
Biosynthesis of Fluorescent β Subunits of C-Phycocyanin from Spirulina subsalsa in Escherichia coli, and Their Antioxidant Properties

Abstract
:1. Introduction
2. Results
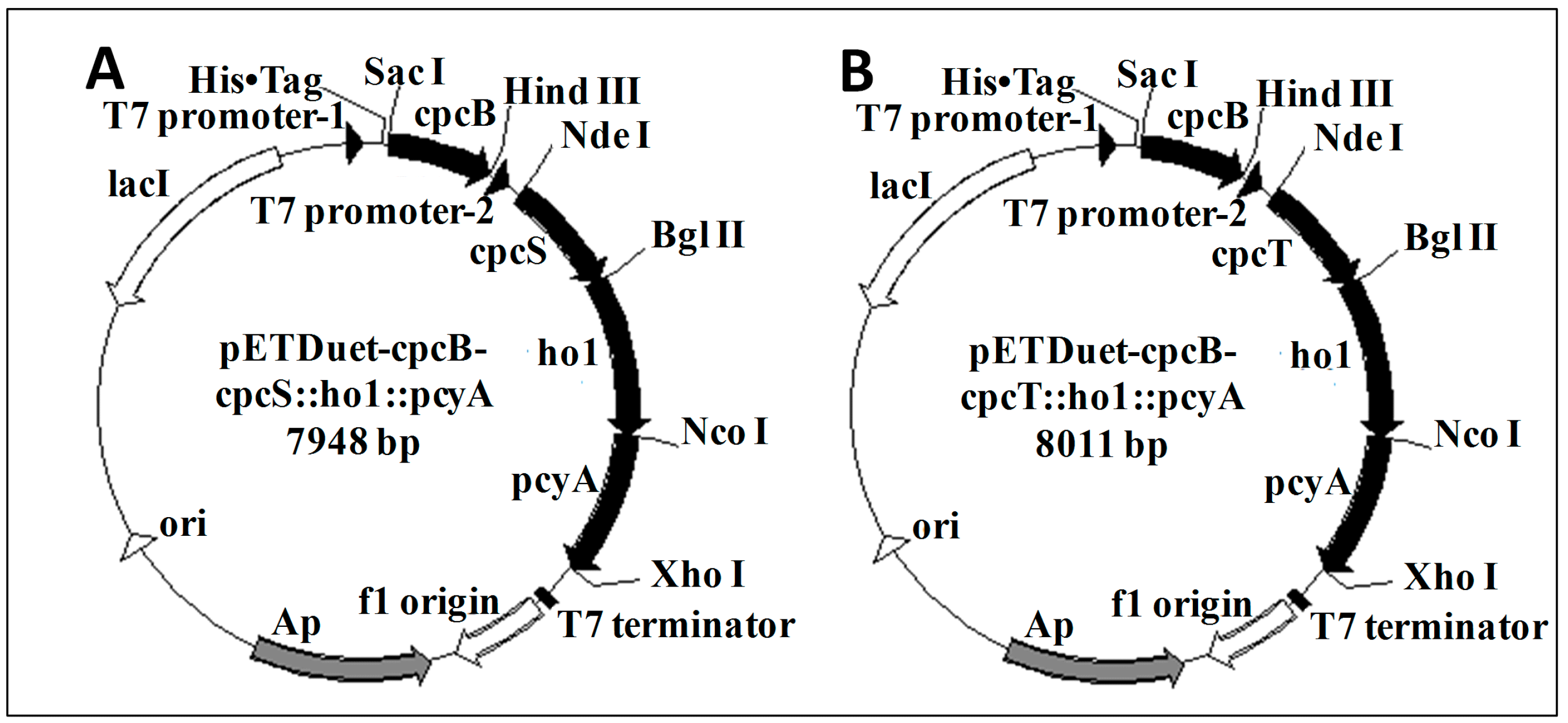
2.1. Cloning of the cpcB Gene and Plasmid Construction
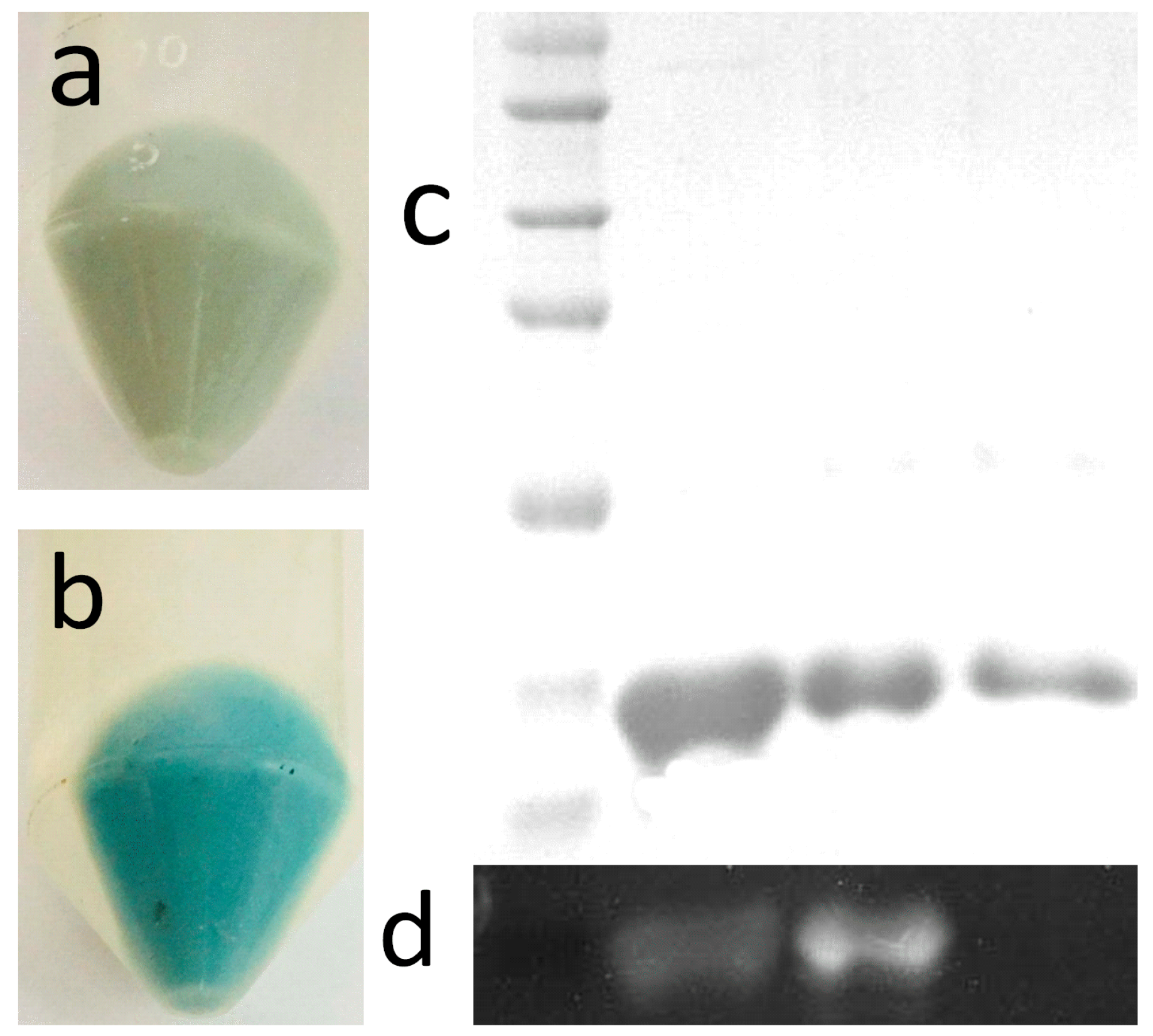
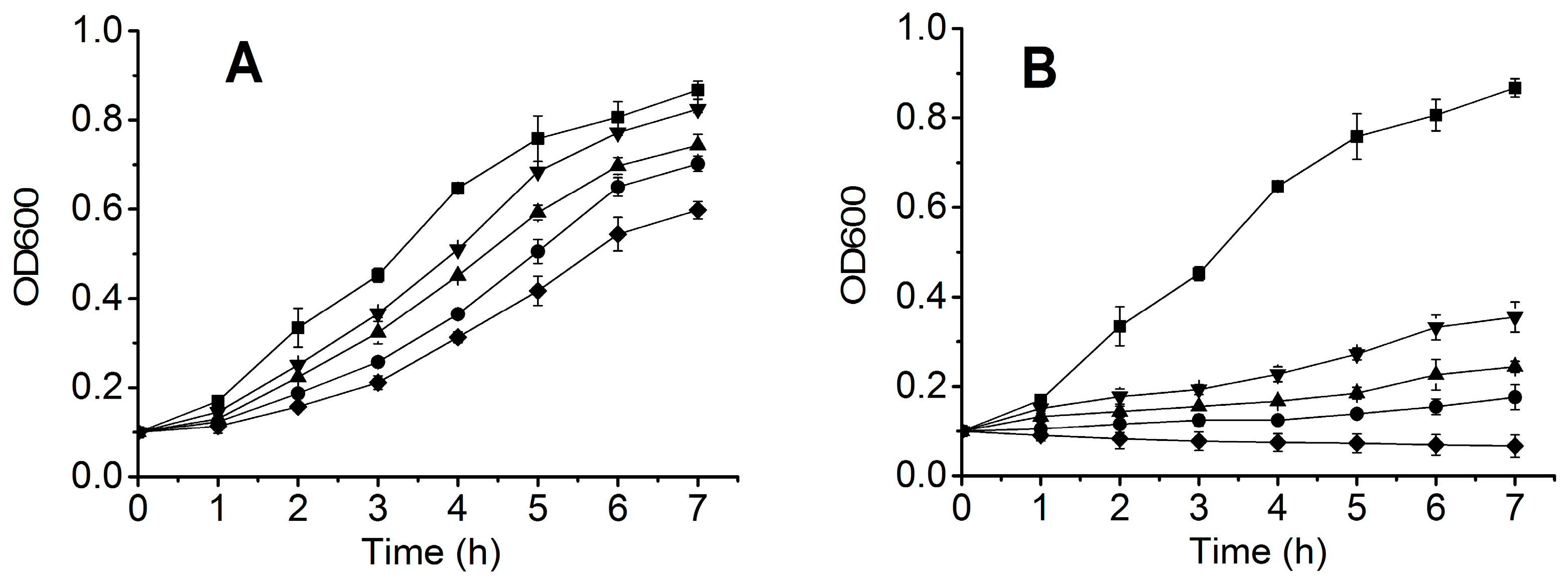
2.2. Expression and Purification
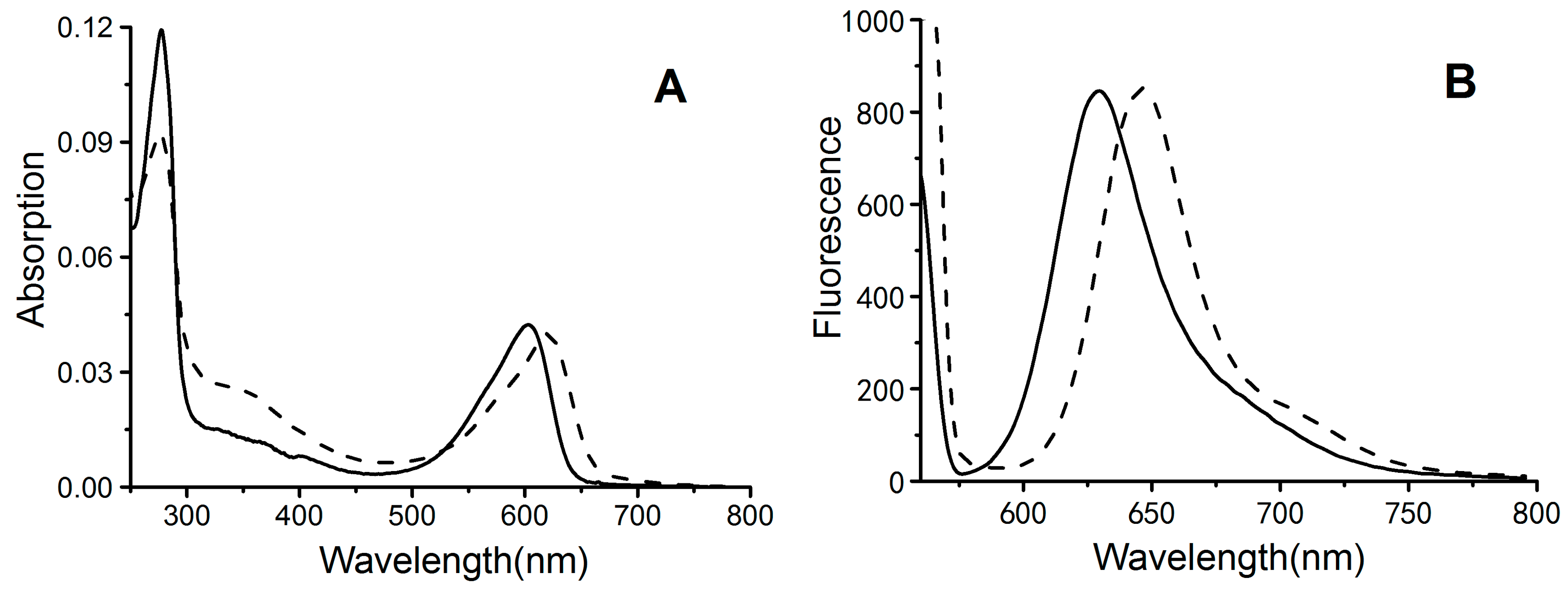
2.3. Absorbance and Fluorescence Spectrometry
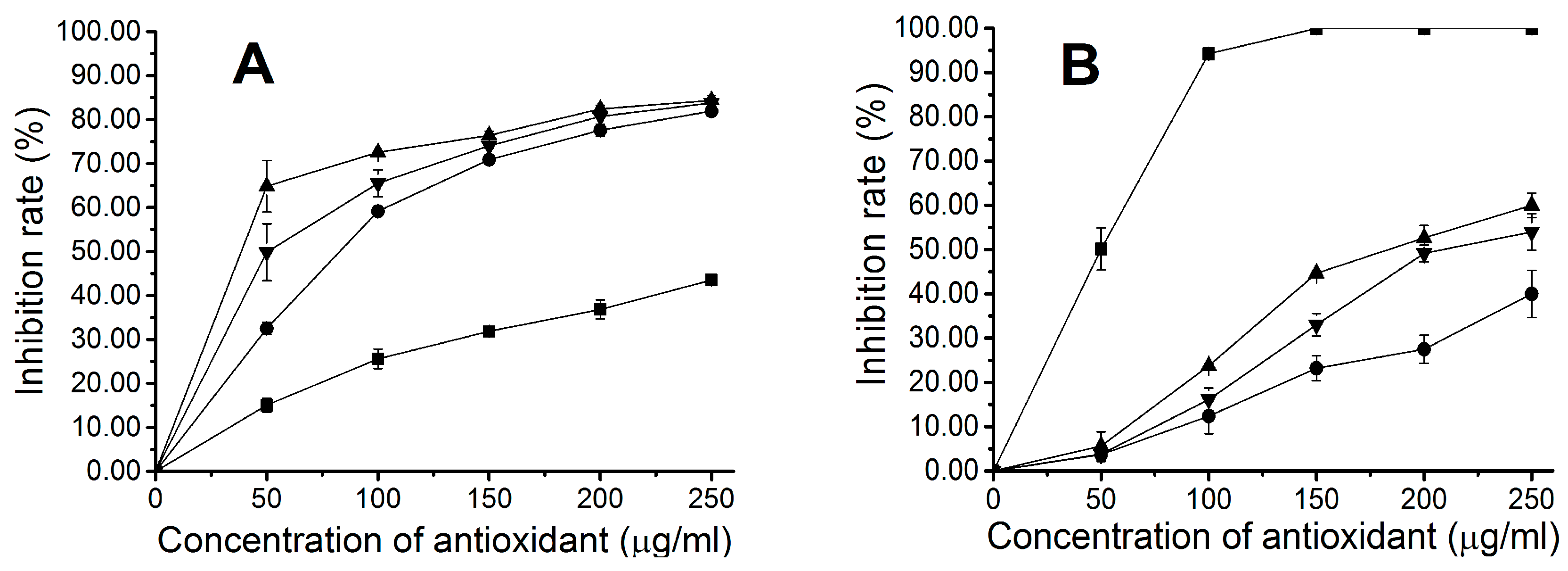
2.4. Antioxidant Activity of Fluorescent Phycocyanin
3. Discussion
4. Materials and Methods
4.1. Cyanobacterial Cultivation and DNA Extraction
4.2. Primer Design and PCR Amplification
4.3. Construction of the Expression Vector and Transformation
4.4. Protein Expression and Purification
4.5. Electrophoretic Analysis
4.6. Spectral Analyses
4.7. Determination of Hydroxyl Radical Scavenging Activity
4.8. Determination of DPPH Free Radical Scavenging Activity
4.9. Inhibition of the Cellular Oxidative Injury by the Recombinant Biliproteins
4.10. Statistics and Data Processing
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Glazer, A.N. Adaptive variations in phycobilisome structure. In Advances in Molecular & Cell Biology; Barber, J., Ed.; JAI Press: London, UK, 1994; Volume 10, pp. 119–149. [Google Scholar]
- Gantt, B.; Grabowski, B.; Cunningham, F.X. Antenna systems of red algae: Phycobilisomes with photosystem II and chlorophyll complexes with photosystem I. In Light-Harvesting Antennas in Photosynthesis; Green, B., Parson, W., Eds.; Kluwer: Dordrecht, The Netherlands, 2003; pp. 307–322. [Google Scholar]
