
Investigation of the Microbial Diversity in the Oryza sativa Cultivation Environment and Artificial Transplantation of Microorganisms to Improve Sustainable Mycobiota

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
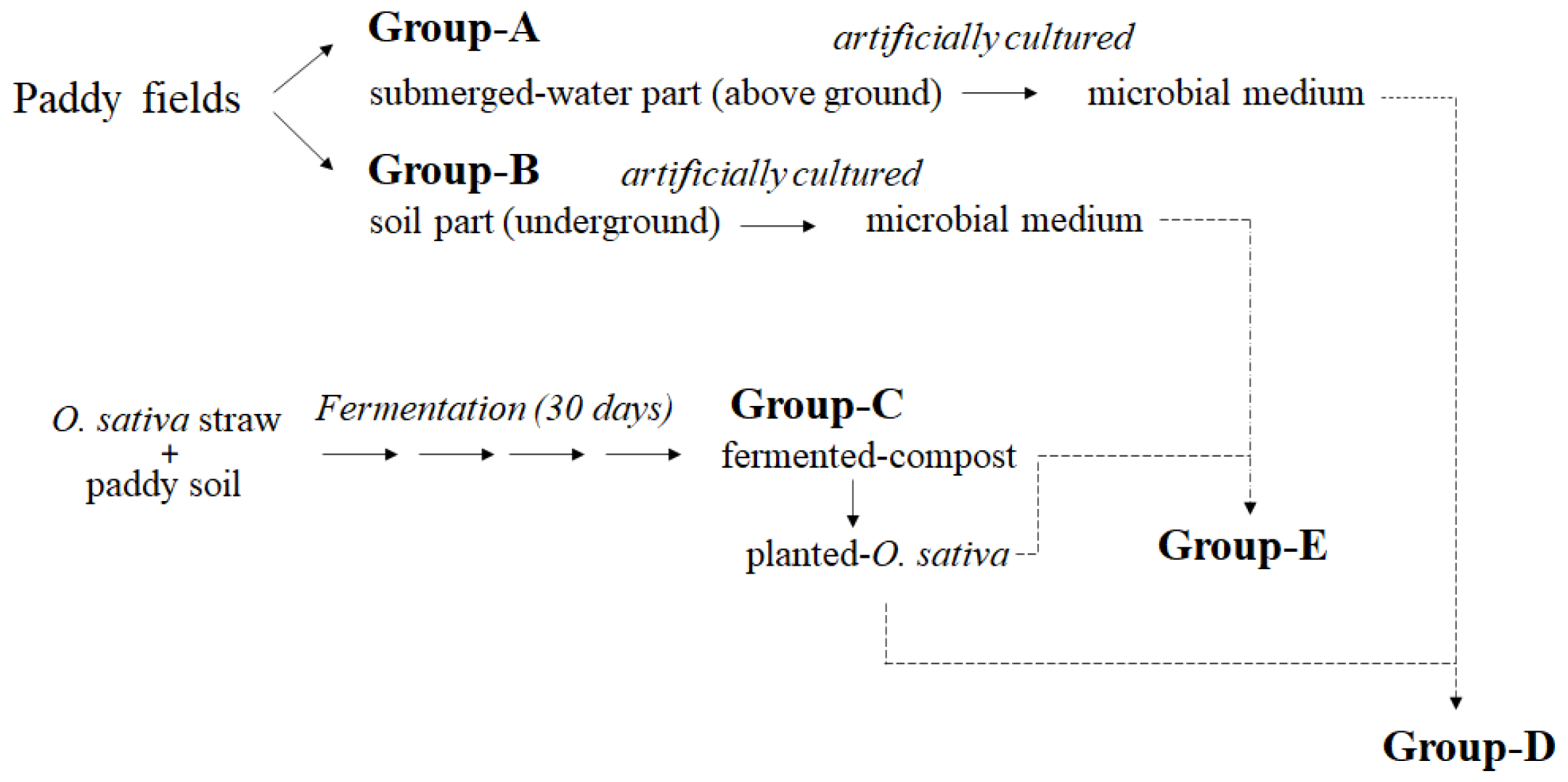
2.1. Compost Preparation
2.2. Microbial Cultivation
2.3. Microbial Transplantation into Planted O. sativa
2.4. Assay for Mycobiota with Next-Generation Sequencing
2.5. Assay for Phytohormones
2.6. Statistical Analysis
3. Results
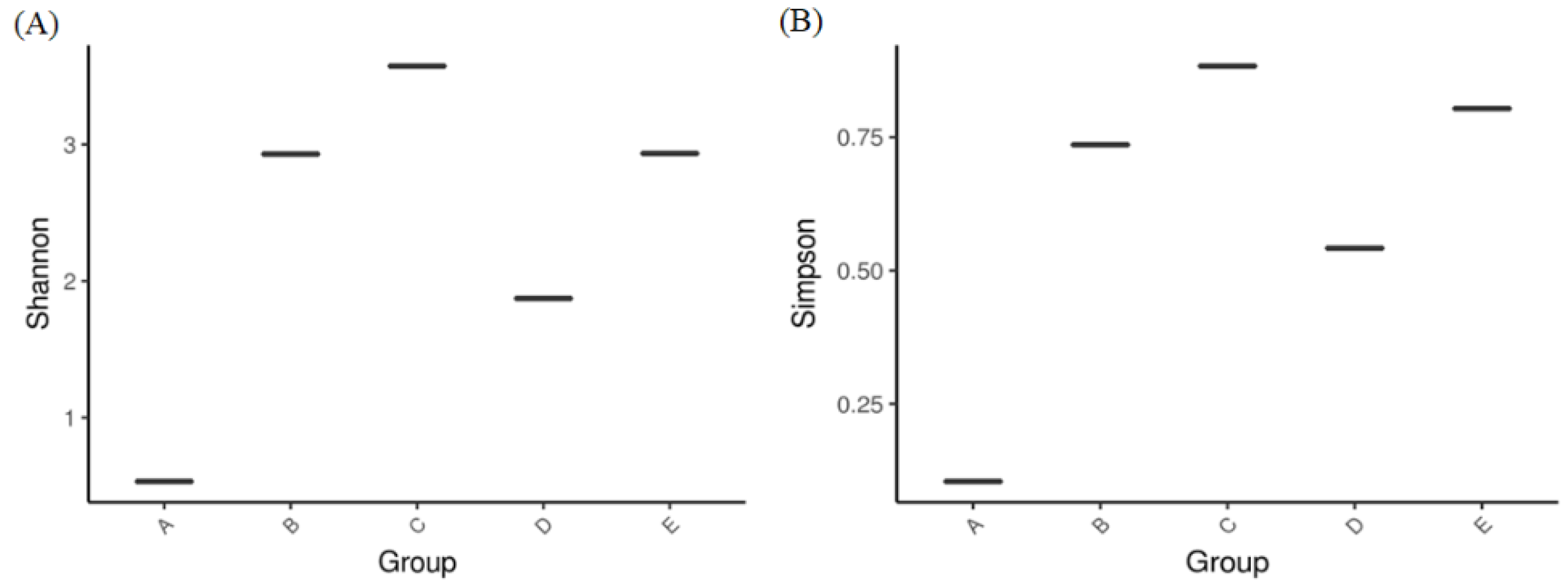
3.1. The Investigation of Mycobiota in Different Samples
3.2. Root Development of O. sativa Treated with Microbial Transplantation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zheng, H.; Tang, F.; Lin, Y.; Xu, Z.; Xie, Z.; Tian, J. Solid-state anaerobic digestion of rice straw pretreated with swine manure digested effluent. J. Clean. Prod. 2022, 348, 131252. [Google Scholar] [CrossRef]
- Cañon, C.; Sanchez, N.; Cobo, M. Sustainable production of ethyl levulinate by levulinic acid esterification obtained from Colombian rice straw. J. Clean. Prod. 2022, 377, 134276. [Google Scholar] [CrossRef]
- Nazar, M.; Xu, L.; Ullah, M.W.; Moradian, J.M.; Wang, Y.; Sethuparhy, S.; Iqbal, B.; Nawaz, M.Z.; Zhu, D. Biological delignification of rice straw using laccase from Bacillus ligniniphilus L1 for bioethanol production: A clean approach for agro-biomass utilization. J. Clean. Prod. 2022, 360, 132171. [Google Scholar] [CrossRef]
- Molaverdi, M.; Mirmohamadsadeghi, S.; Karimi, K.; Aghbashlo, M.; Tabatabaei, M. Efficient ethanol production from rice straw through cellulose restructuring and high solids loading fermentation by Mucor indicus. J. Clean. Prod. 2022, 339, 130702. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Chen, X.; Xiong, L.; Guo, H.; Yao, S.; Wang, M.; Chen, X.; Huang, C. Anaerobic digestion of rice straw pretreatment liquor without detoxification for continuous biogas production using a 100 L internal circulation reactor. J. Clean. Prod. 2022, 349, 131450. [Google Scholar] [CrossRef]
- Singh, N.K.; Singh, R. Hydrogen recovery cascade from pretreated rice straw and its fermentative residuals using step-up potential-based sulfate reducing bacteria-bioelectrochemical system. J. Clean. Prod. 2023, 408, 137167. [Google Scholar] [CrossRef]
