Preparation and Characterization of Cellulose Triacetate as Support for Lecitase Ultra Immobilization

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
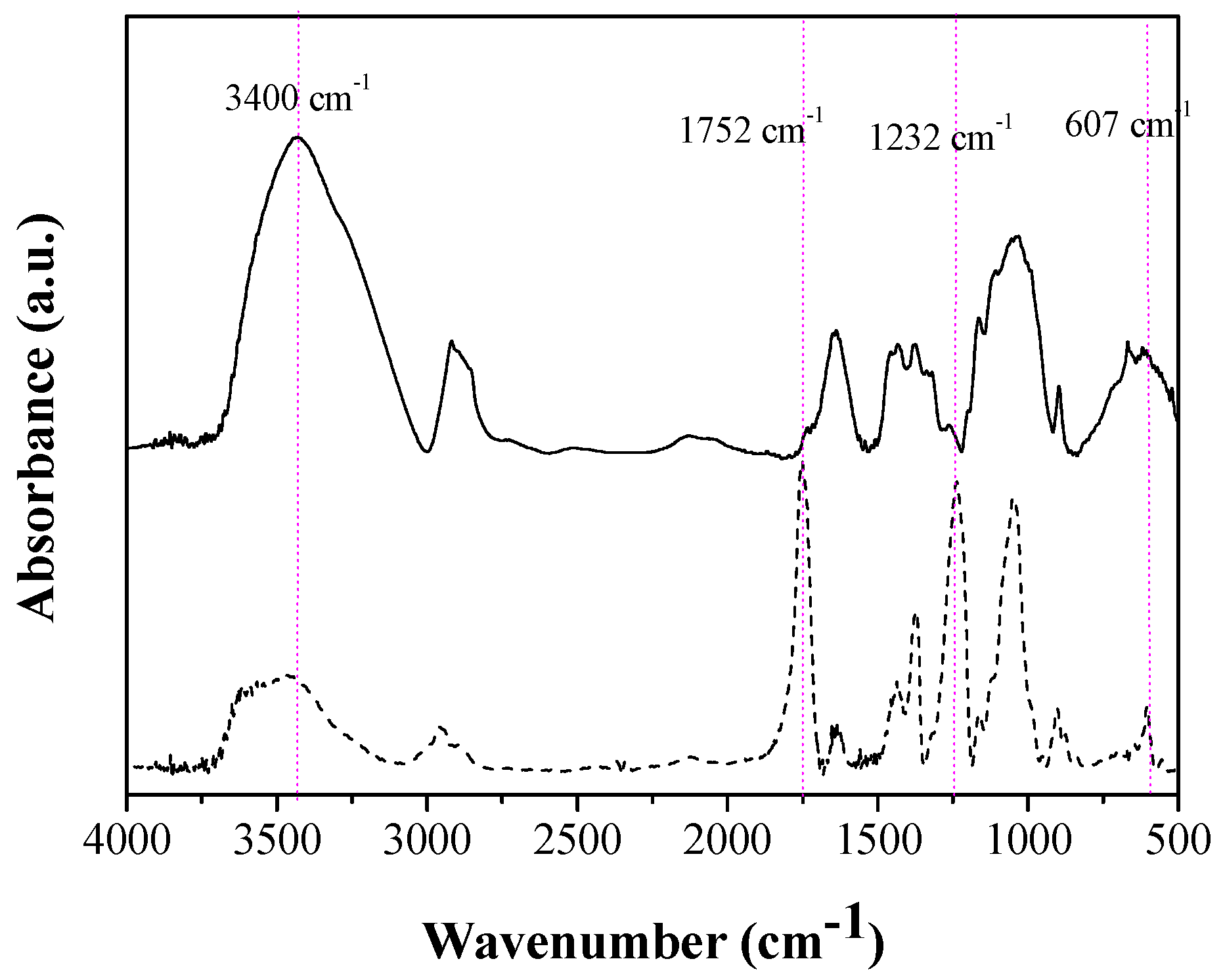
2.2. Production of CA and Characterization of the Material Obtained
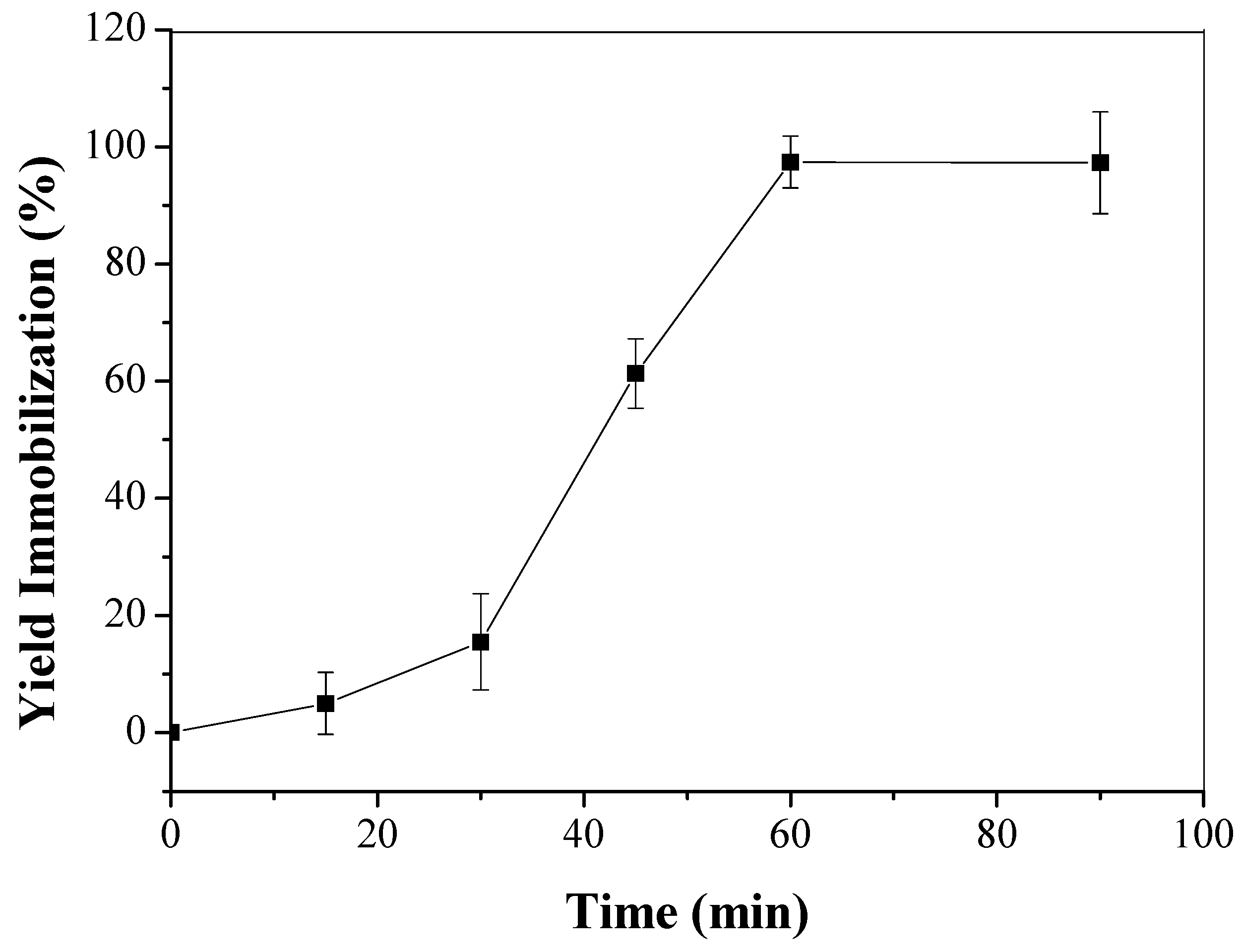
2.3. Immobilization of LU on CTA
2.4. Enzymatic Activity Assay and Protein Dosage
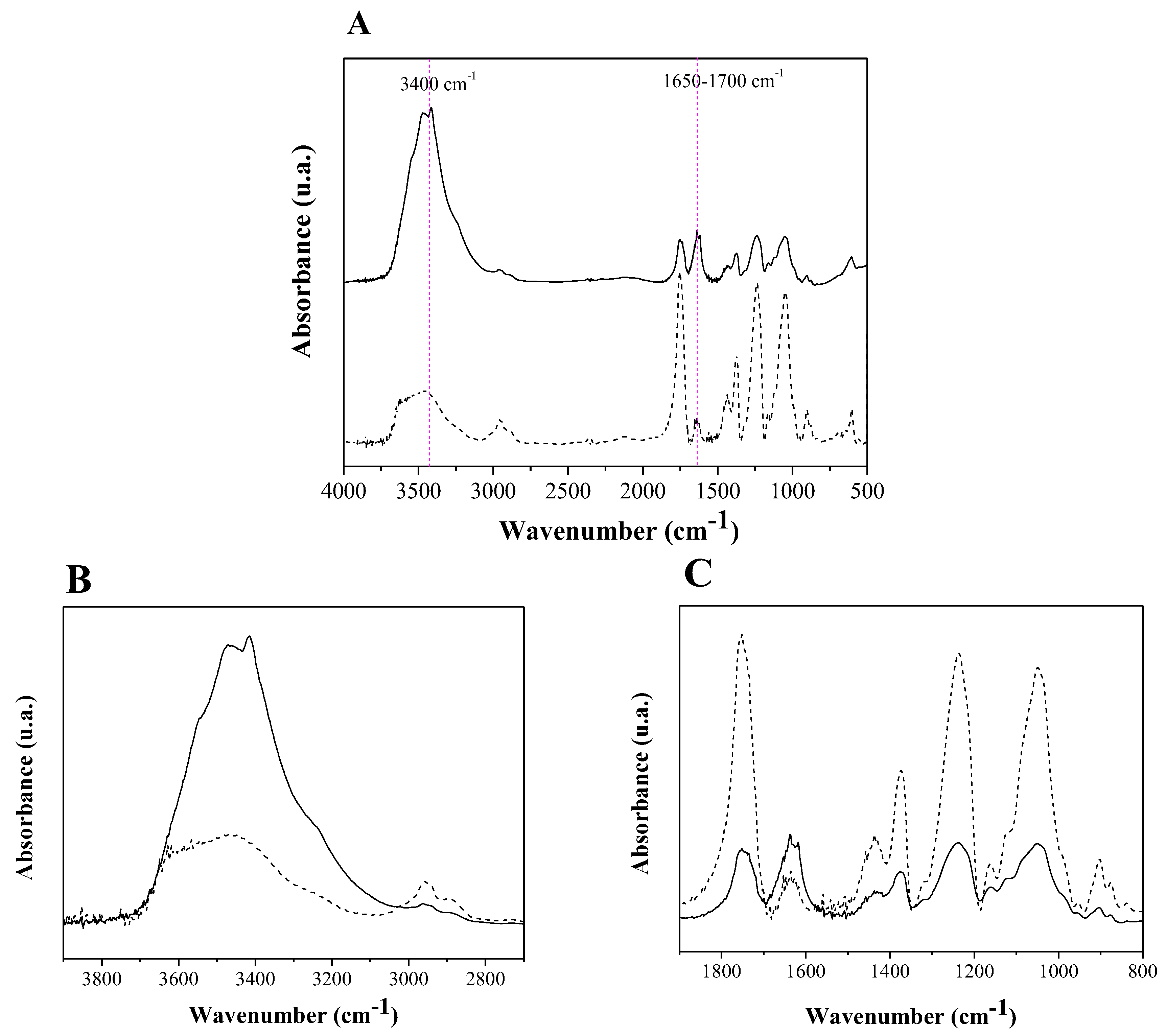
2.5. Biochemical Characterization of Lecitase Ultra Immobilized in CTA (CTA-LU)
2.5.1. Effect of pH and Temperature on Stability of Soluble LU and CTA-LU
2.5.2. Effect of Organic Solvents on Stability of CTA-LU
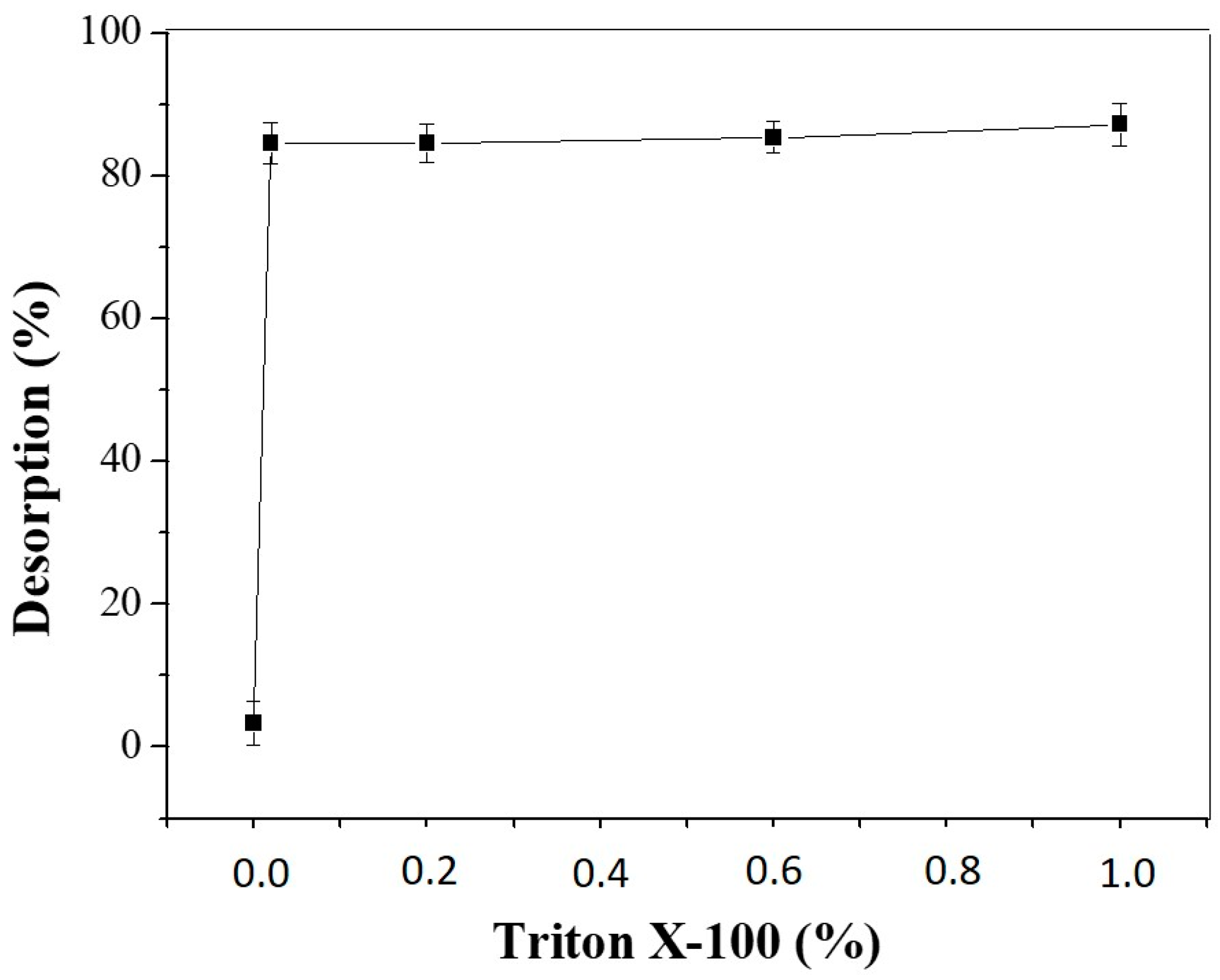
2.5.3. Desorption of LU from CTA
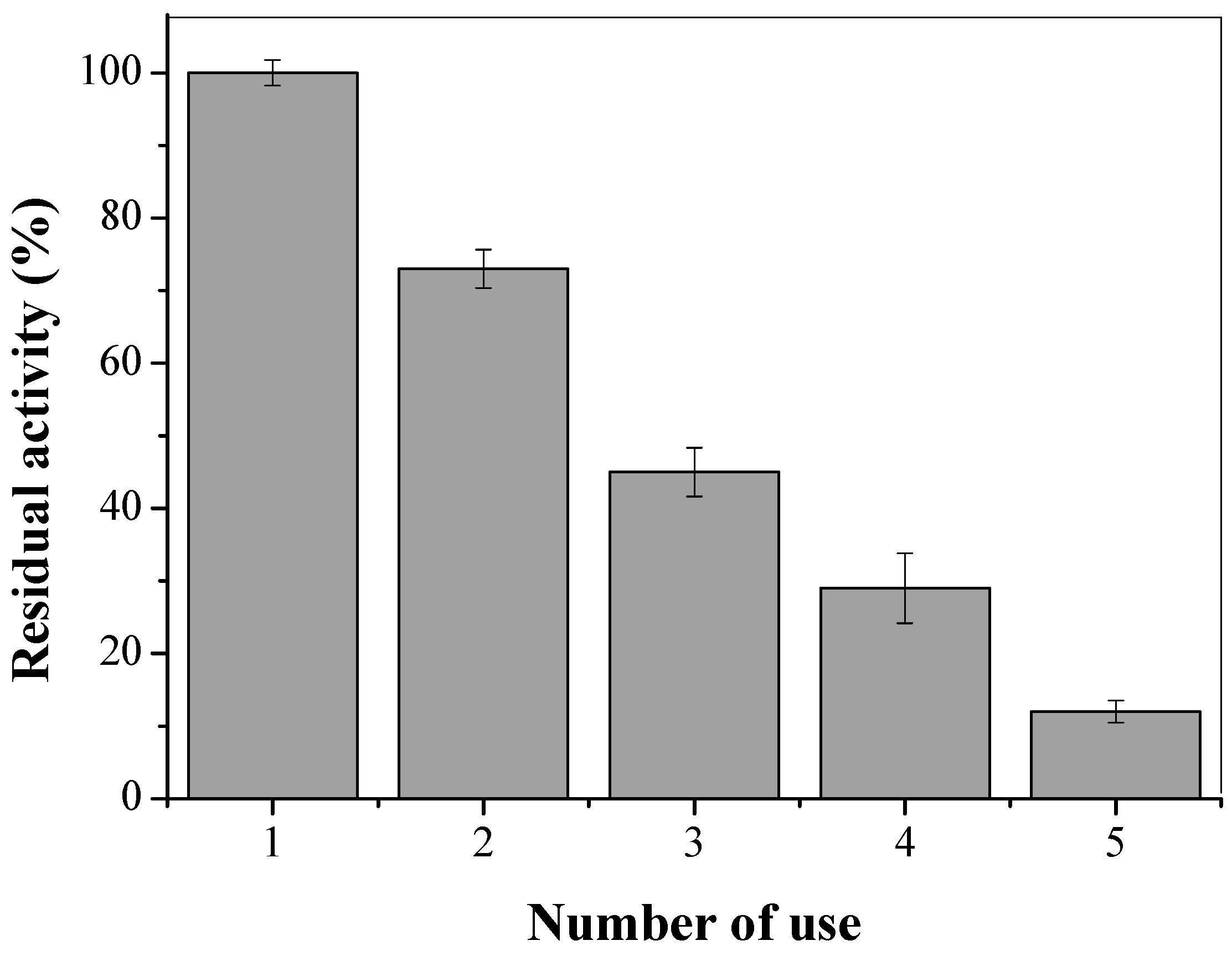
2.6. Reusability of CTA-LU
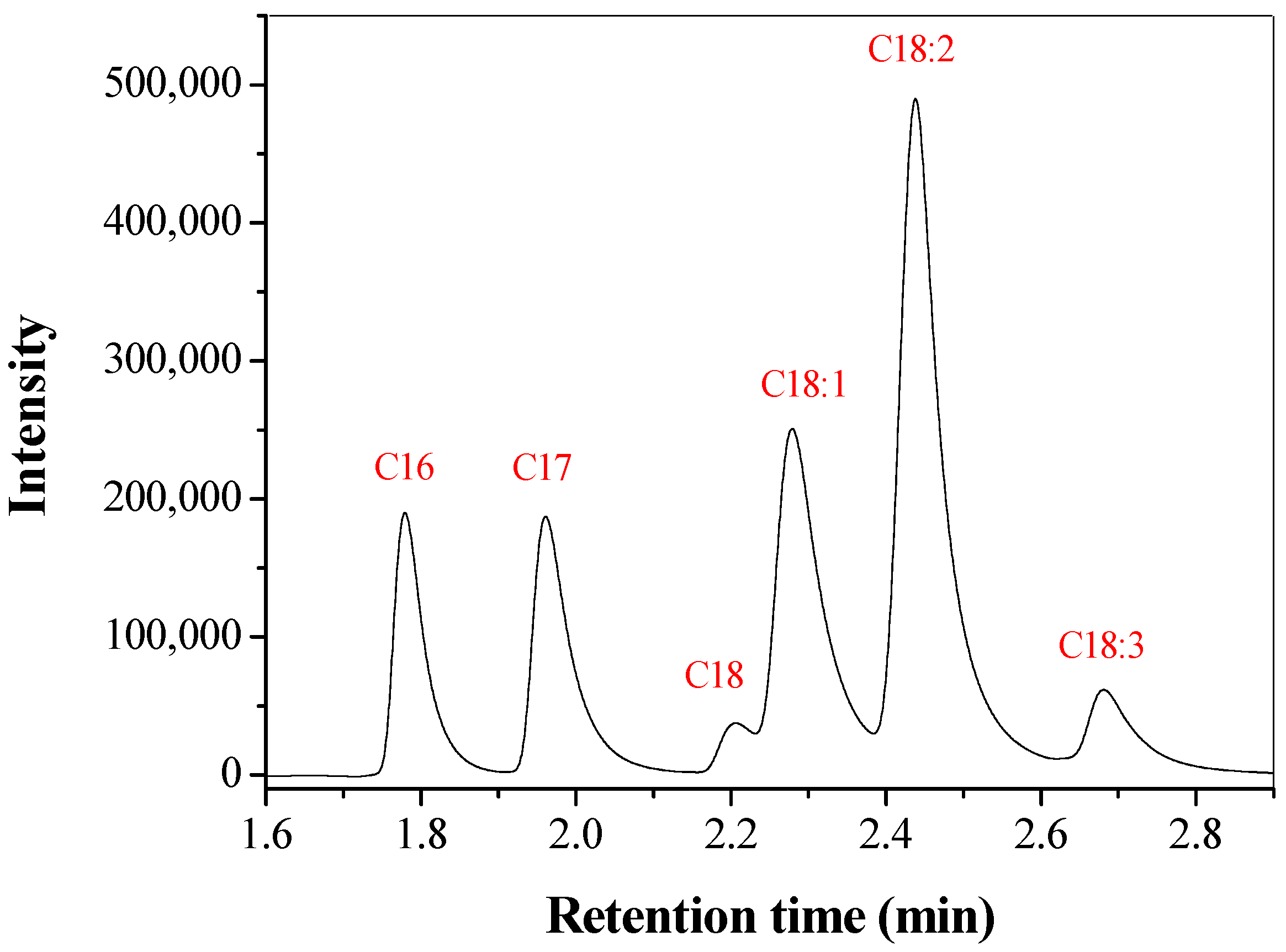
2.7. Methanolysis Product of Soybean Oil Catalyzed by LU Immobilized in CTA
3. Results and Discussion
3.1. Production and Characterization of CA
3.2. Enzyme Immobilization
3.3. Biochemical Characterization of CTA-LU
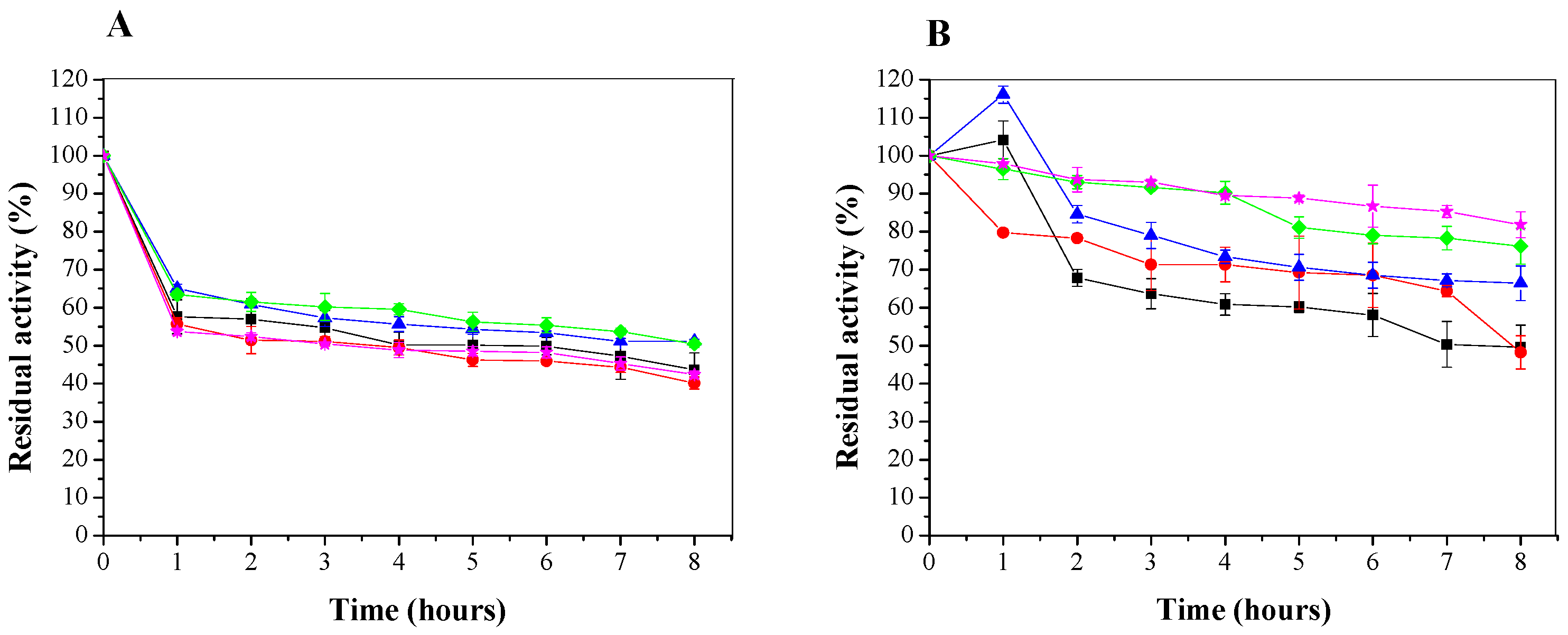
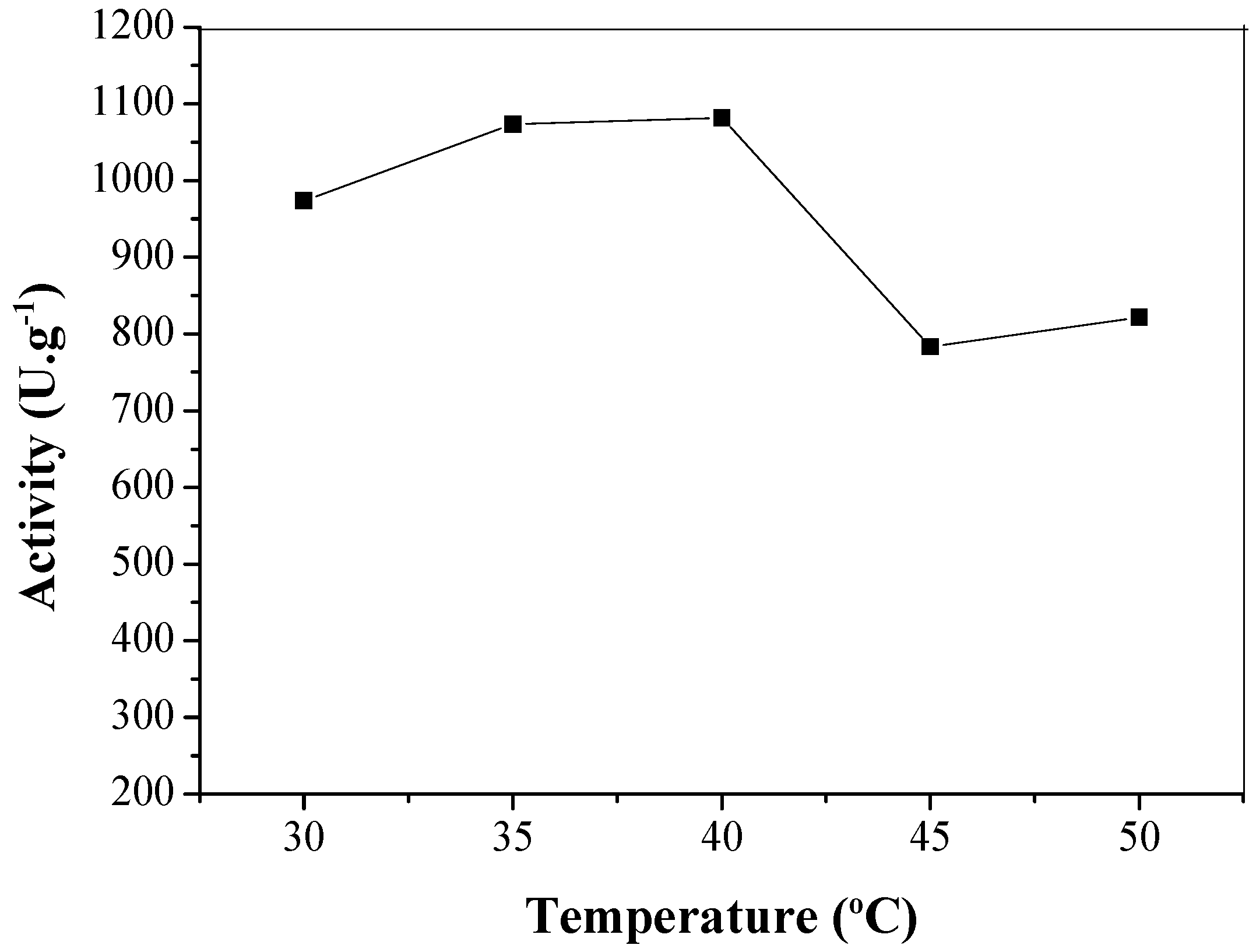
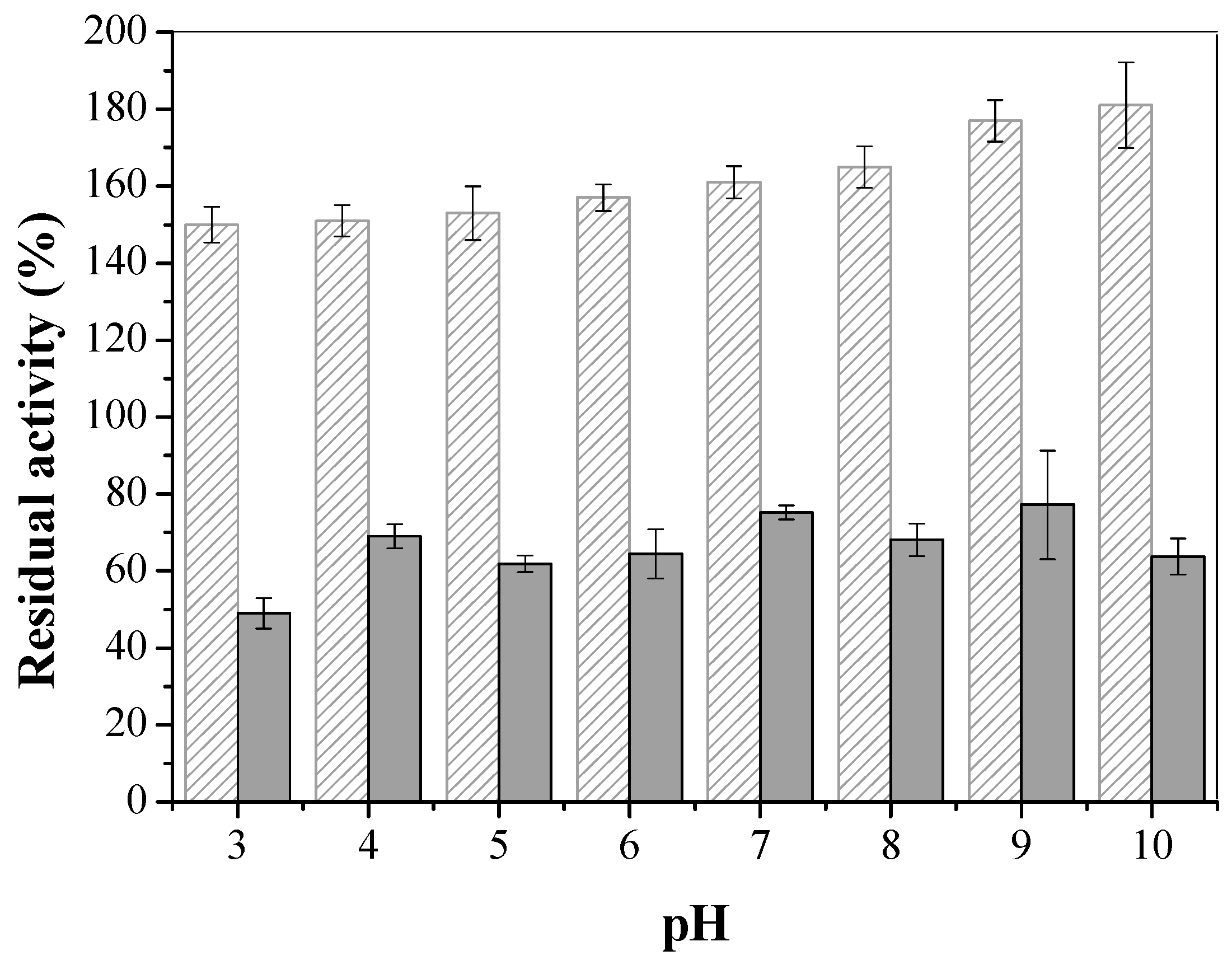
3.3.1. Temperature and pH Effects on Stability of Soluble LU and CTA-LU
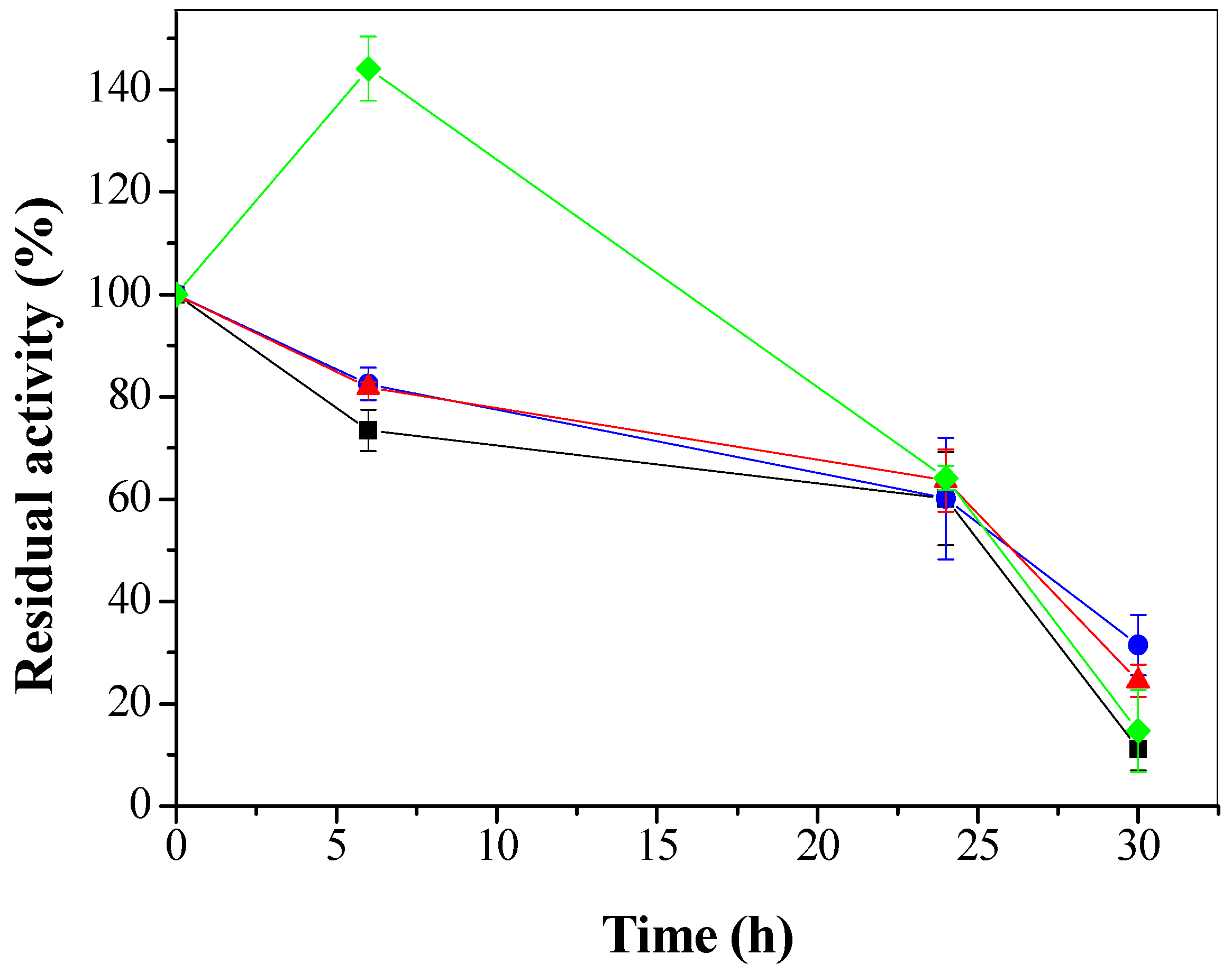
3.3.2. Effect of Organic Solvents on Stability of CTA-LU
3.3.3. Enzymatic Desorption
3.4. CTA-LU Reuse
3.5. Methanolysis Product of Soybean Oil Catalyzed by LU Immobilized in CTA
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ribeiro, S.D.; Guimes, R.F.; Meneguin, A.B.; Prezotti, F.G.; Boni, F.I.; Cury, B.S.F.; Gremião, M.P.D. Cellulose triacetate films obtained from sugarcane bagasse: Evaluation as coating and mucoadhesive material for drug delivery systems. Carbohydr. Polym. 2016, 152, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Filho, G.; Ribeiro, S.D.; da Silva Meireles, C.; da Silva, L.G.; Ruggiero, R.; Ferreira, M.F.; Cerqueira, D.A.; de Assunção, R.M.N.; Zeni, M.; Polleto, P. Release of doxycycline through cellulose acetate symmetric and asymmetric membranes produced from recycled agroindustrial residue: Sugarcane bagasse. Ind. Crops Prod. 2011, 33, 566–571. [Google Scholar] [CrossRef]
- Candido, R.G.; Godoy, G.G.; Gonçalves, A.R. Characterization and application of cellulose acetate synthesized from sugarcane bagasse. Carbohydr. Polym. 2017, 167, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.N.; Dowd, M.K.; Selling, G.W.; Biswas, A. Synthesis of cellulose acetate from cotton byproducts. Carbohydr. Polym. 2010, 80, 449–452. [Google Scholar] [CrossRef]
- Je, H.H.; Noh, S.; Hong, S.; Ju, Y.; Kim, J.; Hwanga, D.S. Cellulose nanofibers for magnetically-separable and highly loaded enzyme immobilization. Chem. Eng. J. 2017, 323, 425–433. [Google Scholar] [CrossRef]