- Grossman, A.R.; Schaefer, M.R.; Chiang, G.G.; Collier, J.L. The phycobilisome, a light-harvesting complex responsive to environmental conditions. Microbiol. Rev. 1993, 57, 725–749. [Google Scholar] [PubMed]
- Sidler, W.A. The Molecular Biology of Cyanobacteria; Springer Netherlands: Dordrecht, The Netherlands, 1994; pp. 139–216. [Google Scholar]
- Gao, X.; Wei, T.D.; Zhang, N.; Xie, B.B.; Su, H.N.; Zhang, X.Y.; Chen, X.L.; Zhou, B.C.; Wang, Z.X.; Wu, J.W.; et al. Molecular insights into the terminal energy acceptor in cyanobacterial phycobilisome. Mol. Microbiol. 2012, 85, 907–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazer, A.N. Phycobiliproteins. In Chemicals from Microalgae; Cohen, Z., Ed.; Taylor and Francis Ltd.: Oxford, UK, 1999; pp. 262–280. [Google Scholar]
- Beale, S.I. Biosynthesis of phycobilins. Chem. Rev. 1993, 93, 785–802. [Google Scholar] [CrossRef]
- Frankenberg, N.; Lagarias, J.C. Biosynthesis and Biological Functions of Bilins. In The Porphyrin Handbook; Kadish, K.M., Smith, K.M., Guilard, R., Eds.; Academic Press: Amsterdam, The Netherlands, 2003; pp. 211–236. [Google Scholar]
- Fairchild, C.D.; Zhao, J.; Zhou, J.; Colson, S.E.; Bryant, D.A.; Glazer, A.N. Phycocyanin alpha-subunit phycocyanobilin lyase. Proc. Natl. Acad. Sci. USA 1992, 89, 7017–7021. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.H.; Su, P.; Tu, J.M.; Wang, X.; Liu, H.; Plöscher, M.; Eichacker, L.; Yang, B.; Zhou, M.; Scheer, H. Phycobilin: Cystein-84 biliprotein lyase, a near-universal lyase for cysteine-84-binding sites in cyanobacterial phycobiliproteins. Proc. Natl. Acad. Sci. USA 2007, 104, 14300–14305. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Chang, K.; Luo, J.; Zhou, M.; Scheer, H.; Zhao, K.H. Modular Generation of Fluorescent Phycobiliproteins. Photochem. Photobiol. Sci. 2013, 12, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Penton-Rol, G.; Marin-Prida, J.; Pardo-Andreu, G.; Martinez-Sanchez, G.; Acosta-Medina, E.F.; Valdivia-Acosta, A.; Lagumersindez-Denis, N.; Rodriguez-Jimenez, E.; Llopiz-Arzuaga, A.; Lopez-Saura, P.A.; et al. C-Phycocyanin is neuroprotective against global cerebral ischemia/reperfusion injury in gerbils. Brain Res. Bull. 2011, 86, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Vadiraja, B.B.; Gaikwad, N.W.; Madyastha, K.M. Hepatoprotective effect of C-phycocyanin: Protection for carbon tetrachloride and R-(+)-pulegone-mediated hepatotoxicty in rats. Biochem. Biophys. Res. Commun. 1998, 249, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Rojas, B.; Medina-Campos, O.N.; Hernandez-Pando, R.; Negrette-Guzman, M.; Huerta-Yepez, S.; Pedraza-Chaverri, J. C-phycocyanin prevents cisplatin-induced nephrotoxicity through inhibition of oxidative stress. Food Funct. 