, Y. Heat Stress Diminishes Gonadotropin Receptor Expression and Enhances Susceptibility to Apoptosis of Rat Granulosa Cells. Reproduction 2005, 129, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.H.; Miyano, T. Interaction between Growing Oocytes and Granulosa Cells in Vitro. Reprod. Med. Biol. 2020, 19, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z. Effect of Heat Stress on Reproduction in Dairy Cows: Insights into the Cellular and Molecular Responses of the Oocyte. Annu. Rev. Anim. Biosci. 2017, 5, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, H.; Tian, Z.; Wu, Y.; Wang, Y.; Fang, Y.; Lin, L.; Han, Y.; Wu, S.; Haq, I.; et al. Effects of Chronic Heat Stress on Granulosa Cell Apoptosis and Follicular Atresia in Mouse Ovary. J. Anim. Sci. Biotechnol. 2016, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Bridges, P.J.; Brusie, M.A.; Fortune, J.E. Elevated Temperature (Heat Stress) in Vitro Reduces Androstenedione and Estradiol and Increases Progesterone Secretion by Follicular Cells from Bovine Dominant Follicles. Domest. Anim. Endocrinol. 2005, 29, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Luo, P.; Yang, Y.; Fu, X.; Li, B.; Shen, X.; Xu, D.; Huang, Y.; Tian, Y.; Liu, W. Drp1 Regulated PINK1-Dependent Mitophagy Protected Duck Follicular Granulosa Cells from Acute Heat Stress Injury. Poult. Sci. 2024, 103, 103247. [Google Scholar] [CrossRef] [PubMed]
- Alemu, T.W.; Pandey, H.O.; Salilew Wondim, D.; Gebremedhn, S.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Oxidative and Endoplasmic Reticulum Stress Defense Mechanisms of Bovine Granulosa Cells Exposed to Heat Stress. Riogenology 2018, 110, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Bauer, M. Heat Shock Proteins in Porcine Ovary: Synthesis, Accumulation and Regulation by Stress and Hormones. Cell Stress. Chaperones 2011, 16, 379–387. [Google Scholar] [CrossRef]
- Inoue, N.; Maeda, A.; Matsuda-Minehata, F.; Fukuta, K.; Manabe, N. Expression and Localization of Fas Ligand and Fas During Atresia in Porcine Ovarian Follicles. J. Reprod. Dev. 2006, 52, 723–730. [Google Scholar] [CrossRef]
- Ni, Z.; Li, Y.; Song, D.; Ding, J.; Mei, S.; Sun, S.; Cheng, W.; Yu, J.; Zhou, L.; Kuang, Y.; et al. Iron-Overloaded Follicular Fluid Increases the Risk of Endometriosis-Related Infertility by Triggering Granulosa Cell Ferroptosis and Oocyte Dysmaturity. Cell Death Dis. 2022, 13, 579. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, M.; Gong, N.; Tian, P.; Deng, H. Se Improves GPX4 Expression and SOD Activity to Alleviate Heat-Stress-Induced Ferroptosis-like Death in Goat Mammary Epithelial Cells. Anim. Cells Syst. 2021, 25, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Billig, H.; Furuta, I.; Hsueh, A.J. Estrogens Inhibit and Androgens Enhance Ovarian Granulosa Cell Apoptosis. Endocrinology 1993, 133, 2204–2212. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z.; Meidan, R.; Shaham-Albalancy, A.; Braw-Tal, R.; Wolfenson, D. Delayed Effect of Heat Stress on Steroid Production in Medium-Sized and Preovulatory Bovine Follicles. Reproduction 2001, 121, 745–751. [Google Scholar] [CrossRef]
- De Castro, E.; Paula, L.A.; Andrzejewski, J.; Julian, D.; Spicer, L.J.; Hansen, P.J. Oxygen and Steroid Concentrations in Preovulatory Follicles of Lactating Dairy Cows Exposed to Acute Heat Stress. Theriogenology 2008, 69, 805–813. [Google Scholar] [CrossRef] [PubMed]
- López-Gatius, F.; López-Béjar, M.; Fenech, M.; Hunter, R.H.F. Ovulation Failure and Double Ovulation in Dairy Cattle: Risk Factors and Effects. Theriogenology 2005, 63, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Mani, A.M.; Fenwick, M.A.; Cheng, Z.; Sharma, M.K.; Singh, D.; Wathes, D.C. IGF1 Induces Up-Regulation of Steroidogenic and Apoptotic Regulatory Genes via Activation of Phosphatidylinositol-Dependent Kinase/AKT in Bovine Granulosa Cells. Reproduction 2010, 139, 139–151. [Google Scholar] [CrossRef]
- Li, L.; Wu, J.; Luo, M.; Sun, Y.; Wang, G. The Effect of Heat Stress on Gene Expression, Synthesis of Steroids, and Apoptosis in Bovine Granulosa Cells. Cell Stress. Chaperones 2016, 21, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Vanselow, J.; Vernunft, A.; Koczan, D.; Spitschak, M.; Kuhla, B. Exposure of Lactating Dairy Cows to Acute Pre-Ovulatory Heat Stress Affects Granulosa Cell-Specific Gene Expression Profiles in Dominant Follicles. PLoS ONE 2016, 11, e0160600. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Lew, B.J.; Thatcher, W.W.; Graber, Y.; Meidan, R. Seasonal and Acute Heat Stress Effects on Steroid Production by Dominant Follicles in Cows. Anim. Reprod. Sci. 1997, 47, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Dou, J.; Wang, Y.; Jiang, X.; Khan, M.Z.; Luo, H.; Usman, T.; Zhu, H. Evaluation of Heat Stress Effects on Cellular and Transcriptional Adaptation of Bovine Granulosa Cells. J. Anim. Sci. Biotechnol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Faheem, M.S.; Dessouki, S.M.; Abdel- Rahman, F.E.S.; Ghanem, N. Physiological and Molecular Aspects of Heat-Treated Cultured Granulosa Cells of Egyptian Buffalo (Bubalus Bubalis). Anim. Reprod. Sci. 2021, 224, 106665. [Google Scholar] [CrossRef] [PubMed]