- Prasetyo, E.N.; Semlitsch, S.; Nyanhongo, G.S.; Lemmouchi, Y.; Guebitza, G.M. Laccase functionalized cellulose acetate for the removal of toxic combustion products. React. Funct. Polym. 2015, 97, 12–18. [Google Scholar] [CrossRef]
- Huang, W.; Zhan, Y.; Shi, X.; Chen, J.; Deng, H.; Du, Y. Controllable immobilization of naringinase on electrospun cellulose acetate nanofibers and their application to juice debittering. Int. J. Biol. Macromol. 2017, 98, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Meireles, C. Síntese e Caracterização de Membranas de Acetato de Celulose, Obtido do Bagaço de Cana-de-Açúcar, e Blendas de Acetato de Celulose com Poliestireno de Copos Plásticos Descartados. Ph.D. Thesis, Federal University of Uberlandia, Uberlandia, Brazil, 2007. [Google Scholar]
- Zhou, X.; Lin, X.; White, K.L.; Lin, S.; Wu, H.; Cao, S.; Huang, L.; Chen, L. Effect of the degree of substitution on the hydrophobicity of acetylated cellulose for production of liquid marbles. Cellulose 2016, 23, 811–821. [Google Scholar] [CrossRef]
- Kosaka, P.M.; Kawano, Y.; El Seoud, O.A.; Petri, D.F.S. Catalytic Activity of Lipase Immobilized onto Ultrathin Films of Cellulose Esters. Langmuir 2007, 23, 12167–12173. [Google Scholar] [CrossRef] [PubMed]
- Cerveró, J.M.; Álvarez, J.R.; Luque, S. Novozym 435-catalyzed synthesis of fatty acid ethyl esters from soybean oil for biodiesel production. Biomass Bioenergy 2014, 61, 131–137. [Google Scholar] [CrossRef]
- Haigh, K.F.; Vladisavljević, G.T.; Reynolds, J.C.; Nagy, Z.; Saha, B. Kinetics of the pre-treatment of used cooking oil using Novozyme 435 for biodiesel production. Chem. Eng. Res. Des. 2014, 92, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Martín, E.; Otero, C. Different enzyme requirements for the synthesis of biodiesel: Novozym® 435 and Lipozyme® TL IM. Bioresour. Technol. 2008, 99, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Castillo López, B.; Esteban Cerdán, L.; Robles Medina, A.; Navarro López, E.; Martín Valverde, L.; Hita Peña, E.; González Moreno, P.A.; Molina Grima, E. Production of biodiesel from vegetable oil and microalgae by fatty acid extraction and enzymatic esterification. J. Biosci. Bioeng. 2015, 119, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Verdasco-Martín, C.M.; Villalba, M.; dos Santos, J.C.S.; Tobajas, M.; Fernandez-Lafuente, R.; Otero, C. Effect of chemical modification of Novozym 435 on its performance in the alcoholysis of camelina oil. Biochem. Eng. J. 2016, 111, 75–86. [Google Scholar] [CrossRef]
- Manurung, R.; Hasibuan, R.; Taslim, T.; Rahayu, N.S.; Darusmy, A. Enzymatic Transesterification of DPO to Produce Biodiesel by Using Lipozyme RM IM in Ionic Liquid System. Procedia Soc. Behav. Sci. 2015, 195, 2485–2491. [Google Scholar] [CrossRef]
- Ungcharoenwiwat, P.; Canyuk, B.; H-Kittikun, A. Synthesis of jatropha oil based wax esters using an immobilized lipase from Burkholderia sp. EQ3 and Lipozyme RM IM. Process Biochem. 2016, 51, 392–398. [Google Scholar] [CrossRef]
