The Immunomodulatory and Neuroprotective Effects of Mesenchymal Stem Cells (MSCs) in Experimental Autoimmune Encephalomyelitis (EAE): A Model of Multiple Sclerosis (MS)


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Autoimmune Encephalomyelitis (EAE)
2.1. Immune Cells in the Experimental Autoimmune Encephalomyelitis (EAE) Model
2.1.1. T Helper 1 (Th1) and T Helper (Th17) Cells
2.1.2. T Helper 9 (Th9) Cells
2.1.3. γδ T Cells
2.1.4. Regulatory T Cells (Tregs)
2.1.5. CD8+ T Cells
3. Mesenchymal Stem Cells (MSCs)
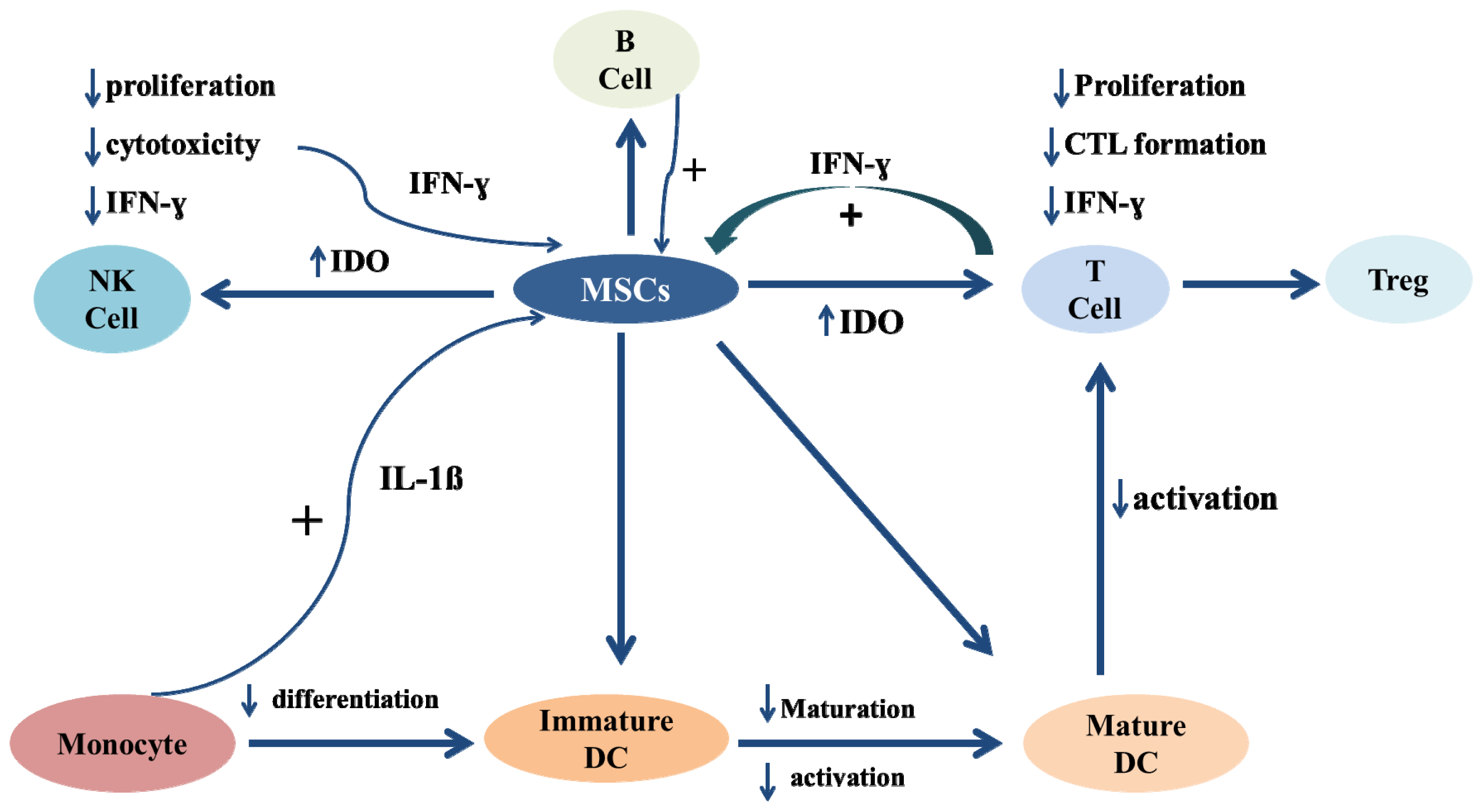
Immunosuppressive Characteristics of Mesenchymal Stem Cells
4. The Use of MSCs in Clinical Experimental Autoimmune Encephalomyelitis (EAE)
4.1. Immune Modulatory Effect of MSCs in EAE
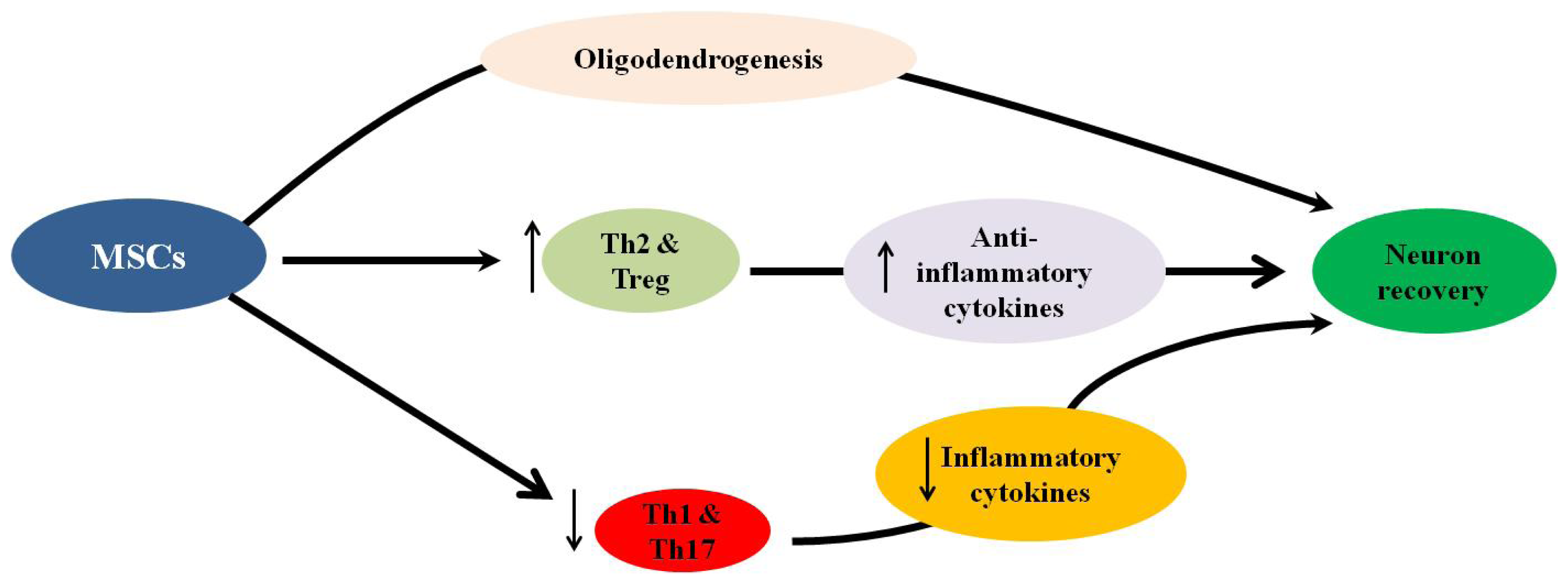
4.2. Neuroprotective Properties of MSCs
5. Conclusions
Acknowledgements
- Conflict of InterestThe authors have no conflicts of interest to declare.
