Abstract
The evolution of azole resistance in fungal pathogens presents a major challenge in both crop production and human health. Apple orchards across the world are faced with the emergence of azole fungicide resistance in the apple scab pathogen Venturia inaequalis. Target site point mutations observed in this fungus to date cannot fully explain the reduction in sensitivity to azole fungicides. Here, polygenic resistance to tebuconazole was studied across a population of V. inaequalis. Genotyping by sequencing allowed Quantitative Trait Loci (QTLs) mapping to identify the genetic components controlling this fungicide resistance. Dose-dependent genetic resistance was identified, with distinct genetic components contributing to fungicide resistance at different exposure levels. A QTL within linkage group seven explained 65% of the variation in the effective dose required to reduce growth by 50% (ED50). This locus was also involved in resistance at lower fungicide doses (ED10). A second QTL in linkage group one was associated with dose-dependent resistance, explaining 34% of variation at low fungicide doses (ED10), but did not contribute to resistance at higher doses (ED50 and ED90). Within QTL regions, non-synonymous mutations were observed in several ATP-Binding Cassette and Major Facilitator SuperFamily transporter genes. These findings provide insight into the mechanisms of fungicide resistance that have evolved in horticultural pathogens. Identification of resistance gene candidates supports the development of molecular diagnostics to inform management practices.
1. Introduction
Apple scab, caused by the fungal plant pathogen Venturia inaequalis, is one of the most damaging diseases impacting apple production [1,2,3]. Infections via airborne spores result in characteristic “scab” like lesions on both leaves and fruit; these lesions render fruits unacceptable for direct-to-consumer retail. Extensive control measures are, therefore, required to prevent scab outbreaks and mitigate the severe economic impact of the disease. It is estimated that apple producers in the eastern United States alone spend in excess of USD 18.6 million per year on scab control [4].
Many fungicides are registered for the control of apple scab, and growers are advised to utilise products with differing modes of action to prevent the development of fungicide resistance [5]. Despite this, V. inaequalis has evolved resistance to successive classes of fungicides going back to the 1950s [6]. This resistance has often developed quickly after the introduction of a new active and been maintained in the pathogen population thereafter [6]. Combined with tightening regulations on pesticide use, fungicide resistance has left growers reliant on a small number of active ingredients.
Currently, demethylase inhibitors are considered the most effective class of fungicide available for scab control [7]. Sterol DeMethylase Inhibitors (DMIs/azoles) target an essential precursor for ergosterol biosynthesis in fungi (which does not occur in animals): the enzyme 14 α-demethylase, which is encoded by the CYtochrome P450 monooxygenase (CYP) superfamily gene CYP51 (ERG11) [8]. Non-competitive direct binding of DMIs to 14 α-demethylase leads to the accumulation of 14-methylated sterols, which in turn disrupt the cell membrane, slowing fungal growth [9]. DMIs are known for their strong post-infection activity, resulting in a reduction in spore production, and can also prevent scab lesions from developing if applied within 48–72 h of infection [7,10,11,12,13]. Given the limited number of fungicides that are still both available and effective against apple scab, it is necessary to monitor for resistance. If V. inaequalis evolves widespread resistance to DMIs before effective substitutes become available, growers stand to suffer substantial economic losses. An understanding of the genetics behind fungicide resistance is an essential prerequisite in order to design effective anti-resistance strategies and ensure the sustainable use of fungicides.
Reducing V. inaequalis sensitivity to DMIs has been a concern for apple growers since the mid-1980s, and reports from around the globe indicate that DMI resistance is becoming more prevalent [14,15,16,17,18,19,20,21,22,23,24,25,26,27]. V. inaequalis isolates with reduced sensitivity to DMIs were first identified within six years of DMI fungicide deployment in Canada [28]. A build-up of resistance was observed, progressing in a step-like manner and eventually resulting in a complete breakdown of scab control in the tenth year [28]. By contrast, in the eastern United States, it took more than 25 years for DMI-resistant strains of V. inaequalis to become established; Cox has subsequently reported that 73 of 93 surveyed orchards in the region could now be considered to have ‘practical resistance’ to the DMI ‘myclobutanil’ [4,29,30]. However, V. inaequalis populations considered to be practically resistant to myclobutanil were found to be manageable using other DMIs such as difenoconazole. Resistance to DMI fungicides in scab is widely considered to be incremental, quantitative, and/or dose-dependent [4].
Target-site point mutations of the CYP51 gene can explain resistance to DMIs in many fungal species, e.g., Candida albicans [8,31,32,33,34,35,36,37]. However, CYP51 point mutations that confer DMI resistance are generally not conserved amongst phytopathogenic fungi [32,38]. Yaegashi et al. were the first to report a point mutation of CYP51 associated with reduced DMI susceptibility in V. inaequalis (Y133F) [39]. However, this mutation could not fully explain variation in DMI sensitivity. Similar observations were made by Hoffmeister et al., who were able to associate several CYP51 mutations with higher EC50 values (half maximal effective concentration) but also found examples of wild-type CYP51 isolates with increased EC50 for DMI fungicides [40]. Thus, it is clear that CYP51 mutations contribute to DMI resistance in apple scab; however, they cannot fully explain all the resistance observed [34,39,40,41]. Indeed, previous studies have found that, in some orchards, none of the DMI-resistant V. inaequalis isolates sampled contained a CYP51 mutation [41,42]. This suggests that multiple mechanisms of DMI resistance have evolved in V. inaequalis.
Cordero-Limon et al. found that the progeny of a cross between fungicide-sensitive and -resistant V. inaqualis strains displayed quantitative variation in DMI sensitivity [43]. They concluded that a minimum of two genetic loci were involved in DMI resistance in V. inaequalis; however, the basis of this resistance remains uncharacterised. If DMI resistance in V. inaequalis is polygenic in nature, this could explain why field isolates exhibit a spectrum of sensitivity levels rather than a binary sensitive and insensitive phenotype [17,41,44]. However, DMI resistance QTLs, outside of the CYP51 gene or immediate upstream region, have not previously been reported in apple scab [39,40,41,42]. Here, we investigate the hypothesis that V. inaequalis resistance to the demethylase inhibitor fungicide ‘tebuconazole’ is a polygenic trait, controlled by genomic regions distinct to those involved in established CYP51 resistance. To test this, we sequenced the progeny of a cross between resistant and sensitive isolates and mapped genetic loci associated with fungicide resistance.
2. Materials and Methods
2.1. Generation of a Mapping Population and Measurement of Sensitivity
A V. inaequalis mapping population was generated and assayed for fungicide susceptibility by Cordero-Limon et al. [43]. Briefly, a susceptible isolate ‘AF28’ (ED50 = 0.12 mg L−1 (ED50 = the effective dose required to cause a 50% reduction in mycelial growth)), derived from Ash Farm, Worcestershire, UK, and never treated with fungicides, was crossed with a resistant isolate ‘Spartan 1’ (ED50 = 3.36 mg L−1) derived from a Kent orchard where fungicide resistance had been reported. Crossing of the two isolates was performed by sub-culturing mycelial plugs of each parent onto the same leaf decoction plate, chilled to 4 °C to facilitate sexual reproduction.
A total of 81 progeny strains were assessed for their level of susceptibility to tebuconazole. The commercial fungicide Folicur 25 EW (250 g L−1 tebuconazole) from Bayer Crop Science UK, Cambridge, UK, was used to amend PDA media (39 g L−1) (Oxoid, Basingstoke, UK), onto which 4 mm diameter circular plugs of each isolate were sub-cultured. Fungal growth was recorded at four fungicide concentrations (4 mg L−1, 1 mg L−1, 0.1 mg L−1, and 0.01 mg L−1) as well as on control plates containing no fungicide. These doses captured the declining phase of dose–response curves for both resistant ‘Spartan 1’ and susceptible ‘AF28’ parent isolates and were selected based on discriminatory doses used in previous studies of DMI resistance in V. inaequalis [45,46,47,48]. Two perpendicular measurements of colony diameter were taken for each colony three weeks after subculturing. The entire experiment was repeated once; within each replicate, each combination of isolate and concentration was represented by one plate containing three to four mycelial plugs, which were considered pseudoreplicates.
2.2. Calculation of Effective Dosages
Effective dose of fungicide required to reduce colony growth by 10, 50, and 90% were estimated for each isolate. The growth area of each colony was estimated, assuming an oval shape and subtracting the initial plug area. From this, mean colony growth measurements were calculated for each treatment within each repeat of the total assessment. These data were transformed into a percentage of maximum growth for each treatment (mean growth on fungicide-free control plates). The upper asymptote of each response curve could then be fixed at 100 (growth unimpeded by fungicide) and lower asymptotes fixed at zero (a lethally high dose). The R package ‘drc’ was used to model the dose–response curve for each isolate [49]. A two-parameter log-logistic model was determined to best-fit the data through assessment of AIC values generated from exploratory fitting of different dose–response models. The ‘ED’ function within the ‘drc’ package was used to estimate absolute ED10, ED50, and ED90 values for each strain (effective dose required for a 10, 50, and 90% reduction in growth, respectively) along with corresponding standard errors and upper and lower limits (Table S1).