- Walder, F.; Schmid, M.W.; Riedo, J.; Valzano-Held, A.Y.; Banerjee, S.; Buchi, L.; Bucheli, T.D.; van der Heijden, M.G.A. Soil microbiome signatures are associated with pesticide residues in arable landscapes. Soil Biol. Biochem. 2022, 174, 108830. [Google Scholar] [CrossRef]
- Rahman, N.S.N.A.; Hamid, N.W.A.; Nadarajah, K. Effects of abiotic stress on soil microbiome. Int. J. Mol. Sci. 2021, 22, 9036. [Google Scholar] [CrossRef]
- Song, Y.; Li, X.; Yao, S.; Yang, X.; Jiang, X. Correlations between soil metabolomics and bacterial community structures in the pepper rhizosphere under plastic greenhouse cultivation. Sci. Total Environ. 2020, 728, 138439. [Google Scholar] [CrossRef] [PubMed]
- Treesubsuntorn, C.; Dhurakit, P.; Khaksar, G.; Thiravetyan, P. Effect of microorganisms on reducing cadmium uptake and toxicity in rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2018, 25, 25690–25701. [Google Scholar] [CrossRef]
- Korenblum, E.; Aharoni, A. Phytobiome metabolism: Beneficial soil microbes steer crop plants secondary metabolism. Pest Manag. Sci. 2019, 75, 2378–2384. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd-Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef] [PubMed]
- Toan, N.S.; Nguyen, T.D.P.; Thu, T.T.N.; Lim, D.T.; Dong, P.D.; Gia, N.T.; Khoo, K.S.; Chew, K.W.; Show, P.L. Soil mineralization as effects of plant growth promoting bacteria isolated from microalgae in wastewater and rice straw application in a long-term paddy rice in Central Viet Nam. Environ. Technol. Innov. 2021, 24, 101982. [Google Scholar] [CrossRef]
- Liang, Y.; Jiang, Y.; Wang, F.; Wen, C.; Deng, Y.; Xue, K.; Qin, Y.; Yang, Y.; Wu, L.; Zhou, J.; et al. Long-term soil transplant regulating climate change with latitude significantly alters microbial temporal turnover. ISME J. 2015, 9, 2561–2572. [Google Scholar] [CrossRef] [PubMed]
- Khaskheli, M.; Wu, L.; Chen, G.; Chen, L.; Hussain, S.; Song, D.; Liu, S.; Feng, G. Isolation and characterization of root-associated bacterial endophytes and their biocontrol potential against major fungal phytopathogens of rice (Oryza sativa L.). Pathogens 2020, 9, 172. [Google Scholar] [PubMed]
- Amprayn, K.O.; Rose, M.T.; Kecskes, M.; Pereg, L.; Nguyen, H.T.; Kennedy, I.R. Plant growth promoting characteristics of soil yeast (Candida tropicalis HY) and its effectiveness for promoting rice growth. Appl. Soil Ecol. 2012, 61, 295–299. [Google Scholar] [CrossRef]
- Lu, H.; Qi, X.; Ur Rahman, S.; Qiao, D.; Li, P.; Han, Y.; Zhao, Z. Rice physiological response with Bacillus subtilis and Saccharomyces cerevisiae inoculation into soil under reclaimed water-fresh water combined irrigation. Water 2021, 13, 773. [Google Scholar] [CrossRef]