- Guzeloglu, A.; Ambrose, J.D.; Kassa, T.; Diaz, T.; Thatcher, M.J.; Thatcher, W.W. Long-Term Follicular Dynamics and Biochemical Characteristics of Dominant Follicles in Dairy Cows Subjected to Acute Heat Stress. Anim. Reprod. Sci. 2001, 66, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V. Effect of Two Types of Stress (Heat Shock/High Temperature and Malnutrition/Serum Deprivation) on Porcine Ovarian Cell Functions and Their Response to Hormones. J. Exp. Biol. 2010, 213, 2125–2130. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, S.; Cai, L.; Ma, W.; Shi, Z. Lipopolysaccharide and Heat Stress Impair the Estradiol Biosynthesis in Granulosa Cells via Increase of HSP70 and Inhibition of Smad3 Phosphorylation and Nuclear Translocation. Cell. Signal. 2017, 30, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Li, L.; Xiao, C.; Sun, Y.; Wang, G.-L. Heat Stress Impairs Mice Granulosa Cell Function by Diminishing Steroids Production and Inducing Apoptosis. Mol. Cell Biochem. 2016, 412, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.K.; Humphreys, G.I.; Nardulli, A.M. 17β-Estradiol Increases Expression of the Oxidative Stress Response and DNA Repair Protein Apurinic Endonuclease (Ape1) in the Cerebral Cortex of Female Mice Following Hypoxia. J. Steroid Biochem. Mol. Biol. 2013, 138, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Aten, R.F.; Behrman, H.R. Heat Shock Protein Induction Blocks Hormone-Sensitive Steroidogenesis in Rat Luteal Cells. Steroids 1994, 59, 4–9. [Google Scholar] [CrossRef]
- An, G.; Chen, X.; Li, C.; Zhang, L.; Wei, M.; Chen, J.; Ma, Q.; Yang, D.; Wang, J. Pathophysiological Changes in Female Rats with Estrous Cycle Disorder Induced by Long-Term Heat Stress. BioMed Res. Int. 2020, 2020, 4701563. [Google Scholar] [CrossRef]
- Lin, H.; De Vos, D.; Decuypere, E.; Buyse, J. Dynamic Changes in Parameters of Redox Balance after Mild Heat Stress in Aged Laying Hens (Gallus Gallus Domesticus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 147, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-M.; Liu, L.-P.; Yin, B.; Liu, Y.-Y.; Dong, W.-W.; Gong, S.; Zhang, J.; Tan, J.-H. Heat Stress Decreases Egg Production of Laying Hens by Inducing Apoptosis of Follicular Cells via Activating the FasL/Fas and TNF-α Systems. Poult. Sci. 2020, 99, 6084–6093. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Hu, M.; Gu, L.; Lei, M.; Chen, Z.; Zhu, H.; Chen, R. Effect of Heat Stress on Egg Production, Steroid Hormone Synthesis, and Related Gene Expression in Chicken Preovulatory Follicular Granulosa Cells. Animals 2022, 12, 1467. [Google Scholar] [CrossRef]
- Shen, X.; Zhao, X.; He, H.; Zhang, Y.; Zhu, Q.; Yin, H. Transcriptome Profiling Reveals SLC5A5 Regulates Chicken Ovarian Follicle Granulosa Cell Proliferation, Apoptosis, and Steroid Hormone Synthesis. Poult. Sci. 2024, 103, 103241. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Huang, X.; Chen, S.; Li, X.; Fu, X.; Xu, D.; Tian, Y.; Liu, W.; Huang, Y. The Effect of Heat Stress on Proliferation, Synthesis of Steroids, and Gene Expression of Duck Granulosa Cells. Anim. Sci. J. 2021, 92, e13617. [Google Scholar] [CrossRef]
- Wise, M.E.; Armstrong, D.V.; Huber, J.T.; Hunter, R.; Wiersma, F. Hormonal Alterations in the Lactating Dairy Cow in Response to Thermal Stress. J. Dairy Sci. 1988, 71, 2480–2485. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Hu, L.; Luo, H.; Abbas, Z.; Umer, S.; Zhao, S.; Xu, Q.; Khan, A.; Wang, Y.; Zhu, H.; et al. Investigation of Metabolome Underlying the Biological Mechanisms of Acute Heat Stressed Granulosa Cells. Int. J. Mol. Sci. 2022, 23, 2146. [Google Scholar] [CrossRef] [PubMed]
- Abdelnour, S.A.; Swelum, A.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Abdo, M. Cellular and Functional Adaptation to Thermal Stress in Ovarian Granulosa Cells in Mammals. J. Therm. Biol. 2020, 92, 102688. [Google Scholar] [CrossRef] [PubMed]
- Nteeba, J.; Sanz-Fernandez, M.V.; Rhoads, R.P.; Baumgard, L.H.; Ross, J.W.; Keating, A.F. Heat Stress Alters Ovarian Insulin-Mediated Phosphatidylinositol-3 Kinase and Steroidogenic Signaling in Gilt Ovaries1. Biol. Reprod. 2015, 92, 148. [Google Scholar] [CrossRef] [PubMed]
- Bidne, K.L.; Romoser, M.R.; Ross, J.W.; Baumgard, L.H.; Keating, A.F. Heat Stress during the Luteal Phase Decreases Luteal Size but Does Not Affect Circulating Progesterone in Gilts1. J. Anim. Sci. 2019, 97, 4314–4322. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J. Progesterone as a Regulator of Granulosa Cell Viability. J. Steroid Biochem. Mol. Biol. 2003, 85, 167–173. [Google Scholar] [CrossRef]
- Peluso, J.J.; Pappalardo, A. Progesterone Mediates Its Anti-Mitogenic and Anti-Apoptotic Actions in Rat Granulosa Cells Through a Progesterone-Binding Protein With Gamma Aminobutyric AcidA Receptor-Like Features1. Biol. Reprod. 1998, 58, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Pappalardo, A. Progesterone Regulates Granulosa Cell Viability through a Protein Kinase G-Dependent Mechanism That May Involve 14-3-3σ1. Biol. Reprod. 2004, 71, 1870–1878. [Google Scholar] [CrossRef] [PubMed]