Isolates with fewer than two doses in the declining phase of the response curve were discarded, as it was not possible to model the shape of the curve in this instance. Additionally, one datapoint, for isolate ‘RS11’ at zero fungicide concentration, was discarded as anomalous; this measurement for RS11 was far below the other replicates at zero fungicide concentration and values at higher fungicide doses.
2.3. DNA Extraction and Sequencing
DNA extractions were performed via a cetyltrimethylammonium bromide (CTAB)/chloroform–isoamyl alcohol extraction method. Frozen haploid mycelia were first homogenized via grinding in a geno/grinder® (Cole-Parmer, St. Neots, UK) with 2 ball bearings in 2 mL Eppendorf tubes at 1500 rpm in 20 s bursts for 2 min chilling with liquid nitrogen between bursts. Following this, samples were gently mixed at Room Temperature (RT) in 1.5 mL lysis buffer (Table S2), alone for 30 min, followed by an additional 30 min of incubation after the addition of 20 µL of proteinase K (Qiagen, Venlo, The Netherlands). Samples were then cooled on ice for 5 min, before 250 µL of 5 M potassium acetate was added, cooling then continued for an additional 5 min. Following this, samples were centrifuged for 12 min at 5000× g, and the supernatants were transferred to safe lock tubes.
To wash samples, a 1:1 volume of Phenol/Chloroform/Isoamyalcohol 100 mM Tris-EDTA pH 8 (P:C:I) (Thermo Fisher Scientific, Waltham, MA, USA) was added; samples were then inverted for 2 min and centrifuged for a further 10 min at 4000× g before transfer of supernatants to fresh tubes. This wash step was repeated three times per sample, followed by a fourth wash using Chloroform/Isoamyl alcohol (C:I) (Sigma-Aldrich, Saint Louis, MO, USA) in place of P:C:I.
To precipitate samples, 200 µL of sodium acetate (3 M, pH 5.2) and 800 µL of isopropanol were added per ~1 mL of supernatant, and samples were inverted at RT for 10 min. Samples were then centrifuged at 8000× g for 30 min before supernatants were removed, and the resulting pellets were washed three times in 1.5 mL of 70% ethanol through resuspension, centrifugation at 13,000× g for 5 min, and disposal of supernatants. Samples were then left to dry at RT for 30 min before addition of 100 µL of 10 mM Tris pH 8.5 and incubation at RT for 30 min to dissolve the extracted DNA.
Final DNA quality was assessed using a Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Purification, fragmentation, DNA library construction, and sequencing on an Illumina NovaSeq platform using paired-end chemistry (PE150) were performed by the commercial sequencing company Novogene.
2.4. Calling Single Nucleotide Polymorphism Markers
In order to genotype isolates, SNPs were identified using the GATK4 variant calling pipeline [50]. Raw sequencing reads were first trimmed and adapters removed using Trimmomatic v0.39 [51]. Additionally, sequencing depth for each isolate was assessed using the K-mer Analysis Toolkit v2.4.2 function ‘kat plot spectra-cn’ [52]. Trimmed reads were then aligned to the V. inaequalis genome generated by Passey et al. (Genbank accession: GCA_003351075.1) using Bowtie 2 v2.4.1 [53,54]. Multimapping reads, discordant reads, and duplicates were removed, and read group and sample names were added to each mapped read using SAMtools v1.1 [53,55]. Following this, Picard tools v2.26.11 was used to create a sequence dictionary for the reference scab sequence, and SNPs were called by GATK ‘HaplotypeCaller’. GATK ‘combineGVCFs’ was used to combine samples and allow joint genotyping via GATK ‘GenotypeGVCFs’.
Vcflib toolkit v0.1.16 and GATK4 were then used to perform filtering and removal of erroneous or low-quality SNPs (successful genotyped in <95% of individuals, quality score of <30 (base call accuracy of 99.9%), read depth of <5, maximum allele frequency of >60%, minimum allele frequency of <40%, or a minor allele count of <3). Following this, call data were filtered using GATK ‘VariantFiltration‘, retaining: QUAL < 30.0; QD < 2.0; SOR > 3.0; FS > 60.0; MQ < 40.0; MQRankSum < −12.5; ReadPosRankSum < −8.0. The resulting SNPs were plotted using R v4.3.0 [56].
2.5. Linkage Map Generation
Joinmap 5 was used to calculate a genetic linkage map for V. inaequalis [57]. Prior to mapping, SNPs with missing data, or those that were homozygous within the population, were excluded. Additionally, a principal component analysis was performed to identify any rogue genotypes across the isolates. Filtered SNPs were first converted from variant call to HapMap format using Tassel v5.2.81 and, subsequently, from HapMap to raw mapmaker format via a custom Python script (https://github.com/harrisonlab/nano_diagnositcs/blob/master/V_inaequalis/hapmap2mapmaker.py, accessed on 20 November 2023). Mapmaker files were then input to Joinmap 5 [57]. In order to reduce the computational resources required to calculate the linkage map, the ‘exclude similar loci’ function was used to exclude identical SNPs, resulting in a minimal set of 861 informative markers. Ten linkage groups were calculated at LOD = 3.4. The order of SNPs within these groups was then curated to ensure congruity with the known physical position of SNPs in reference genome contigs [54].
2.6. Mapping of Quantitative Trait Loci
Each of 861 informative SNPs was tested for association with differences in effective dose between genotypes. To facilitate this, effective dose scores (ED10, ED50, and ED90) were square root transformed to normalise the distribution of phenotypic data before one-way ANalysis Of VAriance (ANOVA) with Benjamini–Hochberg multiple test correction was performed to identify SNP markers significantly associated with effective dose (p ≤ 0.05). Significant SNPs were taken forward for QTL identification. QTL regions were defined surrounding a given QTL’s focal SNP (the SNP exhibiting the highest association with the fungicide resistance trait) and delimited by flanking SNPs on either side with comparatively lower associations.
Effect size and Proportion of Variance Explained (PVE) were calculated based upon the focal SNP within QTL regions. To estimate effect size, the difference in mean effective dose between isolates carrying the resistant parent genotype and those carrying the susceptible parent genotype was divided by the standard deviation for isolates carrying the susceptible genotype.
PVE was estimated by dividing the sum of squares (SSQ) for effective dose of isolates carrying a variant SNP marker by the sum of squares across all isolates.
A two-way ANOVA was subsequently performed for the principal markers 009.1_102387 and 005.1_648012 to test for the presence of statistical interactions between QTLs.
2.7. Candidate Gene Characterisation
Gene models and functional annotations associated with the reference V. inaequalis assembly were used to identify candidate resistance genes within QTL regions [54]. Putative proteins were investigated where genes fell within two QTL regions: firstly, between the SNP markers 009.1_97361 and 009.1_402743 (positions 97,361 and 402,742 of the contig Genbank accession: QFBF01000009.1); and secondly, between SNP markers 005.1_642152 and 005.1_899960 (positions 642,152 and 899,960 of the contig Genbank accession: QFBF01000005.1). Amino acid sequences were aligned to the non-redundant protein sequences (nr) database using the NCBI BLASTp web tool with default parameters to search for homologous proteins [58]. Additionally, protein families, domains, and functional sites were classified by amino acid searches using the European Bioinformatics Institute InterPro and PfamScan web tools with default parameters [59,60].
3. Results
3.1. Progeny Isolates Segregated by Tebuconazole Sensitivity
Sensitivity to tebuconazole within the crossing population ranged from 0.015 mg L−1 to 2.63 mg L−1 (measured as ED50). The distribution of sensitivity values was continuous across the population, and isolates could not be divided into discrete groups (Figure 1). The fungicide-sensitive parent (AF28) had an ED50 of 0.27 mg L−1 and ranked as the 24th least sensitive isolate. In contrast, the resistant parent (Spartan 1) had one of the lowest sensitivities within the population (ED50 = 1.84 mg L−1), second only to one progeny isolate. ‘Spartan 1’ was the least sensitive isolate as measured by ED90; however, there were 16 isolates with a higher ED10 value. ‘AF28’ had the 18th lowest ED10 value and 29th lowest ED90 value. ‘Spartan 1’ was consistently less susceptible than ‘AF28’.
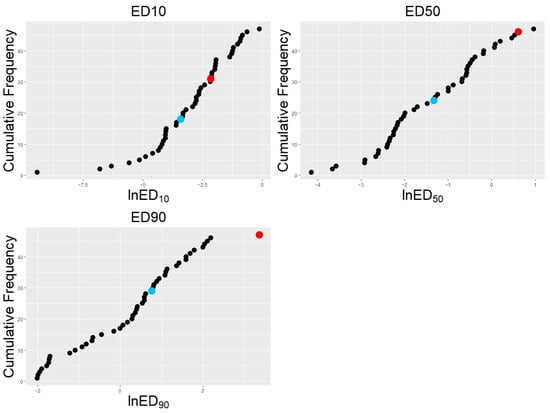
Figure 1.
Cumulative frequency of Venturia inaequalis tebuconazole effective doses: Sensitivity of 47 apple scab isolates plotted with reducing susceptibility to tebuconazole from left to right; susceptibility defined as the effective dose required to cause a 10, 50, or 90% reduction in mycelial growth: ED10, ED50, and ED90. Progeny isolates are shown in black; parent isolates are shown in blue (Sensitive parent ‘AF28’ with lower effective dosages) and red (resistant parent ‘Spartan 1’ with higher effective dosages). Raw data were generated as part of previous work by Cordero-Limon et al. [43].