2014, 5, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Riss, J.; Decorde, K.; Sutra, T.; Delage, M.; Baccou, J.C.; Jouy, N.; Brune, J.P.; Oreal, H.; Cristol, J.P.; Rouanet, J.M. Phycobiliprotein C-phycocyanin from Spirulina platensis is powerfully responsible for reducing oxidative stress and NADPH oxidase expression induced by an atherogenic diet in hamsters. J. Agric. Food Chem. 2007, 55, 7962–7967. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.P.; Sivakumar, J.; Thankappan, B.; Anbarasu, K. C-phycocyanin modulates selenite-induced cataractogenesis in rats. Biol. Trace Elem. Res. 2013, 151, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lissi, E.; Pizarro, M.; Aspee, A.; Romay, C. Kinetics of Phycocyanine Bilin Groups Destruction by Peroxyl Radicals. Free Radic. Biol. Med. 2000, 28, 1051–1055. [Google Scholar] [CrossRef]
- Ge, B.; Qin, S.; Han, L.; Lin, F.; Ren, Y. Antioxidant properties of recombinant allophycocyanin expressed in Escherichia coli. J. Photochem. Photobiol. B 2006, 84, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Pleonsil, P.; Soogarun, S.; Suwanwong, Y. Anti-oxidant activity of holo- and apo-c-phycocyanin and their protective effects on human erythrocytes. Int. J. Biol. Macromol. 2013, 60, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.Y.; Zhang, W.J.; Zhang, X.W.; Li, Y.X.; Wang, J.F.; Lin, H.Z.; Tang, X.X.; Qin, S. A potent anti-oxidant property: Fluorescent recombinant alpha-phycocyanin of Spirulina. J. Appl. Microbiol. 