Antifungal Activity of Thapsia villosa Essential Oil against Candida, Cryptococcus, Malassezia, Aspergillus and Dermatophyte Species




Abstract
:1. Introduction
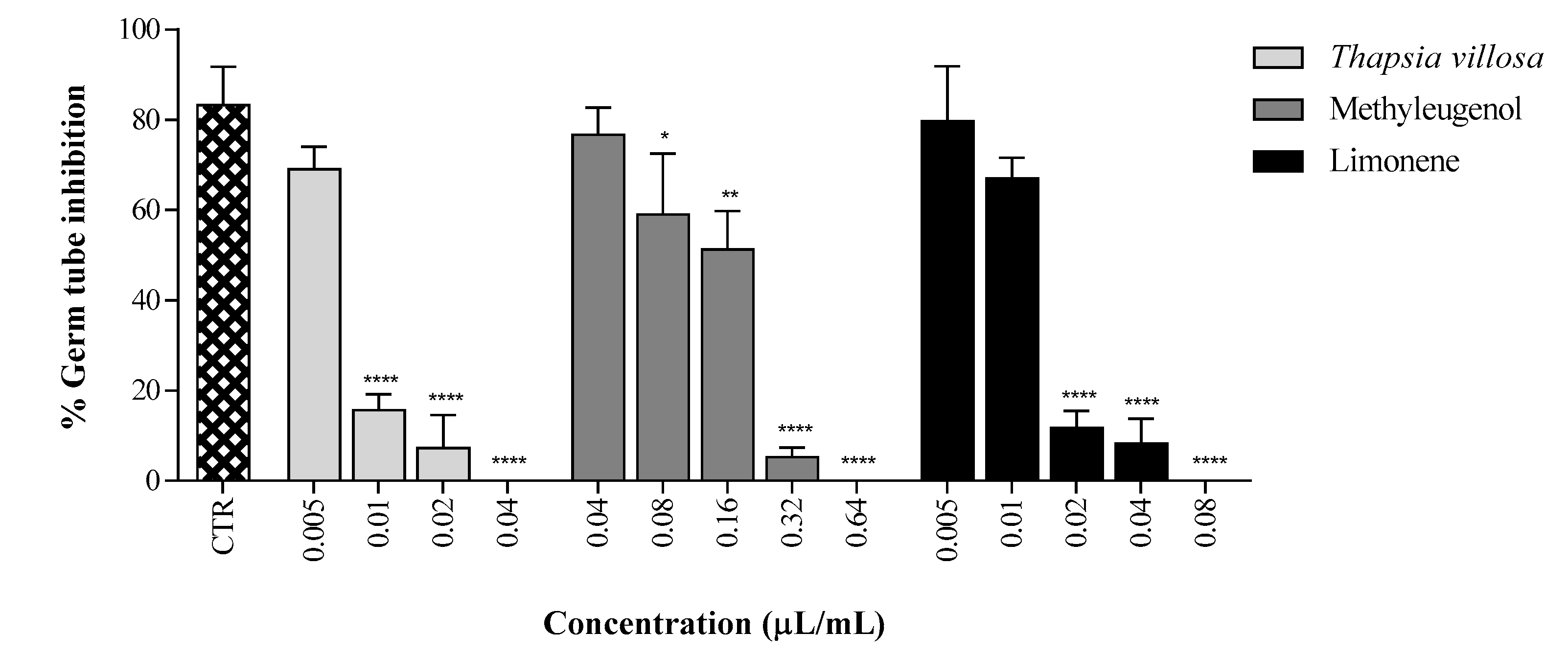
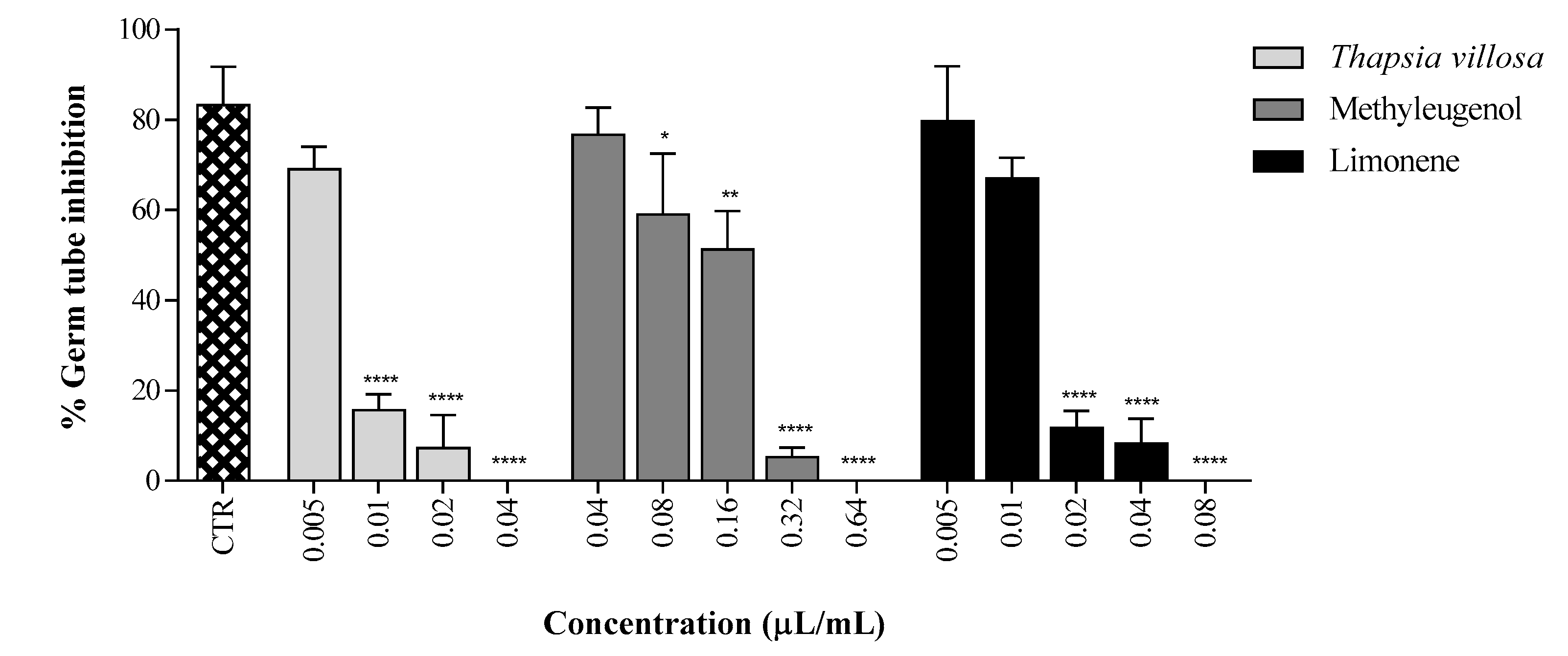
2. Results and Discussion
3. Material and Methods
3.1. Fungal Organisms
3.2. Isolation of Essential Oil from Plant Material and Reference Compounds
3.3. Chemical Composition
3.3.1. Gas Chromatography (GC)
3.3.2. Gas Chromatography-Mass Spectrometry (GC-MS)
3.3.3. Identification of Individual Components
3.4. Antifungal Activity
3.5. Interaction Test by Checkboard Microdilution Assay
3.6. Germ Tube Inhibition Assay
3.7. Haemolytic Activity
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Franz, C.; Novak, J. Sources of essential oils. In Handbook of Essential Oils: Science, Technology, and Applications; Baser, C.K.H., Buchbauer, G., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2010. [Google Scholar]
- Buchbauer, G. The detailed analysis of essential oil leads to the understanding of their properties. Perfum. Flavorist 2000, 25, 64–67. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.; Lass-Flörl, C. Changing epidemiology of systemic fungal infections. Clin. Microbiol. Infect. 2008, 14, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.D.; Warnock, D.W. Fungal Infection: Diagnosis and Management, 4th ed.; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Denning, D.W.; Hope, W.W. Therapy for fungal diseases: Opportunities and priorities. Trends Microbiol. 2010, 18, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A. Antifungal drug resistance: Mechanism, epidemiology and consequences for treatment. Am. J. Med. 2012, 125, 3–137. [Google Scholar] [CrossRef] [PubMed]
- Oroojalian, F.; Kasra-Kermanshahi, R.; Azizi, M.; Bassami, M.R. Phytochemical composition of the essential oils from three Apiaceae species and their antibacterial effects on food-borne pathogens. Food Chem. 2010, 120, 765–770. [Google Scholar] [CrossRef]
- Popović, V.; Petrović, S.; Pavlović, M.; Milenković, M.; Couladis, M.; Tzakou, O.; Duraki, S.; Niketić, M. Essential oil from the underground parts of Laserpitium zernyi: Potential source of alfa-bisabolol and its antimicrobial activity. Nat. Prod. Commun. 2010, 5, 307–310. [Google Scholar] [PubMed]