- Yergeau, E.; Bell, T.H.; Champagne, J.; Maynard, C.; Tardif, S.; Tremblay, J.; Greer, C.W. Transplanting soil microbiomes leads to lasting effects on Willow growth, but not on the rhizosphere microbiome. Front. Microbiol. 2015, 6, 1436. [Google Scholar] [CrossRef] [PubMed]
- Roca-Pérez, L.; Martínez, C.; Marcilla, P.; Boluda, R. Composting rice straw with sewage sludge and compost effects on the soil-plant system. Chemosphere 2009, 75, 781–787. [Google Scholar] [CrossRef]
- Foong, S.Y.; Chan, Y.H.; Chin, B.L.F.; Lock, S.S.M.; Yee, C.Y.; Yiin, C.L.; Peng, W.; Lam, S.S. Production of biochar from rice straw and its application for wastewater remediation—An overview. Bioresor. Technol. 2022, 360, 127588. [Google Scholar] [CrossRef]
- Qin, X.; Li, Y.; Wang, H.; Liu, C.; Li, J.; Wan, Y.; Gao, Q.; Fan, F.; Liao, Y. Long-term effect of biochar application on yield-scaled greenhouse gas emissions in a rice paddy cropping system: A four-year case study in south China. Sci. Total Environ. 2016, 569–570, 1390–1401. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.; Patel, N.; Patel, A.; Desai, C.; Sharma, S.; Dedania, S.; Rudaykiya, D.M.; Vishwakarma, A.S.; Vahora, S.; Narra, M. Rapid decomposition of rice straw by application of novel microbial consortium and study its microbial community dynamics. World J. Microbiol. Biotechnol. 2022, 38, 212. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Lu, Y.; Wan, Y.; Wang, B.; Nie, J.; Li, Y.; Liao, Y. Rice straw application improves yield marginally and increases carbon footprint of double cropping paddy rice (Oryza sativa L.). Field Crop. Res. 2023, 291, 108796. [Google Scholar] [CrossRef]
- Hannula, S.E.; Morriën, E. Will fungi solve the carbon dilemma? Geoderma 2022, 413, 115767. [Google Scholar] [CrossRef]
- Charters, M.D.; Sait, S.; Field, K.J. Aphid herbivory drives asymmetry in carbon for nutrient exchange between plants and an arbuscular mycorrhizal fungus. Curr. Biol. 2020, 30, 1801–1808. [Google Scholar] [CrossRef] [PubMed]
- Averill, C.; Anthony, M.A.; Baldrian, P.; Finkbeiner, F.; van den Hoogen, J.; Kiers, T.; Kohout, P.; Hirt, E.; Smithe, G.R.; Crowther, T.W. Defending earth’s terrestrial microbiome. Nat. Microbiol. 2022, 7, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Yu, S.; Cai, Z.; Liu, J.; Wang, T.; Qi, B.; Wang, Z.; Wang, S.; Yang, A.; Zhu, G.; et al. Inhibition of polycyclic aromatic hydrocarbon (PAHs) release from sediments in an integrated rice and crab coculture system by rice straw biochar. J. Clean. Prod. 2022, 367, 133058. [Google Scholar] [CrossRef]
