Abstract
Pythium-induced damping-off of cucumber is a major constraint to cucumber production in different parts of the world. Although chemical fungicides are used for managing this disease, they have many drawbacks to the environment. The ability of the antagonistic fungi isolated from the rhizosphere and endosphere of Dactyloctenium robecchii and Moraea sisyrinchium in the control of soilborne pathogen Pythium aphanidermatum was inspected. Native Trichoderma isolates, Trichoderma ghanense and Trichoderma citrinoviride, were isolated from plant stem and soil samples collected from Al-Seeb, Oman. Using a dual culture technique, the antagonistic activity of the fungal isolates against P. aphanidermatum was examined in vitro. Among Trichoderma isolates, T. ghanense was more efficient in restraining the mycelial growth of P. aphanidermatum, causing an inhibition percentage of 44.6%. Further, T. citrinoviride induced significantly lower cessation of P. aphanidermatum mycelial growth (31.3%). Microscopic and electrolyte leakage inspection of the pathogen mycelia depicted extreme morphological malformations in their mycelium, which can be attributed to the antifungal metabolites of antagonists. Greenhouse studies demonstrated the effectivity of T. ghanense in controlling Pythium damping-off of cucumber plants, where the number of surviving plants was over 90% when the biocontrol agents were used compared to 0 in the control plants. Furthermore, treatment of the plants with the antagonists promoted growth characteristics of plants compared to uninoculated plants. This included improvements in shoot and root lengths, leaf length and width, and dry weight. These findings suggest that T. ghanense and T. citrinoviride can be developed as alternatives to synthetic chemical fungicides to manage soilborne pathogens of cucumber. This research is also the first to clarify the biocontrol ability of T. citrinoviride and T. ghanense against cucumber damping-off caused by P. aphanidermatum.
1. Introduction
Cucumber (Cucumis sativus L., family Cucurbitaceae), one of the most important vegetable crops, is grown extensively worldwide [1]. Cucumber fruits are popular around the world for their crispy texture and special flavor. They are an excellent source of vitamins, proteins, minerals, and antioxidants and thus deliver various health benefits to the human body [2,3]. They possess anti-diabetic, lipid-lowering, and antioxidant properties. Moreover, they have Cucurbitacin B and C that prevent tumor growth and guard the liver against inflammation [3,4,5]. The leaves, stems, and roots of the cucumber are commonly employed in Chinese traditional medicine as anti-diarrheal, detoxicant, and anti-gonorrheal medicines [6]. Moreover, the seeds of cucumbers have anti-inflammatory, anti-fever, and antidiabetic effects [7]. Cucumber is one of the three most widely cultivated vegetable crops in Oman. Despite that, this crop is greatly vulnerable to a vast range of diseases leading to economic losses [8].
Plant diseases are believed to be the primary cause of global food production decline, which is estimated to be between 10% and 40%, as they directly contribute to the destruction of natural resources in agriculture [9]. Fungi are the most varied group of plant disease agents and are responsible for one-third of all crop losses each year among soil-borne pathogens, which affect both the natural and production ecosystems [10,11,12]. The dispersion of several phytopathogenic fungi, including Pythium, Fusarium, and Rhizoctonia, owing to the changes initiated in farming, has deleterious impacts on crops of economic importance [13,14,15,16]. Some of these pathogens, such as Pythium species, are widely distributed in different parts of the world. They have been shown to result in losses that can reach 75% of cucumbers in greenhouses. The Pythium species and other pathogens are particularly difficult to control since each crop may be susceptible to a variety of pathogen species, and they frequently persist in soil for several years through the production of long-lasting spores called oospores [17]. It has been shown that oospores can persist in the soil for up to 4 years in the absence of a host plant [18]. Even when using the normal techniques, they are typically difficult to control.
Even though fungicides are useful for treating a variety of diseases, frequent use of fungicides can have harmful effects on the environment and the survival of helpful rhizosphere microbes [19,20]. Indecisive utilization of fungicides forces pathogens to go through genetic mutation, which ultimately results in the development of fungicide-resistant biotypes [21,22,23]. Fungicide resistance has been reported against several chemical fungicides, especially following their frequent use. Botrytis cinerea, for example, a major cause of pre- and post-harvest losses in fruit and vegetable production, can adapt to fungicide treatments through mutations, resulting in resistance and loss of fungicide efficacy [24]. The demand for pesticide-free food among consumers and the rising cost of pesticides have sparked the hunt for alternatives to these goods. Therefore, it is essential to develop accessible and environmentally safe non-chemical techniques for the control of plant diseases. Due to this, artificial fungicides have been replaced by biological control, and utilizing antagonistic microorganisms to manage diseases has resulted in significant success [25,26,27,28].
The efficacy and practicability of non-pathogenic antagonistic fungi and bacteria have been the subject of several studies over the past several decades to commercialize them as biocontrol agents [29,30]. Through conducted studies, a plethora of fungal strains have been created as biocontrol agents, which effectively manage soil-borne diseases of important crops [29,31]. Among the non-pathogenic fungi, Trichoderma spp. is a usual saprophytic filamentous fungus that interacts with the soil ecosystem and foliar environment [10]. It plays a role as a biocontrol agent towards varied phytopathogens, causing numerous diseases in crop plants [32,33,34]. It can establish itself in different pathosystems, has the lowest effect on the soil balance, and does not destroy beneficial organisms that help control diseases [23]. It controls phytopathogens by different mechanisms comprising the generation of antifungal compounds, competition for space and nutrients in the rhizosphere, mycoparasitism, and the improvement of host mechanisms for defense [23,35,36].
Numerous Trichoderma species have undergone extensive research and are employed for the biological control of a wide range of aerial-borne and soil-borne pathogens, namely Fusarium oxysporum, Rhizoctonia solani, Pythium aphanidermatum, Alternaria alternata and Macrophomina phaseolina [33,37,38,39,40,41,42,43,44,45,46]. The management capacity varies among Trichoderma species and depends on the target disease [34]. The investigation for potential biocontrol agents against cucumber soil-borne disease in Oman resulted in the contemplation of the Trichoderma species, which are native to different regions in Oman. Moraea sisyrinchium and Dactyloctenium robecchii are native plant species to the Arabian Peninsula. M. sisyrinchium and D. robecchii were considered the most interesting species, especially in terms of their flexibility to grow in various habitats such as hills, mountain floodplains, and saline lands [47,48]. We hypothesized that antagonistic fungi isolated from these plants could enhance pathogen tolerance in cucumber plants under unfavorable conditions. The following were the study goals: (1) isolation of Trichoderma species from the rhizosphere and endosphere of Dactyloctenium robecchii and Moraea sisyrinchium, (2) evaluating the native Trichoderma species’ antagonistic effects against P. aphanidermatum, and (3) investigating how soil treatment with certain Trichoderma species affects cucumber growth and Pythium damping-off control.
