
Strategies to Increase the Value of Pomaces with Fermentation


 ,
,  ,
, 
 ,
, 

Abstract
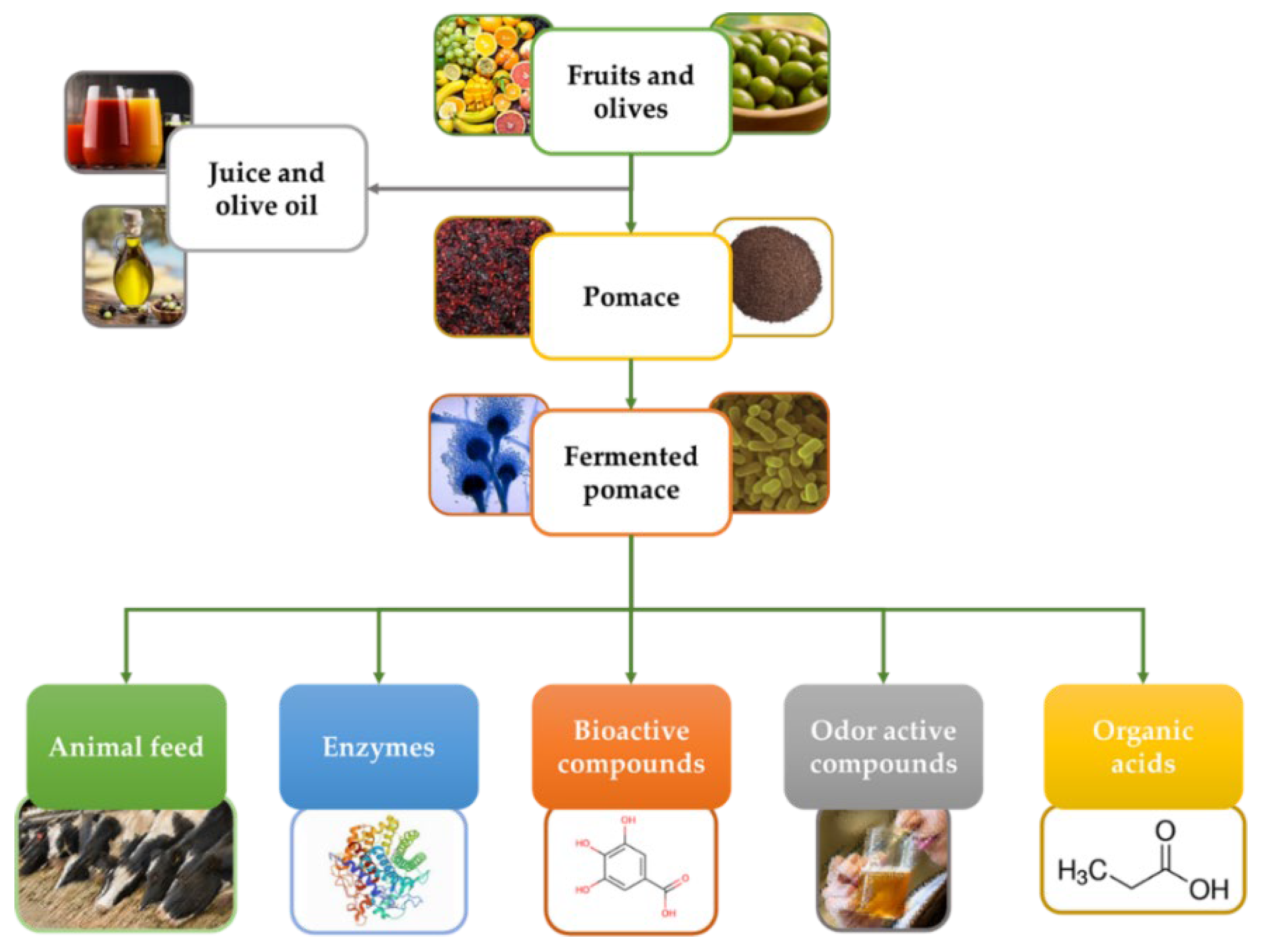
:1. Introduction
2. Utilization in Silage or as a Feed Supplement for Animal Production
3. Enzyme Production and Potential Applications
4. Release and Production of Bioactive Compounds
5. Production of Organic Acids
6. Production of Bioflavors
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bordiga, M.; Travaglia, F.; Locatelli, M. Valorisation of grape pomace: An approach that is increasingly reaching its maturity—A review. Int. J. Food Sci. Technol. 2019, 54, 933–942. [Google Scholar] [CrossRef]
- Gassara, F.; Brar, S.K.; Pelletier, F.; Verma, M.; Godbout, S.; Tyagi, R.D. Pomace waste management scenarios in Québec-Impact on greenhouse gas emissions. J. Hazard. Mater. 2011, 192, 1178–1185. [Google Scholar] [CrossRef]
- Difonzo, G.; Troilo, M.; Squeo, G.; Pasqualone, A.; Caponio, F. Functional compounds from olive pomace to obtain high-added value foods—A review. J. Sci. Food Agric. 2021, 101, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Perussello, C.A.; Zhang, Z.; Marzocchella, A.; Tiwari, B.K. Valorization of Apple Pomace by Extraction of Valuable Compounds. Compr. Rev. Food Sci. Food Saf. 2017, 16, 776–796. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.; Wang, Z.; Feng, Y.; Kim, Y.T.; Stewart, A.C.; O’Keefe, S.F.; Neilson, A.P.; He, Z.; Huang, H. Grape Pomace And Its Secondary Waste Management: Biochar Production for a Broad Range of Lead (Pb) Removal From Water. Environ. Res. 2020, 186, 109442. [Google Scholar] [CrossRef]
- IMARC Fruit Juice Market Size, Share, Trends & Forecast 2021–2026. Available online: https://www.imarcgroup.com/fruit-juice-manufacturing-plant (accessed on 13 October 2021).
- Statista Inc. Juices—Worldwide. Available online: https://www.statista.com/outlook/cmo/non-alcoholic-drinks/juices/worldwide (accessed on 13 October 2021).
- European Commission EU Agricultural Outlook 2019-30: Olive Oil Consumption to Rise in Non-Producing EU Countries. Available online: https://ec.europa.eu/info/news/eu-agricultural-outlook-2019-2030-olive-oil-consumption-rise-non-producing-eu-countries-2019-dec-10_en (accessed on 13 October 2021).