References
- Noseworthy, J.H.; Lucchinetti, C.; Rodriguez, M.; Weinshenker, B.G. Multiple sclerosis. N. Engl. J. Med 2000, 343, 938–952. [Google Scholar]
- Frohman, E.M.; Racke, M.K.; Raine, C.S. Multiple sclerosis—The plaque and its pathogenesis. N. Engl. J. Med 2006, 354, 942–955. [Google Scholar]
- Martin, R. Immunological aspects of experimental allergic encephalomyelitis and multiple sclerosis and their application for new therapeutic strategies. J. Neural Transm. Suppl 1997, 49, 53–67. [Google Scholar]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar]
- Ebers, G.C. Environmental factors and multiple sclerosis. Lancet Neurol 2008, 7, 268–277. [Google Scholar]
- Duquette, P.; Pleines, J.; Girard, M.; Charest, L.; Senecal-Quevillon, M.; Masse, C. The increased susceptibility of women to multiple sclerosis. Can. J. Neurol. Sci 1992, 19, 466–471. [Google Scholar]
- McDonald, W.I.; Sears, T.A. The effects of experimental demyelination on conduction in the central nervous system. Brain 1970, 93, 583–598. [Google Scholar]
- Kuehn, B.M. Scientists probe strategies to repair neuron damage in multiple sclerosis. J. Am. Med. Assoc 2011, 305. [Google Scholar]
- Karussis, D.; Grigoriadis, S.; Polyzoidou, E.; Grigoriadis, N.; Slavin, S.; Abramsky, O. Neuroprotection in multiple sclerosis. Clin. Neurol. Neurosurg 2006, 108, 250–254. [Google Scholar]
- Steinman, L. Multiple sclerosis: A two-stage disease. Nat. Immunol 2001, 2, 762–764. [Google Scholar]
- Gehrmann, J.; Banati, R.B.; Cuzner, M.L.; Kreutzberg, G.W.; Newcombe, J. Amyloid precursor protein (APP) expression in multiple sclerosis lesions. Glia 1995, 15, 141–151. [Google Scholar]
- Foong, J.; Rozewicz, L.; Davie, C.A.; Thompson, A.J.; Miller, D.H.; Ron, M.A. Correlates of executive function in multiple sclerosis: The use of magnetic resonance spectroscopy as an index of focal pathology. J. Neuropsychiatry Clin. Neurosci 1999, 11, 45–50. [Google Scholar]
- Trapp, B.D.; Peterson, J.; Ransohoff, R.M.; Rudick, R.; Mork, S.; Bo, L. Axonal transection in the lesions of multiple sclerosis. N. Engl. J. Med 1998, 338, 278–285. [Google Scholar]
- Vladimirova, O.; O’Connor, J.; Cahill, A.; Alder, H.; Butunoi, C.; Kalman, B. Oxidative damage to DNA in plaques of MS brains. Mult. Scler 1998, 4, 413–418. [Google Scholar]
- Lovas, G.; Szilagyi, N.; Majtenyi, K.; Palkovits, M.; Komoly, S. Axonal changes in chronic demyelinated cervical spinal cord plaques. Brain 2000, 123, 308–317. [Google Scholar]
- Evangelou, N.; Esiri, M.M.; Smith, S.; Palace, J.; Matthews, P.M. Quantitative pathological evidence for axonal loss in normal appearing white matter in multiple sclerosis. Ann. Neurol 2000, 47, 391–395. [Google Scholar]
- Bai, L.; Lennon, D.P.; Eaton, V.; Maier, K.; Caplan, A.I.; Miller, S.D.; Miller, R.H. Human bone marrow-derived mesenchymal stem cells induce Th2-polarized immune response and promote endogenous repair in animal models of multiple sclerosis. Glia 2009, 57, 1192–1203. [Google Scholar]
- Constantin, G.; Marconi, S.; Rossi, B.; Angiari, S.; Calderan, L.; Anghileri, E.; Gini, B.; Bach, S.D.; Martinello, M.; Bifari, F.; et al. Adipose-derived mesenchymal stem cells ameliorate chronic experimental autoimmune encephalomyelitis. Stem. Cells 2009, 27, 2624–2635. [Google Scholar]
- Einstein, O.; Grigoriadis, N.; Mizrachi-Kol, R.; Reinhartz, E.; Polyzoidou, E.; Lavon, I.; Milonas, I.; Karussis, D.; Abramsky, O.; Ben-Hur, T. Transplanted neural precursor cells reduce brain inflammation to attenuate chronic experimental autoimmune encephalomyelitis. Exp. Neurol 2006, 198, 275–284. [Google Scholar]
- Gerdoni, E.; Gallo, B.; Casazza, S.; Musio, S.; Bonanni, I.; Pedemonte, E.; Mantegazza, R.; Frassoni, F.; Mancardi, G.; Pedotti, R.; et al. Mesenchymal stem cells effectively modulate pathogenic immune response in experimental autoimmune encephalomyelitis. Ann. Neurol 2007, 61, 219–227. [Google Scholar]
- Grigoriadis, N.; Lourbopoulos, A.; Lagoudaki, R.; Frischer, J.M.; Polyzoidou, E.; Touloumi, O.; Simeonidou, C.; Deretzi, G.; Kountouras, J.; Spandou, E.; et al. Variable behavior and complications of autologous bone marrow mesenchymal stem cells transplanted in experimental autoimmune encephalomyelitis. Exp. Neurol 2011, 230, 78–89. [Google Scholar]
- Karussis, D.; Kassis, I.; Kurkalli, B.G.; Slavin, S. Immunomodulation and neuroprotection with mesenchymal bone marrow stem cells (MSCs): A proposed treatment for multiple sclerosis and other neuroimmunological/neurodegenerative diseases. J. Neurol. Sci 2008, 265, 131–135. [Google Scholar]
- Kassis, I.; Grigoriadis, N.; Gowda-Kurkalli, B.; Mizrachi-Kol, R.; Ben-Hur, T.; Slavin, S.; Abramsky, O.; Karussis, D. Neuroprotection and immunomodulation with mesenchymal stem cells in chronic experimental autoimmune encephalomyelitis. Arch. Neurol 2008, 65, 753–761. [Google Scholar]
- Lu, Z.; Hu, X.; Zhu, C.; Wang, D.; Zheng, X.; Liu, Q. Overexpression of CNTF in Mesenchymal Stem Cells reduces demyelination and induces clinical recovery in experimental autoimmune encephalomyelitis mice. J. Neuroimmunol 2009, 206, 58–69. [Google Scholar]
- Matysiak, M.; Orlowski, W.; Fortak-Michalska, M.; Jurewicz, A.; Selmaj, K. Immunoregulatory function of bone marrow mesenchymal stem cells in EAE depends on their differentiation state and secretion of PGE2. J. Neuroimmunol 2011, 233, 106–111. [Google Scholar]
- Rafei, M.; Birman, E.; Forner, K.; Galipeau, J. Allogeneic mesenchymal stem cells for treatment of experimental autoimmune encephalomyelitis. Mol. Ther 2009, 17, 1799–1803. [Google Scholar]
- Rafei, M.; Campeau, P.M.; Aguilar-Mahecha, A.; Buchanan, M.; Williams, P.; Birman, E.; Yuan, S.; Young, Y.K.; Boivin, M.N.; Forner, K.; et al. Mesenchymal stromal cells ameliorate experimental autoimmune encephalomyelitis by inhibiting CD4 Th17 T cells in a CC chemokine ligand 2-dependent manner. J. Immunol 2009, 182, 5994–6002. [Google Scholar]
- Zappia, E.; Casazza, S.; Pedemonte, E.; Benvenuto, F.; Bonanni, I.; Gerdoni, E.; Giunti, D.; Ceravolo, A.; Cazzanti, F.; Frassoni, F.; et al. Mesenchymal stem cells ameliorate experimental autoimmune encephalomyelitis inducing T-cell anergy. Blood 2005, 106, 1755–1761. [Google Scholar]
- Stromnes, I.M.; Goverman, J.M. Active induction of experimental allergic encephalomyelitis. Nat. Protoc 2006, 1, 1810–1819. [Google Scholar]
- Furtado, G.C.; Marcondes, M.C.; Latkowski, J.A.; Tsai, J.; Wensky, A.; Lafaille, J.J. Swift entry of myelin-specific T lymphocytes into the central nervous system in spontaneous autoimmune encephalomyelitis. J. Immunol 2008, 181, 4648–4655. [Google Scholar]