Research into these areas will help widen the alternative options available for managing Pythium-induced diseases in vegetable crops, especially with biocontrol isolates from the environment. In addition, it will help reduce the reliance on chemical fungicides.
2. Materials and Methods
2.1. Materials Utilized
The study was undertaken in the Plant Pathology Laboratory at Sultan Qaboos University. Chemicals utilized to make various reagents were acquired from Sigma Aldrich, St. Louis, MO, USA. Media were purchased from Oxoid (Thermo Fisher Scientific, Waltham, MA, USA). All reactions were accomplished utilizing sterilized distilled water. The primers for molecular identification were purchased from Macrogen Inc. (Seoul, Republic of Korea). Cucumber seeds were obtained from a local supplier.
2.2. Collection Site and Isolation of the Fungal Pathogen
The fungal pathogen, Pythium aphanidermatum (isolate no. ON113866), was part of a fungal culture collection of the Department of Plant Sciences (Sultan Qaboos University, Muscat, Oman). It was obtained from our previous studies [49]. On petri dishes containing potato dextrose agar (PDA) medium (Thermo Fisher Scientific, Waltham, MA, USA), the fungal pathogen’s pure culture was transferred and incubated at a temperature of 25 ± 2 °C for further experiments.
2.3. Plant Sampling and Isolation of Rhizospheric and Endophytic Fungi
Dactyloctenium robecchii (family Poaceae) and Moraea sisyrinchium (family Iridaceae) plants were collected from Al-Seeb (23.6473° N, 58.1458° E), Muscat, Oman. In sterile zipper bags, samples of healthy, symptom-free plants and their rhizospheric soils were transferred to the lab using a mobile refrigeration chamber (4 °C). The identification of plants was carried out at the botany herbarium, located at the Life Science Unit, College of Science, Sultan Qaboos University.
The plant leaves, stems, and roots were all similarly dissected (5 mm) by a sterile scalpel shortly after being thoroughly cleaned with tap water. The dissected fragments were surface sterilized as described by Khalil et al. [50]. The dissected fragments were then briefly immersed in sterile distilled water (60 s), 70% ethanol (60 s), 2.5% sodium hypochlorite (4 min), and 70% ethanol (30 s), and eventually rinsed in sterile distilled water three times. The sterilized fragments were then dried employing sterile filter paper. To validate surface sterilization efficiency, 100 µL of the final rise water was dispersed on a potato dextrose agar (PDA) medium and incubated at 25 ± 2 °C for one week.
The sterilized fragments of the leaves, stems, and roots (5 fragments/plate) were placed on the PDA plate supplemented with rifampicin (10 mg/L) and ampicillin (200 mg/L) to prevent bacterial growth and incubated at 25 ± 2 °C. Every day, plates were checked seeking any evidence of fungal growth. Fungal isolates were sub-cultured onto a PDA medium.
Bulk soils from D. robecchii and M. sisyrinchium roots were flaked away and the soils that remained attached to the roots were considered rhizospheric soils [51]. The method described by Dey et al. [52] was used to isolate fungi from rhizospheric soil. Corresponding to the rhizospheric soil, root specimens were agitated at high speed for 90 s to collect rhizospheric soil. Thereafter, 1 g of collected soil was taken and diluted in 10 mL of sterile distilled water and labeled as stock solution. Subsequently, the serial dilution method was used to dilute the stock solution and minimize the fungi in the soil in each dilution. Eventually, 0.1 mL of the solution in each soil dilution was added to the prepared PDA medium, which was then incubated for seven days at 25 °C.
For further examination, pure cultures of the acquired fungus were commonly maintained on PDA slants at 4 °C and categorized according to their cultural appearance. Additionally, the isolated fungi were identified using morphological and molecular analysis.
2.4. Characterization of Fungal Isolates
DNA Extraction, Amplification, and Sequencing
The isolated fungi were identified at the genus level based on cultural and morphological characteristics, and microscopical attributes as stated previously [53,54]. For molecular identification, genomic DNA was extracted from fungal isolates following the protocol of Al-Sadi et al. [55]. The extracted DNA’s quality and quantity were determined by a NanoDropTM 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). DNA was preserved at −20 °C until use. Utilizing the primer combination ITS1 (5′-TCC GTA GGT GAA CCT GCG G-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′); EF1-526F (GTC GTY GTY ATY GGH CAY GT) and EF-jR (GCR TGY TCN CGR GTY TGN CCR TC), we amplified the nuclear ribosomal DNA’s internal transcribed spacer (ITS1-5.8S-ITS2 = ITS) and a portion of the translation elongation factor 1 alpha (EF1α) region, respectively [56]. Polymerase chain reactions were performed using the PuRe Taq Ready-To-Go™ PCR beads (Cytiva, Marlborough, MA, USA), with 1 µL of each primer (10 µM/µL), DNA (1 µL), and 25 µL sterile distilled water (25 µL) following the protocol of Lakhani et al. [57]. On a 1% agarose gel (140 min, 80 V, 400 mA), PCR-amplified products were evaluated for their supposed size and visualized with ethidium bromide under UV illumination. PCR-amplified products were purified and sequenced with the same primers from Macrogen Inc. (Seoul, Republic of Korea). The resulting sequences were deposited in GenBank. The type sequences of closely related species of Trichoderma were selected for phylogenetic analysis. Sequences were aligned using the MAFFT algorithm [58]. The maximum likelihood phylogenetic method was used for phylogenetic analysis using RAxMLHPC2 v. 8.2.4. The resulting phylogenetic tree was visualized in FigTree 1.4.2 [59] and annotated using Adobe Illustrator CC2019.
2.5. Efficacy of Rhizospheric and Endophytic Fungi as Biocontrol Agents against P. aphanidermatum
2.5.1. In Vitro Evaluation of the Fungal Isolates’ Antifungal Activity towards Fungal Pathogens
Two Trichoderma isolates were chosen among the obtained rhizospheric and endophytic fungal isolates and their antagonistic effects against P. aphanidermatum were examined via a dual culture approach [60]. The PDA medium-filled petri plates with a 90 mm diameter were used for this test. The pathogen agar–mycelium cylinder (P. aphanidermatum, 6 mm diameter), cut from the edge of an actively growing fungal colony, was placed on the edge of the plate. The non-pathogen agar–mycelium disc (6 mm diameter) was inoculated on the same day and placed on the opposite edge. Three replicates were arranged for each isolate and plates inoculated only with pathogen were considered as a control. After the inoculation, the plates were kept at 25 °C until the leading edge of the fungus on the control plate reached the edge of the plate. By comparing the percentage of mycelium growth inhibition to the control, the antagonistic activity was shown [61]. A scanning electron microscope (SEM; JEOL JSM-5600, Tokyo, Japan) was also used to see how the selected fungal isolates affected the hyphal morphology of P. aphanidermatum. A protocol defined by Heckman et al. [62] was pursued for the SEM sample preparation.