- Cipolatti, E.P.; Manoel, E.A.; Fernandez-Lafuente, R.; Freire, D.M.G. Support engineering: Relation between development of new supports for immobilization of lipases and their applications. Biotechnol. Res. Innov. 2017, 1–9. [Google Scholar] [CrossRef]
- Badgujar, K.C.; Bhanage, B.M. Synthesis of geranyl acetate in non-aqueous media using immobilized Pseudomonas cepacia lipase on biodegradable polymer film: Kinetic modelling and chain length effect study. Process Biochem. 2014, 49, 1304–1313. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, S.; Kim, S.H.; Kim, J.H.; Yu, H.; Kim, H.J.; Yang, Y.-H.; Kan, E.; Kim, Y.H.; Lee, S.H. Biocompatible cellulose nanocrystals as supports to immobilize lipase. J. Mol. Catal. B Enzym. 2015, 122, 170–178. [Google Scholar] [CrossRef]
- Kosaka, P.M. Aplicações e Caracterização de Ésteres de Celulose. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brasil, 2007. [Google Scholar]
- Kosaka, P.M.; Kawano, Y.; Salvadori, M.C.; Petri, D.F.S. Characterization of Ultrathin Films of Cellulose Esters. Cellulose 2005, 12, 351–359. [Google Scholar] [CrossRef]
- Baron, A.M. Preparação e Caracterização de Lipases Imobilizadas Para Utilização em Biocatálise. Ph.D. Thesis, Federal University of Paraná, Curitiba, Brazil, 2008. [Google Scholar]
- Palomo, J.M.; Segura, R.L.; Mateo, C.; Terreni, M.; Guisan, J.M.; Fernández-Lafuente, R. Synthesis of enantiomerically pure glycidol via a fully enantioselective lipase-catalyzed resolution. Tetrahedron Asymmetry 2005, 16, 869–874. [Google Scholar] [CrossRef]
- Yousefi, M.; Mohammadi, M.; Habibi, Z. Enantioselective resolution of racemic ibuprofen esters using different lipases immobilized on octyl sepharose. J. Mol. Catal. B Enzym. 2014, 104, 87–94. [Google Scholar] [CrossRef]
- Velasco-Lozano, S.; López-Gallego, F.; Rocha-Martin, J.; Guisán, J.M.; Favela-Torres, E. Improving enantioselectivity of lipase from Candida rugosa by carrier-bound and carrier-free immobilization. J. Mol. Catal. B Enzym. 2016, 130, 32–39. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Nigam, S.; Mehrotra, S.; Vani, B.; Mehrotra, R. Lipase Immobilization Techniques for Biodiesel Production: An Overview. IBIMA Publ. Int. J. Renew. Energy Biofuels 2014, 2014. [Google Scholar] [CrossRef]
- Fernandez-Lafuente, R.; Armisén, P.; Sabuquillo, P.; Fernández-Lorente, G.; Guisán, J.M. Immobilization of lipases by selective adsorption on hydrophobic supports. Chem. Phys. Lipids 1998, 93, 185–197. [Google Scholar] [CrossRef]
- Derewenda, U.; Brzozowski, A.M.; Lawson, D.M.; Derewenda, Z.S. Catalysis at the interface: The anatomy of a conformational change in a triglyceride lipase. Biochemistry 1992, 31, 1532–1541. [Google Scholar] [CrossRef] [PubMed]