- Gavahian, M.; Mousavi Khaneghah, A.; Lorenzo, J.M.; Munekata, P.E.S.; Garcia-Mantrana, I.; Collado, M.C.; Meléndez-Martínez, A.J.; Barba, F.J. Health benefits of olive oil and its components: Impacts on gut microbiota antioxidant activities, and prevention of noncommunicable diseases. Trends Food Sci. Technol. 2019, 88, 220–227. [Google Scholar] [CrossRef]
- Jimenez-Lopez, C.; Carpena, M.; Lourenço-Lopes, C.; Gallardo-Gomez, M.; Lorenzo, J.M.; Barba, F.J.; Prieto, M.A.; Simal-Gandara, J. Bioactive Compounds and Quality of Extra Virgin Olive Oil. Foods 2020, 9, 1014. [Google Scholar] [CrossRef]
- Skinner, R.C.; Gigliotti, J.C.; Ku, K.M.; Tou, J.C. A comprehensive analysis of the composition, health benefits, and safety of apple pomace. Nutr. Rev. 2018, 76, 893–909. [Google Scholar] [CrossRef]
- Bobinaitė, R.; Grootaert, C.; Van Camp, J.; Šarkinas, A.; Liaudanskas, M.; Žvikas, V.; Viškelis, P.; Rimantas Venskutonis, P. Chemical composition, antioxidant, antimicrobial and antiproliferative activities of the extracts isolated from the pomace of rowanberry (Sorbus aucuparia L.). Food Res. Int. 2020, 136, 109310. [Google Scholar] [CrossRef] [PubMed]
- Abbasi-Parizad, P.; De Nisi, P.; Adani, F.; Sciarria, T.P.; Squillace, P.; Scarafoni, A.; Iametti, S.; Scaglia, B. Antioxidant and anti-inflammatory activities of the crude extracts of raw and fermented tomato pomace and their correlations with aglycate-polyphenols. Antioxidants 2020, 9, 179. [Google Scholar] [CrossRef] [Green Version]
- Costabile, G.; Vitale, M.; Luongo, D.; Naviglio, D.; Vetrani, C.; Ciciola, P.; Tura, A.; Castello, F.; Mena, P.; Del Rio, D.; et al. Grape pomace polyphenols improve insulin response to a standard meal in healthy individuals: A pilot study. Clin. Nutr. 2019, 38, 2727–2734. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.A.; Gómez-García, R.; Vilas-Boas, A.A.; Madureira, A.R.; Pintado, M.M. Management of Fruit Industrial By-Products—A case Study on Circular Economy Approach. Molecules 2020, 25, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Union The EU Green Deal—A Roadmap to Sustainable Economies. Available online: https://bit.ly/39ydb5i (accessed on 1 December 2021).
- Circular Economy Action Plan. Available online: https://ec.europa.eu/environment/strategy/circular-economy-action-plan_pt (accessed on 1 December 2021).
- Munekata, P.E.S.; Nieto, G.; Pateiro, M.; Lorenzo, J.M. Phenolic compounds obtained from Olea europaea by-products and their use to improve the quality and shelf life of meat and meat products—A review. Antioxidants 2020, 9, 1061. [Google Scholar] [CrossRef] [PubMed]
- Das, A.K.; Nanda, P.K.; Chowdhury, N.R.; Dandapat, P.; Gagaoua, M.; Chauhan, P.; Pateiro, M.; Lorenzo, J.M. Application of Pomegranate by-Products in Muscle Foods: Oxidative Indices, Colour Stability, Shelf Life and Health Benefits. Molecules 2021, 26, 467. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Gullón, P.; Pateiro, M.; Munekata, P.E.S.; Zhang, W.; Lorenzo, J.M. Tomato as potential source of natural additives for meat industry. A review. Antioxidants 2020, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizael, W.C.F.; Costa, R.G.; Cruz, G.R.B.; de Carvalho, F.F.R.; Ribeiro, N.L.; Lima, A.; Domínguez, R.; Lorenzo, J.M. Effect of the use of tomato pomace on feeding and performance of lactating goats. Animals 2020, 10, 1574. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Martino, G. Dietary grape pomace supplementation in dairy cows: Effect on nutritional quality of milk and its derived dairy products. Foods 2020, 9, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiles, A.; Campbell, G.M.; Struck, S.; Rohm, H.; Hernando, I. Fiber from fruit pomace: A review of applications in cereal-based products. Food Rev. Int. 2018, 34, 162–181. [Google Scholar] [CrossRef]
- Gullón, P.; Gullón, B.; Romaní, A.; Rocchetti, G.; Lorenzo, J.M. Smart advanced solvents for bioactive compounds recovery from agri-food by-products: A review. Trends Food Sci. Technol. 2020, 101, 182–197. [Google Scholar] [CrossRef]
- Cattaneo, C.; Lavelli, V.; Proserpio, C.; Laureati, M.; Pagliarini, E. Consumers’ attitude towards food by-products: The influence of food technology neophobia, education and information. Int. J. Food Sci. Technol. 2019, 54, 679–687. [Google Scholar] [CrossRef]
- Perito, M.A.; Di Fonzo, A.; Sansone, M.; Russo, C. Consumer acceptance of food obtained from olive by-products: A survey of Italian consumers. Br. Food J. 2020, 122, 212–226. [Google Scholar] [CrossRef]
- Sabater, C.; Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Margolles, A. Valorization of Vegetable Food Waste and By-Products Through Fermentation Processes. Front. Microbiol. 2020, 11, 2604. [Google Scholar] [CrossRef] [PubMed]
- Koutinas, A.A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Lopez Garcia, I.; Kookos, I.K.; Papanikolaou, S.; Kwan, T.H.; Lin, C.S.K. Valorization of industrial waste and by-product streams via fermentation for the production of chemicals and biopolymers. Chem. Soc. Rev. 2014, 43, 2587–2627. [Google Scholar] [CrossRef]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Xia, G.; Cao, Y. Effects of replacing commercial material with apple pomace on the fermentation quality of total mixed ration silage and its digestibility, nitrogen balance and rumen fermentation in wethers. Grassl. Sci. 2020, 66, 124–131. [Google Scholar] [CrossRef]
- Xia, G.-J.; Fang, J.-C. Intake, digestibility and rumen fermentation pattern in wethers fed total mixed ration silage containing dry or fresh apple pomace. J. Anim. Feed Sci. 2021, 30, 26–32. [Google Scholar] [CrossRef]
- Belém, C.d.S.; de Souza, A.M.; de Lima, P.R.; de Carvalho, F.A.L.; Queiroz, M.A.Á.; da Costa, M.M. Digestibility, fermentation and microbiological characteristics of Calotropis procera silage with different quantities of grape pomace. Cienc. Agrotecnologia 2016, 40, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Shen, Y.; You, M.; Zhang, Y.; Yan, J.; Li, D.; Bai, S. Effect of grape pomace on fermentation quality and aerobic stability of sweet sorghum silage. Anim. Sci. J. 2017, 88, 1523–1530. [Google Scholar] [CrossRef]