2.5.2. Effect of Trichoderma Isolates on Extracellular Conductivity of P. aphanidermatum
The impact of Trichoderma isolates on electrolyte leakage of P. aphanidermatum was studied as described previously [63,64]. 5 mm mycelial discs of Trichoderma isolates (3-day-old, previously grown on PDA) were transferred into 50 mL sterile conical flasks containing 25 mL potato dextrose broth (PDB, Sigma Aldrich, MO, USA) and, subsequently, incubated at 27 °C in a shaker for 7 days. The resulting suspension was centrifuged at 10,000 rpm for 10 min to separate the fungal mycelium and culture filtrate. The culture filtrate was then filtered using Minisart filters with 0.2 μm pore size for the purity of the culture filtrates. The obtained supernatant was preserved at 4 °C. Moreover, Pythium isolate was grown in potato dextrose broth (PDB) for 3 days in an incubator at 27 °C. After that, sterile filter paper was used to collect the Pythium mycelium, which was then thoroughly cleaned with sterile double-distilled water. Following that, 3 mg of the collected mycelium of Pythium was added to the glass vial containing the Trichoderma culture supernatant (20 mL). Eventually, the mycelial suspension was centrifuged at 10,000 rpm for 10 min to collect the supernatant, first immediately after the addition of mycelium (0 h), second after 5 h, and third after 24 h of treatment. A conductivity meter (Mettler Toledo FiveGo™, Herisau, Switzerland) was used to measure conductivity.
2.6. Biocontrol Potential of Trichoderma Isolates against P. aphanidermatum on Cucumber in Pots
Fungal Inoculum Preparation, Experimental Design, and Plant Growth Condition
A pot experiment was outlined using pots (15 cm × 15 cm) containing autoclaved-sterilized potting mixture (300 g/pot) (Bulrush Horticulture, Ireland, UK) to test the efficacy of Trichoderma in promoting plant growth. The potting soil was infested with P. aphanidermatum grown on millet grains at 1% (w/w). 10 mL spore suspensions (1 × 108 spore/mL) of the selected Trichoderma spp. isolates were added to the pots, followed by watering the pots for 3 days before sowing [65]. After disinfecting the cucumber seeds (Cucumis sativus cv. Azza) (AgriMax Company, Barka, Oman) in 1.5% sodium hypochlorite for two minutes and 70% ethanol for one minute, they underwent three rinses in sterile distilled water. The disinfected seeds were then planted in pots (5 seeds/pot) with 5 replicates for each treatment. The growth-promoting experiment comprised 4 treatments: non-infested soil (control), soil treated with P. aphanidermatum only, soil treated with Trichoderma ghanense only, and soil treated with Trichoderma citrinoviride only. The experimental design for the seedling survival experiment is described in Table 1. The pots were incubated in a greenhouse and grown under natural daylight, 25 °C/20 °C (day/night) temperature and 80% relative humidity. Seed germination and disease percentage of the seedlings were examined after 10 days.

Table 1.
Experimental work plan for the seedling survival experiment.
Multiple plant growth characteristics were assessed to ascertain the consequences of each treatment on the plant growth attributes. These metrics, which were measured after ten days, included shoot length, root length, leaf area (length/width), and total plant fresh weight.
2.7. Statistical Analysis
The obtained data (stated as the mean ± SD) were exposed to statistical analysis. R software (version 4.0.3) and Microsoft Excel 2021 were used to perform statistical analysis. At a 5% probability level (p < 0.05), the least significant difference test (LSD) and analysis of variance were used to compare the means. The graphs were created with the aid of the GraphPad Prism program (version 6.01, San Diego, CA, USA).
3. Results
3.1. Identification of the Antagonistic Fungi
The fungal isolates were identified as Trichoderma ghanense (T1) and Trichoderma citrinoviride (T2) using phylogenetic analysis of the combined ITS–EF1α sequence data. Both these species are members of the Harzianum clade. The phylogenetic tree depicted in Figure 1 consists of types of sequences of Trichoderma species, presented as species name, followed by isolate or strain number, ITS accession, and EF1α accession, respectively (Table 2). The Omani isolates are highlighted in green.
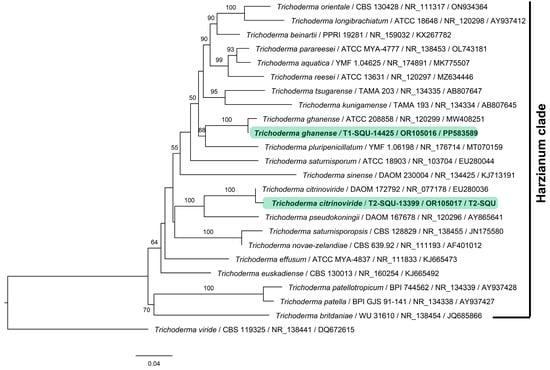
Figure 1.
Maximum likelihood phylogenetic tree based on combined ITS–EF1α dataset obtained from 24 type specimens of Trichoderma species of Harzianum clade. Each leaf in the tree is presented as species name followed by isolate or strain number, ITS accession, and EF1α accession, separated by a/line, respectively. The representative species Trichoderma citrinoviride (T2-SQU-13399) and T. ghanense (T1-SQU-14425) are highlighted in green; bootstrap support values above 50% are consistently significant and are shown.
Table 2.
Identification of potential strains of Trichoderma sp. associated with Dactyloctenium robecchii and Moraea sisyrinchium and its antagonistic activity (+indicates a positive response). EF1α sequences were submitted to GenBank.
3.2. Antifungal Activity of the Trichoderma Isolates against Fungal Pathogen (P. aphanidermatum)
3.2.1. Dual Culture and Scanning Electron Microscope Assays
The Trichoderma isolates were tested for their ability to suppress P. aphanidermatum growth in vitro (Figure 2, Table 2). Among the Trichoderma isolates, T. ghanense (T1) was found to be more efficient in restraining the mycelial growth of P. aphanidermatum compared to T. citrinoviride (T2). T. ghanense caused inhibition percentages of 44.6% for P. aphanidermatum (Figure 3). However, T. citrinoviride (T2) caused a considerably lower suppression of P. aphanidermatum mycelial growth (31.3%). Scanning electron microscopy (SEM) was used to confirm that T. ghanense (T1) and T. citrinoviride (T2) isolates had an impact on the hyphal morphology of P. aphanidermatum (Figure 2). The scanning electron microscope revealed that Trichoderma isolates T1 and T2 significantly altered the hyphal morphology of P. aphanidermatum (Figure 2). When compared to the control, the majority of the observed hyphal patterns were wrinkled and ruptured in both treatments.