- Palomo, J.M.; Munõz, G.; Fernández-Lorente, G.; Mateo, C.; Fernández-Lafuente, R.; Guisán, J.M. Interfacial adsorption of lipases on very hydrophobic support (octadecyl–Sepabeads): Immobilization, hyperactivation and stabilization of the open form of lipases. J. Mol. Catal. B Enzym. 2002, 19–20, 279–286. [Google Scholar] [CrossRef]
- Miled, N.; Beisson, F.; de Caro, J.; de Caro, A.; Arondel, V.; Verger, R. Interfacial catalysis by lipases. J. Mol. Catal. B Enzym. 2001, 11, 165–171. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; Zhao, Q.; Cui, C.; Fu, M.; Zhao, M. Immobilisation of lecitase® ultra for production of diacylglycerols by glycerolysis of soybean oil. Food Chem. 2012, 134, 301–307. [Google Scholar] [CrossRef]
- Dos Santos, J.C.S.; Garcia-Galan, C.; Rodrigues, R.C.; de Sant’Ana, H.B.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Stabilizing hyperactivated lecitase structures through physical treatment with ionic polymers. Process Biochem. 2014, 49, 1511–1515. [Google Scholar] [CrossRef]
- Yang, J.-G.; Wang, Y.-H.; Yang, B.; Mainda, G.; Guo, Y. Degumming of vegetable oil by a new microbial lipase. Food Technol. Biotechnol. 2016, 44, 101–104. [Google Scholar]
- Yang, B.; Wang, Y.-H.; Yang, J.-G. Optimization of enzymatic degumming process for rapeseed oil. J. Am. Oil Chem. Soc. 2006, 83, 653–658. [Google Scholar] [CrossRef]
- Lamas, D.L.; Crapiste, G.H.; Constenla, D.T. Changes in quality and composition of sunflower oil during enzymatic degumming process. LWT Food Sci. Technol. 2014, 58, 71–76. [Google Scholar] [CrossRef]
- Sampaio, K.A.; Zyaykina, N.; Wozniak, B.; Tsukamoto, J.; Greyt, W.D.; Stevens, C.V. Enzymatic degumming: Degumming efficiency versus yield increase. Eur. J. Lipid Sci. Technol. 2015, 117, 81–86. [Google Scholar] [CrossRef]
- Li, Z.; Liu, H.; Zhao, G.; Wang, P.; Wang, L.; Wu, H.; Fang, X.; Sun, X.; Wu, X.; Zheng, Z. Enhancing the performance of a phospholipase A1 for oil degumming by bio-imprinting and immobilization. J. Mol. Catal. B Enzym. 2016, 123, 122–131. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; Zhao, Q.; Zhao, M. Production of palm oil-based diacylglycerol using Lecitase Ultra-catalyzed glycerolysis and molecular distillation. Food Sci. Biotechnol. 2014, 23, 365–371. [Google Scholar] [CrossRef]
- Gonçalves, K.M.; Sutili, F.K.; Júnior, I.I.; Flores, M.C.; Soter de Mariz e Miranda, L.; Leal, I.C.R.; Cordeiro, Y.; Luque, R.; Alves de Souza, R.O.M. A Comprehensive study on the activity and deactivation of immobilized lecitase ultra in esterifications of food waste streams to monoacylglycerols. ChemSusChem 2013, 6, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Hu, C.; Cao, Q.; Yang, X.; Zhao, M. Preparation of diacylglycerol-enriched oil from free fatty acids using lecitase ultra-catalyzed esterification. J. Am. Oil Chem. Soc. 2011, 88, 1557–1565. [Google Scholar] [CrossRef]