- Köksal, Y.; Bölükbaş, B.; Selçuk, Z. An in vitro Evaluation of the Silages in White Mulberry Pomace/Meadow Grass mixtures containing different levels of White Mulberry Pomace. Kocatepe Vet. J. 2021, 14, 309–315. [Google Scholar] [CrossRef]
- Fang, J.; Cao, Y.; Matsuzaki, M.; Suzuki, H.; Kimura, H. Effects of apple pomace-mixed silage on growth performance and meat quality in finishing pigs. Anim. Sci. J. 2016, 87, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- Davies, S.J.; Guroy, D.; Hassaan, M.S.; El-Ajnaf, S.M.; El-Haroun, E. Evaluation of co-fermented apple-pomace, molasses and formic acid generated sardine based fish silages as fishmeal substitutes in diets for juvenile European sea bass (Dicentrachus labrax) production. Aquaculture 2020, 521, 735087. [Google Scholar] [CrossRef]
- Massaro Junior, F.L.; Bumbieris Junior, V.H.; Pereira, E.S.; Zanin, E.; Horst, E.H.; Calixto, O.P.P.; Peixoto, E.L.T.; Galbeiro, S.; Mizubuti, I.Y. Grape pomace silage on growth performance, carcass, and meat quality attributes of lambs. Sci. Agric. 2022, 79, 2022. [Google Scholar] [CrossRef]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I.; et al. Grape pomace improves performance, antioxidant status, fecal microbiota and meat quality of piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gungor, E.; Altop, A.; Erener, G. Effect of raw and fermented grape pomace on the growth performance, antioxidant status, intestinal morphology, and selected bacterial species in broiler chicks. Animals 2021, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Moustafa, A.; Shahin, S.E.; Sherief, W.R.I.A.; Abdallah, K.; Farag, M.F.M.; Nassan, M.A.; Ibrahim, S.M. Impact of Fermented or Enzymatically Fermented Dried Olive Pomace on Growth, Expression of Digestive Enzyme and Glucose Transporter Genes, Oxidative Stability of Frozen Meat, and Economic Efficiency of Broiler Chickens. Front. Vet. Sci. 2021, 8, 644325. [Google Scholar] [CrossRef] [PubMed]
- Tuoxunjiang, H.; Yimamu, A.; Li, X.Q.; Maimaiti, R.; Wang, Y.L. Effect of ensiled tomato pomace on performance and antioxidant status in the peripartum dairy cow. J. Anim. Feed Sci. 2020, 29, 105–114. [Google Scholar] [CrossRef]
- Gungor, E.; Altop, A.; Erener, G.; Coskun, I. Effect of raw and fermented pomegranate pomace on performance, antioxidant activity, intestinal microbiota and morphology in broiler chickens. Arch. Anim. Nutr. 2021, 75, 137–152. [Google Scholar] [CrossRef]
- Siró, I.; Kápolna, E.; Kápolna, B.; Lugasi, A. Functional food. Product development, marketing and consumer acceptance-A review. Appetite 2008, 51, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Sharma, N.; Handa, S. Utilization of horticultural waste (Apple Pomace) for multiple carbohydrase production from Rhizopus delemar F2 under solid state fermentation. J. Genet. Eng. Biotechnol. 2018, 16, 181–189. [Google Scholar] [CrossRef]
- Satapathy, S.; Soren, J.P.; Mondal, K.C.; Srivastava, S.; Pradhan, C.; Sahoo, S.L.; Thatoi, H.; Rout, J.R. Industrially relevant pectinase production from Aspergillus parvisclerotigenus KX928754 using apple pomace as the promising substrate. J. Taibah Univ. Sci. 2021, 15, 347–356. [Google Scholar] [CrossRef]
- Kuvvet, C.; Uzuner, S.; Cekmecelioglu, D. Improvement of Pectinase Production by Co-culture of Bacillus spp. Using Apple Pomace as a Carbon Source. Waste Biomass Valorization 2019, 10, 1241–1249. [Google Scholar] [CrossRef]
- Guleria, S.; Walia, A.; Chauhan, A.; Mahajan, R.; Shirkot, C.K. Mutagenesis of Alkalophilic Cellulosimicrobium sp. CKMX1 for Hyper-Production of Cellulase-Free Xylanase in Solid State Fermentation of Apple Pomace. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 85, 241–252. [Google Scholar] [CrossRef]
- Gassara, F.; Brar, S.K.; Tyagi, R.D.; Verma, M.; Surampalli, R.Y. Screening of agro-industrial wastes to produce ligninolytic enzymes by Phanerochaete chrysosporium. Biochem. Eng. J. 2010, 49, 388–394. [Google Scholar] [CrossRef]
- Singh, R.S.; Chauhan, K.; Kaur, K.; Pandey, A. Statistical optimization of solid-state fermentation for the production of fungal inulinase from apple pomace. Bioresour. Technol. Rep. 2020, 9, 100364. [Google Scholar] [CrossRef]
- Meini, M.R.; Ricardi, L.L.; Romanini, D. Novel Routes for Valorisation of Grape Pomace Through the Production of Bioactives by Aspergillus niger. Waste Biomass Valorization 2020, 11, 6047–6055. [Google Scholar] [CrossRef]
- Kurt, A.S.; Cekmecelioglu, D. Bacterial cellulase production using grape pomace hydrolysate by shake-flask submerged fermentation. Biomass Convers. Biorefinery 2021, 1, 1–8. [Google Scholar] [CrossRef]
- Papadaki, A.; Kachrimanidou, V.; Papanikolaou, S.; Philippoussis, A.; Diamantopoulou, P. Upgrading grape pomace through Pleurotus spp. cultivation for the production of enzymes and fruiting bodies. Microorganisms 2019, 7, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teles, A.S.C.; Chávez, D.W.H.; Oliveira, R.A.; Bon, E.P.S.; Terzi, S.C.; Souza, E.F.; Gottschalk, L.M.F.; Tonon, R.V. Use of grape pomace for the production of hydrolytic enzymes by solid-state fermentation and recovery of its bioactive compounds. Food Res. Int. 2019, 120, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, E.; Kontogiannopoulos, K.N.; Assimopoulou, A.N.; Mantzouridou, F.T. Feasibility of multi-hydrolytic enzymes production from optimized grape pomace residues and wheat bran mixture using Aspergillus niger in an integrated citric acid-enzymes production process. Bioresour. Technol. 2020, 309, 123317. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.E.; Fathy, S.A.; Rashad, M.M.; Ezz, M.K.; Mohammed, A.T. Purification and characterization of a novel tannase produced by Kluyveromyces marxianus using olive pomace as solid support, and its promising role in gallic acid production. Int. J. Biol. Macromol. 2018, 107, 2342–2350. [Google Scholar] [CrossRef]