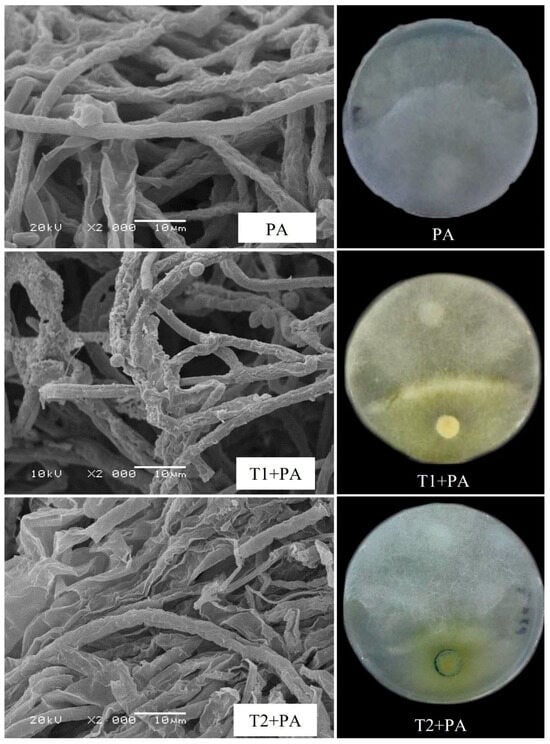
Figure 2.
Effect of Trichoderma ghanense (T1) and Trichoderma citrinoviride (T2) on Pythium aphanidermatum (PA) mycelial morphology depicted using dual culture assay and scanning electron microscope (SEM). Distorted mycelial structure, wrinkled, ruptured, or shrunken patterns in T1 + PA and T2 + PA plates. Typical hypha patterns were visible in the control (PA).
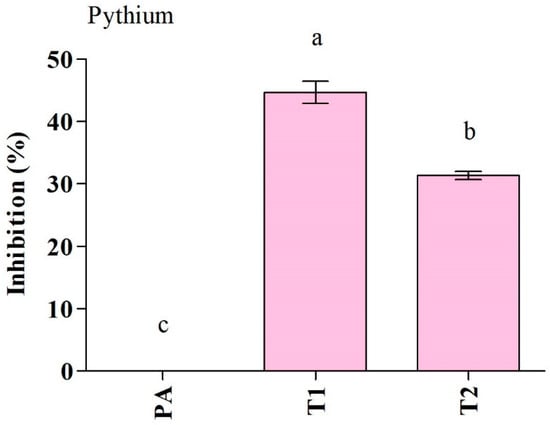
Figure 3.
Antagonistic activity of Trichoderma ghanense (T1) and Trichoderma citrinoviride (T2) and their effect on the inhibition of Pythium aphanidermatum (PA) growth. Values show the means ± standard error (n = 3) and significant differences at p < 0.05 are indicated by different lowercase letters above the columns.
3.2.2. Extracellular Conductivity
Exposure of P. aphanidermatum to the culture filtrate of T. ghanense and T. citrinoviride for 5 and 24 h led to enhanced levels of extracellular conductivity in comparison to the control, which depicted cellular electrolyte leakage from P. aphanidermatum owing to losing cell wall/cell membrane integrity. Among the antagonists, T. ghanense depicted the maximum discharge of electrolytes from the fungal mycelium (Figure 4).
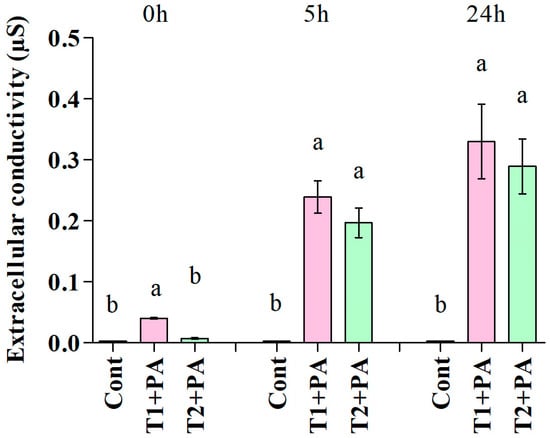
Figure 4.
Effect of antifungal metabolites (Trichoderma ghanense, T1; Trichoderma citrinoviride, T2) on extracellular conductivity in the suspension of the pathogen (Pythium aphanidermatum, PA) after 0, 5, and 24 h of treatment. Values show the means ± standard error (n = 3) and significant differences at p < 0.05 are indicated by different lowercase letters above the columns.
3.3. Effect of Trichoderma Isolates on Cucumber Growth and Damping-Off Disease
The impact of the Trichoderma isolates on cucumber seedlings was investigated (Figure 5 and Figure 6). The unfavorable consequences of P. aphanidermatum invasion resulted in the mitigation of seed germination and growth parameters; namely, shoot length, root length, leaf area (length/width), and total fresh weight of the cucumber plants, as compared to non-diseased, un-treated cucumber plants. However, disease severity was considerably decreased in antagonist-treated plants. Trichoderma-treated cucumber plants emerged higher and stronger than plants treated with pathogen only, and the seeds had 100% germination in control and Trichoderma-treated plants. In comparison to plants exposed to P. aphanidermatum, Trichoderma treatment increased shoot length, root length, leaf area (length/width), and total fresh weight of the inoculated plants. When compared to similar untreated sick plants, the shoot length increased by 100% in the T. ghanense treatment group and by 100% in the T. citrinoviride treatment group (p < 0.05). Similarly, in the T. ghanense and T. citrinoviride treatment groups, compared to the diseased plants, the overall plant fresh weight of Trichoderma-treated plants increased by 100% (Figure 6). As depicted in Figure 7, the survival percentage of diseased plants inoculated by T. ghanense and T. citrinoviride improved by 98.92% and 98.75%, respectively, in contrast to the untreated diseased plants.

Figure 5.
Growth-promoting impact of Trichoderma isolates on cucumber plants treated with fungal pathogens over ten days (A,B). Treatment: control (Cont), Trichoderma ghanense (T1), Trichoderma citrinoviride (T2), Pythium aphanidermatum (PA).