- Tavares, A.C.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Lopes, C.; Canhoto, J.; Salgueiro, L.R. Essential oils of Distichoselinum tenuifolium: Chemical composition, cytotoxicity, antifungal and anti-inflammatory properties. J. Etnopharmacol. 2010, 130, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Siljegović, J.; Glamoclija, J.; Sokovic, M.; Vuckovic, I.; Tesevic, V.; Milosavljević, S.; Stesević, D. Composition and antimicrobial activity of Seseli montanum subsp. tommasinii essential oil. Nat. Prod. Commun. 2011, 6, 263–266. [Google Scholar] [PubMed]
- Valente, J.; Zuzarte, M.; Resende, R.; Gonçalves, M.J.; Cavaleiro, C.; Pereira, C.F.; Cruz, M.T.; Salgueiro, L. Daucus carota subsp. gummifer essential oil as a natural source of antifungal and anti-inflammatory drugs. Ind. Crops Prod. 2015, 65, 361–366. [Google Scholar] [CrossRef]
- Abderrahim, O.; Martin, G.J.; Abdelazzi, A. Botanical identification and ethnomedicinal uses of some underground parts of medicinal plants collected and traded in Marrakech region. J. Med. Plants Res. 2013, 7, 2165–2169. [Google Scholar] [CrossRef]
- Casiglia, S.; Riccobono, L.; Maurizio, B.; Rosselli, S.; Senatore, F.; Senatore, F. Chemical composition of the essential oil from Thapsia garganica (Apiaceae) grown wild in Sicily and its antimicrobial activity. Nat. Prod. Res. 2016, 30, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, C.M.; Denmeade, S.R.; Isaacs, J.T.; Gady, A.; Olsen, C.E.; Christensen, S.B. Design, synthesis and pharmacological evaluation of thapsigargin analogues for targeting apoptosis to prostatic cancer cells. J. Med. Chem. 2001, 44, 4696–4703. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.J.; Cruz, M.T.; Tavares, A.C.; Cavaleiro, C.; Lopes, M.C.; Canhoto, J.; Salgueiro, L. Composition and biological activity of the essential oil from Thapsia minor, a new source of geranyl acetate. Ind. Crops Prod. 2012, 35, 166–171. [Google Scholar] [CrossRef]
- Castroviejo, S.; Laínz, M.; López González, G.; Montserrat, P.; Muñoz Garmendia, F.; Paiva, J.; Villar, L. Araliaceae-Umbelliferae. In Flora Ibérica, Plantas Vasculares de la Península Ibérica, e Islas Baleares; Real Jardín Botánico, CSIC: Madrid, Spain, 2003; Volume X. [Google Scholar]
- Avato, P.; Jacobsen, N.; Smitht, U. Composition of the essential oils of fruits from polyploid types of Thapsia villosa: Chemotaxonomic evaluation. J. Essent. Oil Res. 1996, 8, 123–128. [Google Scholar] [CrossRef]