- O’Connor, R.A.; Prendergast, C.T.; Sabatos, C.A.; Lau, C.W.; Leech, M.D.; Wraith, D.C.; Anderton, S.M. Cutting edge: Th1 cells facilitate the entry of Th17 cells to the central nervous system during experimental autoimmune encephalomyelitis. J. Immunol 2008, 181, 3750–3754. [Google Scholar]
- Steinman, L. Assessment of animal models for MS and demyelinating disease in the design of rational therapy. Neuron 1999, 24, 511–514. [Google Scholar]
- Stromnes, I.M.; Goverman, J.M. Passive induction of experimental allergic encephalomyelitis. Nat. Protoc 2006, 1, 1952–1960. [Google Scholar]
- Teitelbaum, D.; Meshorer, A.; Hirshfeld, T.; Arnon, R.; Sela, M. Suppression of experimental allergic encephalomyelitis by a synthetic polypeptide. Eur. J. Immunol 1971, 1, 242–248. [Google Scholar]
- Yednock, T.A.; Cannon, C.; Fritz, L.C.; Sanchez-Madrid, F.; Steinman, L.; Karin, N. Prevention of experimental autoimmune encephalomyelitis by antibodies against α4β1 integrin. Nature 1992, 356, 63–66. [Google Scholar]
- Sakaguchi, S.; Powrie, F. Emerging challenges in regulatory T cell function and biology. Science 2007, 317, 627–629. [Google Scholar]
- Lopez-Diego, R.S.; Weiner, H.L. Novel therapeutic strategies for multiple sclerosis—A multifaceted adversary. Nat. Rev. Drug Discov 2008, 7, 909–925. [Google Scholar]
- Viglietta, V.; Baecher-Allan, C.; Weiner, H.L.; Hafler, D.A. Loss of functional suppression by CD4+CD25+ regulatory T cells in patients with multiple sclerosis. J. Exp. Med 2004, 199, 971–979. [Google Scholar]
- Leussink, V.I.; Zettl, U.K.; Jander, S.; Pepinsky, R.B.; Lobb, R.R.; Stoll, G.; Toyka, K.V.; Gold, R. Blockade of signaling via the very late antigen (VLA-4) and its counterligand vascular cell adhesion molecule-1 (VCAM-1) causes increased T cell apoptosis in experimental autoimmune neuritis. Acta Neuropathol 2002, 103, 131–136. [Google Scholar]
- Jager, A.; Dardalhon, V.; Sobel, R.A.; Bettelli, E.; Kuchroo, V.K. Th1, Th17, and Th9 effector cells induce experimental autoimmune encephalomyelitis with different pathological phenotypes. J. Immunol 2009, 183, 7169–7177. [Google Scholar]
- Skulina, C.; Schmidt, S.; Dornmair, K.; Babbe, H.; Roers, A.; Rajewsky, K.; Wekerle, H.; Hohlfeld, R.; Goebels, N. Multiple sclerosis: Brain-infiltrating CD8+ T cells persist as clonal expansions in the cerebrospinal fluid and blood. Proc. Natl. Acad. Sci. USA 2004, 101, 2428–2433. [Google Scholar]
- Medana, I.; Martinic, M.A.; Wekerle, H.; Neumann, H. Transection of major histocompatibility complex class I-induced neurites by cytotoxic T lymphocytes. Am. J. Pathol 2001, 159, 809–815. [Google Scholar]
- Giuliani, F.; Goodyer, C.G.; Antel, J.P.; Yong, V.W. Vulnerability of human neurons to T cell-mediated cytotoxicity. J. Immunol 2003, 171, 368–379. [Google Scholar]
- Chitnis, T. The role of CD4 T cells in the pathogenesis of multiple sclerosis. Int. Rev. Neurobiol 2007, 79, 43–72. [Google Scholar]
- Hedegaard, C.J.; Krakauer, M.; Bendtzen, K.; Lund, H.; Sellebjerg, F.; Nielsen, C.H. T helper cell type 1 (Th1), Th2 and Th17 responses to myelin basic protein and disease activity in multiple sclerosis. Immunology 2008, 125, 161–169. [Google Scholar]
- Delgado, S.; Sheremata, W.A. The role of CD4+ T-cells in the development of MS. Neurol. Res 2006, 28, 245–249. [Google Scholar]
- Pettinelli, C.B.; McFarlin, D.E. Adoptive transfer of experimental allergic encephalomyelitis in SJL/J mice after in vitro activation of lymph node cells by myelin basic protein: Requirement for Lyt 1+ 2- T lymphocytes. J. Immunol 1981, 127, 1420–1423. [Google Scholar]
- McDonald, A.H.; Swanborg, R.H. Antigen-specific inhibition of immune interferon production by suppressor cells of autoimmune encephalomyelitis. J. Immunol 1988, 140, 1132–1138. [Google Scholar]
- Ando, D.G.; Clayton, J.; Kono, D.; Urban, J.L.; Sercarz, E.E. Encephalitogenic T cells in the B10.PL model of experimental allergic encephalomyelitis (EAE) are of the Th-1 lymphokine subtype. Cell Immunol 1989, 124, 132–143. [Google Scholar]
- Waldburger, K.E.; Hastings, R.C.; Schaub, R.G.; Goldman, S.J.; Leonard, J.P. Adoptive transfer of experimental allergic encephalomyelitis after in vitro treatment with recombinant murine interleukin-12. Preferential expansion of interferon-gamma-producing cells and increased expression of macrophage-associated inducible nitric oxide synthase as immunomodulatory mechanisms. Am. J. Pathol 1996, 148, 375–382. [Google Scholar]
- Yura, M.; Takahashi, I.; Serada, M.; Koshio, T.; Nakagami, K.; Yuki, Y.; Kiyono, H. Role of MOG-stimulated Th1 type “light up” (GFP+) CD4+ T cells for the development of experimental autoimmune encephalomyelitis (EAE). J. Autoimmun 2001, 17, 17–25. [Google Scholar]
- Lovett-Racke, A.E.; Rocchini, A.E.; Choy, J.; Northrop, S.C.; Hussain, R.Z.; Ratts, R.B.; Sikder, D.; Racke, M.K. Silencing T-bet defines a critical role in the differentiation of autoreactive T lymphocytes. Immunity 2004, 21, 719–731. [Google Scholar]
- Gocke, A.R.; Cravens, P.D.; Ben, L.H.; Hussain, R.Z.; Northrop, S.C.; Racke, M.K.; Lovett-Racke, A.E. T-bet regulates the fate of Th1 and Th17 lymphocytes in autoimmunity. J. Immunol 2007, 178, 1341–1348. [Google Scholar]
- Hemmer, B.; Cepok, S.; Zhou, D.; Sommer, N. Multiple sclerosis—A coordinated immune attack across the blood brain barrier. Curr. Neurovasc. Res 2004, 1, 141–150. [Google Scholar]
- Racke, M.K.; Bonomo, A.; Scott, D.E.; Cannella, B.; Levine, A.; Raine, C.S.; Shevach, E.M.; Rocken, M. Cytokine-induced immune deviation as a therapy for inflammatory autoimmune disease. J. Exp. Med 1994, 180, 1961–1966. [Google Scholar]
- Racke, M.K.; Burnett, D.; Pak, S.H.; Albert, P.S.; Cannella, B.; Raine, C.S.; McFarlin, D.E.; Scott, D.E. Retinoid treatment of experimental allergic encephalomyelitis. IL-4 production correlates with improved disease course. J. Immunol 1995, 154, 450–458. [Google Scholar]
- Willenborg, D.O.; Fordham, S.; Bernard, C.C.; Cowden, W.B.; Ramshaw, I.A. IFN-gamma plays a critical down-regulatory role in the induction and effector phase of myelin oligodendrocyte glycoprotein-induced autoimmune encephalomyelitis. J. Immunol 1996, 157, 3223–3227. [Google Scholar]
- Lublin, F.D.; Knobler, R.L.; Kalman, B.; Goldhaber, M.; Marini, J.; Perrault, M.; D’Imperio, C.; Joseph, J.; Alkan, S.S.; Korngold, R. Monoclonal anti-gamma interferon antibodies enhance experimental allergic encephalomyelitis. Autoimmunity 1993, 16, 267–274. [Google Scholar]
- Ferber, I.A.; Brocke, S.; Taylor-Edwards, C.; Ridgway, W.; Dinisco, C.; Steinman, L.; Dalton, D.; Fathman, C.G. Mice with a disrupted IFN-gamma gene are susceptible to the induction of experimental autoimmune encephalomyelitis (EAE). J. Immunol 1996, 156, 5–7. [Google Scholar]
- Heremans, H.; Dillen, C.; Groenen, M.; Martens, E.; Billiau, A. Chronic relapsing experimental autoimmune encephalomyelitis (CREAE) in mice: Enhancement by monoclonal antibodies against interferon-γ. Eur. J. Immunol 1996, 26, 2393–2398. [Google Scholar]