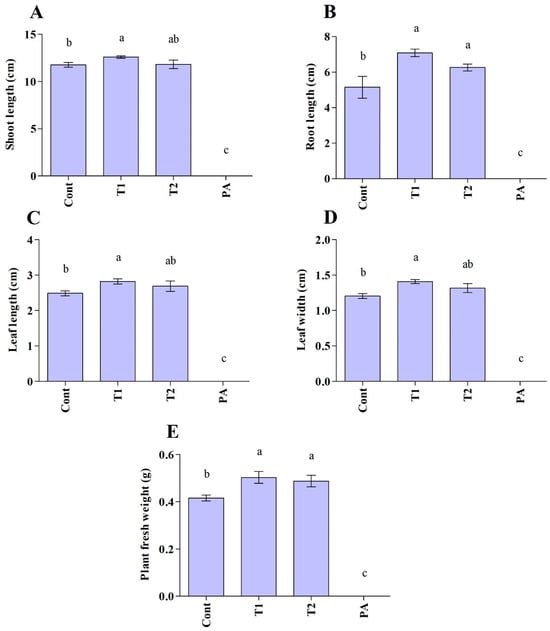
Figure 6.
Plant growth-promoting effect of Trichoderma isolates on the cucumber plants under normal and disease conditions after ten days (A–E). Treatment groups: control (Cont), Trichoderma ghanense (T1), Trichoderma citrinoviride (T2), Pythium aphanidermatum (PA). Values show the means ± standard error (n = 5) and significant differences at p < 0.05 are indicated by different lowercase letters above the columns.
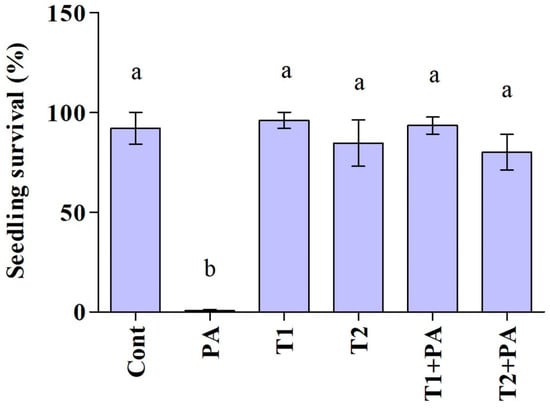
Figure 7.
Biocontrol potential (survival percentage) of the Trichoderma isolates on the Pythium aphanidermatum causal agent of cucumber damping-off in pot experiment over ten days. Treatment groups: control (Cont), Pythium aphanidermatum (PA), Trichoderma ghanense (T1), Trichoderma citrinoviride (T2), Trichoderma ghanense (T1) + Pythium aphanidermatum (PA), and Trichoderma citrinoviride (T2) + Pythium aphanidermatum (PA). Values show the means ± standard error (n = 5) and significant differences at p < 0.05 are indicated by different lowercase letters above the columns.
4. Discussion
Plant diseases caused by pathogenic microorganisms induce enormous losses that are reckoned to range between 13 and 22% [66,67]. Given that chemical control has many side effects [68], the use of biological control is promising [69]. Cucumber damping-off is an extremely severe disease resulting in serious losses to cucumber production. Managing cucumber damping-off through the use of chemical fungicides has both benefits and drawbacks. Controlling damping-off via biological methods offers a safe alternative [70]. Trichoderma is a biocontrol fungus that is found all over the world. Trichoderma has enormous practical value and potential in the realm of biological plant disease control [9]. Trichoderma has been studied for its ability to control plant diseases all over the world. T. viride and T. harzianum have been shown to be effective in inhibiting around 30 plant pathogenic fungi, including Fusarium, Pythium and Rhizoctonia. Rhizoctonia solani, Pythium aphanidermatum, Sclerotinia sclerotiorum, and Colletotrichum spp., which are all controlled by the Trichoderma spp. [71]. A study by Paulitz et al. [72] demonstrated the efficiency of T. harzianum in reducing Pythium damping-off of cucumbers in greenhouse experiments. Trichoderma is a good substitute for chemical pesticides that could be more trustworthy and ecologically safe, along with economically endurable [73]. The current study identified T. ghanense and T. citrinoviride as endophytic and rhizospheric fungi in D. robecchii and M. sisyrinchium, respectively. T. ghanense and T. citrinoviride were identified at the species level based on sequences of the ITS, which was useful in distinguishing these species from other Trichoderma species. These two Trichoderma species were not isolated previously from D. robecchii and M. sisyrinchium.
In this study, T. ghanense and T. citrinoviride reduced the growth of P. aphanidermatum in vitro. A study conducted by Park et al. [74] depicted the effective mycoparasitic activity of T. citrinoviride via dual culture assays against ginseng pathogens, including Rhizoctonia solani, Botrytis cinerea, and Pythium spp. Their results showed that the cell wall degrading enzyme, endoglucanase, contributed to the antagonistic activity of T. citrinoviride toward pathogens. Moreover, a study by Khadka et al. [75] revealed reduced mycelial growth of Rhizoctonia solani by T. ghanense.
Mycoparasitism is a key biological control mechanism for Trichoderma. Trichoderma can parasitize 18 different Pythium, Phytophthora, Rhizoctonia, and Peronospora genera. They can infiltrate or injure the mycelium, causing pathogen cells to enlarge, distort, and rupture [76,77]. In addition to dual culture assays, scanning electron microscopy observations of pathogenic fungal mycelia from the border of inhibition zones exhibited serious structural modifications in mycelial structure, which demonstrated that Trichoderma metabolites possibly attack the cell wall/cell membrane. According to Halifu et al. [78], Trichoderma spp. can permeate into a pathogen body by producing enzymes such as chitinase, cellulase, and xylanase, and grow thoroughly within mycelium via demolishing the cell wall.
The use of culture filtrates from T. ghanense and T. citrinoviride led to the leakage of electrolytes from the P. aphanidermatum mycelium. Extracellular conductivity of T. ghanense and T. citrinoviride supernatants that were treated with P. aphanidermatum mycelium was escalated with the proceeding of time in comparison to 0 min. Antifungal compounds mediated the action. Under in vivo conditions, the interaction’s influence was noticeable for diminished fungal diseases. Zhang et al. [79] indicated that the culture filtrate of Trichoderma spp. negatively influenced the growth of Sclerotinia sclerotiorum.
Trichoderma is a saprophytic fungus with rapid mycelial development and high environmental adaptation. It can prevent pathogenic fungi from invading plant roots. It can also rapidly compete for the nutrients essential for disease fungal growth, resulting in nutritional shortage and preventing pathogen fungi growth and reproduction [80,81]. In the bioassay experiment, it was found that T. ghanense and T. citrinoviride were very efficient in diminishing the severity of the damping-off disease of cucumber as induced by P. aphanidermatum. With or without Pythium infection, Trichoderma inoculation led to higher growth and better health in the cucumber plants. Compared to plants without Trichoderma, inoculated plants maintained greater height, root length, leaf area, and plant/root weight. In addition, Trichoderma isolates suppressed Pythium-induced damping-off of cucumber. Various mechanisms have been suggested to clarify the beneficial effects of Trichoderma spp. on plants. One feature is connected to the broad range of metabolites that they generate. It has been confirmed that these metabolites not only interfere with the growth of the pathogen but also enhance disease endurance by inducing the defense responses in the host plant. Moreover, these metabolites have been shown to improve the growth of plants, which enables plants to tolerate infection by compensating for the lost tissues due to pathogen invasion [78,82,83].