- Avato, P.; Trabace, G.; Smitht, U. Essential oils from fruits of three types of Thapsia villosa. Phytochemistry 1996, 43, 609–612. [Google Scholar] [CrossRef]
- Avato, P.; Cornetti, C.; Andersen, A.; Smitt, U.W.; Christensen, S.B. Localization of the acyl groups in proazulene guaianolides from Thapsia transtagana and Thapsia garganica. J. Nat. Prod. 1993, 56, 411–415. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, A.; Manzoor, N.; Khan, L.A. Evolution of ergosterol biosynthesis inhibitors as fungicidal against Candida. Microb. Pathog. 2010, 48, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, A.; Khan, L.A.; Manzoor, N. Synergy of eugenol and methyleugenol with fluconazole against clinical Candida isolates. J. Med. Microbiol. 2010, 59, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Nishida, R. Methyl eugenol: Its occurrence, distribution, and role in nature, especially in relation to insect behavior and pollination. J. Insect. Sci. 2012, 12, 1–74. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.K.; Silva, J.R.; Nascimento, S.B.; da Luz, S.F.; Meireles, E.N.; Alves, C.N.; Ramos, A.R.; Maia, J.G. Antifungal activity and computational study of constituents from Piper divaricatum essential oil against Fusarium infection in black pepper. Molecules 2014, 19, 17926–17942. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.K. Chemical composition, in vitro antimicrobial and antioxidant activities of the essential oils of Ocimum Gratissimum, O. Sanctum and their major constituents. Indian J. Pharm. Sci. 2013, 8, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and dermatophyte species. J. Med. Microbiol. 2009, 58, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Chee, H.Y.; Kim, H.; Lee, M.H. In vitro antifungal activity of limonene against Trichophyton rubrum. Mycobiology 2009, 37, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Vimal, M.; Vijaya, P.P.; Mumtaj, P.; Seema Farhath, M.S. Antibacterial activity of selected compounds of essential oils indigenous plants. J. Chem. Pharm. Res. 2013, 5, 248–253. [Google Scholar]
- Espina, L.; Gelaw, T.K.; Lamo-Castellví, S.; Pagán, R.; García-Gonzalo, D. Mechanism of Bacterial inactivation by (+)-limonene and its potential use in food preservation combined processes. PLoS ONE 2013, 8, e56769. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. D-Limonene: Safety and clinical applications. Altern. Med. Rev. 2007, 12, 259–264. [Google Scholar] [PubMed]