- Sritongon, N.; Sarin, P.; Theerakulpisut, P.; Riddech, N. The effect of salinity on soil chemical characteristics, enzyme activity and bacterial community composition in rice rhizospheres in Northeastern Thailand. Sci. Rep. 2022, 12, 20360. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Khippal, A.K.; Prajapat, K.; Barman, A.; Singh, G.; Rai, A.K.; Ahlawat, O.P.; Verma, R.P.S.; Kumari, K.; Singh, G. Influence of tillage and residue management practices on productivity, sustainability, and soil biological properties of rice-barley cropping systems in indo-gangetic plain of India. Front. Microbiol. 2023, 14, 1130397. [Google Scholar] [CrossRef]
- Tang, H.; Li, C.; Xiao, X.; Shi, L.; Cheng, K.; Wen, L.; Li, Y. Effects of shor-term manure nitrogen in put on soil microbial community structure and diversity in a double-cropping paddy field of southern China. Sci. Rep. 2020, 10, 13540. [Google Scholar] [CrossRef]
- Haiming, T.; Xiaoping, X.; Chao, L.; Xiaochen, P.; Kaikai, C.; Weiyan, L.; Ke, W. Microbial carbon source utilization in tice rhizosphere and nonrhizosphere soils with short-term manure N input rate in paddy field. Sci. Rep. 2020, 10, 6487. [Google Scholar] [CrossRef]
- Agrahari, R.K.; Singh, P.; Koyama, H.; Panda, S.K. Plant-microbe interactions for sustainable agriculture in the post-genomic era. Curr. Genom. 2020, 21, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Alzandi, A.A.; Naguib, D.M. Effect of yeast application on soil health and root metabolic status of corn seedlings under drought stress. Arch. Microbiol. 2022, 204, 233. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.A.; Delgado-Baquerizo, M.; Duarte, E.; Marigliano, O.; Gorgen, C.; Maestre, F.T.; Eisenhauer, N. Global projections of the soil microbiome in the Anthropocene. Glob. Ecol. Biogeogr. 2021, 30, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Z.; Kowalchuk, G.A.; Xu, Z.; Fu, X.; Kuramae, E.E. Succession of the resident soil microbial community in response to periodic inoculations. Appl. Environ. Microbiol. 2021, 87, e00046-21. [Google Scholar] [CrossRef] [PubMed]