- Luciano, A.M.; Peluso, J.J. Effect of in Vivo Gonadotropin Treatment on the Ability of Progesterone, Estrogen, and Cyclic Adenosine 5′-Monophosphate to Inhibit Insulin-Dependent Granulosa Cell Mitosis in Vitro1. Biol. Reprod. 1995, 53, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J.; Luciano, A.M.; Pappalardo, A.; White, B.A. Cellular and Molecular Mechanisms That Mediate Insulin-Dependent Rat Granulosa Cell Mitosis1. Biol. Reprod. 1995, 52, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Luciano, A.M.; Pappalardo, A.; Ray, C.; Peluso, J.J. Epidermal Growth Factor Inhibits Large Granulosa Cell Apoptosis by Stimulating Progesterone Synthesis and Regulating the Distribution of Intracellular Free Calcium1. Biol. Reprod. 1994, 51, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Zhang, M.; Yu, H.; Mustafa, S.; Shafiq, M.; Wei, Q.; Wang, W.; Jan, M.; Mao, D. Heat Exposure Affected the Reproductive Performance of Pregnant Mice: Enhancement of Autophagy and Alteration of Subcellular Structure in the Corpus Luteum. Reprod. Biol. 2019, 19, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zeitoun, M.; Derar, D.; Ali, A.; Alharbi, Y. Expression of Hormones, Cytokines, and Antioxidants in Heat-Stressed Subfertile Female Dromedaries. Animals 2022, 12, 2125. [Google Scholar] [CrossRef] [PubMed]
- Amitha, J.P.; Krishnan, G.; Bagath, M.; Sejian, V.; Bhatta, R. Heat Stress Impact on the Expression Patterns of Different Reproduction Related Genes in Malabari Goats. Theriogenology 2019, 131, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Roth, Z.; Meidan, R. Impaired Reproduction in Heat-Stressed Cattle: Basic and Applied Aspects. Anim. Reprod. Sci. 2000, 60–61, 535–547. [Google Scholar] [CrossRef]
- Yang, F.; Chen, Y.; Liu, Q.; Dai, S.; Zeng, S. Dynamics and Regulations of BimEL Ser65 and Thr112 Phosphorylation in Porcine Granulosa Cells during Follicular Atresia. Cells 2020, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Margolin, Y.; Aten, R.F.; Behrman, H.R. Antigonadotropic and Antisteroidogenic Actions of Peroxide in Rat Granulosa Cells*. Endocrinology 1990, 127, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.A.; Woodcock, J.R.; Kent, T.R. Effect of Activin A and Inhibin A on Expression of the Inhibin/Activin β-B-Subunit and Gonadotropin Receptors in Granulosa Cells of the Hen. General. Comp. Endocrinol. 2006, 147, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.; Nagaoka, K.; Watanabe, G. Yolk Immunoreactive Corticosterone in Hierarchical Follicles of Japanese Quail (Coturnix Japonica) Exposed to Heat Challenge. General. Comp. Endocrinol. 2019, 279, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Kikusato, M.; Toyomizu, M. Crucial Role of Membrane Potential in Heat Stress-Induced Overproduction of Reactive Oxygen Species in Avian Skeletal Muscle Mitochondria. PLoS ONE 2013, 8, e64412. [Google Scholar] [CrossRef]
- Khan, A.; Khan, M.Z.; Dou, J.; Umer, S.; Xu, H.; Sammad, A.; Zhu, H.-B.; Wang, Y. RNAi-Mediated Silencing of Catalase Gene Promotes Apoptosis and Impairs Proliferation of Bovine Granulosa Cells under Heat Stress. Animals 2020, 10, 1060. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; He, C.-J.; Ji, P.-Y.; Zhuo, Z.-Y.; Tian, X.-Z.; Wang, F.; Tan, D.-X.; Liu, G.-S. Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress. Int. J. Mol. Sci. 2014, 15, 21090–21104. [Google Scholar] [CrossRef] [PubMed]
- Kayampilly, P.P.; Menon, K.M.J. Follicle-Stimulating Hormone Inhibits Adenosine 5′-Monophosphate-Activated Protein Kinase Activation and Promotes Cell Proliferation of Primary Granulosa Cells in Culture through an Akt-Dependent Pathway. Endocrinology 2009, 150, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Q.; Li, R.-F.; Xi, S.-M.; Hu, S.; Jia, Z.-Q.; Li, S.-P.; Wen, X.-L.; Song, Y.-K.; Li, S.; Li, S.-P.; et al. Systematical Analysis of Impacts of Heat Stress on the Proliferation, Apoptosis and Metabolism of Mouse Hepatocyte. J. Physiol. Sci. 2012, 62, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.-S.; Dighe, P.A.; Mezera, V.; Monternier, P.-A.; Brand, M.D. Production of Superoxide and Hydrogen Peroxide from Specific Mitochondrial Sites under Different Bioenergetic Conditions. J. Biol. Chem. 2017, 292, 16804–16809. [Google Scholar] [CrossRef] [PubMed]
- Jendrach, M.; Mai, S.; Pohl, S.; Vöth, M.; Bereiter-Hahn, J. Short- and Long-Term Alterations of Mitochondrial Morphology, Dynamics and mtDNA after Transient Oxidative Stress. Mitochondrion 2008, 8, 293–304. [Google Scholar] [CrossRef]
- Wrzecińska, M.; Kowalczyk, A.; Kordan, W.; Cwynar, P.; Czerniawska-Piątkowska, E. Disorder of Biological Quality and Autophagy Process in Bovine Oocytes Exposed to Heat Stress and the Effectiveness of In Vitro Fertilization. Int. J. Mol. Sci. 2023, 24, 11164. [Google Scholar] [CrossRef]