- Koroch, A.R.; Juliani, H.R.; Zygadlo, A.J. Bioactivity of essential oils and their components. In Flavour and Fragrances—Chemistry, Bioprocessing and Sustainability; Berger, R.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Saville, S.P.; Lazzell, A.L.; Bryant, A.P.; Fretzen, A.; Monreal, A.; Solberg, E.O.; Monteagudo, C.; Lopez-Ribot, J.L.; Milne, G.T. Inhibition of filamentation can be used to treat disseminated candidiasis. Antimicrob. Agents Chemother. 2006, 50, 3312–3316. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Vale-Silva, L.; Gonçalves, M.J.; Cavaleiro, C.; Vaz, S.; Canhoto, J.; Pinto, E.; Salgueiro, L. Antifungal activity of phenolic-rich Lavandula multifida L. essential oil. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Cavaleiro, C.; Salgueiro, L.; Gonçalves, M.J.; Hrimpeng, K.; Pinto, J.; Pinto, E. Antifungal activity of the essential oil of Angelica major against Candida, Cryptococcus, Aspergillus and dermatophyte species. J. Nat. Med. 2015, 69, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Pinto, E.; Salgueiro, L. Natural products: An alternative to conventional therapy for dermatophytosis? Mycopathologia 2017, 182, 143–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, J-H.; Lee, J-S. Chemical composition and antifungal activity of plant essential oils against Malassezia furfur. Korean J. Microbiol. Biotechnol. 2010, 38, 315–321. [Google Scholar]
- Council of Europe. European Pharmacopoeia, 3rd ed.; Council of Europe: Strasbourg, France, 1997. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatogra-phy/Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2004. [Google Scholar]
- Wiley. Wiley Registry of Mass Spectral Data Information; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- Joulain, D.; Konig, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; E.B. Verlag: Hamburg, Germany, 1998. [Google Scholar]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard-Third Edition M27-A3 and Third Informational Supplement M27-S3; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Conidium-Forming Filamentous Fungi; Approved Standard-Second edition M38-A2; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Mukherjee, P.K.; Sheehan, D.J.; Hitchcock, C.A.; Ghannoum, M.A. Combination treatment of invasive fungal infections. Clin. Microbiol. Rev. 2005, 18, 163–194. [Google Scholar] [CrossRef] [PubMed]