5. Conclusions
The study focused on investigating the suppressive effects of Trichoderma species against Pythium aphanidermatum. Along with enhancing cucumber growth, Trichoderma ghanense and T. citrinoviride were found to suppress Pythium damping-off in cucumber. To our knowledge, this study is the first to investigate the suppressive effects of T. ghanense and T. citrinoviride on the damping-off disease of cucumber caused by P. aphanidermatum. The results of this study indicate that T. ghanense and T. citrinoviride are beneficial for C. sativus under disease attack and can be used in the management of Pythium diseases. Overall, employing these Trichoderma species to manage Pythium damping-off is not only associated with retarding fungal growth, but it can also reduce the usage of fungicides, and be used as potential biocontrol agents in organic farms.
Author Contributions
A.M.A.-S., M.N.A.-Y., D.A.-M., M.A.-K. and R.V. planned the study. B.K.A.-S., D.A.-M. and M.A.-K. conducted the main experiments. A.M.A.-S., B.K.A.-S., S.H. and E.A.K. performed the analysis and wrote the first draft of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Oman Animal and Plant Genetic Resources Center (Mawarid) (AGREEMENT NO: MoHERI/BFP/MoHERI-OAPGRC/01/2020) and Sultan Qaboos University (project # CR/AGR/CROP/21/02).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
All procedures were conducted according to the guidelines. The collected samples from Al-Seeb required no permission.
Data Availability Statement
The ITS sequence data generated during the current study are available in GenBank under the accession number OR105016 and OR105017.
Acknowledgments
Thanks to Sultan Qaboos University for providing equipped laboratories for the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jia, H.; Wang, H. Introductory Chapter: Studies on Cucumber. In Cucumber Economic Values and Its Cultivation and Breeding; Wang, H., Ed.; IntechOpen: Rijeka, Croatia, 2021; Chapter 1. [Google Scholar]
- Qian, C.; Ren, N.; Wang, J.; Xu, Q.; Chen, X.; Qi, X. Effects of exogenous application of CPPU, NAA and GA4+ 7 on parthenocarpy and fruit quality in cucumber (Cucumis sativus L.). Food Chem. 2018, 243, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Qing, Z.; Shi, Y.; Han, L.; Li, P.; Zha, Z.; Liu, C.; Liu, X.; Huang, P.; Liu, Y.; Tang, Q. Identification of seven undescribed cucurbitacins in Cucumis sativus (cucumber) and their cytotoxic activity. Phytochemistry 2022, 197, 113123. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Zhang, L.; Zhou, Y.; Fang, Q. Cucurbitacin B alleviates cerebral ischemia/reperfusion injury by inhibiting NLRP3 inflammasome-mediated inflammation and reducing oxidative stress. Biosci. Biotechnol. Biochem. 2022, 86, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Involvement of auxin in growth and stress response of cucumber. Veg. Res. 2022, 2, 13. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Nema, N.K.; Maity, N.; Sarkar, B.K. Phytochemical and therapeutic potential of cucumber. Fitoterapia 2013, 84, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Mandey, J.; Wolayan, F.; Pontoh, C.; Kowel, Y. Effect of orally administrated of cucumber (Cucumis sativus L.) seed juice on the performance and carcass parameters of broiler chickens. IOP Conf. Ser. Earth Environ. Sci. 2020, 492, 012025. [Google Scholar] [CrossRef]
- Ni, L.; Punja, Z.K. Management of fungal diseases on cucumber (Cucumis sativus L.) and tomato (Solanum lycopersicum L.) crops in greenhouses using Bacillus subtilis. In Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Islam, M.T., Rahman, M.M., Pandey, P., Boehme, M.H., Haesaert, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–28. [Google Scholar]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Redda, E.T.; Ma, J.; Mei, J.; Li, M.; Wu, B.; Jiang, X. Antagonistic potential of different isolates of Trichoderma against Fusarium oxysporum, Rhizoctonia solani, and Botrytis cinerea. Eur. J. Exp. Biol. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef]
- Peng, Y.; Li, S.J.; Yan, J.; Tang, Y.; Cheng, J.P.; Gao, A.J.; Yao, X.; Ruan, J.J.; Xu, B.L. Research Progress on Phytopathogenic Fungi and Their Role as Biocontrol Agents. Front. Microbiol. 2021, 12, 670135. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L.; Toxicology, C. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Kuang, Y.; Splivallo, R.; Chatterjee, P.; Karlovsky, P. Interactions among filamentous fungi Aspergillus niger, Fusarium verticillioides and Clonostachys rosea: Fungal biomass, diversity of secreted metabolites and fumonisin production. BMC Microbiol. 2016, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Khan, S.U.; Khan, W.U.; Saleh, T.A.; Khan, M.H.U.; Ullah, S.; Ali, A.; Ikram, M. Antagonist effects of strains of Bacillus spp. against Rhizoctonia solani for their protection against several plant diseases: Alternatives to chemical pesticides. C. R. Biol. 2019, 342, 124–135. [Google Scholar] [CrossRef]
- Ma, M.; Taylor, P.W.; Chen, D.; Vaghefi, N.; He, J.-Z. Major Soilborne Pathogens of Field Processing Tomatoes and Management Strategies. Microorganism 2023, 11, 263. [Google Scholar] [CrossRef] [PubMed]
- Babadoost, M. Oomycete Diseases of Cucurbits: History, Significance, and Management. In Horticultural Reviews; Jules, J., Ed.; 2016; Volume 44, pp. 279–314. [Google Scholar]
- Rosslenbroich, H.-J.; Stuebler, D. Botrytis cinerea—History of chemical control and novel fungicides for its management. Crop Prot. 2000, 19, 557–561. [Google Scholar] [CrossRef]
- Leroux, P. Chemical control of Botrytis cinerea and its resistance to chemical fungicides. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Meszka, B.; Broniarek-Niemiec, A.; Bielenin, A. The status of dodine resistance of Venturia inaequalis populations in Poland. Phytopathol. Pol. 2008, 47, 57–61. [Google Scholar]
- Matson, M.E.; Small, I.M.; Fry, W.E.; Judelson, H.S. Metalaxyl resistance in Phytophthora infestans: Assessing role of RPA190 gene and diversity within clonal lineages. Phytopathology 2015, 105, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A.J.P. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Muthukumar, A.; Eswaran, A.; Sanjeevkumas, K. Exploitation of Trichoderma species on the growth of Pythium aphanidermatum in chilli. Braz. J. Microbiol. 2011, 42, 1598–1607. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Collinge, D.B.; Jensen, D.F.; Rabiey, M.; Sarrocco, S.; Shaw, M.W.; Shaw, R.H. Biological control of plant diseases—What has been achieved and what is the direction? Plant Pathol. 2022, 71, 1024–1047. [Google Scholar] [CrossRef]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Sederoff, H.; Chiang, V.L.; Borriss, R. Microbial interactions within multiple-strain biological control agents impact soil-borne plant disease. Front. Microbiol. 2020, 11, 585404. [Google Scholar] [CrossRef]