- Chen, K.-L.; Wang, H.-L.; Jiang, L.-Z.; Qian, Y.; Yang, C.-X.; Chang, W.-W.; Zhong, J.-F.; Xing, G.-D. Heat Stress Induces Apoptosis through Disruption of Dynamic Mitochondrial Networks in Dairy Cow Mammary Epithelial Cells. Vitr. Cell. Dev. Biol. Anim. 2020, 56, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Bei, M.; Wang, Q.; Yu, W.; Han, L.; Yu, J. Effects of Heat Stress on Ovarian Development and the Expression of HSP Genes in Mice. J. Therm. Biol. 2020, 89, 102532. [Google Scholar] [CrossRef] [PubMed]
- Song, E.; Tang, S.; Xu, J.; Yin, B.; Bao, E.; Hartung, J. Lenti-siRNA Hsp60 Promote Bax in Mitochondria and Induces Apoptosis during Heat Stress. Biochem. Biophys. Res. Commun. 2016, 481, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhn, S.; Gad, A.; Aglan, H.S.; Laurincik, J.; Prochazka, R.; Salilew-Wondim, D.; Hoelker, M.; Schellander, K.; Tesfaye, D. Extracellular Vesicles Shuttle Protective Messages against Heat Stress in Bovine Granulosa Cells. Sci. Rep. 2020, 10, 15824. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Swelum, A.A.-A.; Elsafadi, M.; Mahmood, A.; Osama, A.; Shikshaky, H.; Alfayez, M.; Alowaimer, A.N.; Magdeldin, S. Thermotolerance and Plasticity of Camel Somatic Cells Exposed to Acute and Chronic Heat Stress. J. Adv. Res. 2020, 22, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Steiner, C.-W.; Gröger, M.; Winkler, S.; Graninger, W.; Smolen, J.; Xu, Q.; Steiner, G. Activation of Fas Inhibits Heat-induced Activation of HSF1 and Up-regulation of Hsp70. FASEB J. 1999, 13, 833–842. [Google Scholar] [CrossRef]
- Peter, M.E.; Krammer, P.H. The CD95(APO-1/Fas) DISC and Beyond. Cell Death Differ. 2003, 10, 26–35. [Google Scholar] [CrossRef]
- Kim, J.-M.; Yoon, Y.-D.; Tsang, B.K. Involvement of the Fas/Fas Ligand System in P53-Mediated Granulosa Cell Apoptosis during Follicular Development and Atresia. Endocrinology 1999, 140, 2307–2317. [Google Scholar] [CrossRef] [PubMed]
- Denning, T.L.; Takaishi, H.; Crowe, S.E.; Boldogh, I.; Jevnikar, A.; Ernst, P.B. Oxidative Stress Induces the Expression of Fas and Fas Ligand and Apoptosis in Murine Intestinal Epithelial Cells. Free. Radic. Biol. Med. 2002, 33, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Stahl, B.C.; Dewolf, W.C.; Morgentaler, A. P53 and Fas Are Sequential Mechanisms of Testicular Germ Cell A Poptosis. J. Androl. 2002, 23, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Itami, N.; Shirasuna, K.; Kuwayama, T.; Iwata, H. Short-Term Heat Stress Induces Mitochondrial Degradation and Biogenesis and Enhances Mitochondrial Quality in Porcine Oocytes. J. Therm. Biol. 2018, 74, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome c Promotes Caspase-9 Activation by Inducing Nucleotide Binding to Apaf-1. J. Biol. Chem. 2000, 275, 31199–31203. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M.; Kaufman, R.J. ER Stress and the Unfolded Protein Response. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2005, 569, 29–63. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.-H.; Lin, F.-L.; Hou, S.-M.; Liu, J.-F. Hyperthermia Induces Apoptosis through Endoplasmic Reticulum and Reactive Oxygen Species in Human Osteosarcoma Cells. Int. J. Mol. Sci. 2014, 15, 17380–17395. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Yang, Y.; Li, X.; Chen, F.; Cui, C.; Hu, L.; Li, Q.; Liu, W.; Jin, Y. Endoplasmic Reticulum Stress Is Involved in Granulosa Cell Apoptosis during Follicular Atresia in Goat Ovaries. Mol. Reprod. Devel 2012, 79, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Jin, E.; Yin, Q.; Che, C.; He, S. Boron Attenuates Heat Stress–Induced Apoptosis by Inhibiting Endoplasmic Reticulum Stress in Mouse Granulosa Cells. Biol. Trace Elem. Res. 2021, 199, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Yin, Q.; Jin, E.; Chen, H.; He, S. Selenium Attenuates Chronic Heat Stress-Induced Apoptosis via the Inhibition of Endoplasmic Reticulum Stress in Mouse Granulosa Cells. Molecules 2020, 25, 557. [Google Scholar] [CrossRef]
- Hayes, J.D.; McMahon, M. NRF2 and KEAP1 Mutations: Permanent Activation of an Adaptive Response in Cancer. Trends Biochem. Sci. 2009, 34, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Li, J.; Johnson, D.A.; Stein, T.D.; Kraft, A.D.; Calkins, M.J.; Jakel, R.J.; Johnson, J.A. Nrf2, a Multi-organ Protector? FASEB J. 2005, 19, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Khadrawy, O.; Gebremedhn, S.; Salilew-Wondim, D.; Taqi, M.; Neuhoff, C.; Tholen, E.; Hoelker, M.; Schellander, K.; Tesfaye, D. Endogenous and Exogenous Modulation of Nrf2 Mediated Oxidative Stress Response in Bovine Granulosa Cells: Potential Implication for Ovarian Function. Int. J. Mol. Sci. 2019, 20, 1635. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, L.; Li, M.; Li, L.; Wang, L. Heme Oxygenase 1 Regulates Apoptosis Induced by Heat Stress in Bovine Ovarian Granulosa Cells via the ERK1/2 Pathway. J. Cell Physiol. 2019, 234, 3961–3972. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Elsheikh, N.A.H.; Li, C.; Yang, F.; Wang, G.; Li, L. HO-1 Reduces Heat Stress-Induced Apoptosis in Bovine Granulosa Cells by Suppressing Oxidative Stress. Aging 2019, 11, 5535–5547. [Google Scholar] [CrossRef] [PubMed]