- Kazerooni, E.; Maharachchikumbura, S.; Al-Sadi, A.; Rashid, U.; Kim, I.D.; Kang, S.M.; Lee, I.J. Effects of the rhizosphere fungus Cunninghamella bertholletiae on the Solanum lycopersicum response to diverse abiotic stresses. Int. J. Mol. Sci. 2022, 23, 8909. [Google Scholar] [CrossRef]
- De Oliveira, T.B.; Junior, R.B.; Silva, L.G.; Rosa-Magri, M.M. Rhizosphere yeast Torulaspora globose with plant growth promotion traits and improvement of the development of tomato seedling. s under greenhouse conditions. Afric. J. Agric. Res. 2019, 14, 935–942. [Google Scholar]
- Junker, K.; Chailyan, A.; Hesselbart, A.; Forster, J.; Wendland, J. Multi-omics characterization of the necrotrophic mycoparasite Saccharomycopsis schoenii. PLoS Pathog. 2019, 15, e1007692. [Google Scholar] [CrossRef]
- Giri, R.; Sharma, R.K. Fungal pretreatment of lignocellulosic biomass for the production of plant hormone by Pichia fermentans under submerged conditions. Bioresour. Bioprocess. 2020, 7, 30. [Google Scholar] [CrossRef]
- Manzotti, A.; Bergna, A.; Burow, M.; Jorgensen, H.J.L.; Cernava, T.; Berg, G.; Collinge, D.B.; Jensen, B. Insights into the community structure and lifestyle of the fungal root endophytes of tomato by combining amplicon sequencing and isolation approaches with phytohormone profiling. FEMS Microbiol. Ecol. 2020, 96, fiaa052. [Google Scholar] [CrossRef]
- Ferraz, P.; Cassio, F.; Lucas, C. Potential of yeasts as biocontrol agents of the phytopathogen causing cacao witches’ broom disease: Is microbial warfare a solution? Front. Microbiol. 2019, 10, 1766. [Google Scholar] [CrossRef]
- Carla Baron, N.; de Souza Pollo, A.; Rigobelo, E.C. Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 2020, 8, e9005. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.F.; Sun, P.F.; Lu, H.Y.; Wei, J.Y.; Xiao, H.S.; Fang, W.T.; Cheng, B.Y.; Chou, J.Y. Plant growth-promoting traits of yeasts isolated from the phyllosphere and rhizosphere of Drosera spatulate Lab. Fungal Biol. 2016, 120, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ye, T.; Liu, G.; Xu, X.; Zheng, Y.; Wang, W. Synthesis and bioactivity of indoleacetic acid-carbendazim and its effects on Cylindrocladium parasiticum. Pestic. Biochem. Physiol. 2019, 158, 128–134. [Google Scholar] [CrossRef]
- Nartey, L.K.; Pu, Q.; Zhu, W.; Zhang, S.; Li, J.; Yao, Y.; Hu, X. Antagonistic and plant growth promotion effects of Mucor moelleri, a potential biocontrol agent. Microbiol. Res. 2021, 255, 126922. [Google Scholar] [CrossRef] [PubMed]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef]