- Chu, C.Q.; Wittmer, S.; Dalton, D.K. Failure to suppress the expansion of the activated CD4 T cell population in interferon γ-deficient mice leads to exacerbation of experimental autoimmune encephalomyelitis. J. Exp. Med 2000, 192, 123–128. [Google Scholar]
- Becher, B.; Durell, B.G.; Noelle, R.J. Experimental autoimmune encephalitis and inflammation in the absence of interleukin-12. J. Clin. Invest 2002, 110, 493–497. [Google Scholar]
- Gran, B.; Zhang, G.X.; Yu, S.; Li, J.; Chen, X.H.; Ventura, E.S.; Kamoun, M.; Rostami, A. IL-12p35-deficient mice are susceptible to experimental autoimmune encephalomyelitis: Evidence for redundancy in the IL-12 system in the induction of central nervous system autoimmune demyelination. J. Immunol 2002, 169, 7104–7110. [Google Scholar]
- Cua, D.J.; Sherlock, J.; Chen, Y.; Murphy, C.A.; Joyce, B.; Seymour, B.; Lucian, L.; To, W.; Kwan, S.; Churakova, T.; et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 2003, 421, 744–748. [Google Scholar]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med 2005, 201, 233–240. [Google Scholar]
- Mangan, P.R.; Harrington, L.E.; O’Quinn, D.B.; Helms, W.S.; Bullard, D.C.; Elson, C.O.; Hatton, R.D.; Wahl, S.M.; Schoeb, T.R.; Weaver, C.T. Transforming growth factor-β induces development of the T(H)17 lineage. Nature 2006, 441, 231–234. [Google Scholar]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol 2005, 6, 1123–1132. [Google Scholar]
- Durelli, L.; Conti, L.; Clerico, M.; Boselli, D.; Contessa, G.; Ripellino, P.; Ferrero, B.; Eid, P.; Novelli, F. T-helper 17 cells expand in multiple sclerosis and are inhibited by interferon-β. Ann. Neurol 2009, 65, 499–509. [Google Scholar]
- Tzartos, J.S.; Friese, M.A.; Craner, M.J.; Palace, J.; Newcombe, J.; Esiri, M.M.; Fugger, L. Interleukin-17 production in central nervous system-infiltrating T cells and glial cells is associated with active disease in multiple sclerosis. Am. J. Pathol 2008, 172, 146–155. [Google Scholar]
- Kebir, H.; Kreymborg, K.; Ifergan, I.; Dodelet-Devillers, A.; Cayrol, R.; Bernard, M.; Giuliani, F.; Arbour, N.; Becher, B.; Prat, A. Human TH17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation. Nat. Med 2007, 13, 1173–1175. [Google Scholar]
- Ransohoff, R.M. Immunology: In the beginning. Nature 2009, 462, 41–42. [Google Scholar]
- Tesmer, L.A.; Lundy, S.K.; Sarkar, S.; Fox, D.A. Th17 cells in human disease. Immunol. Rev 2008, 223, 87–113. [Google Scholar]
- Hofstetter, H.H.; Ibrahim, S.M.; Koczan, D.; Kruse, N.; Weishaupt, A.; Toyka, K.V.; Gold, R. Therapeutic efficacy of IL-17 neutralization in murine experimental autoimmune encephalomyelitis. Cell Immunol 2005, 237, 123–130. [Google Scholar]
- Komiyama, Y.; Nakae, S.; Matsuki, T.; Nambu, A.; Ishigame, H.; Kakuta, S.; Sudo, K.; Iwakura, Y. IL-17 plays an important role in the development of experimental autoimmune encephalomyelitis. J. Immunol 2006, 177, 566–573. [Google Scholar]
- Haak, S.; Croxford, A.L.; Kreymborg, K.; Heppner, F.L.; Pouly, S.; Becher, B.; Waisman, A. IL-17A and IL-17F do not contribute vitally to autoimmune neuro-inflammation in mice. J. Clin. Invest 2009, 119, 61–69. [Google Scholar]
- Nowak, E.C.; Weaver, C.T.; Turner, H.; Begum-Haque, S.; Becher, B.; Schreiner, B.; Coyle, A.J.; Kasper, L.H.; Noelle, R.J. IL-9 as a mediator of Th17-driven inflammatory disease. J. Exp. Med 2009, 206, 1653–1660. [Google Scholar] [Green Version]
- Sutton, C.; Brereton, C.; Keogh, B.; Mills, K.H.; Lavelle, E.C. A crucial role for interleukin (IL)-1 in the induction of IL-17-producing T cells that mediate autoimmune encephalomyelitis. J. Exp. Med 2006, 203, 1685–1691. [Google Scholar]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-β-induced Foxp3+ T cells and, together with TGF-β, generates IL-9+ IL-10+ Foxp3(−) effector T cells. Nat. Immunol 2008, 9, 1347–1355. [Google Scholar]
- Veldhoen, M.; Uyttenhove, C.; van Snick, J.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-β ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol 2008, 9, 1341–1346. [Google Scholar]
- Soroosh, P.; Doherty, T.A. Th9 and allergic disease. Immunology 2009, 127, 450–458. [Google Scholar]
- Wucherpfennig, K.W.; Newcombe, J.; Li, H.; Keddy, C.; Cuzner, M.L.; Hafler, D.A. Gamma delta T-cell receptor repertoire in acute multiple sclerosis lesions. Proc. Natl. Acad. Sci. USA 1992, 89, 4588–4592. [Google Scholar]
- Shimonkevitz, R.; Colburn, C.; Burnham, J.A.; Murray, R.S.; Kotzin, B.L. Clonal expansions of activated gamma/delta T cells in recent-onset multiple sclerosis. Proc. Natl. Acad. Sci. USA 1993, 90, 923–927. [Google Scholar]
- Lockhart, E.; Green, A.M.; Flynn, J.L. IL-17 production is dominated by γδ T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J. Immunol 2006, 177, 4662–4669. [Google Scholar]
- Shibata, K.; Yamada, H.; Hara, H.; Kishihara, K.; Yoshikai, Y. Resident Vδ1+ γδ T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. J. Immunol 2007, 178, 4466–4472. [Google Scholar]
- Sutton, C.E.; Lalor, S.J.; Sweeney, C.M.; Brereton, C.F.; Lavelle, E.C.; Mills, K.H. Interleukin-1 and IL-23 induce innate IL-17 production from γδ T cells, amplifying Th17 responses and autoimmunity. Immunity 2009, 31, 331–341. [Google Scholar]
- Kobayashi, Y.; Kawai, K.; Ito, K.; Honda, H.; Sobue, G.; Yoshikai, Y. Aggravation of murine experimental allergic encephalomyelitis by administration of T-cell receptor γδ-specific antibody. J. Neuroimmunol 1997, 73, 169–174. [Google Scholar]
- Ponomarev, E.D.; Dittel, B.N. γδ T cells regulate the extent and duration of inflammation in the central nervous system by a Fas ligand-dependent mechanism. J. Immunol 2005, 174, 4678–4687. [Google Scholar]
- Ponomarev, E.D.; Novikova, M.; Yassai, M.; Szczepanik, M.; Gorski, J.; Dittel, B.N. γδ T cell regulation of IFN-γ production by central nervous system-infiltrating encephalitogenic T cells: Correlation with recovery from experimental autoimmune encephalomyelitis. J. Immunol 2004, 173, 1587–1595. [Google Scholar]
- Rajan, A.J.; Gao, Y.L.; Raine, C.S.; Brosnan, C.F. A pathogenic role for gamma delta T cells in relapsing-remitting experimental allergic encephalomyelitis in the SJL mouse. J. Immunol 1996, 157, 941–949. [Google Scholar]
- Rajan, A.J.; Klein, J.D.; Brosnan, C.F. The effect of γδ T cell depletion on cytokine gene expression in experimental allergic encephalomyelitis. J. Immunol 1998, 160, 5955–5962. [Google Scholar]
- Spahn, T.W.; Issazadah, S.; Salvin, A.J.; Weiner, H.L. Decreased severity of myelin oligodendrocyte glycoprotein peptide 33–35-induced experimental autoimmune encephalomyelitis in mice with a disrupted TCR δ chain gene. Eur. J. Immunol 1999, 29, 4060–4071. [Google Scholar]