2009, 106, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Li, P.; Chen, X.; Chao, X. Combinatorial biosynthesis of Synechocystis PCC6803 phycocyanin holo-alpha-subunit (CpcA) in Escherichia coli and its activities. Appl. Microbiol. Biotechnol. 2016, 100, 5375–5388. [Google Scholar] [CrossRef] [PubMed]
- Cherdkiatikul, T.; Suwanwong, Y. Production of the alpha and beta Subunits of Spirulina Allophycocyanin and C-Phycocyanin in Escherichia coli: A Comparative Study of Their Antioxidant Activities. J. Biomol. Screen. 2014, 19, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Glazer, A.N. Phycobiliproteins—A family of valuable, widely used fluorophores. J. Appl. Phycol. 1994, 6, 105–112. [Google Scholar] [CrossRef]
- Rahman, D.Y.; Sarian, F.D.; van Wijk, A.; Martinez-Garcia, M.; van der Maarel, M. Thermostable phycocyanin from the red microalga Cyanidioschyzon merolae, a new natural blue food colorant. J. Appl. Phycol. 2017, 29, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; Gonzalez, R.; Ledon, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Oliinyk, O.S.; Chernov, K.G.; Verkhusha, V.V. Bacterial Phytochromes, Cyanobacteriochromes and Allophycocyanins as a Source of Near-Infrared Fluorescent Probes. Int. J. Mol. Sci. 2017, 18, 1691. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, J.; Molinajijón, E.; Medinacampos, O.N.; Rodríguezmuñoz, R.; Reyes, J.L.; Loredo, M.L.; Barreraoviedo, D.; Pinzón, E.; Rodríguezrangel, D.S.; Pedrazachaverri, J. Curcumin prevents cisplatin-induced decrease in the tight and adherens junctions: Relation to oxidative stress. Food Funct. 