- Hermosa, R.; Rubio, M.B.; Cardoza, R.E.; Nicolas, C.; Monte, E.; Gutierrez, S. The contribution of Trichoderma to balancing the costs of plant growth and defense. Int. Microbiol. 2013, 16, 69–80. [Google Scholar]
- Nakayan, P.; Hameed, A.; Singh, S.; Young, L.S.; Hung, M.H.; Young, C.C. Phosphate-solubilizing soil yeast Meyerozyma guilliermondii CC1 improves maize (Zea mays L.) productivity and minimizes requisite chemical fertilization. Plant Soil 2013, 373, 301–315. [Google Scholar] [CrossRef]
- Hillman, E.T.; Li, M.; Hooker, C.A.; Englaender, J.A.; Wheeldon, I.; Solomon, K.V. Hydrolysis of lignocellulose by anaerobic fungi produces free sugars and organic acids for two-stage fine chemical production with Kluyveromyces marxianus. Biotechnol. Prog. 2021, 37, e3172. [Google Scholar] [CrossRef]
- Goh, Y.K.; Marzuki, N.F.; Tuan Pa, T.N.F.; Goh, T.K.; Kee, Z.S.; Goh, Y.K.; Yusof, M.T.; Vujanovic, V.; Goh, K.J. Biocontrol and plant-growth-promoting traits of Talaromyces apiculatus and Clonostachys rosea consortium against Ganoderma basal stem rot disease of oil palm. Microorganisms 2020, 8, 1138. [Google Scholar] [CrossRef]
- Kaur, J.; Anand, V.; Srivastava, S.; Bist, V.; Tripathi, P.; Naseem, M.; Nand, S.; Khare, P.; Srivastava, P.K.; Bisht, S.; et al. Yeast strain Debaryomyces hansenii for amelioration of arsenic stress in rice. Ecotoxicol. Environ. Saf. 2020, 195, 110480. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.K.; Shenoy, B.D.; Gupta, M.; Vaish, A.; Mannan, S.; Singh, N.; Teware, S.K.; Tripathi, R.D. Stimulatory effects of arsenic-tolerant soil fungi on plant growth promotion and soil properties. Microbes Envrion. 2012, 27, 477–482. [Google Scholar] [CrossRef] [PubMed]
- El-Tarabily, K.A.; Sivasithamparam, K. Potential of yeasts as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Mycoscience 2006, 47, 25–35. [Google Scholar] [CrossRef]
- Rao, R.P.; Hunter, A.; Kashpur, O.; Normanly, J. Aberrant synthesis of indole 3-acetic acid in Saccharomyces cerevisiae triggers morphogenic transition, a virulence trait of pathogenic fungi. Genetics 2020, 185, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Rayle, D.L.; Cleland, R.E. The acid growth theory of auxin-induced cell elongation is alive and well. Plant Physiol. 1992, 99, 1271–1274. [Google Scholar] [CrossRef]
- Susilowati, D.N.; Riyanti, E.I.; Setyowati, M.; Nulya, K. Indole-3-acetic acid producing bacteria and its application on the growth of rice. AIP Conf. Proc. 2018, 2002, 020016. [Google Scholar] [CrossRef]