- Balla, G.; Jacob, H.S.; Balla, J.; Rosenberg, M.; Nath, K.; Apple, F.; Eaton, J.W.; Vercellotti, G.M. Ferritin: A Cytoprotective Antioxidant Strategem of Endothelium. J. Biol. Chem. 1992, 267, 18148–18153. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-T.; Yen, C.-J.; Chang, C.-C.; Huang, K.-T.; Chen, K.-H.; Zhang, R.-Y.; Lee, P.-Y.; Miaw, S.-C.; Huang, J.-W.; Chiang, C.-K.; et al. Ferritin Heavy Chain Mediates the Protective Effect of Heme Oxygenase-1 against Oxidative Stress. Biochim. Biophys. Acta (BBA) General. Subj. 2015, 1850, 2506–2517. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct Evidence That Sulfhydryl Groups of Keap1 Are the Sensors Regulating Induction of Phase 2 Enzymes That Protect against Carcinogens and Oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.H. Glucose Availability Is a Decisive Factor for Nrf2-Mediated Gene Expression. Redox Biol. 2013, 1, 359–365. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; O’Connor, T.; Yamamoto, M. Keap1 Regulates Both Cytoplasmic-nuclear Shuttling and Degradation of Nrf2 in Response to Electrophiles. Genes. Cells 2003, 8, 379–391. [Google Scholar] [CrossRef]
- Cui, W.; Bai, Y.; Luo, P.; Miao, L.; Cai, L. Preventive and Therapeutic Effects of MG132 by Activating Nrf2-ARE Signaling Pathway on Oxidative Stress-Induced Cardiovascular and Renal Injury. Oxidative Med. Cell. Longev. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Baird, L.; Dinkova-Kostova, A.T. The Cytoprotective Role of the Keap1–Nrf2 Pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef]
- Jang, H.J.; Hong, E.M.; Kim, M.; Kim, J.H.; Jang, J.; Park, S.W.; Byun, H.W.; Koh, D.H.; Choi, M.H.; Kae, S.H.; et al. Simvastatin Induces Heme Oxygenase-1 via NF-E2-Related Factor 2 (Nrf2) Activation through ERK and PI3K/Akt Pathway in Colon Cancer. Oncotarget 2016, 7, 46219–46229. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Ma, Y.D.; Liu, S.Q.; Huang, R.W.; Chen, J.J.; An, L.L.; Wu, J. SARS-CoV-2 Structural Proteins Modulated Blood-Testis Barrier-Related Proteins through Autophagy in the Primary Sertoli Cells. Viruses 2023, 15, 1272. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Li, H.; Graham, E.T.; Deik, A.A.; Eaton, J.K.; Wang, W.; Sandoval-Gomez, G.; Clish, C.B.; Doench, J.G.; Schreiber, S.L. Cytochrome P450 Oxidoreductase Contributes Tophospholipid Peroxidation in Ferroptosis. Nat. Chem. Biol. 2020, 16, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cai, X.; Qiu, M.; Deng, C.; Xue, H.; Zhang, J.; Yang, W.; XianZhong, W. Heat Stress Induces Ferroptosis of Porcine Sertoli Cells by Enhancing CYP2C9-Ras- JNK Axis. Theriogenology 2024, 215, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, W.; Zhou, H.; Wu, Q.; Duan, M.; Liu, C.; Wu, H.; Deng, W.; Shen, D.; Tang, Q. Ferritinophagy-Mediated Ferroptosis Is Involved in Sepsis-Induced Cardiac Injury. Free. Radic. Biol. Med. 2020, 160, 303–318. [Google Scholar] [CrossRef]
- Zheng, D.-W.; Lei, Q.; Zhu, J.-Y.; Fan, J.-X.; Li, C.-X.; Li, C.; Xu, Z.; Cheng, S.-X.; Zhang, X.-Z. Switching Apoptosis to Ferroptosis: Metal–Organic Network for High-Efficiency Anticancer Therapy. Nano Lett. 2017, 17, 284–291. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Yao, W.; Li, Q.; Liu, H.; Pan, Z. Initiation of Follicular Atresia: Gene Networks during Early Atresia in Pig Ovaries. Reproduction 2018, 156, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Hassannia, B.; Wiernicki, B.; Ingold, I.; Qu, F.; Van Herck, S.; Tyurina, Y.Y.; Bayır, H.; Abhari, B.A.; Angeli, J.P.F.; Choi, S.M.; et al. Nano-Targeted Induction of Dual Ferroptotic Mechanisms Eradicates High-Risk Neuroblastoma. J. Clin. Investig. 2018, 128, 3341–3355. [Google Scholar] [CrossRef] [PubMed]
- Suttner, D.M.; Dennery, P.A. Reversal of HO-1 Related Cytoprotection with Increased Expression Is Due to Reactive Iron. FASEB J. 1999, 13, 1800–1809. [Google Scholar] [CrossRef]
- Sun, X.; Ou, Z.; Chen, R.; Niu, X.; Chen, D.; Kang, R.; Tang, D. Activation of the p62-Keap1-NRF2 Pathway Protects against Ferroptosis in Hepatocellular Carcinoma Cells. Hepatology 2016, 63, 173–184. [Google Scholar] [CrossRef]
- Fan, Z.; Wirth, A.-K.; Chen, D.; Wruck, C.J.; Rauh, M.; Buchfelder, M.; Savaskan, N. Nrf2-Keap1 Pathway Promotes Cell Proliferation and Diminishes Ferroptosis. Oncogenesis 2017, 6, e371. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, X.-L.; Zeng, H.-F.; Han, Z.-Y. Methionine Alleviates Heat Stress-Induced Ferroptosis in Bovine Mammary Epithelial Cells through the Nrf2 Pathway. Ecotoxicol. Environ. Saf. 2023, 256, 114889. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Biswas, G.; Avadhani, N.G. Mitochondria-Targeted Heme Oxygenase-1 Induces Oxidative Stress and Mitochondrial Dysfunction in Macrophages, Kidney Fibroblasts and in Chronic Alcohol Hepatotoxicity. Redox Biol. 2014, 2, 273–283. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Taxonomic Group | Types/Strains | Heat Stress Environmental Settings | Main Findings | References |
|---|---|---|---|---|
| 1. Bovine | Dairy cattle | 36 °C and relative humidity 60% for five days | • Heat stress reduces E2 levels in GCs Heat stress has a delayed effect on steroid hormone secretion | [14] |
| Bovine | / | • FSH synergizes with IGF1 to increase cell number and CYP19A1 mRNA expression in bovine granulosa cells | [15] | |