3.2. Whole-Genome Resequencing Identified Single Nucleotide Polymorphisms across the Mapping Population
Whole-genome skim sequencing was performed for 52 apple scab progeny isolates and the two parents of the mapping population, ‘AF28’ and ‘Spartan 1’. An average of 9,115,913 raw reads were generated for each isolate, with a minimum coverage of ~15× achieved for the 54 isolates, including the two parents. An average of 89.9% of paired trimmed reads aligned to the reference assembly.
A total of 603,595 SNP sites were called across the ~72 Mb V. inaequalis reference genome, a SNP, on average, every 119 bp. Of these, a total of 46,727 were high-quality polymorphic SNPs, representing an average of one SNP per 1540 bp interval. These SNPs fell across 90 of the 238 contigs within the reference assembly, with these 90 contigs representing 89% of the total assembly (64,327,219 bp of 72,310,420 bp).
3.3. Single Nucleotide Polymorphisms Mapped to Nine Linkage Groups
A total of 861 informative SNP markers were used to generate a linkage map consisting of 10 groups for V. inaequalis, which has seven chromosomes. The size of linkage groups ranged from 17,896,836 bp for linkage group one, represented by 241 informative SNPs, to 612,523 bp for linkage group ten, represented by one informative SNP (Table 1).

Table 1.
Venturia inaequalis linkage groups: The sizes of 10 assembled linkage groups are given along with the number of Single Nucleotide Polymorphisms (SNPs) these contain and the number of informative SNPs representing segregation.
3.4. Two Dose-Dependent Quantitative Trait Loci for Tebuconazole Sensitivity Were Identified
Genomic regions associated with tebuconazole sensitivity were determined through QTL mapping of genotypic and phenotypic data. SNPs significantly associated with reduced tebuconazole sensitivity were identified and mapped to V. inaequalis linkage groups, with significance assessed at ED10, ED50, and ED90 levels. From this, two QTLs were identified at ED10, each comprising a central ‘focal SNP’ flanked by SNPs whose association diminishes as the genetic distance from the focal SNP increases (Figure 2). Identified focal SNPs fell within linkage group one (LG1) (p = 0.044) and linkage group seven (LG7) (p = 0.0007).
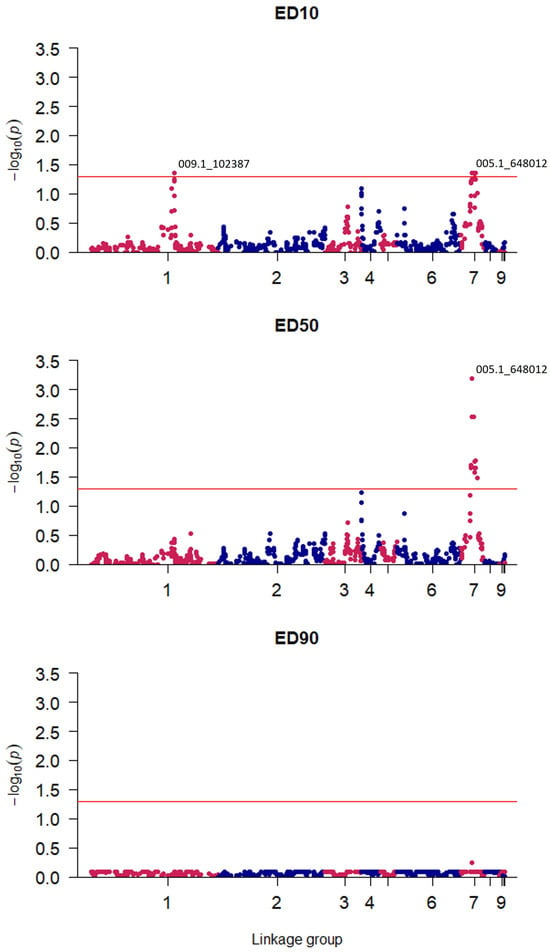
Figure 2.
Quantitative Trait Locus (QTL) mapping identifies a dose-dependent response to tebuconazole sensitivity over ED10, ED50, and ED90: 861 informative Single Nucleotide Polymorphism (SNP) markers are plotted by significance of their association at a given effective dose (ED10, ED50, or ED90) against their estimated position within the Venturia inaequalis genome in one of the ten linkage groups. Significance of p = 0.05 is indicated by a red horizontal line. The ID of the most significant SNP marker is indicated for each identified QTL.
A dose-dependent genetic response was observed; the QTL in LG7 was found to be significant at both ED10 and ED50, whilst the QTL in LG1 was only significant at ED10. The LG1 QTL had an effect size of −0.9 SD at ED10 and explained an estimated 34% of observed variance. The effect size of this LG1 QTL was negative as the resistant parent carried a susceptible variant marker at this QTL. Meanwhile, the LG7 QTL had an effect size of 1.36 SD and explained 36% of the variance in ED10; in contrast, 65% of variance in ED50 was explained by the LG7 QTL, which had an effect size of 2.18 SD. Given an overall ED50 range of 4.71 mg L−1, these QTLs were associated with major effect alleles. A third putative QTL in LG4 fell slightly below the threshold for significance (p = 0.059) at ED50, with an effect size of 0.64 SD, and explained 26% of the variance.
Interaction between QTLs was investigated. Isolates carrying resistance alleles at both QTLs were observed to have reduced susceptibility at ED10 and ED50 versus isolates carrying either an LG1 resistance allele or an LG7 resistance allele alone (Figure 3). A two-way ANOVA, considering the focal SNPs for each of the two QTLs, found no evidence of an interaction effect associated with tebuconazole sensitivity (Table 2); this indicates that the two QTLs have an additive effect for resistance at these doses.
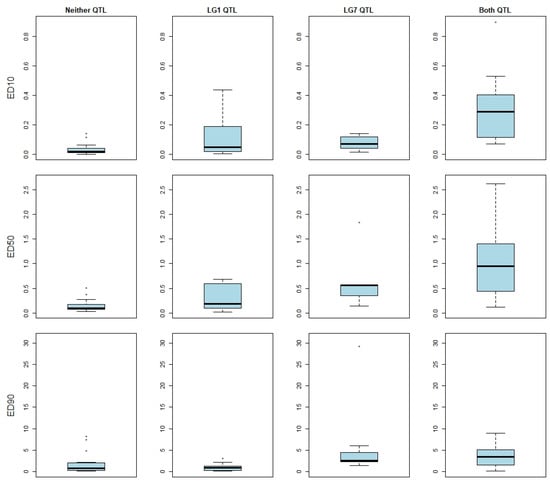
Figure 3.
Resistance Quantitative Trait Loci (QTLs) contribute additively to tebuconazole susceptibility at low doses: Contribution of combinations of QTLs to tebuconazole susceptibility, defined as the effective dose required to cause a 10, 50, or 90% reduction in mycelial growth (ED10, ED50, and ED90). Sensitivities for apple scab isolates carrying neither resistant allele (n = 7), carrying only a linkage group one resistance allele (n = 12), carrying only a linkage group seven allele (n = 16), and carrying both resistant alleles (n = 12) are plotted left to right, respectively.
Table 2.
Assessment of non-additive interactions between Quantitative Trait Loci (QTLs): Two-way ANalysis Of VAriance (ANOVA) for linkage group one (LG1) and linkage group seven (LG7) QTLs. p-values for LG1 QTL, LG7 QTL, and interaction (LG1 QTL: LG7 QTL) are given for effective dose required to cause a 10, 50, or 90% reduction in mycelial growth (ED10, ED50, and ED90), as well as the percentage of variability explained by these two QTLs, determined using the R-squared value.
3.5. Quantitative Trait Loci Location and Gene Content
Genes models were investigated within the two QTL regions predicted. The QTL in LG7 spanned 257 kb of the V. inaequalis genome and was associated with a greater difference in fungicide sensitivity than any other locus identified. A total of 70 genes were predicted within this region (Table S3), including six with InterPro Gene Ontology (GO) term annotations for transmembrane transport. Additionally, three putative proteins returned annotations/sequence similarity matches related to the regulation of transcription. These annotations were investigated in detail as improved efflux of toxins and altered expression of CYP51 are two possible fungicide resistance mechanisms. Twenty gene models in this QTL region contained non-synonymous SNPs, resulting in amino acid changes in corresponding putative proteins. These included three transmembrane transport genes with the following non-synonymous SNPs: A610T and L615F substitutions in a putative voltage-gated chloride channel protein (Accession: RDI87354.1); a Q1154E substitution in a putative ATP-Binding Cassette (ABC) transporter protein (Accession: RDI87357.1); as well as T489A, H501R and A774E substitutions in a putative active sulfate transmembrane transporter (Accession: RDI87081.1). Additionally, all three transcription-associated genes carried non-synonymous SNPs: P174T and A458T substitutions in a putative zinc finger protein (Accession: RDI87174.1); a H47R substitution in a putative zinc finger protein (Accession: RDI87163.1); and a T403A substitution in a putative helicase (Accession: RDI87242.1).