- Subedi, P.; Gattoni, K.; Liu, W.; Lawrence, K.S.; Park, S.-W. Current utility of plant growth-promoting rhizobacteria as biological control agents towards plant-parasitic nematodes. Plants 2020, 9, 1167. [Google Scholar] [CrossRef]
- Savita, A.; Giri, B.; Prasad, R.; Wu, Q.; Varma, A. Fungi as biological control agents. In Biofertilizers for Sustainable Agriculture and Environment; Springer: Cham, Switzerland, 2019; Volume 55. [Google Scholar]
- Singh, A.; Srivastava, M.; Kumar, V.; Sharma, A.; Pandey, S.; Shahid, M. Exploration and interaction of Trichoderma species and their metabolites by confrontation assay against Pythium aphanidermatum. Int. J. Sci. Res. 2014, 3, 44–48. [Google Scholar]
- El_Komy, M.H.; Saleh, A.A.; Eranthodi, A.; Molan, Y.Y. Characterization of novel Trichoderma asperellum isolates to select effective biocontrol agents against tomato Fusarium wilt. Plant Pathol. J. 2015, 31, 50. [Google Scholar] [CrossRef]
- Galarza, L.; Akagi, Y.; Takao, K.; Kim, C.S.; Maekawa, N.; Itai, A.; Peralta, E.; Santos, E.; Kodama, M. Characterization of Trichoderma species isolated in Ecuador and their antagonistic activities against phytopathogenic fungi from Ecuador and Japan. J. Gen. Plant Pathol. 2015, 81, 201–210. [Google Scholar] [CrossRef]
- Woo, S.L.; Lorito, M. Exploiting the interactions between fungal antagonists, pathogens and the plant for biocontrol. In Novel Biotechnologies for Biocontrol Agent Enhancement and Management; Springer: Berlin/Heidelberg, Germany, 2007; pp. 107–130. [Google Scholar]
- Hoyos-Carvajal, L.; Duque, G.; Orduz, S. Antagonismo in vitro de Trichoderma spp. sobre aislamientos de Sclerotinia spp. y Rhizoctonia spp. Rev. Colomb. Cienc. Hortícolas 2008, 2, 76–86. [Google Scholar] [CrossRef]
- Hjeljord, L.G.; Tronsmo, A. Effect of germination initiation on competitive capacity of Trichoderma atroviride P1 conidia. J. Phytopathol. 2003, 93, 1593–1598. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Le, H.; Black, L.; Sikora, R. Evaluation of Trichoderma spp. for biocontrol of tomato sudden caused by Pythium aphanidermatum following flooding in tropical hot season. Commun. Agric. Appl. Biol. Sci. 2003, 68, 463–474. [Google Scholar] [PubMed]
- Watanabe, S.; Kumakura, K.; Kato, H.; Iyozumi, H.; Togawa, M.; Nagayama, K. Identification of Trichoderma SKT-1, a biological control agent against seedborne pathogens of rice. J. Gen. Plant Pathol. 2005, 71, 351–356. [Google Scholar] [CrossRef]
- Karthikeyan, M.; Radhika, K.; Mathiyazhagan, S.; Bhaskaran, R.; Samiyappan, R.; Velazhahan, R. Induction of phenolics and defense-related enzymes in coconut (Cocos nucifera L.) roots treated with biocontrol agents. Braz. J. Plant Physiol. 2006, 18, 367–377. [Google Scholar] [CrossRef]
- Sharma, K.; Mishra, A.K.; Misra, R.S. Morphological, biochemical and molecular characterization of Trichoderma harzianum isolates for their efficacy as biocontrol agents. J. Phytopathol. 2009, 157, 51–56. [Google Scholar] [CrossRef]
- Gveroska, B.; Ziberoski, J. Trichoderma harzianum as a biocontrol agent against Alternaria alternata on tobacco. ATI-Appl. Technol. Innov. 2012, 7, 67–76. [Google Scholar] [CrossRef]
- Kumar, M.; Ashraf, S. Role of Trichoderma spp. as a Biocontrol Agent of Fungal Plant Pathogens. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 497–506. [Google Scholar]
- Swehla, A.; Pandey, A.K.; Nair, R.M. Bioactivity of Trichoderma harzianum isolates against the fungal root rot pathogens with special reference to Macrophomina phaseolina causing dry root rot of mungbean. Indian Phytopathol. 2020, 73, 787–792. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, W.-Y. Trichoderma brevicrassum strain TC967 with capacities of diminishing cucumber disease caused by Rhizoctonia solani and promoting plant growth. Biol. Control 2020, 142, 104151. [Google Scholar] [CrossRef]
- Sánchez-Montesinos, B.; Santos, M.; Moreno-Gavíra, A.; Marín-Rodulfo, T.; Gea, F.J.; Diánez, F. Biological control of fungal diseases by Trichoderma aggressivum f. europaeum and its compatibility with fungicides. J. Fungi 2021, 7, 598. [Google Scholar] [CrossRef]
- Ferjani, H.; Bencheikh, Z. Diversity analysis of spontaneous populations of Moraea sisyrunchium (Iridacea) in different bioclimates of Tunisia. J. Anim. Plant Sci. 2022, 52, 9495–9502. [Google Scholar]
- Quattrocchi, U. CRC World Dictionary of Grasses: Common Names, Scientific Names, Eponyms, Synonyms, and Etymology-3 Volume set; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Al-Daghari, D.S.S.; Al-Sadi, A.M.; Al-Mahmooli, I.H.; Janke, R.; Velazhahan, R. Biological Control Efficacy of Indigenous Antagonistic Bacteria Isolated from the Rhizosphere of Cabbage Grown in Biofumigated Soil against Pythium aphanidermatum Damping-Off of Cucumber. Agriculture 2023, 13, 626. [Google Scholar] [CrossRef]
- Khalil, A.M.A.; Hassan, S.E.-D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.-D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and characterization of fungal endophytes isolated from medicinal plant Ephedra pachyclada as plant growth-promoting. J. Biomol. 2021, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Sarkar, A.; Dutta, S. Characterisation of plant growth promoting traits and stress tolerant properties of Aspergillus terreus (MCC 1819) isolated from the rhizosphere of Aloe barbadensis Mill. and its application. J. Pharmacogn. Phytochem. 2019, 8, 2303–2310. [Google Scholar]
- Cai, F.; Dou, K.; Wang, P.; Chenthamara, K.; Chen, J.; Druzhinina, I.S. The Current State of Trichoderma Taxonomy and Species Identification. In Advances in Trichoderma Biology for Agricultural Applications; Amaresan, N., Sankaranarayanan, A., Dwivedi, M.K., Druzhinina, I.S., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 3–35. [Google Scholar]
- Mazrou, Y.S.; Makhlouf, A.H.; Elseehy, M.M.; Awad, M.F.; Hassan, M.M. Antagonistic activity and molecular characterization of biological control agent Trichoderma harzianum from Saudi Arabia. Egypt. J. Biol. Pest Control 2020, 30, 4. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Al-Masoodi, R.S.; Al-Ismaili, M.; Al-Mahmooli, I.H. Population structure and development of resistance to hymexazol among Fusarium solani populations from date palm, citrus and cucumber. J. Phytopathol. 2015, 163, 947–955. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38—Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Lakhani, H.N.; Vakharia, D.N.; Hassan, M.M.; Eissa, R.A. Fingerprinting and molecular comparison among two parental strains of Trichoderma spp. and their corresponding fusants produced by protoplast fusion. Biotechnol. Biotechnol. Equip. 2016, 30, 1065–1074. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary 623 Biology, University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 3 January 2022).
