
Single-Particle Characterization of SARS-CoV-2 Isoelectric Point and Comparison to Variants of Interest


Abstract
:

Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aboubakr, A.H.; Sharafeldin, T.A.; Goyal, S.M. Stability of SARS-CoV-2 and other coronaviruses in the environment and on common touch surfaces and the influence of climatic conditions: A review. Transbound. Emerg. Dis. 2021, 68, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Joonaki, E.; Hassanpouryouzband, A.; Heldt, C.L.; Areo, O. Surface Chemistry Can Unlock Drivers of Surface Stability of SARS-CoV-2 in a Variety of Environmental Conditions. Chem 2020, 6, 2135–2146. [Google Scholar] [CrossRef]
- Vega, E.; Garland, J.; Pillai, S.D. Electrostatic Forces Control Nonspecific Virus Attachment to Lettuce. J. Food Prot. 2008, 71, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Van Oss, C. Acid—base interfacial interactions in aqueous media. Colloids Surfaces A Physicochem. Eng. Asp. 1993, 78, 1–49. [Google Scholar] [CrossRef]
- Dang, H.T.; Tarabara, V.V. Virus deposition onto polyelectrolyte-coated surfaces: A study with bacteriophage MS2. J. Colloid Interface Sci. 2019, 540, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Chrysikopoulos, C.V.; Syngouna, V.I. Attachment of bacteriophages MS2 and ΦX174 onto kaolinite and montmorillonite: Extended-DLVO interactions. Colloids Surfaces B Biointerfaces 2012, 92, 74–83. [Google Scholar] [CrossRef]
- Boone, S.A.; Gerba, C.P. Significance of Fomites in the Spread of Respiratory and Enteric Viral Disease. Appl. Environ. Microbiol. 2007, 73, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Zhu, X.; Jańczewski, D.; Lee, S.S.C.; He, T.; Teo, S.L.M.; Vancso, G.J. Measuring protein isoelectric points by AFM-based force spectroscopy using trace amounts of sample. Nat. Nanotechnol. 2016, 11, 817–823. [Google Scholar] [CrossRef]
- Michen, B.; Graule, T. Isoelectric points of viruses. J. Appl. Microbiol. 2009, 109, 388–397. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S. DLS and zeta potential—What they are and what they are not? J. Control. Release 2016, 235, 337–351. [Google Scholar] [CrossRef]
- Samandoulgou, I.; Fliss, I.; Jean, J. Zeta Potential and Aggregation of Virus-Like Particle of Human Norovirus and Feline Calicivirus under Different Physicochemical Conditions. Food Environ. Virol. 2015, 7, 249–260. [Google Scholar] [CrossRef]
- Dika, C.; Duval, J.F.; Francius, G.; Perrin, A.; Gantzer, C. Isoelectric point is an inadequate descriptor of MS2, Phi X 174 and PRD1 phages adhesion on abiotic surfaces. J. Colloid Interface Sci. 2015, 446, 327–334. [Google Scholar] [CrossRef]
- Heffron, J.; Mayer, B.K. Improved Virus Isoelectric Point Estimation by Exclusion of Known and Predicted Genome-Binding Regions. Appl. Environ. Microbiol. 2020, 86, e01674-20. [Google Scholar] [CrossRef]
- Krebs, F.; Scheller, C.; Grove-Heike, K.; Pohl, L.; Watzig, H. Isoelectric point determination by imaged CIEF of commercially available SARS-CoV-2 proteins and the hACE2 receptor. Electrophoresis 2021, 42, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Hristova, S.H.; Zhivkov, A.M. Isoelectric point of free and adsorbed cytochrome c determined by various methods. Colloids Surfaces B Biointerfaces 2019, 174, 87–94. [Google Scholar] [CrossRef]
- Mi, X.; Bromley, E.K.; Joshi, P.U.; Long, F.; Heldt, C.L. Virus Isoelectric Point Determination Using Single-Particle Chemical Force Microscopy. Langmuir 2019, 36, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Mi, X.; Heldt, C.L. Single-particle chemical force microscopy to characterize virus surface chemistry. BioTechniques 2020, 69, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Weichert, W.S.; Parker, J.; Wahid, A.; Chang, S.-F.; Meier, E.; Parrish, C.R. Assaying for Structural Variation in the Parvovirus Capsid and Its Role in Infection. Virology 1998, 250, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Swiss Institute of Bioinformations (SIB). ExPASy: ProtParam Tool. Available online: https://web.expasy.org/protparam/ (accessed on 15 May 2021).