- Liang, B.; Wang, H.; Yang, C.; Wang, L.; Qi, L.; Guo, Z.; Chen, X. Salicylic acid is required for broad-spectrum disease resistance in rice. Int. J. Mol. Sci. 2022, 23, 1354. [Google Scholar] [CrossRef] [PubMed]
- Bowya, T.; Balachandar, D. Harnessing PGPR inoculation through exogenous foliar application of salicylic acid and microbial extracts for improving rice growth. J. Basic Microbiol. 2020, 60, 95–961. [Google Scholar] [CrossRef]
- De Meyer, G.; Capieau, K.; Audenaert, K.; Buchala, A.; Métraux, J.P.; Höfte, M. Nanogram amounts of salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 activate the systemic acquired resistance pathway in bean. Mol. Plant Microb. Interact. 1999, 12, 450–458. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-derived exosomal microRNAs shape the gut microbiota. Cell Host Microbe 2018, 24, 637–652. [Google Scholar] [CrossRef]
- Que, Y.; Huang, D.; Gong, S.; Zhang, X.; Yuan, B.; Xue, M.; Shi, W.; Zeng, F.; Liu, M.; Chen, T.; et al. Indole-3-carboxylic acid from the endophytic fungus Lasiodiplodia pseudotheobromae LPS-1 as a synergist enhancing the antagonism of jasmonic acid against Blumeria graminis on wheat. Front. Cell. Infect. Microbiol. 2022, 12, 898500. [Google Scholar] [CrossRef] [PubMed]
- Růžička, K.; Strader, L.C.; Bailly, A.; Yang, H.; Blakeslee, J.; Langowski, L.; Nejedla, E.; Fujita, H.; Itoh, H.; Syono, K.; et al. Arabidopsis PIS1 encodes the ABCG37 transporter of auxinic compounds including the auxin precursor indole-3-butyric acid. Proc. Natl. Acad. Sci. USA 2010, 107, 10749–10753. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Plant Height (cm) | Panicles/Plant (Numbers) | Panicle Weight (g) | Panicle Length (cm) | Seed/Panicle (Numbers) |
|---|---|---|---|---|---|
| Group-C | 84.6 ± 7.8 a | 7.1 ± 2.4 b | 9.11 ± 3.7 | 15. 2± 3.1 a | 14.6 ± 4.1 b |
| Group-D | 80.1 ± 10.4 a | 9.2 ± 3.3 ab | 13.11 ± 2.2 | 17.5 ± 4.2 a | 23.3 ± 5.7 ab |
| Group-E | 82.4 ± 8.2 a | 10.3 ± 1.6 a | 12.76 ± 2.6 | 16.3 ± 3.5 a | 29.5 ± 3.9 a |
| Metabolites * | Group C | Group D | Group E |
|---|---|---|---|
| Concentration (nM) | |||
| 1-Aminocyclopropanecarboxylic acid | 31.7 ± 6.3 b | 43.7 ± 8.2 ab | 53.1 ± 13 a |
| Indole-3-carboxaldehyde | 14.5 ± 3.2 b | 28.2 ± 14.7 a | 31.4 ± 4.3 a |
| Indole-3-acetic acid | 2.1 ± 0.2 b | 16.3 ± 3.5 a | 21.9 ± 7.1 a |