- Leite, P.; Salgado, J.M.; Venâncio, A.; Domínguez, J.M.; Belo, I. Ultrasounds pretreatment of olive pomace to improve xylanase and cellulase production by solid-state fermentation. Bioresour. Technol. 2016, 214, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, F.; Salgado, J.M.; Abrunhosa, L.; Pérez-Rodríguez, N.; Domínguez, J.M.; Venâncio, A.; Belo, I. Optimization of lipase production by solid-state fermentation of olive pomace: From flask to laboratory-scale packed-bed bioreactor. Bioprocess Biosyst. Eng. 2017, 40, 1123–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, A.; Chalima, A.; Topakas, E. Biorefinery of exhausted olive pomace through the production of polygalacturonases and omega-3 fatty acids by Crypthecodinium cohnii. Algal Res. 2021, 59, 102470. [Google Scholar] [CrossRef]
- Belmessikh, A.; Boukhalfa, H.; Mechakra-Maza, A.; Gheribi-Aoulmi, Z.; Amrane, A. Statistical optimization of culture medium for neutral protease production by Aspergillus oryzae. Comparative study between solid and submerged fermentations on tomato pomace. J. Taiwan Inst. Chem. Eng. 2013, 44, 377–385. [Google Scholar] [CrossRef]
- Boukhalfa-lezzar, H.; Copinet, E.; Duchiron, F.; Aicha, M. Utilization of tomato pomace as a substrate for neutral protease production by Aspergillus oryzae 2220 on solid-state fermentation. Artic. Int. J. Adv. Res. 2014, 2, 338–346. [Google Scholar]
- Iandolo, D.; Piscitelli, A.; Sannia, G.; Faraco, V. Enzyme production by solid substrate fermentation of pleurotus ostreatus and trametes versicolor on tomato pomace. Appl. Biochem. Biotechnol. 2011, 163, 40–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torkashvand, N.; Mousivand, M.; Hashemi, M. Canola meal and tomato pomace as novel substrates for production of thermostable Bacillus subtilis T4b xylanase with unique properties. Biomass Convers. Biorefinery 2020, 1–13. [Google Scholar] [CrossRef]
- Mahmoodi, M.; Najafpour, G.D.; Mohammadi, M. Production of pectinases for quality apple juice through fermentation of orange pomace. J. Food Sci. Technol. 2017, 54, 4123–4128. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodi, M.; Najafpour, G.D.; Mohammadi, M. Bioconversion of agroindustrial wastes to pectinases enzyme via solid state fermentation in trays and rotating drum bioreactors. Biocatal. Agric. Biotechnol. 2019, 21, 101280. [Google Scholar] [CrossRef]
- Singh, R.S.; Chauhan, K.; Singh, J.; Pandey, A.; Larroche, C. Solid-state fermentation of carrot pomace for the production of inulinase by Penicillium oxalicum BGPUP-4. Food Technol. Biotechnol. 2018, 56, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Dey, T.B.; Banerjee, R. Application of decolourized and partially purified polygalacturonase and α-amylase in apple juice clarification. Brazilian J. Microbiol. 2014, 45, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Cuprys, A.; Thomson, P.; Suresh, G.; Roussi, T.; Brar, S.K.; Drogui, P. Potential of agro-industrial produced laccase to remove ciprofloxacin. Environ. Sci. Pollut. Res. 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of microbial enzymes in food industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial applications of enzymes: Recent advances, techniques, and outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Gligor, O.; Mocan, A.; Moldovan, C.; Locatelli, M.; Crișan, G.; Ferreira, I.C.F.R. Enzyme-assisted extractions of polyphenols—A comprehensive review. Trends Food Sci. Technol. 2019, 88, 302–315. [Google Scholar] [CrossRef]
- Dzah, C.S.; Duan, Y.; Zhang, H.; Serwah Boateng, N.A.; Ma, H. Latest developments in polyphenol recovery and purification from plant by-products: A review. Trends Food Sci. Technol. 2020, 99, 375–388. [Google Scholar] [CrossRef]
- Zambrano, C.; Kotogán, A.; Bencsik, O.; Papp, T.; Vágvölgyi, C.; Mondal, K.C.; Krisch, J.; Takó, M. Mobilization of phenolic antioxidants from grape, apple and pitahaya residues via solid state fungal fermentation and carbohydrase treatment. LWT Food Sci. Technol. 2018, 89, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Dulf, F.V.; Vodnar, D.C.; Toşa, M.I.; Dulf, E.H. Simultaneous enrichment of grape pomace with γ-linolenic acid and carotenoids by solid-state fermentation with Zygomycetes fungi and antioxidant potential of the bioprocessed substrates. Food Chem. 2020, 310, 125927. [Google Scholar] [CrossRef] [PubMed]
- Bucić-Kojić, A.; Fernandes, F.; Silva, T.; Planinić, M.; Tišma, M.; Šelo, G.; Šibalić, D.; Pereira, D.M.; Andrade, P.B. Enhancement of the anti-inflammatory properties of grape pomace treated by: Trametes versicolor. Food Funct. 2020, 11, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Lohani, U.C.; Muthukumarappan, K. Effect of sequential treatments of fermentation and ultrasonication followed by extrusion on bioactive content of apple pomace and textural, functional properties of its extrudates. Int. J. Food Sci. Technol. 2016, 51, 1811–1819. [Google Scholar] [CrossRef]
- Madrera, R.R.; Pando Bedriñana, R.; Suárez Valles, B. Enhancement of the nutritional properties of apple pomace by fermentation with autochthonous yeasts. LWT Food Sci. Technol. 2017, 79, 27–33. [Google Scholar] [CrossRef]
- Liu, L.; You, Y.; Deng, H.; Guo, Y.; Meng, Y. Promoting hydrolysis of apple pomace by pectinase and cellulase to produce microbial oils using engineered Yarrowia lipolytica. Biomass Bioenergy 2019, 126, 62–69. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Toşa, M.I. Total Phenolic Contents, Antioxidant Activities, and Lipid Fractions from Berry Pomaces Obtained by Solid-State Fermentation of Two Sambucus Species with Aspergillus niger. J. Agric. Food Chem. 2015, 63, 3489–3500. [Google Scholar] [CrossRef] [PubMed]