- Odyniec, A.; Szczepanik, M.; Mycko, M.P.; Stasiolek, M.; Raine, C.S.; Selmaj, K.W. γδ T cells enhance the expression of experimental autoimmune encephalomyelitis by promoting antigen presentation and IL-12 production. J. Immunol 2004, 173, 682–694. [Google Scholar]
- Cardona, A.E.; Teale, J.M. γ/δ T cell-deficient mice exhibit reduced disease severity and decreased inflammatory response in the brain in murine neurocysticercosis. J. Immunol 2002, 169, 3163–3171. [Google Scholar]
- Hori, S.; Sakaguchi, S. Foxp3: A critical regulator of the development and function of regulatory T cells. Microbes Infect 2004, 6, 745–751. [Google Scholar]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol 2003, 4, 330–336. [Google Scholar]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol 1995, 155, 1151–1164. [Google Scholar]
- Roncarolo, M.G.; Gregori, S.; Battaglia, M.; Bacchetta, R.; Fleischhauer, K.; Levings, M.K. Interleukin-10-secreting type 1 regulatory T cells in rodents and humans. Immunol. Rev 2006, 212, 28–50. [Google Scholar]
- Martinez-Forero, I.; Garcia-Munoz, R.; Martinez-Pasamar, S.; Inoges, S.; Lopez-Diaz de Cerio, A.; Palacios, R.; Sepulcre, J.; Moreno, B.; Gonzalez, Z.; Fernandez-Diez, B.; et al. IL-10 suppressor activity and ex vivo Tr1 cell function are impaired in multiple sclerosis. Eur. J. Immunol 2008, 38, 576–586. [Google Scholar]
- Astier, A.L.; Meiffren, G.; Freeman, S.; Hafler, D.A. Alterations in CD46-mediated Tr1 regulatory T cells in patients with multiple sclerosis. J. Clin. Invest 2006, 116, 3252–3257. [Google Scholar]
- Venken, K.; Hellings, N.; Thewissen, M.; Somers, V.; Hensen, K.; Rummens, J.L.; Medaer, R.; Hupperts, R.; Stinissen, P. Compromised CD4+ CD25(high) regulatory T-cell function in patients with relapsing-remitting multiple sclerosis is correlated with a reduced frequency of FOXP3-positive cells and reduced FOXP3 expression at the single-cell level. Immunology 2008, 123, 79–89. [Google Scholar]
- Venken, K.; Hellings, N.; Hensen, K.; Rummens, J.L.; Medaer, R.; D’Hooghe, M.B.; Dubois, B.; Raus, J.; Stinissen, P. Secondary progressive in contrast to relapsing-remitting multiple sclerosis patients show a normal CD4+CD25+ regulatory T-cell function and FOXP3 expression. J. Neurosci. Res 2006, 83, 1432–1446. [Google Scholar]
- Lafaille, J.J.; Nagashima, K.; Katsuki, M.; Tonegawa, S. High incidence of spontaneous autoimmune encephalomyelitis in immunodeficient anti-myelin basic protein T cell receptor transgenic mice. Cell 1994, 78, 399–408. [Google Scholar]
- Olivares-Villagomez, D.; Wang, Y.; Lafaille, J.J. Regulatory CD4(+) T cells expressing endogenous T cell receptor chains protect myelin basic protein-specific transgenic mice from spontaneous autoimmune encephalomyelitis. J. Exp. Med 1998, 188, 1883–1894. [Google Scholar]
- Kohm, A.P.; Carpentier, P.A.; Anger, H.A.; Miller, S.D. Cutting edge: CD4+CD25+ regulatory T cells suppress antigen-specific autoreactive immune responses and central nervous system inflammation during active experimental autoimmune encephalomyelitis. J. Immunol 2002, 169, 4712–4716. [Google Scholar]
- Friese, M.A.; Fugger, L. Pathogenic CD8(+) T cells in multiple sclerosis. Ann. Neurol 2009, 66, 132–141. [Google Scholar]
- Weiss, H.A.; Millward, J.M.; Owens, T. CD8+ T cells in inflammatory demyelinating disease. J. Neuroimmunol 2007, 191, 79–85. [Google Scholar]
- Najafian, N.; Chitnis, T.; Salama, A.D.; Zhu, B.; Benou, C.; Yuan, X.; Clarkson, M.R.; Sayegh, M.H.; Khoury, S.J. Regulatory functions of CD8+CD28- T cells in an autoimmune disease model. J. Clin. Invest 2003, 112, 1037–1048. [Google Scholar]
- Lee, Y.H.; Ishida, Y.; Rifa’i, M.; Shi, Z.; Isobe, K.; Suzuki, H. Essential role of CD8+CD122+ regulatory T cells in the recovery from experimental autoimmune encephalomyelitis. J. Immunol 2008, 180, 825–832. [Google Scholar]
- Liu, Z.; Tugulea, S.; Cortesini, R.; Suciu-Foca, N. Specific suppression of T helper alloreactivity by allo-MHC class I-restricted CD8+CD28- T cells. Int. Immunol 1998, 10, 775–783. [Google Scholar]
- Ciubotariu, R.; Colovai, A.I.; Pennesi, G.; Liu, Z.; Smith, D.; Berlocco, P.; Cortesini, R.; Suciu-Foca, N. Specific suppression of human CD4+ Th cell responses to pig MHC antigens by CD8+CD28- regulatory T cells. J. Immunol 1998, 161, 5193–5202. [Google Scholar]
- Jiang, S.; Tugulea, S.; Pennesi, G.; Liu, Z.; Mulder, A.; Lederman, S.; Harris, P.; Cortesini, R.; Suciu-Foca, N. Induction of MHC-class I restricted human suppressor T cells by peptide priming in vitro. Hum. Immunol 1998, 59, 690–699. [Google Scholar]
- Li, J.; Liu, Z.; Jiang, S.; Cortesini, R.; Lederman, S.; Suciu-Foca, N. T suppressor lymphocytes inhibit NF-κB-mediated transcription of CD86 gene in APC. J. Immunol 1999, 163, 6386–6392. [Google Scholar]
- Colovai, A.I.; Liu, Z.; Ciubotariu, R.; Lederman, S.; Cortesini, R.; Suciu-Foca, N. Induction of xenoreactive CD4+ T-cell anergy by suppressor CD8+CD28- T cells. Transplantation 2000, 69, 1304–1310. [Google Scholar]
- Rifa’i, M.; Kawamoto, Y.; Nakashima, I.; Suzuki, H. Essential roles of CD8+CD122+ regulatory T cells in the maintenance of T cell homeostasis. J. Exp. Med 2004, 200, 1123–1134. [Google Scholar]
- Endharti, A.T.; Rifa, I.M.; Shi, Z.; Fukuoka, Y.; Nakahara, Y.; Kawamoto, Y.; Takeda, K.; Isobe, K.; Suzuki, H. Cutting edge: CD8+CD122+ regulatory T cells produce IL-10 to suppress IFN-γ production and proliferation of CD8+ T cells. J. Immunol 2005, 175, 7093–7097. [Google Scholar]
- Linker, R.A.; Rott, E.; Hofstetter, H.H.; Hanke, T.; Toyka, K.V.; Gold, R. EAE in beta-2 microglobulin-deficient mice: Axonal damage is not dependent on MHC-I restricted immune responses. Neurobiol. Dis 2005, 19, 218–228. [Google Scholar]
- Jiang, H.; Ware, R.; Stall, A.; Flaherty, L.; Chess, L.; Pernis, B. Murine CD8+ T cells that specifically delete autologous CD4+ T cells expressing Vβ8 TCR: A role of the Qa-1 molecule. Immunity 1995, 2, 185–194. [Google Scholar]
- Jiang, H.; Braunstein, N.S.; Yu, B.; Winchester, R.; Chess, L. CD8+ T cells control the TH phenotype of MBP-reactive CD4+ T cells in EAE mice. Proc. Natl. Acad. Sci. USA 2001, 98, 6301–6306. [Google Scholar]
- Jiang, H.; Curran, S.; Ruiz-Vazquez, E.; Liang, B.; Winchester, R.; Chess, L. Regulatory CD8+ T cells fine-tune the myelin basic protein-reactive T cell receptor Vβ repertoire during experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2003, 100, 8378–8383. [Google Scholar]
- Hu, D.; Ikizawa, K.; Lu, L.; Sanchirico, M.E.; Shinohara, M.L.; Cantor, H. Analysis of regulatory CD8 T cells in Qa-1-deficient mice. Nat. Immunol 2004, 5, 516–523. [Google Scholar]