| 3-Indolebutyric acid | 0.17 ± 0.06 b | 0.55 ± 0.28 a | 0.44 ± 0.03 a |
| N6-(delta 2-Isopentenyl)-adenine | 0.04 ± 0.00 | 0.06 ± 0.00 | 0.06 ± 0.03 |
| Kinetin | 0.17 ± 0.03 | 0.16 ± 0.01 | 0.18 ± 0.01 |
| trans-Zeatin | 0.02 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 |
| cis-Zeatin | 0.07 ± 0.01 | 0.10 ± 0.09 | 0.11 ± 0.05 |
| N6-isopentenyladenosine | 0.03 ± 0.01 b | 0.14 ± 0.09 a | 0.14 ± 0.00 a |
| Salicylic acid | 30.5 ± 5.8 c | 179.8 ± 26.8 b | 290.3 ± 92.8 a |
| (±)-Jasmonic acid | 2.5 ± 0.8 | 2.2 ± 0.3 | 3.3 ± 0.8 |
| Dihydrojasmonic Acid | 0.59 ± 0.14 | 0.78 ± 0.41 | 1.34 ± 0.25 |
| N-((-)-jasmonoyl)-S-isoleucine | 0.13 ± 0.04 | 0.21 ± 0.01 | 0.23 ± 0.01 |
| Methyl 3-indolylacetate | N/D | N/D | N/D |
| Methyl salicylate | N/D | N/D | N/D |
| DL-Dihydrozeatin | N/D | N/D | N/D |
| Methyl jasmonate | N/D | N/D | N/D |
| trans-Zeatin-riboside | N/D | N/D | N/D |
| (+)-Abscisic acid | N/D | N/D | N/D |
| Gibberellin A7 | N/D | N/D | N/D |
| Gibberellin A4 | N/D | N/D | N/D |
| Gibberellin A3 | N/D | N/D | N/D |
| Gibberellin A1 | N/D | N/D | N/D |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.-C.; Zheng, Y.-J.; Lin, Y.-C.; Huang, C.-H.; Shen, T.-L.; Hsu, Y.-C.; Lee, B.-H. Investigation of the Microbial Diversity in the Oryza sativa Cultivation Environment and Artificial Transplantation of Microorganisms to Improve Sustainable Mycobiota. J. Fungi 2024, 10, 412. https://doi.org/10.3390/jof10060412
Shi Y-C, Zheng Y-J, Lin Y-C, Huang C-H, Shen T-L, Hsu Y-C, Lee B-H. Investigation of the Microbial Diversity in the Oryza sativa Cultivation Environment and Artificial Transplantation of Microorganisms to Improve Sustainable Mycobiota. Journal of Fungi. 2024; 10(6):412. https://doi.org/10.3390/jof10060412
Chicago/Turabian StyleShi, Yeu-Ching, Yu-Juan Zheng, Yi-Ching Lin, Cheng-Hao Huang, Tang-Long Shen, Yu-Chia Hsu, and Bao-Hong Lee. 2024. "Investigation of the Microbial Diversity in the Oryza sativa Cultivation Environment and Artificial Transplantation of Microorganisms to Improve Sustainable Mycobiota" Journal of Fungi 10, no. 6: 412. https://doi.org/10.3390/jof10060412
APA StyleShi, Y.-C., Zheng, Y.-J., Lin, Y.-C., Huang, C.-H., Shen, T.-L., Hsu, Y.-C., & Lee, B.-H. (2024). Investigation of the Microbial Diversity in the Oryza sativa Cultivation Environment and Artificial Transplantation of Microorganisms to Improve Sustainable Mycobiota. Journal of Fungi, 10(6), 412. https://doi.org/10.3390/jof10060412