- Fathy, S.A.; Mahmoud, A.E.; Rashad, M.M.; Ezz, M.K.; Mohammed, A.T. Improving the nutritive value of olive pomace by solid state fermentation of Kluyveromyces marxianus with simultaneous production of gallic acid. Int. J. Recycl. Org. Waste Agric. 2018, 7, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Paz, A.; Karnaouri, A.; Templis, C.C.; Papayannakos, N.; Topakas, E. Valorization of exhausted olive pomace for the production of omega-3 fatty acids by Crypthecodinium cohnii. Waste Manag. 2020, 118, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Diaconeasa, Z.; Socaciu, C. Liberation and recovery of phenolic antioxidants and lipids in chokeberry (Aronia melanocarpa) pomace by solid-state bioprocessing using Aspergillus niger and Rhizopus oligosporus strains. LWT 2018, 87, 241–249. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Socaciu, C. Effects of solid-state fermentation with two filamentous fungi on the total phenolic contents, flavonoids, antioxidant activities and lipid fractions of plum fruit (Prunus domestica L.) by-products. Food Chem. 2016, 209, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Pintea, A. Phenolic compounds, flavonoids, lipids and antioxidant potential of apricot (Prunus armeniaca L.) pomace fermented by two filamentous fungal strains in solid state system. Chem. Cent. J. 2017, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Jiang, J.; Yue, Y.; Feng, Z.; Chen, J.; Ye, X. Influence of mixed probiotics on the the bioactive composition, antioxidant activity and appearance of fermented red bayberry pomace. LWT 2020, 133, 110076. [Google Scholar] [CrossRef]
- Cheng, Y.; Wu, T.; Tang, S.; Liang, F.; Fang, Y.; Cao, W.; Pan, S.; Xu, X. Fermented blueberry pomace ameliorates intestinal barrier function through the NF-κB-MLCK signaling pathway in high-fat diet mice. Food Funct. 2020, 11, 3167–3179. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Tang, S.; Huang, Y.; Liang, F.; Fang, Y.; Pan, S.; Wu, T.; Xu, X. Lactobacillus casei-fermented blueberry pomace augments sIgA production in high-fat diet mice by improving intestinal microbiota. Food Funct. 2020, 11, 6552–6564. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, F.; Chai, Z.; Liu, M.; Battino, M.; Meng, X. Mixed fermentation of blueberry pomace with L. rhamnosus GG and L. plantarum-1: Enhance the active ingredient, antioxidant activity and health-promoting benefits. Food Chem. Toxicol. 2019, 131, 110541. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Pardo, F.A.; Nakajima, V.M.; Macedo, G.A.; Macedo, J.A.; Martínez, J. Extraction of phenolic compounds from dry and fermented orange pomace using supercritical CO2 and cosolvents. Food Bioprod. Process. 2017, 101, 1–10. [Google Scholar] [CrossRef]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic acids and potential for modifying the avian gastrointestinal tract and reducing pathogens and disease. Front. Vet. Sci. 2018, 5, 216. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.Z.; Mujumdar, A.S.; Pan, Z.; Vidyarthi, S.K.; Xu, J.; Zielinska, M.; Xiao, H.W. Emerging chemical and physical disinfection technologies of fruits and vegetables: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2020, 60, 2481–2508. [Google Scholar] [CrossRef] [PubMed]
- Halla, N.; Fernandes, I.P.; Heleno, S.A.; Costa, P.; Boucherit-Otmani, Z.; Boucherit, K.; Rodrigues, A.E.; Ferreira, I.C.F.R.; Barreiro, M.F. Cosmetics preservation: A review on present strategies. Molecules 2018, 23, 1571. [Google Scholar] [CrossRef] [Green Version]
- Golmohammadzadeh, R.; Faraji, F.; Rashchi, F. Recovery of lithium and cobalt from spent lithium ion batteries (LIBs) using organic acids as leaching reagents: A review. Resour. Conserv. Recycl. 2018, 136, 418–435. [Google Scholar] [CrossRef]
- Vashisht, A.; Thakur, K.; Kauldhar, B.S.; Kumar, V.; Yadav, S.K. Waste valorization: Identification of an ethanol tolerant bacterium Acetobacter pasteurianus SKYAA25 for acetic acid production from apple pomace. Sci. Total Environ. 2019, 690, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Parmar, I.; Rupasinghe, H.P.V. Bio-conversion of apple pomace into ethanol and acetic acid: Enzymatic hydrolysis and fermentation. Bioresour. Technol. 2013, 130, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Rudziak, A.; Kieliszek, M. Optimization of propionic acid production in apple pomace extract with Propionibacterium freudenreichii. Prep. Biochem. Biotechnol. 2019, 49, 974–986. [Google Scholar] [CrossRef] [PubMed]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Kot, A.M.; Kieliszek, M.; Bonin, S. Use of Propionibacterium freudenreichii T82 strain for effective biosynthesis of propionic acid and trehalose in a medium with apple pomace extract and potato wastewater. Molecules 2021, 26, 3965. [Google Scholar] [CrossRef]
- Ali, S.R.; Anwar, Z.; Irshad, M.; Mukhtar, S.; Warraich, N.T. Bio-synthesis of citric acid from single and co-culture-based fermentation technology using agro-wastes. J. Radiat. Res. Appl. Sci. 2016, 9, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Das, R.K.; Brar, S.K.; Verma, M. A fermentative approach towards optimizing directed biosynthesis of fumaric acid by Rhizopus oryzae 1526 utilizing apple industry waste biomass. Fungal Biol. 2015, 119, 1279–1290. [Google Scholar] [CrossRef]
- Das, R.K.; Lonappan, L.; Brar, S.K.; Verma, M. Bio-conversion of apple pomace into fumaric acid in a rotating drum type solid-state bench scale fermenter and study of the different underlying mechanisms. RSC Adv. 2015, 5, 104472–104479. [Google Scholar] [CrossRef] [Green Version]
- Madrera, R.R.; Bedriñana, R.P.; Valles, B.S. Production and characterization of aroma compounds from apple pomace by solid-state fermentation with selected yeasts. LWT Food Sci. Technol. 2015, 64, 1342–1353. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Guido, A.; Liberatore, C.M.; Ganino, T.; Lazzi, C.; Chiancone, B. From byproduct to resource: Fermented apple pomace as beer flavoring. Foods 2019, 8, 309. [Google Scholar] [CrossRef] [Green Version]
- Hadj Saadoun, J.; Ricci, A.; Cirlini, M.; Bancalari, E.; Bernini, V.; Galaverna, G.; Neviani, E.; Lazzi, C. Production and recovery of volatile compounds from fermented fruit by-products with Lacticaseibacillus rhamnosus. Food Bioprod. Process. 2021, 128, 215–226. [Google Scholar] [CrossRef]