- Mishra, M.K.; Harini, M.; Kumaraguru, T.; Lakshmi Prasanna, T.; Fadnavis, N.W. A porous vessel bioreactor for gel entrapped biocatalysts: Kinetic resolution of trans-methyl (4-methoxyphenyl)glycidate by Lecitase® Ultra in gelatin organogel (Gelozyme). J. Mol. Catal. B Enzym. 2011, 71, 56–62. [Google Scholar] [CrossRef]
- Mishra, M.K.; Kumaraguru, T.; Sheelu, G.; Fadnavis, N.W. Lipase activity of Lecitase® Ultra: Characterization and applications in enantioselective reactions. Tetrahedron Asymmetry 2009, 20, 2854–2860. [Google Scholar] [CrossRef]
- Hirata, D.B.; Albuquerque, T.L.; Rueda, N.; Virgen-Ortíz, J.J.; Tacias-Pascacio, V.G.; Fernandez-Lafuente, R. Evaluation of different immobilized lipases in transesterification reactions using tributyrin: Advantages of the heterofunctional octyl agarose beads. J. Mol. Catal. B Enzym. 2016, 133, 117–123. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Virgen-Ortíz, J.J.; Jiménez-Pérez, M.; Yates, M.; Torrestiana-Sanchez, B.; Rosales-Quintero, A.; Fernandez-Lafuente, R. Evaluation of different lipase biocatalysts in the production of biodiesel from used cooking oil: Critical role of the immobilization support. Fuel 2017, 200, 1–10. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Palomo, J.M.; Guisan, J.M.; Fernandez-Lafuente, R. Effect of the immobilization protocol in the activity, stability, and enantioslectivity of Lecitase® Ultra. J. Mol. Catal. B Enzym. 2007, 47, 99–104. [Google Scholar] [CrossRef]
- Ikeda, Y.; Kurokawa, Y. Synthesis of geranyl acetate by lipase entrap-immobilized in cellulose acetate-TiO2 gel fiber. J. Am. Oil Chem. Soc. 2001, 78, 1099–1103. [Google Scholar] [CrossRef]
- Singh, A.K.; Mukhopadhyay, M. Immobilization of Candida antarctica lipase onto cellulose acetate-coated Fe2O3 nanoparticles for glycerolysis of olive oil. Korean J. Chem. Eng. 2014, 31, 1225–1232. [Google Scholar] [CrossRef]
- Cerqueira, D.A.; Rodrigues Filho, G.; da Silva Meireles, C. Optimization of sugarcane bagasse cellulose acetylation. Carbohydr. Polym. 2007, 69, 579–582. [Google Scholar] [CrossRef]
- Hurtubise, F.G. The analytical and structural aspects of the infrared spectroscopy of cellulose acetate. Tappi 1962, 45. [Google Scholar]
- Cerqueira, D.A.; Rodrigues Filho, G.; de A. Carvalho, R.; Valente, A.J.M. Caracterização de acetato de celulose obtido a partir do bagaço de cana-de-açúcar por 1H-RMN. Polímeros 2010, 20, 85–91. [Google Scholar] [CrossRef]
- Pereira, M.G.; Facchini, F.D.A.; Filó, L.E.C.; Polizeli, A.M.; Vici, A.C.; Jorge, J.A.; Fernandez-Lorente, G.; Pessela, B.C.; Guisan, J.M.; Polizeli, M.; et al. Immobilized lipase from Hypocrea pseudokoningii on hydrophobic and ionic supports: Determination of thermal and organic solvent stabilities for applications in the oleochemical industry. Process Biochem. 2015, 50, 561–570. [Google Scholar] [CrossRef]