- Pandey, L.M. Surface engineering of personal protective equipments (PPEs) to prevent the contagious infections of SARS-CoV-2. Surf. Eng. 2020, 36, 901–907. [Google Scholar] [CrossRef]
- Scheller, C.; Krebs, F.; Minkner, R.; Astner, I.; Gil-Moles, M.; Watzig, H. Physicochemical properties of SARS-CoV-2 for drug targeting, virus inactivation and attenuation, vaccine formulation and quality control. Electrophoresis 2020, 41, 1137–1151. [Google Scholar] [CrossRef] [PubMed]
- Gobeil, S.M.; Janowska, K.; McDowell, S.; Mansouri, K.; Parks, R.; Stalls, V.; Kopp, M.F.; Manne, K.; Saunders, K.; Edwards, R.J.; et al. Effect of natural mutations of SARS-CoV-2 on spike structure, conformation and antigenicity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Heldt, C.L.; Zahid, A.; Vijayaragavan, K.S.; Mi, X. Experimental and computational surface hydrophobicity analysis of a non-enveloped virus and proteins. Colloids Surf. B Biointerfaces 2017, 153, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292 e6. [Google Scholar] [PubMed]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
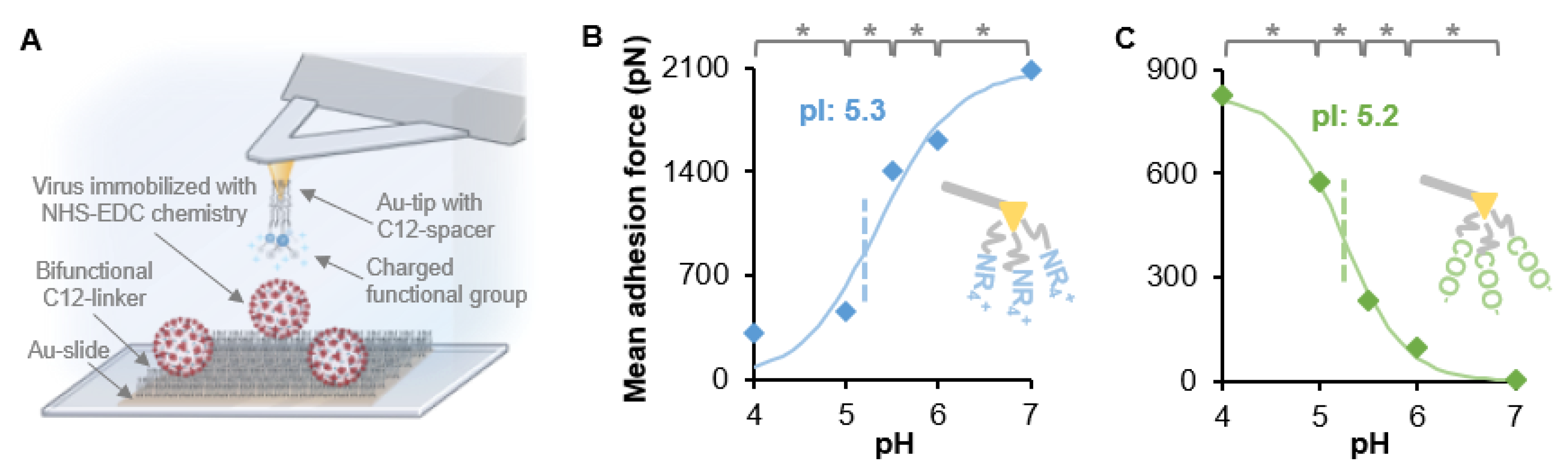{kind=link}
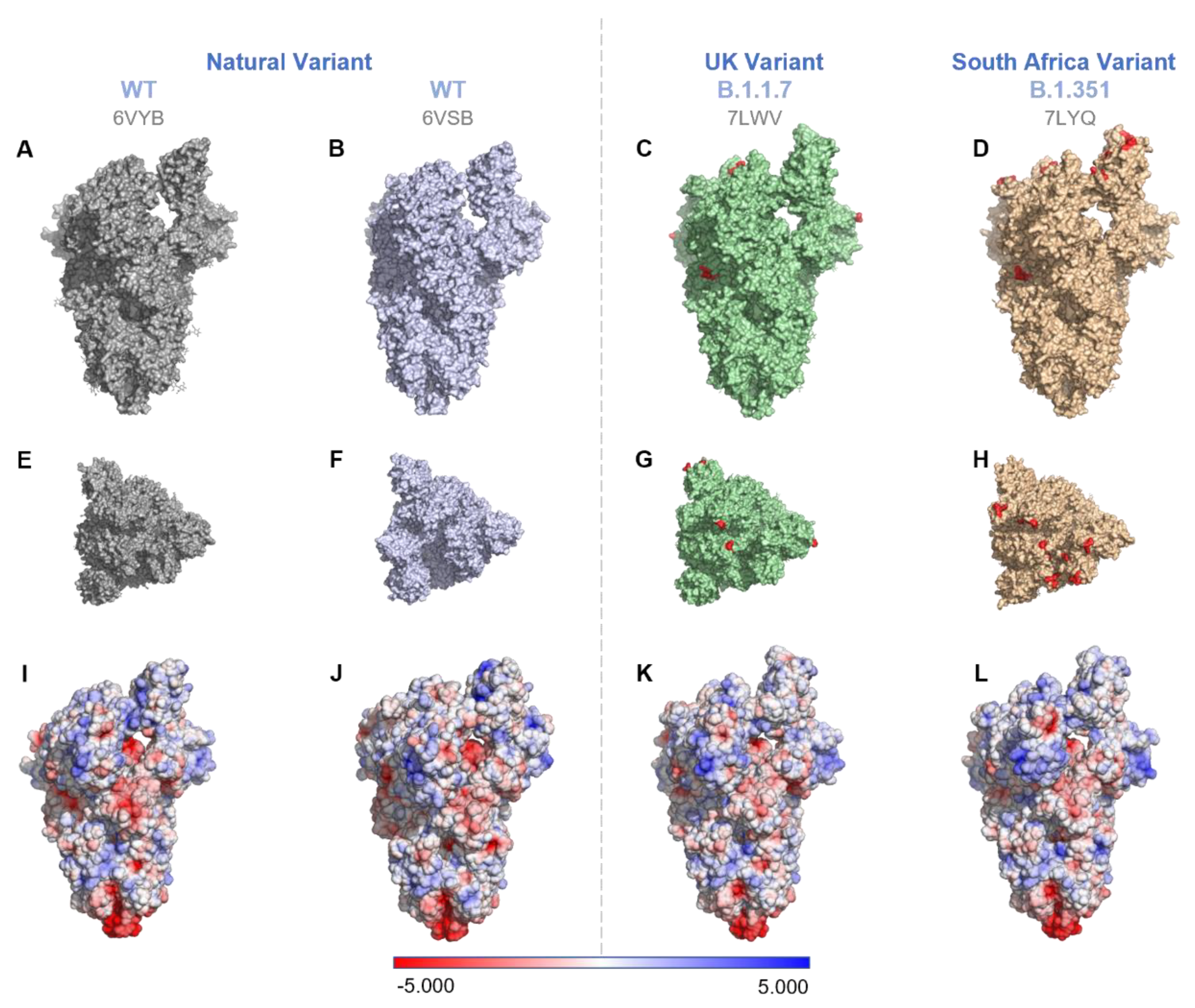{kind=link}
| Calculation/Method | Protein | IEP | Reference |
|---|---|---|---|
| Amino acid sequence | SARS-CoV-2 spike glycoprotein | 5.9 | [20] |
| ProtParam | SARS-CoV-2 spike glycoprotein | 6.24 | [21] |
| ProtParam | His-tagged SARS-CoV-2 RBD 1 | 8.91 | [14] |
| CIEF | His-tagged SARS-CoV-2 RBD 1 | 7.36–9.85 | [14] |
| CIEF | S1/S2 subunit with His-tag | 4.41–5.87 | [14] |