- Antonić, B.; Jančíková, S.; Dordević, D.; Tremlová, B. Grape pomace valorization: A systematic review and meta-analysis. Foods 2020, 9, 1627. [Google Scholar] [CrossRef]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Yeni, F.; Yavaş, S.; Alpas, H.; Soyer, Y. Most Common Foodborne Pathogens and Mycotoxins on Fresh Produce: A Review of Recent Outbreaks. Crit. Rev. Food Sci. Nutr. 2016, 56, 1532–1544. [Google Scholar] [CrossRef]


{kind=link}
| Source | Experimental Conditions | Effect in Silage Characteristics | Ref. |
|---|---|---|---|
| Apple pomace | Alfalfa hay, timothy hay, soybean meal, and vitamin and mineral supplement, pomace (0, 5, 10, and 20%), 3.4–22.4 °C, and 60 days | Increased pH, ethanol, acetic acid, and ammonia nitrogen levels; reduced lactic acid content | [30] |
| Apple pomace | Maize, wheat bran, soybean meal, timothy hay, alfalfa hay and vitamin-mineral supplement, pomace (20% in silage), 4.4–25.8 °C, and 60 days | Increased pH and ethanol content; reduced lactic and acetic acid, and ammonia nitrogen contents | [31] |
| Grape pomace | Calotropis procera, pomace (0, 10, 20, 40% in silage), and 90 days | Increased ethanol, acetic, propionic and butyric acid contents, effluent loss and gas loss; reduced soluble carbohydrate and lactic acid content and digestibility; no effect in pH | [32] |
| Grape pomace | Sweet sorghum silage, pomace (0, 5, 10, and 15% in silage), and 90 days | Increased acetic acid (only 10%) and total polyphenol content; reduced water-soluble carbohydrates, lactic acid (only 15% treatment), and butyric acid contents; no effect in dry matter and protein neutral and detergent fiber contents, pH, and ammonia nitrogen level | [33] |
| White mulberry pomace | Meadow grass, pomace (0, 25, 50, 75, and 100% in silage), and 60 days | Increased gas production, organic matter digestibility, and metabolizable energy | [34] |
| Source | Experimental Conditions | Animals and Study Characteristics | Effect in Animal Production and Related Food | Ref. |
|---|---|---|---|---|
| Apple pomace | Silage: alfalfa hay, timothy hay, soybean meal, and vitamin and mineral supplement, pomace (0, 5, 10, and 20%), 3.4–22.4 °C, and 60 days | Suffolk wethers (4 animals), initial weight 50.3 kg, and 21 days of experiment | Reduced digestibility, gross energy, and nitrogen retention; no effect in feed intake and fiber content | [30] |
| Apple pomace | Silage: maize, wheat bran, soyabean meal, timothy hay, alfalfa hay and vitamin-mineral supplement, pomace (20% in silage), 4.4–25.8 °C, and 60 days | Suffolk wethers (4 animals), initial weight 65.3 kg, and 21 days of experiment | Increased organic acid content (except propionic and butyric); reduced digestibility and nitrogen retention; no effect in feed intake | [31] |
| Apple pomace | Silage: control feed, pomace (14.8% in silage), 9.7–20.1 °C, and 21 days | Male Yorkshire × Duroc × Landrace pigs (10 animals), initial weight 70 kg, and 53 days of experiment | Animals: increased feed efficiency; reduced average daily feed intake; no effect in finished body weight, average, daily gain, carcass weight, back fat thickness or dressing ratio Back fat: increased moisture, linoleic acid (C18:2n6), linolenic acid (C18:3) and arachidic acid; reduced water holding capacity, palmitic acid (C16:0), palmitoleic acid (C16:1) and heptadecenoic acid (C17:1) proportion | [35] |
| Apple pomace | Silage: minced sardine, pomace (15%), Lactobacillus plantarum (starter culture), 35 °C for 7 days | Juvenile European sea bass fish (240 animals), initial weight 15 g, and 9 weeks of experiment | Increased feed conversion ratio, relative average daily feed intake, leukocyte count, and carcass composition (moisture, lipid and ash contents); reduced final body weight, weight gain, specific growth rate, protein efficiency, apparent net protein utilization, and microvilli density | [36] |
| Grape pomace | Silage: sorghum, pomace (0, 10, 20, and 30%), and 7 months | Male mixed breed lambs (24 animals), initial weight 21.5 kg, and 35 days of experiment | No effect in performance, carcass composition, and meat quality | [37] |
| Grape pomace | Silage: corn, water, starter culture, and pomace (43.6 g/kg feed) | Landrace × Large White − Duroc − Pietrain piglets (24 animals), and 15 days of experiment | Animals: increased antioxidant defense system response, average daily gain, growth of facultative probiotic bacteria, and LAB; reduced oxidative stress and pathogen Meat: increased omega-3 fatty acids content; reduced n-6/n-3 ratio | [38] |
| Grape pomace | SSF: 1 kg substrate, Aspergillus niger, 30 °C, and 7 days; pomace (15 g/kg feed) | Male Ross 308 broiler chicks (140 animals), and 42 days of experiment | Animals: increased body weight and serum CAT level; reduced Clostridium perfringens count in cecum; no effect in feed intake, feed conversion ratio, serum GPx and SOD, other microorganism in cecum, and intestinal morphology Liver: no effect in pH and color | [39] |
| Olive pomace | SSF: Two-step fermentation: Bacillus subtilis var. natto N21, 37 °C, 2 days; Lactobacillus casei, 25–35 °C, and 5 days; pomace (7.5, 15, and 30%) | Male Ross 308 broiler chicks (1400 animals), initial weight 44–47 g, and 42 days of experiment | Animals: increased feed conversion ratio, antioxidant status and defense system response; reduced body weight gain, protein efficiency ratio, nutrient digestibility, serum triglycerides and total cholesterol; no effect in feed intake, serum LDL cholesterol, ALT and AST Breast meat: increased GPx and SOD; reduced fat and cholesterol content, and lipid oxidation status; no effect in moisture and protein | [40] |
| Tomato pomace | SSF: pomace (10% in silage), Lactobacillus plantarum (starter culture), and 30 days | Pregnant Holstein dairy cows (50 animals), initial weight 710–715 kg, 7 days of experiment | Animals: increased feed intake and digestibility, blood cholesterol and HDL, IgA, IgG, IgM, and antioxidant defense system response; no effect in feed intake, digestibility, milk yield and composition Milk: increased vitamin A, C, and E contents; no effect milk yield and composition | [41] |