The LG7 QTL is located 435 kb from the V. inaequalis CYP51 gene, which encodes 14-α sterol demethylase; the enzyme responsible for catalysing a key demethylation step in the synthesis of ergosterol and the target of demethylase inhibitor fungicides. Cordero-Limon et al. previously sequenced the CYP51 genes of ‘AF28’ and ‘Spartan 1’, confirming that there were no point mutations within the gene sequences of the parent isolates [13]. The most significant marker in the LG7 QTL, ‘005.1_648012’, is located a large distance (683 kb) upstream of the CYP51 gene location. Although SNPs between marker ‘005.1_648012’ and CYP51 were significantly associated with tebuconazole resistance, this was to a lesser extent than the focal SNP and in a pattern consistent with linkage to a neighbouring allele, indicating the LG7 QTL represents a novel resistance locus.
The second resistance QTL was identified in LG1 and spanned 305 kb; 39 gene models were predicted within this QTL region, 19 of which carried non-synonymous SNPs (Table S4). Three of these genes encoded putative Major Facilitator Superfamily (MFS) transporter proteins, two of which carried SNPs resulting in amino acid changes: an I796M substitution in the protein Vi05172_g3976 (Accession: RDI85792.1) as well as V223G and G326D substitutions in Vi05172_g3984 (Accession: RDI85945.1). No transcription-related annotations were identified within the LG1 QTL.
Taken together, we identify 39 genes that demonstrate non-synonymous mutations within QTL regions. Through the identification of putative transmembrane transport proteins and transcription factors, we prioritise a subset of these as candidates that may contribute to resistance for future study. These include six gene candidates for LG7 QTL resistance and two gene candidates for LG1 QTL resistance.
4. Discussion
Here, we have performed the first QTL mapping study of fungicide resistance in V. inaequalis and demonstrated that multiple loci are associated with resistance to the DMI tebuconazole. These results support our original hypothesis that DMI resistance in the apple scab pathogen population is a polygenic trait [43]. Moreover, both genetic loci identified were observed to meditate dose-dependent resistance. Finally, we identify putative transporter proteins in both QTL regions, which may underly this novel dose-dependent resistance mechanism in V. inaequalis.
4.1. Multiple Loci Contribute to the Level of Tebuconazole Sensitivity in the Apple Scab Pathogen
A progressive increase in resistance to DMI fungicides has been reported in apple scab pathogen populations since the mid-1980s [14,15,17,61]. The QTL mapping results presented here support the hypothesis that, at lower dosages, DMI resistance in V. inaequalis is due to the additive effects of multiple loci (Figure 3, Table 2). This may explain the relatively delayed development of DMI resistance in V. inaequalis compared to resistance against other chemical families [6]. The evolution of polygenic resistance typically takes longer to arise than monogenic resistance, as at each step, any fitness disadvantage conferred by a resistance mutation must be compensated for, and each iterative step must be advantageous for the mutation to persist [62].
Those QTLs identified here in V. inaequalis are dose-dependent; a resistance QTL was identified in linkage group one, which was important only at lower fungicide concentrations (ED10), whilst a QTL within linkage group seven was associated with resistance at both ED10 and ED50. Whereas there were 23 progeny (42.6%) with greater susceptibility than the susceptible parent ‘AF28’ (ED50), only one isolate was more resistant than the resistant parent ‘Spartan 1’ (ED50). This asymmetric distribution is inconsistent with purely additive QTL effects [43]. Additionally, dominance effects between alleles are unlikely, given that V. inaequalis is haploid outside of a very brief diploid stage during sexual reproduction. Epistatic effects are, therefore, likely to underpin the distribution of susceptibilities at higher fungicide concentrations. This asymmetric distribution was present only in ED50 and ED90 estimates and was not observed for ED10 (Figure 1). Therefore, the data and analyses show that resistances associated with the LG1 and LG7 QTLs behave in an additive fashion at low fungicide concentrations, whereas at high fungicide concentrations, only the LG7 QTL, carried by ‘Spartan 1’, has an impact. In a single-gene scenario, no progeny isolates are expected to significantly exceed the parental ED50.
The resistant parent ‘Spartan 1’ carries a susceptible variant genotype for the LG1 QTL, whilst the susceptible parent ‘AF28’ has a resistant genotype at this locus. This explains why, for ED10, certain progeny isolates were observed to exhibit higher levels of resistance than the resistant parent, whilst others displayed greater susceptibility than the susceptible parent. This result underscores the complexity of DMI fungicide resistance in the apple scab pathogen. Additionally, several loci were identified that appeared to be associated with resistance but fell short of statistical significance in this study, for example, within linkage group four at ED50 and linkage group three at ED10 (Figure 2). Clearly, there is scope for evolution of the pathogen to further increase the resistance levels of even the most tolerant isolates. This study demonstrates polygenic resistance in a mapping population generated from the isolate ‘Spartan 1’, which was sampled from a single UK orchard suffering from fungicide-resistant scab outbreaks. As such, further research is required to assess the prevalence of the observed resistance across field populations in the UK and globally. Furthermore, it will be important to determine whether the resistance and SNPs described here co-occur with SNPs identified in previous studies (e.g., CYP51 Y133F) [39,40]. Such monitoring and appropriate mitigation measures are particularly important for sexual species, such as V. inaequalis, to prevent the stacking of resistance mutations in pathogen populations.
4.2. Quantitative Trait Loci Indicate a Novel Mechanism of Resistance
Azole resistance in fungal pathogens has been attributed to several mechanisms, including point mutations in CYP51, duplication of CYP51, overexpression of CYP51, modified efflux through ABC transporters, alternative modes of sterol biosynthesis, and changes in membrane composition [8,31,32,33,34,35,36,37,38,39,40,41,63,64,65,66,67,68,69,70]. Of these, point mutations and altered expression of the CYP51 gene and upstream region have previously been investigated in V. inaequalis [39,40,41,42]. Mutations in CYP51 can explain DMI resistance in many fungal species, including plant and human pathogens such as Aspergillus fumigatus, Blumeria graminis, Candida albicans, Cercospora beticola, and Erysiphe necator [32,34,39,40,41]. However, none of the common CYP51 point mutations associated with DMI resistance were identified in the resistant parent ‘Spartan 1’ (e.g., Y133F). Of the two resistance QTLs identified in this study, one is in a separate linkage group to CYP51, whilst another, strongly associated with resistance at both ED10 and ED50, is located 435 kb from the CYP51 gene. This lack of proximity to the CYP51 gene indicates that, in these QTLs, we have identified discrete alleles associated with resistance.
Insertions < 1000 bp upstream of CYP51 have previously been associated with resistance to the DMI difenoconazole in V. inaequalis [41,42]. However, regulatory elements were not anticipated at a distance of 435 kb or more from the CYP51 gene, as all DMI resistance mutations reported in V. inaequalis to date have been within the gene coding region itself or immediately upstream (<2 kb). Nonetheless, we cannot rule out the possibility that the QTLs identified here represent distal regulatory elements of CYP51. Indeed, putative proteins encoded in one QTL returned GO terms related to DNA-templated transcription and had sequence similarity to transcription factor Zn [43]. Additionally, many genes in the QTL regions are of unknown function, and several lacked homologs outside of V. inaequalis. These genes may encode a trans-regulatory transcription factor that affects CYP51, or they play a role in regulating other genes, leading to resistance.
Overexpression of CYP51 has been implicated in DMI fungicide resistance in several phytopathogenic fungi [31,71,72,73,74,75,76]. In Erysiphe necator, increased CYP51 copy number enhances fungicide resistance, whilst in Aspergillus fumigatus, a second gene encoding the CYP51 protein has been discovered [69,70]. Increased expression of the CYP51 gene has also been investigated in V. inaequalis; Villani et al. (2016) identified a 169 bp repeatable element carrying transcription factor binding sites upstream of CYP51, which was associated with a 9–13-fold increase in gene expression and resistance to the DMI difenoconazole [42]. However, neither the presence of these insertions nor CYP51 overexpression could be associated with myclobutanil resistance, suggesting that different mechanisms may influence resistance to different DMI-class fungicides [42].
4.3. Dose-Dependent Resistance May Result from Variant Efflux Transporter Genes
A resistance QTL was identified in LG1, which was important only at lower fungicide concentrations (ED10). Within this QTL, two genes encoding putative MFS transporter proteins contained non-synonymous SNPs, resulting in changes to the translated protein sequence and potentially protein function. Upregulation, altered specificity, or altered efficiency of efflux proteins, that detoxify the fungus at lower quantities of fungicide but are overwhelmed at higher concentrations, could explain the dose-dependent nature of resistance. Alterations to transport efflux could also explain the cross-resistance between different DMI fungicides, as previously observed between myclobutanil and tebuconazole in the ‘AF28’ × ‘Spartan 1’ population by Cordero-Limon et al., enabling detoxification of several different fungicides [44,72]. Active efflux transporters, such as ABC and MFS proteins, have remarkably broad substrate specificity, and upregulation of ABC and MFS transporters is known to be involved in the development of resistance to DMIs in other fungi [63,64,77,78,79,80]. For example, enhanced efflux adaptations have been observed in fungicide-resistant strains of Candida spp., Botrytis cinerea, and Z. tritici [81,82,83,84,85,86]. Additionally, laboratory mutants of V. inaequalis have been generated which exhibit increased efflux efficiency associated with DMI resistance [87]. Vijaya Palani and Lalithakumari were able to generate DMI-resistant V. inaequalis isolates through chemical mutagenesis, resulting in an increased energy-dependent efflux rate [87]. However, changes in chemical transport have not yet been associated with V. inaequalis fungicide resistance in the field. As such, transporter genes carrying non-synonymous SNPs in this study represent good resistance gene candidates for future investigation. We cannot rule out the contribution of the other genes possessing non-synonymous SNPs within this region. However, we suggest prioritisation of the identified putative transporter genes for functional validation to assess their role in resistance.