- Marichal, P.; Gorrens, J.; Vancutsem, J.; Vandenbossche, H. Culture Media for the study of the effects of azole derivatives on germ tube formation and hyphal growth of Candida albicans. Mykosen 1986, 29, 76–81. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |

{kind=link}
| Compound * | RI | RI | % | % |
|---|---|---|---|---|
| SPB-1 a | SW 10 b | A | B | |
| α-Thujene | 922 | 1029 | 0.1 | t |
| α-Pinene | 930 | 1030 | 0.1 | t |
| Sabinene | 964 | 1128 | 0.1 | t |
| β-Pinene | 970 | 1118 | 0.1 | t |
| Myrcene | 980 | 1161 | 0.2 | 0.3 |
| p-Cymene | 1012 | 1272 | 0.3 | 0.3 |
| Limonene | 1020 | 1206 | 56.0 | 57.5 |
| γ-Terpinene | 1046 | 1249 | 0.5 | 0.3 |
| Nonanal | 1084 | 1393 | 0.1 | t |
| Linalool | 1084 | 1541 | 0.1 | 0.1 |
| Terpinen-4-ol | 1158 | 1597 | t | t |
| α-Terpineol | 1169 | 1692 | t | t |
| cis-Carveol | 1206 | 1860 | 0.1 | 0.1 |
| Carvone | 1212 | 1728 | 0.1 | 0.1 |
| E-p-Menth-1(7),8-dien-2-ol | 1167 | 1790 | t | t |
| Z-p-Menth-1(7),8-dien-2-ol | 1240 | 1555 | t | 0.1 |
| Methyleugenol | 1368 | 2012 | 35.0 | 35.9 |
| E-Caryophyllene | 1408 | 1590 | 0.1 | t |
| Germacrene-d | 1466 | 1699 | 0.1 | 0.1 |
| Myristicine | 1485 | 2253 | 0.1 | t |
| Elemicine | 1516 | 2253 | 1.2 | 1.0 |
| Monoterpene hydrocarbons | 57.4 | 58.6 | ||
| Oxygen containing monoterpenes | 0.5 | 0.6 | ||
| Sesquiterpene hydrocarbons | 0.2 | 0.1 | ||
| Phenylpropanoids | 36.3 | 37.0 | ||
| Other compounds | 0.1 | t | ||
| Total identified | 94.5 | 96.3 |
| Essential Oil | Methyleugenol | (R)-(+)-Limonene | Fluconazole | Amphotericin B | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC a | MFC a | MIC a | MFC a | MIC a | MFC a | MIC b | MFC b | MIC b | MFC b | |||
| Yeasts | ATCC | Candida albicans ATCC 10231 | 1.25 | 1.25–2.5 | 0.64 | 0.64 | 0.32 | 0.32 | 1 | >128 | - | - |
| C. krusei ATCC 6258 | 0.64 | 1.25 | 0.64 | 0.64 | 0.16 | 0.16 | 64 | 64–128 | - | - | ||
| C. tropicalis ATCC 13803 | 0.64–1.25 | 1.25 | 0.64 | 0.64 | 0.64 | 0.64 | 4 | >128 | - | - | ||
| C. parapsilosis ATCC 90018 | 1.25 | 1.25–2.5 | 0.32–0.64 | 0.64 | 0.64 | 0.64 | 1 | 1–2 | - | - | ||
| Cryptococcus neoformans CECT 1078 | 0.16 | 0.16 | 0.32 | 0.32–0.64 | 0.08 | 0.08 | 16 | 128 | - | - | ||
| Clinical isolates | C. albicans D5 | 1.25 | 1.25 | 0.64 | 0.64 | 0.16 | 0.16 | 64 | >128 | - | - | |
| C. albicans M1 | 1.25 | 2.5 | 0.64 | 0.64 | 0.64 | 0.64 | 2 | 128 | - | - | ||