- Pencreac’h, G.; Baratti, J.C. Hydrolysis of p-nitrophenyl palmitate in n-heptane by the Pseudomonas cepacia lipase: A simple test for the determination of lipase activity in organic media. Enzym. Microb. Technol. 1996, 18, 417–422. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Batista, K.A.; Purcena, L.L.A.; Alves, G.L.; Fernandes, K.F. A pectin-lipase derivative as alternative copolymer for lipase assay. J. Mol. Catal. B Enzym. 2014, 102, 25–32. [Google Scholar] [CrossRef]
- Ribeiro, E.A.M.; Filho, G.R.; Vieira, J.G.; de Sousa, R.M.F.; de Assunção, R.M.N.; da Silva Meireles, C.; Duarte, J.; Zeni, M. Characterization of asymmetric membranes of cellulose acetate from recycling of residue corn stover for use in ultrafiltration. Quim. Nova 2014, 37, 385–391. [Google Scholar] [CrossRef]
- Da Cruz, A.C.; da Silva Meireles, C.; Ribeiro, S.D.; Rodrigues Filho, G.; de Assunção, R.M.N.; Cerqueira, D.A.; Zeni, M.; Poletto, P. Utilização do acetato de celulose produzido a partir da celulose extraída do caroço de manga como matriz para produção de sistemas microparticulados. Quim. Nova 2011, 34, 385–389. [Google Scholar] [CrossRef]
- Ferreira Júnior, M.F.; Mundim, E.A.R.; Filho, G.R.; da Silva Meireles, C.; Cerqueira, D.A.; de Assunção, R.M.N.; Marcolin, M.; Zeni, M. SEM study of the morphology of asymmetric cellulose acetate membranes produced from recycled agro-industrial residues: Sugarcane bagasse and mango seeds. Polym. Bull. 2011, 66, 377–389. [Google Scholar] [CrossRef]
- Fischer, S.; Thümmler, K.; Volkert, B.; Hettrich, K.; Schmidt, I.; Fischer, K. Properties and applications of cellulose acetate. Macromol. Symp. 2008, 262, 89–96. [Google Scholar] [CrossRef]
- Kamide, K.; Okajima, K.; Saito, M. Nuclear Magnetic Resonance Study of Thermodynamic Interaction between Cellulose Acetate and Solvent. Polym. J. 1981, 13, 115–125. [Google Scholar] [CrossRef]
- Puleo, A.C.; Paul, D.R.; Kelley, S.S. The effect of degree of acetylation on gas sorption and transport behavior in cellulose acetate. J. Membr. Sci. 1989, 47, 301–332. [Google Scholar] [CrossRef]
- Manoel, E.A.; dos Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports involves the open form of the enzyme. Enzym. Microb. Technol. 2015, 71, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Aybastıer, Ö.; Demir, C. Optimization of immobilization conditions of Thermomyces lanuginosus lipase on styrene–Divinylbenzene copolymer using response surface methodology. J. Mol. Catal. B Enzym. 2010, 63, 170–178. [Google Scholar] [CrossRef]
- Cipolatti, E.P.; Moreno-Pérez, S.; de Andrade Souza, L.T.; Valério, A.; Guisán, J.M.; de Araújo, P.H.H.; Sayer, C.; Ninow, J.L.; de Oliveira, D.; Pessela, B.C. Synthesis and modification of polyurethane for immobilization of Thermomyces lanuginosus (TLL) lipase for ethanolysis of fish oil in solvent free system. J. Mol. Catal. B Enzym. 2015, 122, 163–169. [Google Scholar] [CrossRef]