The QTL within LG7, which is associated with resistance at both ED10 and ED50, also contained genes encoding putative transporters. However, in this QTL, only one putative transporter protein had broad specificity: an ABC transporter that carries amino acid substitutions in the resistant parent ‘Spartan 1’. This protein, Vi05172_g2818, has sequence similarity to ABC multidrug transporter-like proteins present in many other species and may underly the resistance associated with the LG7 QTL. Whist there is little literature concerning the function of ABC transporters in V. inaequalis, ABC transporters have been demonstrated to be involved in resistance to azoles in other species, such as Penicillium digitatum and Botrytis cinerea [64,78,88].
Unfortunately, if resistance indeed derives from improved efflux transporter activity, this may reduce the effectiveness of mixed fungicide applications combining different modes of action. Cross-resistance is most common between chemicals with the same target site (i.e., Other DMIs); however, enhanced efflux can also cause cross-resistance between different fungicide classes [43,89,90].
4.4. Alternative Resistance Mechanisms
In addition to transporter proteins and transcription factors, a number of additional protein families may play a role in DMI resistance. It has been proposed that in addition to CYP51, other proteins in the ergosterol biosynthesis pathway could be involved in resistance to demethylase inhibitors. A reduction in the total rates of lipid and ergosterol synthesis may allow efflux at normal rates to prevent the build-up of damaging levels of methylated ergosterol. In Ustilago maydis, a defective desaturase protein in the sterol pathway has been associated with resistance to demethylase inhibitors [66]. In Z. tritici, another methyltransferase in the ergosterol biosynthesis pathway (ERG6), a phospholipid biosynthesis protein, and a polyketide synthase involved in melanisation (PSK1) are proposed to be involved in resistance; it is believed that melanin may be able to adhere to fungicidal chemicals, preventing them from reaching their target [63]. In C. beticola, a group of polyketide synthase genes have been associated with resistance to the DMI tetraconazole and are believed to cause resistance by fortifying cell membranes [67]. As such, similar mechanisms should also be considered for genes in the vicinity of the LG1 and LG7 QTLs. Such annotations were not observed; however, given that many genes lack any functional annotations (Tables S3 and S4, 14 of 39 genes containing non-synonymous SNPs), as gene annotation pipelines improve, we may prioritise additional gene candidates for functional assessment of contribution to DMI resistance.
4.5. Lab-Based vs. In-Field Assessments
In this work, fungicide susceptibility was assessed under lab conditions, care must therefore be taken when relating the observed resistance to the field. We note that the recommended tebuconazole dose for in-field apple scab control is 125 mg L−1, with the application of 1500 L ha−1. However, fungicide exposure under spray conditions differs from exposure under laboratory conditions in infused agar plates. We found that a maximum experimental dose of 4 mg L−1 was sufficient to reduce the growth of ‘Spartan 1’, which was sampled from a resistant field population, by ~60% in a laboratory setting. This level of fungicide exposure is in line with similar lab-based studies. For example, using a discriminatory tebuconazole dose of 3 mg L−1 in vitro, Keinath et al. were able to predict cross-resistance of Stagonosporopsis citrulli isolates to five DMI fungicides in planta, whilst Chen et al. used a discriminatory dose of 5 mg L−1 to investigate tebuconazole resistance in 1118 Fusarium graminearum strains [91,92].
4.6. Implications for Practical Management
The identification of resistance gene candidates underpins the development of molecular diagnostics to inform management practices. Ideally, steps should be taken to tackle the emergence of fungicide resistance whilst the frequency of resistance genes islow, as it may be possible to prevent the fixation of resistance alleles in pathogen populations [89,93]. To achieve this, pre-emptive screening for the emergence of resistance alleles is required. Historically, identification of apple scab has primarily relied upon visual inspection of symptoms or spores; however, more recently, molecular techniques such as real-time PCR and loop-mediated isothermal amplification have been applied to V. inaequalis detection [94,95,96,97,98]. These methods could be adapted to screen for fungicide resistance markers and monitor the build-up of incremental gains in fungicide resistance within pathogen populations. Low-dose fungicide resistance (e.g., LG1 QTL) may be hard to track in the environment through phenotyping alone. Identifying genetic markers underpinning this resistance is, therefore, an important step towards effective resistance monitoring.
V. inaequalis’ ability to rapidly evolve and overcome host resistance genes and fungicides is aided by its two reproductive phases. Repeated cycles of asexual reproduction in each growing season generate an enormous number of progeny and facilitate the rapid spread of adaptive mutational events, whilst sexual reproduction over winter facilitates the combination of novel mutations and can accelerate evolution [99]. As growers seek to maximise the effective lifespan of available fungicides and forestall the evolution of resistance, a key principle is to ‘hit hard and hit early’ [100]. This approach minimizes population sizes, thereby limiting opportunities for adaptive mutation, and prevents a gradual buildup of resistance driven by the selective pressure of sublethal fungicide doses [89]. Past fungicide application strategies for the control of apple scab have come under scrutiny where regimes have not adhered to this approach [93]. Strategies that rely upon the strong post-infection activity of DMIs and seek to reduce the quantity of fungicide used, by spraying ‘curatively’ once the pathogen population is well established, on alternate rows of trees, and/or at reduced doses, are to be discouraged [7,10,11,12,13,93]. The identification here of dose-dependent resistance mechanisms in V. inaequalis underscores the importance of applying the full recommended dose [101]. In theory, the use of high doses should prevent selection for quantitative resistance by killing partially resistant genotypes along with sensitive isolates, although there is limited evidence of this in field settings [93,101,102].
4.7. Summary
We have demonstrated, for the first time, that DMI resistance in V. inaequalis is controlled by multiple loci. We also show that a dose-dependent genetic resistance response exists against azole fungicides, which provides insight into the evolution of fungicide resistance in V. inaequalis and other fungi that are subject to azole fungicides. These findings represent an important step towards understanding the complexity of resistance and improving the control of the apple scab pathogen [96,103]. The identification of novel resistance QTLs highlights putative fungicide resistance genes. Functionally validated resistance genes represent good diagnostic targets for monitoring and surveillance of fungicide resistance in pathogenic fungi, which aids in the early detection and containment of resistant strains [104,105]. Additionally, deciphering the mechanisms behind resistance can be used to model the rate and pattern of resistance development over time. By modulating dose, or choosing combinations of fungicides with different targets or modes of action, it may be possible to overcome or delay the establishment of resistance [102,106].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9121136/s1, Table S1. Estimated ED10, ED50, and ED90 values for each strain: Untransformed estimates of ED10, ED50, and ED90 (effective dose required for a 10, 50, and 90% reduction in growth, respectively) are recorded, along with corresponding standard errors and upper and lower limits; Table S2. Lysis buffer: Constituents of buffers used to prepare lysis buffer for DNA extrac-tion. Fresh lysis buffer was prepared as required prior to DNA extractions by combining stock preparations of BuffersA-E in the ratio 5:5:2:1:1 and adding 10 µL (10 kU) RNase A per 14 mL; Table S3. Interior annotations for Linkage group seven Quantitative Trait Loci (QTL): Gene ID, Genbank accession ID, and any Gene Ontology (GO) terms associated with the predicted protein (InterPro search, default settings) are given for each prediction, starred (*) gene IDs contain non-synonymous Single Nucleotide Polymorphisms; Table S4. Interior annotations for Linkage group one Quantitative Trait Loci (QTL): Gene ID, Genbank accession ID, and any Gene Ontology (GO) terms associated with the predicted protein (InterPro search, default settings) are given for each prediction, starred (*) gene IDs contain non-synonymous Single Nucleotide Polymorphisms.
Author Contributions
Conceptualization, X.X., H.M.C. and A.D.A.; methodology, H.M.C. and X.X.; software, A.D.A. and T.H.; formal analysis, T.H.; investigation, T.H.; writing—original draft preparation, T.H.; writing—review and editing, H.M.C., A.D.A., X.X., M.R.G. and T.H.; supervision, H.M.C., A.D.A., M.R.G. and X.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Biotechnology and Biosciences Research Council (BBSRC) through the Collaborative Training Partnership for Fruit Crop Research (CTP FCR), grant number BB/S507180/1. The APC was funded by RCUK open access block grant C-101746.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available in NCBI SRA archive, BioProject accession PRJNA817384 Venturia inaequalis Map.