| C. dubliniensis CD1 | 1.25 | 1.25 | 0.64 | 0.64 | 0.16 | 0.16 | 1 | >128 | - | - | ||
| C. glabrata D10R | 1.25 | 2.5 | 0.64 | 0.64 | 0.32 | 0.32–0.64 | 32 | 32 | - | - | ||
| Malassezia furfur P26 | 2.5 | 2.5 | - | - | - | - | - | - | - | - | ||
| Filamentous fungi | Dermatophytes | Epidermophyton floccosum FF9 | 0.64 | 0.64 | 0.32 | 0.32 | 0.08 | 0.08 | 16 | 16 | - | - |
| Trichophyton rubrum CECT 2794 | 0.64 | 0.64 | 0.32 | 0.64 | 0.08 | 0.08 | 16 | 64 | - | - | ||
| T. mentagrophytes FF7 | 0.64 | 1.25 | 0.32 | 0.32–0.64 | 0.16 | 0.16 | 16–32 | 32–64 | - | - | ||
| T. mentagrophytes var. interdigitale CECT 2958 | 1.25 | 1.25 | 0.32 | 0.64 | 0.16 | 0.16 | 128 | ≥128 | - | - | ||
| T. verrucosum CECT 2992 | 1.25 | 1.25 | 0.32 | 0.32 | 0.16 | 0.16 | >128 | >128 | - | - | ||
| Microsporum canis FF1 | 0.64 | 0.64 | 0.32 | 0.32 | 0.08 | 0.16 | 128 | 128 | - | - | ||
| M. gypseum CECT 2908 | 1.25 | 1.25 | 0.32 | 0.32–0.64 | 0.08–0.16 | 0.08–0.16 | 128 | >128 | - | - | ||
| Aspergillus species | Aspergillus flavus F44 | 1.25 | >5 | 0.64 | >2.5 | 0.32–0.64 | 0.64 | - | - | 2 | 8 | |
| A. fumigatus ATCC 46645 | 1.25 | 1.25–2.5 | 0.32 | 1.25 | 0.32 | 0.32 | - | - | 2 | 4 | ||
| A. niger ATCC 16404 | 0.64 | ≥5 | 0.64 | >2.5 | 0.32 | 0.64 | - | - | 1–2 | 4 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, E.; Gonçalves, M.-J.; Cavaleiro, C.; Salgueiro, L. Antifungal Activity of Thapsia villosa Essential Oil against Candida, Cryptococcus, Malassezia, Aspergillus and Dermatophyte Species. Molecules 2017, 22, 1595. https://doi.org/10.3390/molecules22101595
Pinto E, Gonçalves M-J, Cavaleiro C, Salgueiro L. Antifungal Activity of Thapsia villosa Essential Oil against Candida, Cryptococcus, Malassezia, Aspergillus and Dermatophyte Species. Molecules. 2017; 22(10):1595. https://doi.org/10.3390/molecules22101595
Chicago/Turabian StylePinto, Eugénia, Maria-José Gonçalves, Carlos Cavaleiro, and Lígia Salgueiro. 2017. "Antifungal Activity of Thapsia villosa Essential Oil against Candida, Cryptococcus, Malassezia, Aspergillus and Dermatophyte Species" Molecules 22, no. 10: 1595. https://doi.org/10.3390/molecules22101595
APA StylePinto, E., Gonçalves, M.-J., Cavaleiro, C., & Salgueiro, L. (2017). Antifungal Activity of Thapsia villosa Essential Oil against Candida, Cryptococcus, Malassezia, Aspergillus and Dermatophyte Species. Molecules, 22(10), 1595. https://doi.org/10.3390/molecules22101595