| Pomegranate pomace | SSF: 100 g substrate, Aspergillus niger, 30 °C, and 7 days; pomace (5 and 10 g/kg feed) | Male Ross 308 broiler chicks (175 animals), initial weight 39 g, and 42 days of experiment | Animals: increased crypt depth; reduced lipid oxidation, Clostridium perfringens in cecum, and villus height; no effect in body weight, feed intake and conversion ratio, carcass characteristics, antioxidant defense system response, and muscularis mucosa thickness Meat and liver: no effect in color and pH | [42] |
| Source | Microorganism | Fermentation Conditions | Enzyme and Enzymatic Activity | Ref. |
|---|---|---|---|---|
| Apple pomace | Phanerochaete chrysosporium BKM-F-1767 | 40 g substrate, 60% moisture, pH 4.5, 37 °C, and 14 days | Lignin peroxidase: 141.4 U/gds Manganese peroxidase: 631.2 U/gds Laccase: 719.9 U/gds | [48] |
| Apple pomace | Rhizopus delemar F2 | 5 g apple pomace, 10 mL moistening agent (6.0 g Na2HPO4, 3.0 g KH2PO4, 0.5 g NaCl, 1.0 g NH2Cl, 2 mL of 1 M MgSO4 and 0.1 mL of 1 M CaCl2) | Amylase: 21.0 U/g Cellulase: 18.2 U/g Pectinase: 61.5 U/g Xylanase: 158.3 U/g | [44] |
| Apple pomace | Aspergillus parvisclerotigenus KX928754 | 5 g substrate, pH 7.0, 168 h, 30 °C, 2% sucrose and 3% peptone, 30 °C, and 168 h | Pectinase: 1366.3 U/mL | [45] |
| Apple pomace | Bacillus subtilis and Bacillus pumilus (20 and 80% in inoculum, respectively) | 15 g substrate/L, 0.2 g/L pectin, 0.2 g/L MgSO4 7H2O, and 0.2 g/L K2HPO4, pH 9.0, 130 rpm, 30 °C, and 24 h | Pectinase: 11.25 U/mL | [46] |
| Apple pomace and dahlia tubers | Mucor circinelloides | 10 g substrate, apple pomace: dahlia tubers (9:1), 83.5% moisture, 0.3% NH4H2PO4, 0.2% KH2PO4 and 0.1% KCl, pH 6.4, 30 °C, 5.8 days | Inulinase: 411.3 U/gds | [49] |
| Apple pomace | Cellulosimicrobium sp. CKMX1 (wild) and its mutant E5 | 10 g substrate, 20 mL basal salt medium, pH 8.0, 35 °C, and 72 h | Xylanase: 418 (wild) and 568 (mutant E5) U/g | [47] |
| Grape pomace | Aspergillus niger NRRL3 | 100 mL modified Czapex minimal medium with grape pomace, 4% tannic acid, pH 5.50, 120 rpm, 30 °C | Tannase: 3.0–4.5 U/mL | [50] |
| Grape pomace | Bacillus subtilis natto DSM 17766 | 15 g/100 mL, 3% H2SO4, pH 6.0, and 7 days | Cellulase: 0.2 U/mL | [51] |
| Grape pomace | Pleurotus ostreatus and Pleurotus pulmonarius | 4 g, 26 °C, 140 rpm, and 15 days | Laccase: 26.2 and 15,273.0 U/g for Pleurotus ostreatus and Pleurotus pulmonarius, respectively | [52] |
| Grape pomace and wheat bran | Aspergillus niger 3T5B8 | Grape pomace: wheat bran (50 and 50%), 60% moisture, 0.91% ammonium sulfate solution, 37 °C, and 96 h | Tannase: 0.30 U/g | [53] |
| White grape pomace, olive mill wastewater, red grape pomace and wheat bran | Aspergillus niger B60 | 50 g substrate, white grape pomace and olive mill wastewater, red grape pomace, and wheat bran (15, 15 and 70% of total substrate, respectively), 30 °C, and 120 h | CMCase: 668 U/g Polygalacturonase: 3151 U/g Amylase: 1099 U/g Xylanase: 579 U/g Protease: 204 U/g | [54] |
| Olive pomace | Kluyveromyces marxianus | 5 g substrate, 45 °C, and 48 h | Tannase: 42.4 U/mg | [55] |
| Exhausted olive pomace | Aspergillus niger CECT 2915 | 10 g substrate, 30 °C, and 6 days | Xylanase: 28 U/g Cellulase: 38 U/g | [56] |
| Olive pomace and wheat bran | Aspergillus ibericus MUM 03.49, Aspergillus niger MUM 03.58, and Aspergillus tubingensis MUM 06.152 | 30 g olive pomace: wheat bran (50 and 50%), 75% moisture, 30 °C, and 7 days | Lipase: 223, 53.6 and 7.6 U/g for A. ibericus, A. niger and A. tubingensis, respectively | [57] |
| Exhausted olive pomace | Crypthecodinium cohnii ATCC 30772 | 5 and 8 g substrate/L, 27 °C, 160 rpm, and 7 days | Pectinase: 37 and 33 U/mL for 8 and 5 g/L olive pomace, respectively | [58] |
| Tomato pomace | Aspergillus oryzae NRRL 2220 in SSF or SmF | 10 g substrate, 19.8 g/L casein, 0.92 g/L NaCl, 30 °C, and 72 h | Protease: 21,309 and 2343.5 U/g for SSF and SmF, respectively | [59] |
| Tomato pomace | Aspergillus oryzae 2220 | 20 g, 50% initial moisture content, pH 6 and 1 mL of 5-day-old inoculum, 30 °C, and 72 h | Protease: 12 U/gds after 42 h | [60] |
| Tomato pomace | Aspergillus oryzae 2220 (static bioreactor) | 5 kg, 10 cm bed, 30 °C, and 44 h | Protease: 13.6 U/gds | [60] |
| Tomato pomace and sorghum stalks | Pleurotus ostreatus and Trametes versicolor | 500 g tomato pomace, 100 g sorghum stalks, and 28 °C | Laccase: 15 and 35 U/g for P. ostreatus (4 days) and T. versicolor (18 days), respectively Protease: 13,000 and 34,000 U/g for P. ostreatus (4 days) and T. versicolor (13 days), respectively Xylanase: 9 and 50 U/g for P. ostreatus (4 days) and T. versicolor (13 days), respectively | [61] |
| Tomato pomace, wheat bran, and canola meal | Bacillus subtilis T4b | Wheat bran 30 g/L, canola meal 40 g/L, and tomato pomace 15 g/L, 180 rpm, 28 °C, and 48 h | Xylanase: 315 U/mL | [62] |
| Orange pomace | Aspergillus niger | 5 g substrate, 30 °C, and 96 h | Pectinase: around 17 U/g (endo+exo enzyme activities) | [63] |
| Orange pomace | Aspergillus niger (tray bioreactor) | 285 g substrate/tray, 30 °C, and 96 h | Pectinase: around 60 U/g (endo+exo enzyme activities) | [64] |
| Orange pomace with sugarcane bagasse | Aspergillus niger (tray bioreactor) | 285 g substrate/tray, 30 °C, and 96 h | Pectinase: around 75 U/g (endo+exo enzyme activities) | [64] |
| Orange pomace | Aspergillus niger (rotating-drum bioreactor) | 285 g substrate/batch, 30 °C, and 96 h | Pectinase: around 40 U/g (endo+exo enzyme activities) | [64] |
| Carrot pomace | Penicillium oxalicum BGPUP-4 | 10 g substrate, 90% moisture, 0.5% inulin, 0.2% NaNO3, 0.2 g/mL KH2PO4, 0.1% KCl, 0.05% MgSO4·7H2O, 0.001% FeSO4 7H2O and 0.2% NH4H2PO4, pH 7.0, 30 °C, and 4 days | Inulinase: 322.10 U/g | [65] |
| Source | Fermentation Conditions | Bioactive Compounds | Outcome | Ref. |