| Dairy cattle | 38 °C or 40.5 °C for 12 h, | • Heat stress induced GC apoptosis through the BAX/BCL-2 pathway Heat stress reduced the steroidogenic gene messenger RNA (mRNA) expression and E2 synthesis | [16] | |
| Dairy cattle | (28 °C, 52% relative humidity, THI = 76) conditions for a total of 95 h | • Acute preovulatory heat stress specifically alters the gene expression profile of granulosa cells but does not induce stress-related genes and pathways | [17] | |
| Holstein dairy cows | Winter maintained at (36, 60% RH, THI = 88.3) for 3 days, 12 h per day; summer | • Seasonal heat stress has less effect on GC (estradiol-producing) biosynthesis capacity | [18] | |
| Dairy cattle | (39, 40 and 41 °C) for 2 h | • HS significantly reduced E2 and P4 levels in bGCs, while increasing intracellular ROS and apoptosis rates • HS can significantly regulate many differentially expressed genes (DEGs), such as BCL2L1, STAR, and CYP11A1, which are involved in apoptosis, steroidogenesis, and oxidative stress | [19] | |
| Egyptian buffalo (Bubalus bubalis) | Heat treatment at 40.5 °C for 24, 48 or 72 h | • Buffalo GCs treated with heat stress for 72 h maintain normal vigor, steroidogenesis, and transcriptional profiles | [20] | |
| Holstein dairy cattle | (43.3 °C) for seven days, eight hours per day | • Estradiol and progesterone levels in follicular fluid of dominant follicles did not differ after HS treatment | [21] | |
| Lactating Holstein cows | / | • Acute heat stress has no significant effect on follicular fluid PO2, estradiol-17b, or progesterone concentrations | [22] | |
| Non-pregnant cows | 43 °C for 2 h | • Acute heat stress decreases P4 concentrations in bovine GCs | [23] | |
| Holstein heifers | Incubate with 0, 2, or 100 ng/mL LH or FSH for 96 h at 37, 39, or 41 °C | • Elevated blood progesterone levels in cattle under high-temperature conditions | [6] | |
| 2. Pig | Slovakian white gilts | 41.5 °C for 48 h | • Hormones (IGF-I, leptin, and FSH) are involved in the control of proliferation, apoptosis, and secretory activity of ovarian cells • Heat stress leads to reduced ovarian cell proliferation and apoptosis and excessive ovarian hormone secretion | [24] |
| Prepubertal gilts | Pre-incubation at 41 °C for 24 h followed by three hours of incubation in a humid environment at (41 °C, 5% CO2) | • LPS and heat stress could impair estradiol biosynthesis in GCs via increased HSP70 and indirect inhibition of Smad3 phosphorylation and nuclear translocation | [25] | |
| Female pigs | (35 °C; 20–35% humidity; n = 6) for either 7 (n = 10) or 35 d (n = 12) | • HS increased mRNA and protein abundance and phosphorylated AKT (pAKT) in ovine CYP19a and STAR | [26] | |
| Malabari female goats | Exposure to outdoor summer heat stress for 45 days (6 h per day) | • HS inhibits FSHR expression | [27] | |
| Gilts | / | • Pro-survival factors such as FSH and IGF-1 upregulate the phosphorylation level of BimEL-Ser65 to inhibit apoptosis in GCs | [28] | |
| 3. Mice and rats | Kunming mice | CO2 incubator at 40 °C or 43 °C | • Heat stress reduces E2 and P4 secretion and mRNA expression of steroid-related genes and induces apoptosis in GCs via the mitochondrial pathway | [29] |
| C57BL/6J mouse | / | • E2 enhances Ape1 expression and reduces oxidative DNA damage | [30] | |
| Wistar female rats | 35 °C and 70% relative humidity conditions for 48 h | • Heat stress reduces the number of FSH receptors on the granulosa cells of the antral follicle and the level of estradiol in the follicular fluid, and strongly inhibits gonadotropin receptor levels and aromatase activity in the granulosa cells | [2] | |
| Female rats | 45 °C water bath incubation for 10 min | • Two transcription-dependent responses: heat shock-induced anti-steroid response and HPS induction, which occur simultaneously | [31] | |
| Sprague Dawley rats | 38 ± 0:5 °C, RH55 ± 5% for 2 h/day for at least 90 days | • Heat stress treatment decreased the growth levels of Hsp70 and CORT, decreased the levels of E2 and LH, increased the levels of FSH and Prl, did not differ in GnRH and T4, and significantly decreased INS | [32] | |
| CF-1 mice | / | • The phosphorylation state of 14-3-3 is regulated by P4 through a PKG-dependent pathway | [33] | |
| Wistar rats | / | • P4 inhibits insulin-dependent mitosis in small GCs | [34] | |
| Wistar rats | / | • P4 acts through its receptor to prevent a redistribution and increase in [Ca2+] that may subsequently result in GC apoptosis | [35] | |
| Mice | Seven-day continuous exposure to 40.5 ± 0.2 °C | • Heat exposure decreases serum progesterone levels and ovarian P450scc expression and can induce autophagic flux and affect progesterone synthesis in mice by decreasing P62 levels, enhancing LC3B lipidation and binding, and increasing Atg7 expression. | [36] | |
| CD rats | / | • The inhibitory effect of peroxides on FSH is not due to ATP depletion | [37] | |
| 4. Poultry | 60-wk-old Hy-Line laying hens | 32 °C (25% RH) for three weeks. | • ROS formation was significantly increased by heat exposure in laying hens | [38] |