Acknowledgments
The authors would like to acknowledge the phenotyping work conducted by Laura Cordero-Limon, Thomas Passey, and Joyce Robinson, which was published as part of a separate publication [43]. The authors would also like to thank Thomas Passey for laboratory support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carisse, O.; Bernier, J. Effect of Environmental Factors on Growth, Pycnidial Production and Spore Germination of Microsphaeropsis Isolates with Biocontrol Potential against Apple Scab. Mycol. Res. 2002, 106, 1455–1462. [Google Scholar] [CrossRef]
- Bowen, J.K.; Mesarich, C.H.; Bus, V.G.M.; Beresford, R.M.; Plummer, K.M.; Templeton, M.D. Venturia inaequalis: The Causal Agent of Apple Scab. Mol. Plant Pathol. 2011, 12, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Chatzidimopoulos, M.; Lioliopoulou, F.; Sotiropoulos, T.; Vellios, E. Efficient Control of Apple Scab with Targeted Spray Applications. Agronomy 2020, 10, 217. [Google Scholar] [CrossRef]
- Cox, K.D. Fungicide Resistance in Venturia inaequalis, the Causal Agent of Apple Scab, in the United States. In Fungicide Resistance in Plant Pathogens: Principles and a Guide to Practical Management; Ishii, H., Hollomon, D.W., Eds.; Springer: Tokyo, Japan, 2015; pp. 433–448. ISBN 978-4-431-55641-1. [Google Scholar]
- FRAG Fungicide Resistance Action Group. Fungicide Resistance Management in Apple and Pear Pathogens. Available online: https://media.ahdb.org.uk/media/Default/Imported%20Publication%20Docs/AHDB%20Cereals%20&%20Oilseeds/Disease/FRAG%20Fungicide%20resistance%20management%20in%20apple%20and%20pear%20pathogens%20(March%202015).pdf (accessed on 10 February 2023).
- MacHardy, W.E. Apple Scab: Biology, Epidemiology, and Management; APS Press: St Paul, MN, USA, 1996; ISBN 0-89054-206-6. [Google Scholar]
- AHDB Agriculture and Horticulture Development Board. Apple Best Practice Guide: Apple Scab—Additional Information. Available online: https://applegrowing.niab.com/apple-scab-additional-information/ (accessed on 2 February 2023).
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The Fungal CYP51s: Their Functions, Structures, Related Drug Resistance, and Inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.E.; Warrilow, A.G.S.; Price, C.L.; Mullins, J.G.L.; Kelly, D.E.; Kelly, S.L. Resistance to Antifungals That Target CYP51. J. Chem. Biol. 2014, 7, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, W.F.S.; Jones, A.L.; Jonker, J.P. Changes in the Susceptibility of Developing Apple Fruit to Venturia inaequalis. Phytopathology 1984, 74, 118–121. [Google Scholar] [CrossRef]
- O’Leary, A.L.; Sutton, T.B. The Influence of Temperature and Moisture on the Quantitative Production of Pseudothecia of Venturia inaequalis. Phytopathology 1986, 76, 199–204. [Google Scholar] [CrossRef]
- Poblete, J.A.; Latorre, B.A. Efecto preventivo y curativo de los fungicidas inhibidores de esteroles en el control de Venturia inaequalis del manzano. Cienc. E Investig. Agrar. 2001, 28, 145–150. [Google Scholar] [CrossRef]
- Cordero-Limon, L. Genetics behind the Variability in Sensitivity to the Demethylation Inhibitor (DMI) Fungicides Myclobutanil and Tebuconazole in Venturia inaequalis. PhD Thesis, University of Reading, Reading, UK, 2018. [Google Scholar]
- Chapman, K.S.; Sundin, G.W.; Beckerman, J.L. Identification of Resistance to Multiple Fungicides in Field Populations of Venturia inaequalis. Plant Dis. 2011, 95, 921–926. [Google Scholar] [CrossRef]
- Stanis, V.F.; Jones, A.L. Reduced Sensitivity to Sterol-Inhibiting Fungicides in Field Isolates of Venturia inaequalis. Phytopathology 1985, 75, 1098–1101. [Google Scholar] [CrossRef]
- Köller, W.; Smith, F.D.; Reynolds, K.L. Phenotypic Instability of Flusilazole Sensitivity in Venturia inaequalis. Plant Pathol. 1991, 40, 608–611. [Google Scholar] [CrossRef]
- Jobin, T.; Carisse, O. Incidence of Myclobutanil- and Kresoxim-Methyl-Insensitive Isolates of Venturia inaequalis in Quebec Orchards. Plant Dis. 2007, 91, 1351–1358. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gao, L.; Berrie, A.; Yang, J.; Xu, X. Within- and between-Orchard Variability in the Sensitivity of Venturia inaequalis to Myclobutanil, a DMI Fungicide, in the UK. Pest. Manag. Sci. 2009, 65, 1241–1249. [Google Scholar] [CrossRef]
- Sombardier, A.; Dufour, M.-C.; Blancard, D.; Corio-Costet, M.-F. Sensitivity of Podosphaera aphanis Isolates to DMI Fungicides: Distribution and Reduced Cross-Sensitivity: Sensitivity of P. Aphanis to DMI Fungicides. Pest. Manag. Sci. 2010, 66, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Stević, M.; Vukša, P.; Elezović, I. Resistance of Venturia inaequalis to Demethylation Inhibiting (DMI) Fungicides. Žemdir-bystė-Agric. 2010, 97, 65–72. [Google Scholar]
- Henríquez, S.J.L.; Sarmiento, V.O.; Alarcón, C.P. Sensitivity of Venturia inaequalis Chilean Isolates to Difenoconazole, Fenarimol, Mancozeb, and Pyrimethanil. Chil. J. Agric. Res. 2011, 71, 39–44. [Google Scholar] [CrossRef]
- Beresford, R.; Wright, P.; Wood, P.; Park, N. Sensitivity of Venturia inaequalis to Myclobutanil Penconazole and Dodine in Relation to Fungicide Use in Hawkes Bay Apple Orchards. N. Z. Plant Prot. 2012, 65, 106–113. [Google Scholar] [CrossRef]
- Mondino, P.; Casanova, L.; Celio, A.; Bentancur, O.; Leoni, C.; Alaniz, S. Sensitivity of Venturia inaequalis to Trifloxystrobin and Difenoconazole in Uruguay. J. Phytopathol. 2014, 163, 1–10. [Google Scholar] [CrossRef]
- Villani, S.M.; Biggs, A.R.; Cooley, D.R.; Raes, J.J.; Cox, K.D. Prevalence of Myclobutanil Resistance and Difenoconazole Insensitivity in Populations of Venturia inaequalis. Plant Dis. 2015, 99, 1526–1536. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Yu, Z.; Gao, L.; Yang, J. Investigating the Sensitivity of Venturia inaequalis Isolates to Difenoconazole and Pyraclostrobin in Apple Orchards in China. Eur. J. Plant Pathol. 2021, 161, 207–217. [Google Scholar] [CrossRef]
- Chatzidimopoulos, M.; Zambounis, A.; Lioliopoulou, F.; Vellios, E. Detection of Venturia inaequalis Isolates with Multiple Resistance in Greece. Microorganisms 2022, 10, 2354. [Google Scholar] [CrossRef] [PubMed]
- Nasonov, A.; Yakuba, G.; Marchenko, N.; Lobodina, E.; Astapchuk, I. Evaluation of Sensitivity of Apple Scab Pathogen to Difenoconazole Using the Discriminatory Dose Technique. BIO Web Conf. 2022, 47, 10002. [Google Scholar] [CrossRef]
- Hildebrand, P.D.; Lockhart, C.L.; Newbery, R.J.; Ross, R.G. Resistance of Venturia inaequalis to Bitertanol and Other Demethylation-Inhibiting Fungicides. Can. J. Plant Pathol. 1988, 10, 311–316. [Google Scholar] [CrossRef]
- Köller, W.; Wilcox, W.F.; Barnard, J.; Jones, A.L.; Braun, P.G. Detection and Quantification of Resistance of Venturia inaequalis Populations to Sterol Demethylation Inhibitors. Phytopathology 1997, 87, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Köller, W.; Wilcox, W.F. Interactive Effects of Dodine and the DMI Fungicide Fenarimol in the Control of Apple Scab. Plant Dis. 2000, 84, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Stammler, G.; Cordero, J.; Koch, A.; Semar, M.; Schlehuber, S. Role of the Y134F Mutation in CYP51 and Overexpression of CYP51 in the Sensitivity Response of Puccinia triticina to Epoxiconazole. Crop. Prot. 2009, 28, 891–897. [Google Scholar] [CrossRef]
- Becher, R.; Wirsel, S.G.R. Fungal Cytochrome P450 Sterol 14α-Demethylase (CYP51) and Azole Resistance in Plant and Human Pathogens. Appl. Microbiol. Biotechnol. 2012, 95, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, H.K.; Medeiros, C.-A.; Craig, I.R.; Stammler, G. Sensitivity of Phakopsora pachyrhizi towards Quinone-Outside-Inhibitors and Demethylation-Inhibitors, and Corresponding Resistance Mechanisms: Sensitivity and Resistance Mechanisms of P. pachyrhizi towards Fungicides. Pest. Manag. Sci. 2014, 70, 378–388. [Google Scholar] [CrossRef]