|---|---|---|---|---|
| Grape pomace | 2 g substrate, Rhizomucor miehei NRRL 5282, 37 °C, and 18 days | Polyphenols | Oven dried: reduction in TPC and FRAP, no effect in DPPH Lyophilized: maximum TPC and FRAP values at day 7, no effect in DPPH | [72] |
| Grape pomace | 10 g substrate (grape pomace:wheat bran; 1:1), Aspergillus niger 3T5B8, 37 °C, and 96 h | Polyphenols | Increased TPC, ABTS, and ORAC | [53] |
| Grape pomace | 50 g substrate, Trametes versicolor TV-6, 5 mycelial plugs, 27 °C, and 15 days | Polyphenols | Reduced 5-lipoxygenase and hyaluronidase activities (up to 4 days of fermentation), and polyphenol content throughout fermentation period | [74] |
| Grape pomace | 60 g substrate, Actinomucor elegans ATCC- 22963 or Umbelopsis isabellina ATCC-36671, 30 °C, and 12 days | γ-Linolenic acid and carotenoids | γ-Linolenic acid: maximum at 4 days for Umbelopsis isabellina and 6 days for Actinomucor elegans Carotenoids: carotene increased throughout fermentation and maximum at 8 days for lutein | [73] |
| Apple pomace | 2 g substrate, Rhizomucor miehei NRRL 5282, 37 °C, and 18 days | Polyphenols | Oven dried: reduced TPC, maximum FRAP value at day 3, no effect in DPPH Lyophilized: slight increase in TPC and DPPH up to day 10, maximum FRAP value at day 10 | [72] |
| Apple pomace | 12.5 g, natural fermentation, 30 °C, and 72 h | Polyphenols | Reduced throughout the fermentation period | [75] |
| Apple pomace | 250 g substrate, Saccharomyces cerevisiae ref: 32, Saccharomycodes bayanus ref: C6, and Hanseniaspora uvarum ref: 62, 25 °C, and 7 days | Fatty acids and polyphenols | Increased fatty acids Slight reduction in polyphenols | [76] |
| Apple pomace | 40, 60 and 80 g substrate/L, Yarrowia lipolytica, 28 °C, and 6 days | Fatty acids | Maximum production after day 3 | [77] |
| Elderberry and dwarf elderberry pomace | 50 g substrate, Aspergillus niger ATCC-6275, 30 °C, and 6 days | Polyphenols and fatty acids | TPC: maximum release up to 3–4 days of fermentation DPPH: maximum after 3–4 days of fermentation Lipids: slight increase in linoleic and oleic acids up to 4 days of fermentation | [78] |
| Olive pomace | 5 g substrate, Kluyveromyces marxianus NRRL Y-8281, 45 °C, and 48 h | Tannic and gallic acids | Reduced tannic acid and increased gallic acid content | [79] |
| Exhausted olive pomace | 5 and 8 g substrate/L, Crypthecodinium cohnii ATCC 30772, 27 °C, 160 rpm, and 7 days | Fatty acids | Increased total lipid and DHA content in dry cells | [58] |
| Exhausted olive pomace | 25 g substrate/L, Crypthecodinium cohnii ATCC 30772, 27 °C, 160 rpm, and 5 days | Fatty acids | High production yield; negative effect of detoxification prior to fermentation | [80] |
| Chokeberry pomace | 40 g substrate, Aspergillus niger ATCC-6275 or Rhizopus oligosporus ATCC-22959, 30 °C, and 12 days | Polyphenols | TPC: maximum at 6 days for or Rhizopus oligosporus and 9 days for Aspergillus niger; DPPH and TEAC: maximum at 6 days for Aspergillus niger and 9 days for Rhizopus oligosporus | [81] |
| Plum pomace | 15 g substrate, Aspergillus niger ATCC-6275 or Rhizopus oligosporus ATCC-22959, 30 °C, and 14 days | Polyphenols | TPC: maximum after 9 days of fermentation; DPPH: maximum at 6 days of fermentation | [82] |
| Apricot pomace | 15 g substrate, Aspergillus niger ATCC-6275 or Rhizopus oligosporus ATCC-22959, 30 °C, and 14 days | Polyphenols | TPC: maximum at 9 days for Rhizopus oligosporus; reduced after 6 days for Aspergillus niger; DPPH: maximum at 2 days for both | [83] |
| Pitahaya pomace | 2 g substrate, Rhizomucor miehei NRRL 5282, 37 °C, and 18 days | Polyphenols | Oven dried: slight decrease in TPC, decreased FRAP, and no effect in DPPH Lyophilized: slight increase in TPC, maximum FRAP value at day 10 and DPPH value at day 15 | [72] |
| Red bayberry pomace | 0.02% live yeast, 25 °C, 16 h followed by 0.1% probiotic mix, 28 °C, 24 h, and let for up to 7 days | Polyphenols | Increased TPC and TFC values; reduced DPPH value | [84] |
| Source | Fermentation Conditions | Organic Acid and Yield | Ref. |
|---|---|---|---|
| Apple pomace | 120 g substrate/L, Acetobacter pasteurianus, 37 °C, 180 rpm, and 24 h | Acetic acid: 52.4 g/100 g DM | [93] |
| Apple pomace | 1.5 L of substrate, Acetobacter aceti, pH 7.0, 28 °C, and 7 days | Acetic acid: 61.4 g/100 g DM | [94] |
| Apple pomace | 14 g substrate/100 g, Propionibacterium freudenreichii, 37 °C, and 120 h | Propionic acid: 38 g/100 g DM | [95] |
| Apple pomace | 250 mL substrate, Propionibacterium freudenreichii, 37 °C, and 120 h | Acetic acid: 5.01 g/L Propionic acid: 14.54 g/L | [96] |
| Apple pomace | 25 g substrate, Aspergillus ornatus and Alternaria alternate, pH 5.0, 30 °C, and 48 h | Citric acid: 0.5 g/L | [97] |
| Apple pomace | 25 g substrate, Rhizopus oryzae, 30 °C, and 14 days | Fumaric acid: 52 g/kg | [98] |
| Apple pomace | 3–4 L working volume, 50% moisture, Rhizopus oryzae, 1.97 atm, and 14 days | Fumaric acid: 138 g/kg | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munekata, P.E.S.; Domínguez, R.; Pateiro, M.; Nawaz, A.; Hano, C.; Walayat, N.; Lorenzo, J.M. Strategies to Increase the Value of Pomaces with Fermentation. Fermentation 2021, 7, 299. https://doi.org/10.3390/fermentation7040299
Munekata PES, Domínguez R, Pateiro M, Nawaz A, Hano C, Walayat N, Lorenzo JM. Strategies to Increase the Value of Pomaces with Fermentation. Fermentation. 2021; 7(4):299. https://doi.org/10.3390/fermentation7040299
Chicago/Turabian StyleMunekata, Paulo E. S., Rubén Domínguez, Mirian Pateiro, Asad Nawaz, Christophe Hano, Noman Walayat, and José M. Lorenzo. 2021. "Strategies to Increase the Value of Pomaces with Fermentation" Fermentation 7, no. 4: 299. https://doi.org/10.3390/fermentation7040299
APA StyleMunekata, P. E. S., Domínguez, R., Pateiro, M., Nawaz, A., Hano, C., Walayat, N., & Lorenzo, J. M. (2021). Strategies to Increase the Value of Pomaces with Fermentation. Fermentation, 7(4), 299. https://doi.org/10.3390/fermentation7040299