| Laying hens | 37 °C with 5% CO2 | • SLC5A5 deficiency inhibits proliferation, steroid hormone synthesis, and secretion and promotes apoptosis in chicken GCs | [39] | |
| Shan Ma ducks | 39 °C, 5% CO2 for 6 h | • The expression of proliferative genes required for the transition from G1 to S phase is inhibited Heat stress inhibits GC estradiol synthesis by downregulating CYP11A1 and CYP19A1 gene expression | [40] | |
| Hy-Line laying hens | 32 °C, 50%RHh for 14 days | • Heat stress enhanced P4 secretion by increasing the expression of StAR, CYP11A1, and 3β-HSD and inhibited estradiol synthesis by suppressing the expression of FSHR and CYP19A1, and upregulation of HSP70 was also observed | [41] | |
| Single-comb White Leghorn hens of the Babcock B 300 | / | • Activin A significantly increased FSHR expression in follicular granulosa cells | [42] | |
| Female Japanese quail of a laying strain | 34 °C for 4 h per day (12:00 to 16:00) for 10 consecutive days | • The expression level of 17β-HSD under heat stress was unchanged in the ovary, but significantly increased in the adrenal gland (p < 0.05) • The effect of heat stress on the ovaries of female quail is mediated by adrenal function | [43] |
| Cellular Regulatory Processes | Types/Strains | Heat Stress Environmental Settings | Main Findings | References |
|---|---|---|---|---|
| 1. Cell proliferation and cycle transition under heat stress invasion | Slovakian white gilts | 41.5 °C | • HS significantly reduced the expression of PCNA and cell cycle protein B1 in GCs, and the addition of FSH alleviated this effect | [24] |
| Bovine | Heat treatment 24 and 48 h | • Reduction in PCNA expression in GCs was observed after treatment with HS | [8] | |
| Holstein cattle | 2 h at 40 °C in humidified air with 5% carbon dioxide. | • Cell cycle arrest in GO/G1 and G2/M phases | [55] | |
| Sheep | 5% CO2 in humidified air at 43 °C for 2 h for 12 days. | • Colony formation rate (CFE) of sheep GCs was significantly reduced under heat stress | [56] | |
| Rats | / | • FSH promotes granulosa cell proliferation by increasing cell cycle protein D2 mRNA expression and through an Akt-dependent pathway | [57] | |
| Mouse | 40, 42, 44, and 46 °C for 20 min at 60% relative humidity | • Low-temperature heat stress (e.g., 40 and 42 °C) significantly induced PCNA orchestration, whereas high-temperature heat stress (e.g., 44 and 46 °C) significantly inhibited PCNA expression | [58] | |
| Shan Ma ducks | 41 and 43 °C for 24 h | • In ducks, during heat stress, GC is blocked at the G1 phase and the expression of proliferation genes is inhibited | [40] | |
| 2. Mitochondria damaged by heat stress: kinetic imbalance and autophagy | Cows | 39.8 and 41.0 °C for 6 and 12 h. | • Heat stress impairs mitochondria and their autophagic processes, leading to an accumulation of damaged mitochondria in the cell | [60] |
| Dairy cow mammary epithelial cells | 42 °C for 2 h | • Mfn1/2 and Opa-1 are downregulated and Drp1 and Fis-1 are upregulated following heat stress | [61] | |
| Duck | 43 °C treatment (0, 60, 90, 120 and 150 min) | • Interfering with PINK1 levels significantly reduced apoptosis levels, but interfering with Drp1 levels did not change significantly | [7] | |
| Porcine | 41.5 °C for 1 h | • Short-term heat stress increases the expression level of HSP72 in porcine oocytes, induces phosphorylation of SIRT1 and AMP-activated protein kinase, and induces mitochondrial degradation and generation | [62] | |
| 3. Heat stress affects the expression of the heat shock protein family in HSPs | ZCK mice | 43 °C at 50% humidity for 0.5 h; 1.5 h per day (12:00–13:30) at 39 °C with 50% humidity for 1, 3 or 6 weeks. | • Small HSP genes increase rapidly after heat treatment and are co-expressed by HSP40, HSP70, and HSP90 family members | [63] |
| Slovakian white gilts | 41.5 °C for 2 days | • IGF-1, leptin, and FSH prevent stress-induced changes in HSPs • HS reduces the accumulation of HSP70 in granulosa cells | [9] | |
| Bovine | Heat treatment 24 and 48 h | • HSP70 protein accumulation increased at 24 and 48 h after heat stress treatment but there was no significant difference in levels between the two and no significant change in mRNA levels • HSP90 expression was significantly upregulated at 24 h after treatment | [8] | |
| Bovine | 42 °C for 24 h | • EVs produced by HS-exposed granulosa cells were enriched in HSP90 and SOD1, and HSP70 and GRP94 were also highly expressed, allowing recipient cells to be robust to subsequent HS | [64] | |
| Camel | After incubation at 45 °C for 2 h or 20 h, place at 38 °C | • Acute heat stress induces the expression of HSPA4, HSPA1B protein, and DNA damage binding protein 1 • Chronic heat stress significantly upregulates the levels of HSP70A1B, HSP90, and HSP90B1, but HSP70 protein levels significantly increase during recovery | [65] | |
| 4. Heat stress triggers the FAS/FASL extrinsic apoptotic pathway | Mature sows | / | • Upregulation of FasL and Fas mRNA and protein levels in granulosa cells during follicular atresia • Fas is transferred from the cytoplasm to the cell membrane region at the same time as the granular cells begin to undergo apoptosis | [10] |