- Chen, M.L.; Yan, B.S.; Kozoriz, D.; Weiner, H.L. Novel CD8+ Treg suppress EAE by TGF-β- and IFN-γ-dependent mechanisms. Eur. J. Immunol 2009, 39, 3423–3435. [Google Scholar]
- Lucchinetti, C.; Bruck, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of multiple sclerosis lesions: Implications for the pathogenesis of demyelination. Ann. Neurol 2000, 47, 707–717. [Google Scholar]
- Breij, E.C.; Brink, B.P.; Veerhuis, R.; van den Berg, C.; Vloet, R.; Yan, R.; Dijkstra, C.D.; van der Valk, P.; Bo, L. Homogeneity of active demyelinating lesions in established multiple sclerosis. Ann. Neurol 2008, 63, 16–25. [Google Scholar]
- Reindl, M.; Linington, C.; Brehm, U.; Egg, R.; Dilitz, E.; Deisenhammer, F.; Poewe, W.; Berger, T. Antibodies against the myelin oligodendrocyte glycoprotein and the myelin basic protein in multiple sclerosis and other neurological diseases: A comparative study. Brain 1999, 122, 2047–2056. [Google Scholar]
- Van der Goes, A.; Kortekaas, M.; Hoekstra, K.; Dijkstra, C.D.; Amor, S. The role of anti-myelin (auto)-antibodies in the phagocytosis of myelin by macrophages. J. Neuroimmunol 1999, 101, 61–67. [Google Scholar]
- Menon, K.K.; Piddlesden, S.J.; Bernard, C.C. Demyelinating antibodies to myelin oligodendrocyte glycoprotein and galactocerebroside induce degradation of myelin basic protein in isolated human myelin. J. Neurochem 1997, 69, 214–222. [Google Scholar]
- Link, H.; Baig, S.; Jiang, Y.P.; Olsson, O.; Hojeberg, B.; Kostulas, V.; Olsson, T. B cells and antibodies in MS. Res. Immunol 1989, 140, 219–226, Discussion 245–218. [Google Scholar]
- Lalive, P.H. Autoantibodies in inflammatory demyelinating diseases of the central nervous system. Swiss Med. Wkly 2008, 138, 692–707. [Google Scholar]
- Vyshkina, T.; Kalman, B. Autoantibodies and neurodegeneration in multiple sclerosis. Lab. Invest 2008, 88, 796–807. [Google Scholar]
- Hauser, S.L.; Waubant, E.; Arnold, D.L.; Vollmer, T.; Antel, J.; Fox, R.J.; Bar-Or, A.; Panzara, M.; Sarkar, N.; Agarwal, S.; et al. B-cell depletion with rituximab in relapsing-remitting multiple sclerosis. N. Engl. J. Med 2008, 358, 676–688. [Google Scholar]
- Harp, C.T.; Lovett-Racke, A.E.; Racke, M.K.; Frohman, E.M.; Monson, N.L. Impact of myelin-specific antigen presenting B cells on T cell activation in multiple sclerosis. Clin. Immunol 2008, 128, 382–391. [Google Scholar]
- Correale, J.; Farez, M. Helminth antigens modulate immune responses in cells from multiple sclerosis patients through TLR2-dependent mechanisms. J. Immunol 2009, 183, 5999–6012. [Google Scholar]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar]
- Zozulya, A.L.; Clarkson, B.D.; Ortler, S.; Fabry, Z.; Wiendl, H. The role of dendritic cells in CNS autoimmunity. J. Mol. Med. (Berl) 2010, 88, 535–544. [Google Scholar]
- Menges, M.; Rossner, S.; Voigtlander, C.; Schindler, H.; Kukutsch, N.A.; Bogdan, C.; Erb, K.; Schuler, G.; Lutz, M.B. Repetitive injections of dendritic cells matured with tumor necrosis factor α induce antigen-specific protection of mice from autoimmunity. J. Exp. Med 2002, 195, 15–21. [Google Scholar]
- Serafini, B.; Rosicarelli, B.; Magliozzi, R.; Stigliano, E.; Capello, E.; Mancardi, G.L.; Aloisi, F. Dendritic cells in multiple sclerosis lesions: Maturation stage, myelin uptake, and interaction with proliferating T cells. J. Neuropathol. Exp. Neurol 2006, 65, 124–141. [Google Scholar]
- Karni, A.; Abraham, M.; Monsonego, A.; Cai, G.; Freeman, G.J.; Hafler, D.; Khoury, S.J.; Weiner, H.L. Innate immunity in multiple sclerosis: Myeloid dendritic cells in secondary progressive multiple sclerosis are activated and drive a proinflammatory immune response. J. Immunol 2006, 177, 4196–4202. [Google Scholar]
- Fabriek, B.O.; van Haastert, E.S.; Galea, I.; Polfliet, M.M.; Dopp, E.D.; van Den Heuvel, M.M.; van Den Berg, T.K.; de Groot, C.J.; van Der Valk, P.; Dijkstra, C.D. CD163-positive perivascular macrophages in the human CNS express molecules for antigen recognition and presentation. Glia 2005, 51, 297–305. [Google Scholar]
- Van Zwam, M.; Huizinga, R.; Melief, M.J.; Wierenga-Wolf, A.F.; van Meurs, M.; Voerman, J.S.; Biber, K.P.; Boddeke, H.W.; Hopken, U.E.; Meisel, C.; et al. Brain antigens in functionally distinct antigen-presenting cell populations in cervical lymph nodes in MS and EAE. J. Mol. Med. (Berl) 2009, 87, 273–286. [Google Scholar]
- Reubinoff, B.E.; Pera, M.F.; Fong, C.Y.; Trounson, A.; Bongso, A. Embryonic stem cell lines from human blastocysts: Somatic differentiation in vitro. Nat. Biotechnol 2000, 18, 399–404. [Google Scholar]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar]
- Kehat, I.; Kenyagin-Karsenti, D.; Snir, M.; Segev, H.; Amit, M.; Gepstein, A.; Livne, E.; Binah, O.; Itskovitz-Eldor, J.; Gepstein, L. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J. Clin. Invest 2001, 108, 407–414. [Google Scholar]
- Lavon, N.; Benvenisty, N. Differentiation and genetic manipulation of human embryonic stem cells and the analysis of the cardiovascular system. Trends Cardiovasc. Med 2003, 13, 47–52. [Google Scholar]
- Augello, A.; Kurth, T.B.; de Bari, C. Mesenchymal stem cells: A perspective from in vitro cultures to in vivo migration and niches. Eur. Cell Mater 2010, 20, 121–133. [Google Scholar]
- Krampera, M.; Pizzolo, G.; Aprili, G.; Franchini, M. Mesenchymal stem cells for bone, cartilage, tendon and skeletal muscle repair. Bone 2006, 39, 678–683. [Google Scholar]
- Li, C.; Zhang, W.; Jiang, X.; Mao, N. Human-placenta-derived mesenchymal stem cells inhibit proliferation and function of allogeneic immune cells. Cell Tissue Res 2007, 330, 437–446. [Google Scholar]
- Zhang, X.; Mitsuru, A.; Igura, K.; Takahashi, K.; Ichinose, S.; Yamaguchi, S.; Takahashi, T.A. Mesenchymal progenitor cells derived from chorionic villi of human placenta for cartilage tissue engineering. Biochem. Biophys. Res. Commun 2006, 340, 944–952. [Google Scholar]
- Chang, C.J.; Yen, M.L.; Chen, Y.C.; Chien, C.C.; Huang, H.I.; Bai, C.H.; Yen, B.L. Placenta-derived multipotent cells exhibit immunosuppressive properties that are enhanced in the presence of interferon-γ. Stem. Cells 2006, 24, 2466–2477. [Google Scholar]
- Jones, B.J.; Brooke, G.; Atkinson, K.; McTaggart, S.J. Immunosuppression by placental indoleamine 2,3-dioxygenase: A role for mesenchymal stem cells. Placenta 2007, 28, 1174–1181. [Google Scholar]
- Brooke, G.; Tong, H.; Levesque, J.P.; Atkinson, K. Molecular trafficking mechanisms of multipotent mesenchymal stem cells derived from human bone marrow and placenta. Stem. Cells Dev 2008, 17, 929–940. [Google Scholar]