| Pango Lineage | Name | Mutations | PDB | Surface Charge (Formal Charge) | Surface Charge (Partial Charge) | RBD Charge |
|---|---|---|---|---|---|---|
| WT (Natural variant) | USA/WA1/2020 | - | 6VYB | −20 | −17 | 9 |
| - | 6VSB | −26 | −23 | 6 | ||
| B.1.1.7 (UK variant) | 20J/501Y.V1 | Δ69/70 Δ144Y N501Y A570D D614G P681H | 7LWV | −12 | −9 | 7 |
| B.1.351 (South Africa variant) | 20H/501.V2 | K417N E484K N501Y D614G | 7LYQ | −7 | −4 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Areo, O.; Joshi, P.U.; Obrenovich, M.; Tayahi, M.; Heldt, C.L. Single-Particle Characterization of SARS-CoV-2 Isoelectric Point and Comparison to Variants of Interest. Microorganisms 2021, 9, 1606. https://doi.org/10.3390/microorganisms9081606
Areo O, Joshi PU, Obrenovich M, Tayahi M, Heldt CL. Single-Particle Characterization of SARS-CoV-2 Isoelectric Point and Comparison to Variants of Interest. Microorganisms. 2021; 9(8):1606. https://doi.org/10.3390/microorganisms9081606
Chicago/Turabian StyleAreo, Oluwatoyin, Pratik U. Joshi, Mark Obrenovich, Moncef Tayahi, and Caryn L. Heldt. 2021. "Single-Particle Characterization of SARS-CoV-2 Isoelectric Point and Comparison to Variants of Interest" Microorganisms 9, no. 8: 1606. https://doi.org/10.3390/microorganisms9081606
APA StyleAreo, O., Joshi, P. U., Obrenovich, M., Tayahi, M., & Heldt, C. L. (2021). Single-Particle Characterization of SARS-CoV-2 Isoelectric Point and Comparison to Variants of Interest. Microorganisms, 9(8), 1606. https://doi.org/10.3390/microorganisms9081606