| Mode-K cells derived from C3H/HeJ mice | / | • ROS-induced expression of Fas and Fas-L | [66] | |
| Mice | / | • Fas and p53 are involved in apoptosis in mouse testicular germ cells | [67] | |
| Laying hens | 35–37 °C for 5 days | • HS triggers follicular cell apoptosis by activating the FasL/Fas and TNF-α systems. | [45] | |
| 5. Heat stress triggers ER endoplasmic reticulum stress | Bovine | Heat treatment 24 and 48 h | • The mRNA expression of GRP78 and GRP94 was significantly increased in bovine GCs, and a significant accumulation of GRP78 protein was observed • Bovine GCs can respond rapidly to heat stress by activating GRP94 expression | [8] |
| Human OS cell line | / | • High temperatures can upregulate the expression of GRP78 and GRP94 genes in cells, accompanied by changes in calcium levels in cell membranes and an increase in the expression and activity of calpain | [68] | |
| Goat | / | • Transcriptional upregulation of GRP78 in GCs during follicular atresia in goats | [69] | |
| Mice | Incubate at 40 °C for 24 h in 5% CO2 | • HS can induce apoptosis of mGCs through the ER stress pathway, and boron can alleviate this to a certain extent | [70] | |
| Mice | Incubate at 39 °C for 0, 6, 12, 24, and 48 h in 5% CO2 | • Selenium is protective against chronic HS-induced apoptosis by inhibiting the ER stress pathway | [71] | |
| 6. Heat stress activates the Nrf2/HO-1 pathway | Bovine | Heat treatment 24 and 48 h | • Nrf2 mRNA expression was elevated after 24 h of HS treatment and then normalized after 48 h | [8] |
| Bovine | / | • Oxidative stimulation activates the expression of Nrf2-associated genes in GCs and represses miRNAs targeting Nrf2, and overexpression of these miRNAs leads to downregulation of Nrf2 expression | [72] | |
| Bovine | 39 and 40 °C for 2 h | • HO-1 expression is significantly increased by heat stress • Silencing of HO-1 increases apoptosis • HO-1 inhibits apoptosis | [73] | |
| Bovine | 40 °C and 42 °C for 2 h | • HO-1 reduces reactive oxygen species production and activates antioxidant responses | [74] | |
| Porcine aortic endothelial cells | / | • Ferritin induction in endothelial cells under oxidative stress induced by H2O2 occurs concomitantly with HO-1 induction | [75] | |
| Mice | / | • The synergistic effect of ferritin heavy chain and HO-1 is one of the key reasons for the potent cytoprotective capacity of HO-1 | [76] |
| Research Fields | Types/Strains | Heat Stress Environmental Settings | Main Findings | References |
|---|---|---|---|---|
| 1. Heat stress and ferroptosis | Human | / | • Elevated lipid peroxide levels induce ferroptosis | [93] |
| Porcine Sertoli cells | 5% CO2 incubator at 44 °C for 30 min | • Expression of GPX4 and ferritin is inhibited under heat stress Increased protein levels of intracellular transferrin receptor 1 (TFR1) | [94] | |
| 2. Ferroptosis and granulocytes | Pig | / | • Within early atretic follicles, there is a significant decrease in the expression of transferrin, a significant increase in the expression of iron chaperonin, and abnormalities in glutathione metabolism | [97] |
| 3. Interaction mechanism | Human | / | • HO-1 acts to detoxify hemoglobin to biliverdin, a process that leads to the release of labile iron | [98] |
| Hamster fibroblast (HA-1) cells | / | • Low levels (<5-fold) of HO-1 expression were cytoprotective, and high levels (>15-fold) of HO-1 expression were associated with significant oxygen cytotoxicity | [99] | |
| Mice | / | • Iron-overloaded follicular fluid induces granulosa cell iron metamorphosis and releases granulosa cell exosomes containing aberrant miRNAs | [11] | |
| Mouse | / | • Mitochondrial targeting of HO-1 reduces cytochrome C oxidase activity, induces ROS production, and leads to an increase in mitochondrial fission preprotein Drp1 and autophagy marker LC-3 in mitochondria | [103] | |
| Hepatocellular carcinoma cells | / | • Activation of the p62-Keap1-NRF2 pathway protects against ferroptosis in hepatocellular carcinoma cells | [100] | |
| Bovine mammary epithelial cells | / | • Interference with Nrf2 in bovine mammary epithelial cells reveals decreased intracellular protein expression of GPX4, SLC7A11, and ferritin heavy chain 1 (FTH1) | [102] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Wu, J.; Wen, Y.; Wu, X.; Bao, H.; Wang, M.; Kang, K. Advances in the Effects of Heat Stress on Ovarian Granulosa Cells: Unveiling Novel Ferroptosis Pathways. Vet. Sci. 2024, 11, 464. https://doi.org/10.3390/vetsci11100464
Zhu Z, Wu J, Wen Y, Wu X, Bao H, Wang M, Kang K. Advances in the Effects of Heat Stress on Ovarian Granulosa Cells: Unveiling Novel Ferroptosis Pathways. Veterinary Sciences. 2024; 11(10):464. https://doi.org/10.3390/vetsci11100464
Chicago/Turabian StyleZhu, Zhen, Jiang Wu, Yuguo Wen, Xiaocheng Wu, Huimingda Bao, Min Wang, and Kai Kang. 2024. "Advances in the Effects of Heat Stress on Ovarian Granulosa Cells: Unveiling Novel Ferroptosis Pathways" Veterinary Sciences 11, no. 10: 464. https://doi.org/10.3390/vetsci11100464