2015, 7, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Blot, N.; Wu, X.J.; Thomas, J.C.; Zhang, J.; Garczarek, L.; Bohm, S.; Tu, J.M.; Zhou, M.; Ploscher, M.; Eichacker, L.; et al. Phycourobilin in trichromatic phycocyanin from oceanic cyanobacteria is formed post-translationally by a phycoerythrobilin lyase-Isomerase. J. Biol. Chem. 2009, 284, 9290–9298. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Vasquez, Y.M.; Dragomani, T.M.; Kronfel, M.L.; Williams, S.R.; Alvey, R.M.; Bryant, D.A.; Schluchter, W.M. Biosynthesis of cyanobacterial phycobiliproteins in Escherichia coli: Chromophorylation efficiency and specificity of all bilin lyases from Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 2010, 76, 2729–2739. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.H.; Zhang, J.; Tu, J.M.; Bohm, S.; Ploscher, M.; Eichacker, L.; Bubenzer, C.; Scheer, H.; Wang, X.; Zhou, M. Lyase activities of CpcS- and CpcT-like proteins from Nostoc PCC7120 and sequential reconstitution of binding sites of phycoerythrocyanin and phycocyanin beta-subunits. J. Biol. Chem. 2007, 282, 34093–34103. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, P.; Pinero, E.; Villar, A.M. Iron-chelating ability and antioxidant properties of phycocyanin isolated from a protean extract of Spirulina platensis. Food Chem. 2008, 110, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Li, D.; Peng, J.; Fang, J.; Zhang, L.; Liu, L. Cloning, expression and antioxidant activity of a novel collagen from Pelodiscus sinensis. World J. Microbiol. Biotechnol. 