- Lee, M.Y.; Huang, J.P.; Chen, Y.Y.; Aplin, J.D.; Wu, Y.H.; Chen, C.Y.; Chen, P.C.; Chen, C.P. Angiogenesis in differentiated placental multipotent mesenchymal stromal cells is dependent on integrin α5β1. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Chen, L.; He, D.M.; Zhang, Y. The differentiation of human placenta-derived mesenchymal stem cells into dopaminergic cells in vitro. Cell Mol. Biol. Lett 2009, 14, 528–536. [Google Scholar]
- Li, G.; Zhang, X.A.; Wang, H.; Wang, X.; Meng, C.L.; Chan, C.Y.; Yew, D.T.; Tsang, K.S.; Li, K.; Tsai, S.N.; et al. Comparative proteomic analysis of mesenchymal stem cells derived from human bone marrow, umbilical cord, and placenta: Implication in the migration. Proteomics 2009, 9, 20–30. [Google Scholar]
- Hiwase, S.D.; Dyson, P.G.; To, L.B.; Lewis, I.D. Cotransplantation of placental mesenchymal stromal cells enhances single and double cord blood engraftment in nonobese diabetic/severe combined immune deficient mice. Stem. Cells 2009, 27, 2293–2300. [Google Scholar]
- Semenov, O.V.; Koestenbauer, S.; Riegel, M.; Zech, N.; Zimmermann, R.; Zisch, A.H.; Malek, A. Multipotent mesenchymal stem cells from human placenta: Critical parameters for isolation and maintenance of stemness after isolation. Am. J. Obstet. Gynecol 2010, 202, 193.e1–193.e13. [Google Scholar]
- Portmann-Lanz, C.B.; Schoeberlein, A.; Huber, A.; Sager, R.; Malek, A.; Holzgreve, W.; Surbek, D.V. Placental mesenchymal stem cells as potential autologous graft for pre- and perinatal neuroregeneration. Am. J. Obstet. Gynecol 2006, 194, 664–673. [Google Scholar]
- Hwang, J.H.; Shim, S.S.; Seok, O.S.; Lee, H.Y.; Woo, S.K.; Kim, B.H.; Song, H.R.; Lee, J.K.; Park, Y.K. Comparison of cytokine expression in mesenchymal stem cells from human placenta, cord blood, and bone marrow. J. Korean Med. Sci 2009, 24, 547–554. [Google Scholar]
- Ilancheran, S.; Michalska, A.; Peh, G.; Wallace, E.M.; Pera, M.; Manuelpillai, U. Stem cells derived from human fetal membranes display multilineage differentiation potential. Biol. Reprod 2007, 77, 577–588. [Google Scholar]
- Deuse, T.; Stubbendorff, M.; Tang-Quan, K.; Phillips, N.; Kay, M.A.; Eiermann, T.; Phan, T.T.; Volk, H.D.; Reichenspurner, H.; Robbins, R.C.; et al. Immunogenicity and immunomodulatory properties of umbilical cord lining mesenchymal stem cells. Cell Transplant 2011, 20, 655–667. [Google Scholar]
- In’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 2003, 102, 1548–1549. [Google Scholar]
- Tomic, S.; Djokic, J.; Vasilijic, S.; Vucevic, D.; Todorovic, V.; Supic, G.; Colic, M. Immunomodulatory properties of mesenchymal stem cells derived from dental pulp and dental follicle are susceptible to activation by toll-like receptor agonists. Stem. Cells Dev 2011, 20, 695–708. [Google Scholar]
- Bartholomew, A.; Sturgeon, C.; Siatskas, M.; Ferrer, K.; McIntosh, K.; Patil, S.; Hardy, W.; Devine, S.; Ucker, D.; Deans, R.; et al. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp. Hematol 2002, 30, 42–48. [Google Scholar]
- Djouad, F.; Bony, C.; Haupl, T.; Uze, G.; Lahlou, N.; Louis-Plence, P.; Apparailly, F.; Canovas, F.; Reme, T.; Sany, J.; et al. Transcriptional profiles discriminate bone marrow-derived and synovium-derived mesenchymal stem cells. Arthritis Res. Ther 2005, 7, R1304–R1315. [Google Scholar]
- Tse, W.T.; Pendleton, J.D.; Beyer, W.M.; Egalka, M.C.; Guinan, E.C. Suppression of allogeneic T-cell proliferation by human marrow stromal cells: Implications in transplantation. Transplantation 2003, 75, 389–397. [Google Scholar]
- Bocelli-Tyndall, C.; Bracci, L.; Schaeren, S.; Feder-Mengus, C.; Barbero, A.; Tyndall, A.; Spagnoli, G.C. Human bone marrow mesenchymal stem cells and chondrocytes promote and/or suppress the in vitro proliferation of lymphocytes stimulated by interleukins 2, 7 and 15. Ann. Rheum. Dis 2009, 68, 1352–1359. [Google Scholar]
- Schurgers, E.; Kelchtermans, H.; Mitera, T.; Geboes, L.; Matthys, P. Discrepancy between the in vitro and in vivo effects of murine mesenchymal stem cells on T-cell proliferation and collagen-induced arthritis. Arthritis Res. Ther 2010, 12. [Google Scholar] [CrossRef]
- Chen, C.P.; Liu, S.H.; Huang, J.P.; Aplin, J.D.; Wu, Y.H.; Chen, P.C.; Hu, C.S.; Ko, C.C.; Lee, M.Y.; Chen, C.Y. Engraftment potential of human placenta-derived mesenchymal stem cells after in utero transplantation in rats. Hum. Reprod 2009, 24, 154–165. [Google Scholar]
- Chang, Y.J.; Hwang, S.M.; Tseng, C.P.; Cheng, F.C.; Huang, S.H.; Hsu, L.F.; Hsu, L.W.; Tsai, M.S. Isolation of mesenchymal stem cells with neurogenic potential from the mesoderm of the amniotic membrane. Cells Tissues Organs 2010, 192, 93–105. [Google Scholar]
- Bailo, M.; Soncini, M.; Vertua, E.; Signoroni, P.B.; Sanzone, S.; Lombardi, G.; Arienti, D.; Calamani, F.; Zatti, D.; Paul, P.; et al. Engraftment potential of human amnion and chorion cells derived from term placenta. Transplantation 2004, 78, 1439–1448. [Google Scholar]
- Wolbank, S.; Peterbauer, A.; Fahrner, M.; Hennerbichler, S.; van Griensven, M.; Stadler, G.; Redl, H.; Gabriel, C. Dose-dependent immunomodulatory effect of human stem cells from amniotic membrane: A comparison with human mesenchymal stem cells from adipose tissue. Tissue Eng 2007, 13, 1173–1183. [Google Scholar]
- Gotherstrom, C.; Ringden, O.; Westgren, M.; Tammik, C.; Le Blanc, K. Immunomodulatory effects of human foetal liver-derived mesenchymal stem cells. Bone Marrow Transplant 2003, 32, 265–272. [Google Scholar]
- Giuliani, M.; Fleury, M.; Vernochet, A.; Ketroussi, F.; Clay, D.; Azzarone, B.; Lataillade, J.J.; Durrbach, A. Long-lasting inhibitory effects of fetal liver mesenchymal stem cells on T-lymphocyte proliferation. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Zheng, Z.H.; Li, X.Y.; Ding, J.; Jia, J.F.; Zhu, P. Allogeneic mesenchymal stem cell and mesenchymal stem cell-differentiated chondrocyte suppress the responses of type II collagen-reactive T cells in rheumatoid arthritis. Rheumatology (Oxford) 2008, 47, 22–30. [Google Scholar]
- Petrini, I.; Pacini, S.; Petrini, M.; Fazzi, R.; Trombi, L.; Galimberti, S. Mesenchymal cells inhibit expansion but not cytotoxicity exerted by gamma-delta T cells. Eur. J. Clin. Invest 2009, 39, 813–818. [Google Scholar]
- Augello, A.; Tasso, R.; Negrini, S.M.; Amateis, A.; Indiveri, F.; Cancedda, R.; Pennesi, G. Bone marrow mesenchymal progenitor cells inhibit lymphocyte proliferation by activation of the programmed death 1 pathway. Eur. Immunol 2005, 35, 1482–1490. [Google Scholar]
- Xue, Q.; Luan, X.Y.; Gu, Y.Z.; Wu, H.Y.; Zhang, G.B.; Yu, G.H.; Zhu, H.T.; Wang, M.; Dong, W.; Geng, Y.J.; et al. The negative co-signaling molecule b7-h4 is expressed by human bone marrow-derived mesenchymal stem cells and mediates its T-cell modulatory activity. Stem. Cells Dev 2010, 19, 27–38. [Google Scholar]
- Glennie, S.; Soeiro, I.; Dyson, P.J.; Lam, E.W.; Dazzi, F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood 2005, 105, 2821–2827. [Google Scholar]