- Nicoletti, G.; Cipolatti, E.P.; Valério, A.; Carbonera, N.T.G.; Soares, N.S.; Theilacker, E.; Ninow, J.L.; de Oliveira, D. Evaluation of different methods for immobilization of Candida antarctica lipase B (CalB lipase) in polyurethane foam and its application in the production of geranyl propionate. Bioprocess Biosyst. Eng. 2015, 38, 1739–1748. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Galan, C.; dos Santos, J.C.S.; Barbosa, O.; Torres, R.; Pereira, E.B.; Corberan, V.C.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Tuning of Lecitase features via solid-phase chemical modification: Effect of the immobilization protocol. Process Biochem. 2014, 49, 604–616. [Google Scholar] [CrossRef]
- Salihu, A.; Alam, M.Z. Solvent tolerant lipases: A review. Process Biochem. 2015, 50, 86–96. [Google Scholar] [CrossRef]
- Cabrera, Z.; Fernandez-Lorente, G.; Fernandez-Lafuente, R.; Palomo, J.M.; Guisan, J.M. Novozym 435 displays very different selectivity compared to lipase from Candida antarctica B adsorbed on other hydrophobic supports. J. Mol. Catal. B Enzym. 2009, 57, 171–176. [Google Scholar] [CrossRef]
- Virgen-Ortíz, J.J.; Tacias-Pascacio, V.G.; Hirata, D.B.; Torrestiana-Sanchez, B.; Rosales-Quintero, A.; Fernandez-Lafuente, R. Relevance of substrates and products on the desorption of lipases physically adsorbed on hydrophobic supports. Enzym. Microb. Technol. 2017, 96, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Turati, D.; Morais Júnior, W.; Terrasan, C.; Moreno-Perez, S.; Pessela, B.; Fernandez-Lorente, G.; Guisan, J.; Carmona, E. Immobilization of Lipase from Penicillium sp. Section Gracilenta (CBMAI 1583) on Different Hydrophobic Supports: Modulation of Functional Properties. Molecules 2017, 22, 339. [Google Scholar] [CrossRef] [PubMed]
- Rueda, N.; dos Santos, C.S.; Rodriguez, M.D.; Albuquerque, T.L.; Barbosa, O.; Torres, R.; Ortiz, C.; Fernandez-Lafuente, R. Reversible immobilization of lipases on octyl-glutamic agarose beads: A mixed adsorption that reinforces enzyme immobilization. J. Mol. Catal. B Enzym. 2016, 128, 10–18. [Google Scholar] [CrossRef]
- Vescovi, V.; Kopp, W.; Guisán, J.M.; Giordano, R.L.C.; Mendes, A.A.; Tardioli, P.W. Improved catalytic properties of Candida antarctica lipase B multi-attached on tailor-made hydrophobic silica containing octyl and multifunctional amino-glutaraldehyde spacer arms. Process Biochem. 2016, 51, 2055–2066. [Google Scholar] [CrossRef]
- Bernal, C.; Illanes, A.; Wilson, L. Heterofunctional Hydrophilic-Hydrophobic Porous Silica as Support for Multipoint Covalent Immobilization of Lipases: Application to Lactulose Palmitate Synthesis. Langmuir 2014, 30, 3557–3566. [Google Scholar] [CrossRef] [PubMed]
- Guajardo, N.; Bernal, C.; Wilson, L.; Cabrera, Z. Selectivity of R-α-monobenzoate glycerol synthesis catalyzed by Candida antarctica lipase B immobilized on heterofunctional supports. Process Biochem. 2015, 50, 1870–1877. [Google Scholar] [CrossRef]