2016, 32, 100. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombinant Proteins | Absorption | Fluorescence | Chromophore-Binding Rate (Amax/A280) | IC50 Values (μg/mL) | |
|---|---|---|---|---|---|
| Hydroxyl Radical Scavenging Activity | DPPH Free Radical Scavenging Activity | ||||
| apo-CpcB | 0 | 0 | 0 | 81.82 ± 0.67 | 352.93 ± 26.30 |
| PCB-CpcB(C-82) | 621 | 646 | 0.45 | 38.72 ± 2.48 | 201.00 ± 5.86 |
| PCB-CpcB(C-153) | 602 | 629 | 0.37 | 51.06 ± 6.74 | 240.34 ± 4.03 |
| Ascorbic acid | - | - | - | - | 49.91 ± 0.32 |
| Mannitol | - | - | - | 245.72 ± 9.43 | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.-J.; Yang, H.; Chen, Y.-T.; Li, P.-P. Biosynthesis of Fluorescent β Subunits of C-Phycocyanin from Spirulina subsalsa in Escherichia coli, and Their Antioxidant Properties. Molecules 2018, 23, 1369. https://doi.org/10.3390/molecules23061369
Wu X-J, Yang H, Chen Y-T, Li P-P. Biosynthesis of Fluorescent β Subunits of C-Phycocyanin from Spirulina subsalsa in Escherichia coli, and Their Antioxidant Properties. Molecules. 2018; 23(6):1369. https://doi.org/10.3390/molecules23061369
Chicago/Turabian StyleWu, Xian-Jun, Hong Yang, Yu-Ting Chen, and Ping-Ping Li. 2018. "Biosynthesis of Fluorescent β Subunits of C-Phycocyanin from Spirulina subsalsa in Escherichia coli, and Their Antioxidant Properties" Molecules 23, no. 6: 1369. https://doi.org/10.3390/molecules23061369