- Frenkel, O.; Cadle-Davidson, L.; Wilcox, W.F.; Milgroom, M.G. Mechanisms of Resistance to an Azole Fungicide in the Grapevine Powdery Mildew Fungus, Erysiphe necator. Phytopathology 2015, 105, 370–377. [Google Scholar] [CrossRef]
- Mair, W.; Lopez-Ruiz, F.; Stammler, G.; Clark, W.; Burnett, F.; Hollomon, D.; Ishii, H.; Thind, T.S.; Brown, J.K.; Fraaije, B.; et al. Proposal for a Unified Nomenclature for Target-site Mutations Associated with Resistance to Fungicides. Pest Manag. Sci. 2016, 72, 1449–1459. [Google Scholar] [CrossRef]
- Huf, A.; Rehfus, A.; Lorenz, K.H.; Bryson, R.; Voegele, R.T.; Stammler, G. Proposal for a New Nomenclature for CYP51 Haplotypes in Zymoseptoria tritici and Analysis of Their Distribution in Europe. Plant Pathol. 2018, 67, 1706–1712. [Google Scholar] [CrossRef]
- Muellender, M.M.; Mahlein, A.; Stammler, G.; Varrelmann, M. Evidence for the Association of Target-site Resistance in Cyp51 with Reduced DMI Sensitivity in European Cercospora beticola Field Isolates. Pest Manag. Sci. 2021, 77, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Cools, H.J.; Hawkins, N.J.; Fraaije, B.A. Constraints on the Evolution of Azole Resistance in Plant Pathogenic Fungi. Plant Pathol. 2013, 62, 36–42. [Google Scholar] [CrossRef]
- Yaegashi, H.; Hirayama, K.; Akahira, T.; Ito, T. Point Mutation in CYP51A1 of Venturia inaequalis Is Associated with Low Sensitivity to Sterol Demethylation Inhibitors. J. Gen. Plant Pathol. 2020, 86, 245–249. [Google Scholar] [CrossRef]
- Hoffmeister, M.; Zito, R.; Böhm, J.; Stammler, G. Mutations in CYP51 of Venturia inaequalis and Their Effects on DMI Sensitivity. J. Plant Dis. Prot. 2021, 128, 1467–1478. [Google Scholar] [CrossRef]
- Schnabel, G.; Jones, A.L. The 14α-Demethylasse (CYP51A1) Gene Is Overexpressed in Venturia inaequalis Strains Resistant to Myclobutanil. Phytopathology 2001, 91, 102–110. [Google Scholar] [CrossRef]
- Villani, S.M.; Hulvey, J.; Hily, J.-M.; Cox, K.D. Overexpression of the CYP51A1 Gene and Repeated Elements Are Associated with Differential Sensitivity to DMI Fungicides in Venturia inaequalis. Phytopathology 2016, 106, 562–571. [Google Scholar] [CrossRef]
- Cordero-Limon, L.; Shaw, M.W.; Passey, T.A.; Robinson, J.D.; Xu, X. Cross-resistance between Myclobutanil and Tebuconazole and the Genetic Basis of Tebuconazole Resistance in Venturia inaequalis. Pest Manag. Sci. 2021, 77, 844–850. [Google Scholar] [CrossRef]
- Xu, X.-M.; Gao, L.-Q.; Yang, J.-R. Are Insensitivities of Venturia inaequalis to Myclobutanil and Fenbuconazole Correlated? Crop. Prot. 2010, 29, 183–189. [Google Scholar] [CrossRef]
- Errampalli, D. Distribution of Myclobutanil Fungicide Sensitivities among Populations of Venturia inaequalis, the Causal Agent of Apple Scab, in Ontario. Acta Hortic. 2004, 638, 157–162. [Google Scholar] [CrossRef]
- Beresford, R.M.; Wright, P.J.; Wood, P.N.; Park, N.M.; Larsen, N.J.; Fisher, B.M. Resistance of Venturia inaequalis to Demethylation Inhibitor and Dodine Fungicides in Four New Zealand Applegrowing Regions. NZPP 2013, 66, 274–283. [Google Scholar] [CrossRef]
- Koller, W. Baseline Sensitivities of Venturia inaequalis to Sterol Demethylation Inhibitors. Plant Dis. 1991, 75, 726. [Google Scholar] [CrossRef]
- Pfeufer, E.E.; Ngugi, H.K. Orchard Factors Associated with Resistance and Cross Resistance to Sterol Demethylation Inhibitor Fungicides in Populations of Venturia inaequalis from Pennsylvania. Phytopathology 2012, 102, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Mapleson, D.; Garcia Accinelli, G.; Kettleborough, G.; Wright, J.; Clavijo, B.J. KAT: A K-Mer Analysis Toolkit to Quality Control NGS Datasets and Genome Assemblies. Bioinformatics 2017, 33, 574–576. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Passey, T.A.J.; Armitage, A.D.; Xu, X. Annotated Draft Genome Sequence of the Apple Scab Pathogen Venturia inaequalis. Microbiol. Resour. Announc. 2018, 7, e01062-18. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Stam, P. Construction of Integrated Genetic Linkage Maps by Means of a New Computer Package: Join Map. Plant J. 1993, 3, 739–744. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.W. A Model of the Evolution of Polygenically Controlled Fungicide Resistance. Plant Pathol. 1989, 38, 44–55. [Google Scholar] [CrossRef]
- Georgopoulos, S.G.; Skylakakis, G. Genetic Variability in the Fungi and the Problem of Fungicide Resistance. Crop. Prot. 1986, 5, 299–305. [Google Scholar] [CrossRef]
- Lendenmann, M.H.; Croll, D.; McDonald, B.A. QTL Mapping of Fungicide Sensitivity Reveals Novel Genes and Pleiotropy with Melanization in the Pathogen Zymoseptoria tritici. Fungal Genet. Biol. 2015, 80, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Nakaune, R.; Adachi, K.; Nawata, O.; Tomiyama, M.; Akutsu, K.; Hibi, T. A Novel ATP-Binding Cassette Transporter Involved in Multidrug Resistance in the Phytopathogenic Fungus Penicillium digitatum. Appl. Environ. Microbiol. 1998, 64, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Anderson, J.B.; Kohn, L.M. Evolution of Drug Resistance in Candida albicans. Annu. Rev. Microbiol. 2002, 56, 139–165. [Google Scholar] [CrossRef]
- Joseph-Home, T.; Manning, N.J.; Hollomon, D.; Kelly, S.L. Defective Sterol Δ 5(6) Desaturase as a Cause of Azole Resistance in Ustilago maydis. FEMS Microbiol. Lett. 1995, 127, 29–34. [Google Scholar] [CrossRef]
- Bolton, M.D.; Ebert, M.K.; Faino, L.; Rivera-Varas, V.; de Jonge, R.; Van de Peer, Y.; Thomma, B.P.H.J.; Secor, G.A. RNA-Sequencing of Cercospora beticola DMI-Sensitive and -Resistant Isolates after Treatment with Tetraconazole Identifies Common and Contrasting Pathway Induction. Fungal Genet. Biol. 2016, 92, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Lindquist, S. Hsp90 Potentiates the Rapid Evolution of New Traits: Drug Resistance in Diverse Fungi. Science 2005, 309, 2185–2189. [Google Scholar] [CrossRef] [PubMed]
- Mellado, E.; Diaz-Guerra, T.M.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Identification of Two Different 14-α Sterol Demethylase-Related Genes (cyp51A and cyp51B) in Aspergillus fumigatus and Other Aspergillus Species. J. Clin. Microbiol. 2001, 39, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Riaz, S.; Morales-Cruz, A.; Amrine, K.C.; McGuire, B.; Gubler, W.D.; Walker, M.A.; Cantu, D. Adaptive Genomic Structural Variation in the Grape Powdery Mildew Pathogen, Erysiphe necator. BMC Genom. 2014, 15, 1081. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, H.; Hasegawa, K.; Nakaune, R.; Lee, Y.J.; Makizumi, Y.; Akutsu, K.; Hibi, T. Tandem Repeat of a Transcriptional Enhancer Upstream of the Sterol 14α-Demethylase Gene (CYP51) in Penicillium digitatum. Appl. Environ. Microbiol. 2000, 66, 3421–3426. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Proffer, T.J.; Jacobs, J.L.; Sundin, G.W. Overexpression of the 14α-Demethylase Target Gene (CYP51) Mediates Fungicide Resistance in Blumeriella jaapii. Appl. Environ. Microbiol. 2006, 72, 2581–2585. [Google Scholar] [CrossRef]
- Nikou, D.; Malandrakis, A.; Konstantakaki, M.; Vontas, J.; Markoglou, A.; Ziogas, B. Molecular Characterization and Detection of Overexpressed C-14 Alpha-Demethylase-Based DMI Resistance in Cercospora beticola Field Isolates. Pestic. Biochem. Physiol. 2009, 95, 18–27. [Google Scholar] [CrossRef]
- Luo, C.-X.; Schnabel, G. The Cytochrome P450 Lanosterol 14α-Demethylase Gene Is a Demethylation Inhibitor Fungicide Resistance Determinant in Monilinia fructicola Field Isolates from Georgia. Appl. Environ. Microbiol. 2008, 74, 359–366. [Google Scholar] [CrossRef]