- Ji, S.H.; Gururani, M.A.; Chun, S.-C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Kunova, A.; Bonaldi, M.; Saracchi, M.; Pizzatti, C.; Chen, X.; Cortesi, P. Selection of Streptomyces against soil borne fungal pathogens by a standardized dual culture assay and evaluation of their effects on seed germination and plant growth. BMC Microbiol. 2016, 16, 272. [Google Scholar] [CrossRef]
- Heckman, C.; Kanagasundaram, S.; Cayer, M.; Paige, J. Preparation of Cultured Cells for Scanning Electron Microscope. Protoc. Exch. 2007. [Google Scholar] [CrossRef]
- Manhas, R.K.; Kaur, T. Biocontrol potential of Streptomyces hydrogenans strain DH16 toward Alternaria brassicicola to control damping off and black leaf spot of Raphanus sativus. Front. Plant Sci. 2016, 7, 1869. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Abo-Elyousr, K.A.M.; Mousa, M.A.A.; Saad, M.M. Use of Trichoderma culture filtrates as a sustainable approach to mitigate early blight disease of tomato and their influence on plant biomarkers and antioxidants production. Front. Plant Sci. 2023, 14, 1192818. [Google Scholar] [CrossRef] [PubMed]
- Benito, E.P.; Ten Have, A.; van’t Klooster, J.W.; van Kan, J.A. Fungal and plant gene expression during synchronized infection of tomato leaves by Botrytis cinerea. Eur. J. Plant Pathol. 1998, 104, 207–220. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-P.; Pan, Z.-C.; Yang, L.-N.; Burdon, J.J.; Friberg, H.; Sui, Q.-j.; Zhan, J. Optimizing plant disease management in agricultural ecosystems through rational in-crop diversification. Front. Plant Sci. 2021, 12, 767209. [Google Scholar] [CrossRef]
- Pathak, V.M.; Verma, V.K.; Rawat, B.S.; Kaur, B.; Babu, N.; Sharma, A.; Dewali, S.; Yadav, M.; Kumari, R.; Singh, S. Current status of pesticide effects on environment, human health and it’s eco-friendly management as bioremediation: A comprehensive review. Front. Microbiol. 2022, 13, 962619. [Google Scholar] [CrossRef] [PubMed]
- He, D.-C.; He, M.-H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef]
- Mahmoud, A.F.; Abdalla, O.A.; Protection, P. Biological control of fungi associated with damping-off and root rot disease of cucumber (Cucumis sativus L.). Arch. Phytopathol. 2021, 54, 870–885. [Google Scholar] [CrossRef]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Front. Microbiol. 2023, 14, 1160551. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Ahmad, J.S.; Baker, R. Integration of Pythium nunn and Trichoderma harzianum isolate T-95 for the biological control of Pythium damping-off of cucumber. Plant Soil 1990, 121, 243–250. [Google Scholar] [CrossRef]
- Huang, X.; Cui, H.; Yang, L.; Lan, T.; Zhang, J.; Cai, Z. The microbial changes during the biological control of cucumber damping-off disease using biocontrol agents and reductive soil disinfestation. Biocontrol 2017, 62, 97–109. [Google Scholar] [CrossRef]
- Park, Y.-H.; Mishra, R.C.; Yoon, S.; Kim, H.; Park, C.; Seo, S.-T.; Bae, H. Endophytic Trichoderma citrinoviride isolated from mountain-cultivated ginseng (Panax ginseng) has great potential as a biocontrol agent against ginseng pathogens. J. Ginseng Res. 2019, 43, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Khadka, R.B.; Rotondo, F.; Miller, S.A. The pursuit in Nepal of native Trichoderma spp. for plant disease biocontrol. PhytoFrontiers 2022, 2, 242–256. [Google Scholar] [CrossRef]
- Risoli, S.; Cotrozzi, L.; Sarrocco, S.; Nuzzaci, M.; Pellegrini, E.; Vitti, A. Trichoderma-induced resistance to Botrytis cinerea in Solanum species: A Meta-Analysis. Plants 2022, 11, 180. [Google Scholar] [CrossRef]
- Aswani, R.; Thomas, R.; Radhakrishnan, E. Induction of plant defense response by endophytic microorganisms. In Biocontrol Mechanisms of Endophytic Microorganisms; Elsevier: Amsterdam, The Netherlands, 2022; pp. 89–115. [Google Scholar]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Zhang, F.; Ge, H.; Zhang, F.; Guo, N.; Wang, Y.; Chen, L.; Ji, X.; Li, C. Biocontrol potential of Trichoderma harzianum isolate T-aloe against Sclerotinia sclerotiorum in soybean. Plant Physiol. Biochem. 2016, 100, 64–74. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R.; Liang, X. Inhibitory mechanism of Trichoderma virens ZT05 on Rhizoctonia solani. Plants 2020, 9, 912. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yan, L.; Zhang, M.; Chang, X.; Zhu, D.; Wei, D.; Naeem, M.; Song, C.; Wu, X.; Liu, T. Changes in the density and composition of rhizosphere pathogenic Fusarium and beneficial Trichoderma contributing to reduced root rot of intercropped soybean. Pathogens 2022, 11, 478. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Commun. 2012, 7, 1934578X1200701133. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).