- Fernandez-Lopez, L.; Pedrero, S.G.; Lopez-Carrobles, N.; Virgen-Ortíz, J.J.; Gorines, B.C.; Otero, C.; Fernandez-Lafuente, R. Physical crosslinking of lipase from Rhizomucor miehei immobilized on octyl agarose via coating with ionic polymers. Process Biochem. 2017, 54, 81–88. [Google Scholar] [CrossRef]
- Santos, R.C.R.; Vieira, R.B.; Valentini, A. Monitoring the conversion of soybean oil to methyl or ethyl esters using the refractive index with correlation gas chromatography. Microchem. J. 2013, 109, 46–50. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Gerken, H.; Zhang, C.; Hu, Q.; Li, Y. Highly-efficient enzymatic conversion of crude algal oils into biodiesel. Bioresour. Technol. 2014, 172, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Talukder, M.M.R.; Wu, J.C.; Fen, N.M.; Melissa, Y.L.S. Two-step lipase catalysis for production of biodiesel. Biochem. Eng. J. 2010, 49, 207–212. [Google Scholar] [CrossRef]
- Purwanto, M.G.M.; Maretha, M.V.; Wahyudi, M.; Goeltom, M.T. Whole Cell Hydrolysis of Sardine (Sardinella Lemuru) Oil Waste Using Mucor Circinelloides NRRL 1405 Immobilized in Poly-urethane Foam. Procedia Chem. 2015, 14, 256–262. [Google Scholar] [CrossRef]
Sample Availability: Samples of the support are available from the authors. |














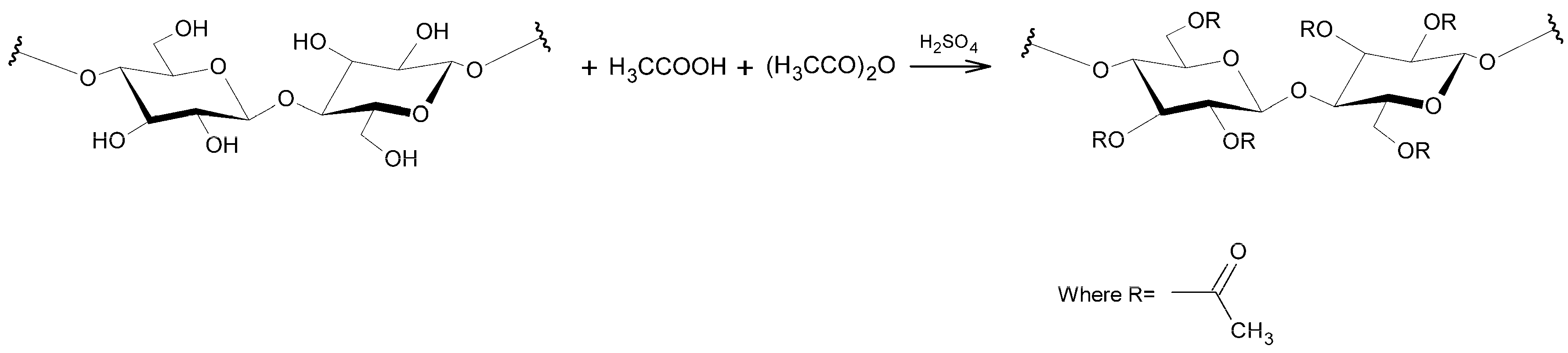{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free Enzyme | Immobilized Enzyme | |||
|---|---|---|---|---|
| T (°C) | kd (h−1) | R2 | kd (h−1) | R2 |
| 30 | 0.062 ± 0.0013 | 0.6591 | 0.050 ± 0.0058 | 0.9199 |
| 35 | 0.058 ± 0.0039 | 0.6791 | 0.036 ± 0.0055 | 0.9568 |
| 40 | 0.069 ± 0.0028 | 0.5706 | 0.024 ± 0.0013 | 0.9825 |
| 45 | 0.071 ± 0.0091 | 0.6526 | 0.076 ± 0.0067 | 0.8887 |
| 50 | 0.077 ± 0.0057 | 0.6506 | 0.064 ± 0.0074 | 0.8225 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, F.B.d.; Morais Júnior, W.G.d.; Silva, C.V.d.; Vieira, A.T.; Batista, A.C.F.; Faria, A.M.d.; Assunção, R.M.N. Preparation and Characterization of Cellulose Triacetate as Support for Lecitase Ultra Immobilization. Molecules 2017, 22, 1930. https://doi.org/10.3390/molecules22111930
Silva FBd, Morais Júnior WGd, Silva CVd, Vieira AT, Batista ACF, Faria AMd, Assunção RMN. Preparation and Characterization of Cellulose Triacetate as Support for Lecitase Ultra Immobilization. Molecules. 2017; 22(11):1930. https://doi.org/10.3390/molecules22111930
Chicago/Turabian StyleSilva, Francielle Batista da, Wilson Galvão de Morais Júnior, Cleuzilene Vieira da Silva, Andressa Tironi Vieira, Antônio Carlos Ferreira Batista, Anízio Márcio de Faria, and Rosana Maria Nascimento Assunção. 2017. "Preparation and Characterization of Cellulose Triacetate as Support for Lecitase Ultra Immobilization" Molecules 22, no. 11: 1930. https://doi.org/10.3390/molecules22111930