- Cools, H.J.; Bayon, C.; Atkins, S.; Lucas, J.A.; Fraaije, B.A. Overexpression of the Sterol 14α-Demethylase Gene (MgCYP51) in Mycosphaerella graminicola Isolates Confers a Novel Azole Fungicide Sensitivity Phenotype. Pest Manag. Sci. 2012, 68, 1034–1040. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Q.; Ruan, R.; Zhang, T.; Zhu, C.; Li, H. PdMLE1, a Specific and Active Transposon Acts as a Promoter and Confers Penicillium digitatum with DMI Resistance. Environ. Microbiol. Rep. 2013, 5, 135–142. [Google Scholar] [CrossRef]
- de Waard, M.A.; Andrade, A.C.; Hayashi, K.; Schoonbeek, H.; Stergiopoulos, I.; Zwiers, L.-H. Impact of Fungal Drug Transporters on Fungicide Sensitivity, Multidrug Resistance and Virulence. Pest Manag. Sci. 2006, 62, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Schoonbeek, H.; De Waard, M.A. Expression of the ABC Transporter BcatrD from Botrytis cinerea Reduces Sensitivity to Sterol Demethylation Inhibitor Fungicides. Pestic. Biochem. Physiol. 2002, 73, 110–121. [Google Scholar] [CrossRef]
- Hulvey, J.; Popko, J.T.; Sang, H.; Berg, A.; Jung, G. Overexpression of ShCYP51B and ShatrD in Sclerotinia homoeocarpa Isolates Exhibiting Practical Field Resistance to a Demethylation Inhibitor Fungicide. Appl. Environ. Microbiol. 2012, 78, 6674–6682. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Zwiers, L.-H.; De Waard, M.A. Secretion of Natural and Synthetic Toxic Compounds from Filamentous Fungi by Membrane Transporters of the ATP-Binding Cassette and Major Facilitator Superfamily. Eur. J. Plant Pathol. 2002, 108, 719–734. [Google Scholar] [CrossRef]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-Mediated Antifungal Drug Resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.-S.; Fillinger, S.; Mernke, D.; Schoonbeek, H.-J.; Pradier, J.-M.; Leroux, P.; De Waard, M.A.; et al. Fungicide-Driven Evolution and Molecular Basis of Multidrug Resistance in Field Populations of the Grey Mould Fungus Botrytis cinerea. PLoS Pathog. 2009, 5, e1000696. [Google Scholar] [CrossRef] [PubMed]
- Leroch, M.; Plesken, C.; Weber, R.W.S.; Kauff, F.; Scalliet, G.; Hahn, M. Gray Mold Populations in German Strawberry Fields Are Resistant to Multiple Fungicides and Dominated by a Novel Clade Closely Related to Botrytis cinerea. Appl. Environ. Microbiol. 2013, 79, 159–167. [Google Scholar] [CrossRef]
- Grabke, A.; Stammler, G. A Botrytis cinerea Population from a Single Strawberry Field in Germany Has a Complex Fungicide Resistance Pattern. Plant Dis. 2015, 99, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Omrane, S.; Sghyer, H.; Audéon, C.; Lanen, C.; Duplaix, C.; Walker, A.-S.; Fillinger, S. Fungicide Efflux and the MgMFS1 Transporter Contribute to the Multidrug Resistance Phenotype in Zymoseptoria tritici Field Isolates: Fungicide Efflux & MgMFS 1 Contribute to MDR in Z. tritici. Environ. Microbiol. 2015, 17, 2805–2823. [Google Scholar] [CrossRef]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; Sklar, L.A.; Cannon, R.D. Targeting Efflux Pumps to Overcome Antifungal Drug Resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef]
- Palani, P.V.; Lalithakumari, D. Resistance of Venturia inaequalis to the Sterol Biosynthesis-Inhibiting Fungicide, Penconazole [1-(2-(2,4-Dichlorophenyl) Pentyl)-1H-1,2,4-Triazole]. Mycol. Res. 1999, 103, 1157–1164. [Google Scholar] [CrossRef]
- Koopman, T.A.; Meitz-Hopkins, J.C.; Tobutt, K.R.; Bester, C.; Lennox, C.L. Pathogenicity and Virulence of South African Isolates of Venturia inaequalis. Eur. J. Plant Pathol. 2022, 164, 45–58. [Google Scholar] [CrossRef]
- Corkley, I.; Fraaije, B.; Hawkins, N. Fungicide Resistance Management: Maximizing the Effective Life of Plant Protection Products. Plant Pathol. 2022, 71, 150–169. [Google Scholar] [CrossRef]
- Van Den Bosch, F.; Paveley, N.; Van Den Berg, F.; Hobbelen, P.; Oliver, R. Mixtures as a Fungicide Resistance Management Tactic. Phytopathology 2014, 104, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wei, J.; Fu, L.; Wang, S.; Liu, J.; Guo, Q.; Jiang, J.; Tian, Y.; Che, Z.; Chen, G.; et al. Tebuconazole Resistance of Fusarium graminearum Field Populations from Wheat in Henan Province. J. Phytopathol. 2021, 169, 525–532. [Google Scholar] [CrossRef]
- Keinath, A.P.; Rennberger, G.; Wechter, P. Widespread Resistance to Tebuconazole and Cross-Resistance to Other DMI Fungicides in Stagonosporopsis Citrulli Isolated from Watermelon in South Carolina. Plant Dis. 2023. PDIS-03-23-0478-RE. [Google Scholar] [CrossRef] [PubMed]
- Beckerman, J.L.; Sundin, G.W.; Rosenberger, D.A. Do Some IPM Concepts Contribute to the Development of Fungicide Resistance? Lessons Learned from the Apple Scab Pathosystem in the United States. Pest Manag. Sci. 2015, 71, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Franco Ortega, S.; Prencipe, S.; Gullino, M.L.; Spadaro, D. New Molecular Tool for a Quick and Easy Detection of Apple Scab in the Field. Agronomy 2020, 10, 581. [Google Scholar] [CrossRef]
- Gadoury, D.M.; Seem, R.C.; MacHardy, W.E.; Wilcox, W.F.; Rosenberger, D.A.; Stensvand, A. A Comparison of Methods Used to Estimate the Maturity and Release of Ascospores of Venturia inaequalis. Plant Dis. 2004, 88, 869–874. [Google Scholar] [CrossRef]
- Meitz-Hopkins, J.C.; Von Diest, S.G.; Koopman, T.A.; Bahramisharif, A.; Lennox, C.L. A Method to Monitor Airborne Venturia inaequalis Ascospores Using Volumetric Spore Traps and Quantitative PCR. Eur. J. Plant Pathol. 2014, 140, 527–541. [Google Scholar] [CrossRef]
- Daniëls, B.; De Landtsheer, A.; Dreesen, R.; Davey, M.W.; Keulemans, J. Real-Time PCR as a Promising Tool to Monitor Growth of Venturia spp. in Scab-Susceptible and -Resistant Apple Leaves. Eur. J. Plant Pathol. 2012, 134, 821–833. [Google Scholar] [CrossRef]
- Prencipe, S.; Sillo, F.; Garibaldi, A.; Gullino, M.L.; Spadaro, D. Development of a Sensitive TaqMan qPCR Assay for Detection and Quantification of Venturia inaequalis in Apple Leaves and Fruit and in Air Samples. Plant Dis. 2020, 104, 2851–2859. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.R.; Godfray, H.C.J.; Burt, A. Sex Increases the Efficacy of Natural Selection in Experimental Yeast Populations. Nature 2005, 434, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P. Address in Pathology, ON CHEMIOTHERAPY: Delivered before the Seventeenth International Congress of Medicine. BMJ 1913, 2, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance in Crop Pathogens: How Can It Be Managed? In FRAC Monograph, 2nd ed.; Fungicide Resistance Action Committee: Brussels, Belgium, 2007; ISBN 90-72398-07-6. [Google Scholar]
- Van Den Bosch, F.; Paveley, N.; Shaw, M.; Hobbelen, P.; Oliver, R. The Dose Rate Debate: Does the Risk of Fungicide Resistance Increase or Decrease with Dose? The Dose Rate Debate. Plant Pathol. 2011, 60, 597–606. [Google Scholar] [CrossRef]
- Koh, H.S.; Sohn, S.H.; Lee, Y.S.; Koh, Y.J.; Song, J.H.; Jung, J.S. Specific and Sensitive Detection of Venturia nashicola, the Scab Fungus of Asian Pears, by Nested PCR. Plant Pathol. J. 2013, 29, 357–363. [Google Scholar] [CrossRef]
- Gutierrez Vazquez, Y.; Adams, I.P.; McGreig, S.; Walshaw, J.; van den Berg, F.; Sanderson, R.; Pufal, H.; Conyers, C.; Langton, D.; Broadhead, R.; et al. Profiling Azole Resistant Haplotypes within Zymoseptoria tritici Populations Using Nanopore Sequencing. Front. Agron. 2022, 4, 943440. [Google Scholar] [CrossRef]
- Giolai, M.; Verweij, W.; Pearson, N.; Nicholson, P.; Leggett, R.M.; Clark, M.D. Air-Seq: Measuring Air Metagenomic Diversity in an Agricultural Ecosystem. bioRxiv 2022. [Google Scholar] [CrossRef]
- van den Bosch, F.; Fraaije, B.; Oliver, R.; van den Berg, F.; Paveley, N. The Use of Mathematical Models to Guide Fungicide Resistance Management Decisions. In Fungicide Resistance in Plant Pathogens; Ishii, H., Hollomon, D.W., Eds.; Springer: Tokyo, Japan, 2015; pp. 49–62. ISBN 978-4-431-55641-1. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).