- Corcione, A.; Benvenuto, F.; Ferretti, E.; Giunti, D.; Cappiello, V.; Cazzanti, F.; Risso, M.; Gualandi, F.; Mancardi, G.L.; Pistoia, V.; et al. Human mesenchymal stem cells modulate B-cell functions. Blood 2006, 107, 367–372. [Google Scholar]
- Asari, S.; Itakura, S.; Ferreri, K.; Liu, C.P.; Kuroda, Y.; Kandeel, F.; Mullen, Y. Mesenchymal stem cells suppress B-cell terminal differentiation. Exp. Hematol 2009, 37, 604–615. [Google Scholar]
- Angoulvant, D.; Clerc, A.; Benchalal, S.; Galambrun, C.; Farre, A.; Bertrand, Y.; Eljaafari, A. Human mesenchymal stem cells suppress induction of cytotoxic response to alloantigens. Biorheology 2004, 41, 469–476. [Google Scholar]
- Rasmusson, I.; Ringden, O.; Sundberg, B.; Le Blanc, K. Mesenchymal stem cells inhibit the formation of cytotoxic T lymphocytes, but not activated cytotoxic T lymphocytes or natural killer cells. Transplantation 2003, 76, 1208–1213. [Google Scholar]
- Spaggiari, G.M.; Capobianco, A.; Becchetti, S.; Mingari, M.C.; Moretta, L. Mesenchymal stem cell-natural killer cell interactions: Evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell proliferation. Blood 2006, 107, 1484–1490. [Google Scholar]
- Poggi, A.; Prevosto, C.; Massaro, A.M.; Negrini, S.; Urbani, S.; Pierri, I.; Saccardi, R.; Gobbi, M.; Zocchi, M.R. Interaction between human NK cells and bone marrow stromal cells inducesNK cell triggering: Role of NKp30 and NKG2D receptors. J. Immunol 2005, 175, 6352–6360. [Google Scholar]
- Hoogduijn, M.J.; Korevaar, S.S.; Engela, A.U.; Weimar, W.; Baan, C.C. Immunological aspects of allogeneic and autologous mesenchymal stem cell therapies. Hum. Gene Ther 2011, 22, 1587–1591. [Google Scholar]
- Crop, M.J.; Korevaar, S.S.; de Kuiper, R.; Ijzermans, J.N.; van Besouw, N.M.; Baan, C.C.; Weimar, W.; Hoogduijn, M.J. Human mesenchymal stem cells are susceptible to lysis by CD8+ T-cells and NK cells. Cell Transplant 2011. [Google Scholar] [CrossRef]
- Gonzalez, M.A.; Gonzalez-Rey, E.; Rico, L.; Buscher, D.; Delgado, M. Adipose-derived mesenchymal stem cells alleviate experimental colitis by inhibiting inflammatory and autoimmune responses. Gastroenterology 2009, 136, 978–989. [Google Scholar]
- Christensen, M.E.; Turner, B.E.; Sinfield, L.J.; Kollar, K.; Cullup, H.; Waterhouse, N.J.; Hart, D.N.; Atkinson, K.; Rice, A.M. Mesenchymal stromal cells transiently alter the inflammatory milieu post-transplant to delay graft-versus-host disease. Haematologica 2010, 95, 2102–2110. [Google Scholar]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar]
- Groh, M.E.; Maitra, B.; Szekely, E.; Koc, O.N. Human mesenchymal stem cells require monocyte-mediated activation to suppress alloreactive T cells. Exp. Hematol 2005, 33, 928–934. [Google Scholar]
- Djouad, F.; Plence, P.; Bony, C.; Tropel, P.; Apparailly, F.; Sany, J.; Noel, D.; Jorgensen, C. Immunosuppressive effect of mesenchymal stem cells favors tumor growth in allogeneic animals. Blood 2003, 102, 3837–3844. [Google Scholar]
- Abumaree, M.; Al Jumah, M.; Pace, R.A.; Kalionis, B. Immunosuppressive properties of mesenchymal stem cells. Stem. Cell Rev 2012, 8, 375–392. [Google Scholar]
- Jiang, X.X.; Zhang, Y.; Liu, B.; Zhang, S.X.; Wu, Y.; Yu, X.D.; Mao, N. Human mesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood 2005, 105, 4120–4126. [Google Scholar]
- Zhang, Y.; Li, C.; Jiang, X.; Zhang, S.; Wu, Y.; Liu, B.; Tang, P.; Mao, N. Human placenta-derived mesenchymal progenitor cells support culture expansion of long-term culture-initiating cells from cord blood CD34+ cells. Exp. Hematol 2004, 32, 657–664. [Google Scholar]
- Xu, G.; Zhang, L.; Ren, G.; Yuan, Z.; Zhang, Y.; Zhao, R.C.; Shi, Y. Immunosuppressive properties of cloned bone marrow mesenchymal stem cells. Cell Res 2007, 17, 240–248. [Google Scholar]
- Bian, L.; Guo, Z.K.; Wang, H.X.; Wang, J.S.; Wang, H.; Li, Q.F.; Yang, Y.F.; Xiao, F.J.; Wu, C.T.; Wang, L.S. In vitro and in vivo immunosuppressive characteristics of hepatocyte growth factor-modified murine mesenchymal stem cells. In Vivo 2009, 23, 21–27. [Google Scholar]
- Cargnoni, A.; Gibelli, L.; Tosini, A.; Signoroni, P.B.; Nassuato, C.; Arienti, D.; Lombardi, G.; Albertini, A.; Wengler, G.S.; Parolini, O. Transplantation of allogeneic and xenogeneic placenta-derived cells reduces bleomycin-induced lung fibrosis. Cell Transplant 2009, 18, 405–422. [Google Scholar]
- Devine, S.M.; Bartholomew, A.M.; Mahmud, N.; Nelson, M.; Patil, S.; Hardy, W.; Sturgeon, C.; Hewett, T.; Chung, T.; Stock, W.; et al. Mesenchymal stem cells are capable of homing to the bone marrow of non-human primates following systemic infusion. Exp. Hematol 2001, 29, 244–255. [Google Scholar]
- Lu, X.; Liu, T.; Gu, L.; Huang, C.; Zhu, H.; Meng, W.; Xi, Y.; Li, S.; Liu, Y. Immunomodulatory effects of mesenchymal stem cells involved in favoring type 2 T cell subsets. Transpl. Immunol 2009, 22, 55–61. [Google Scholar]
- Kopen, G.C.; Prockop, D.J.; Phinney, D.G. Marrow stromal cells migrate throughout forebrain and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse brains. Proc. Natl. Acad. Sci USA 1999, 96, 10711–10716. [Google Scholar]
- Devine, S.M.; Cobbs, C.; Jennings, M.; Bartholomew, A.; Hoffman, R. Mesenchymal stem cells distribute to a wide range of tissues following systemic infusion into nonhuman primates. Blood 2003, 101, 2999–3001. [Google Scholar]
- Sanchez-Ramos, J.; Song, S.; Cardozo-Pelaez, F.; Hazzi, C.; Stedeford, T.; Willing, A.; Freeman, T.B.; Saporta, S.; Janssen, W.; Patel, N.; et al. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp. Neurol 2000, 164, 247–256. [Google Scholar]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res 2000, 61, 364–370. [Google Scholar]
- Woodbury, D.; Reynolds, K.; Black, I.B. Adult bone marrow stromal stem cells express germline, ectodermal, endodermal, and mesodermal genes prior to neurogenesis. J. Neurosci. Res 2002, 69, 908–917. [Google Scholar]
- Azizi, S.A.; Stokes, D.; Augelli, B.J.; DiGirolamo, C.; Prockop, D.J. Engraftment and migration of human bone marrow stromal cells implanted in the brains of albino rats—similarities to astrocyte grafts. Proc. Natl. Acad. Sci. USA 1998, 95, 3908–3913. [Google Scholar]
- Chen, J.; Li, Y.; Wang, L.; Lu, M.; Zhang, X.; Chopp, M. Therapeutic benefit of intracerebral transplantation of bone marrow stromal cells after cerebral ischemia in rats. J. Neurol. Sci 2001, 189, 49–57. [Google Scholar]
- Zhang, J.; Li, Y.; Chen, J.; Yang, M.; Katakowski, M.; Lu, M.; Chopp, M. Expression of insulin-like growth factor 1 and receptor in ischemic rats treated with human marrow stromal cells. Brain Res 2004, 1030, 19–27. [Google Scholar]
- Zhao, L.R.; Duan, W.M.; Reyes, M.; Keene, C.D.; Verfaillie, C.M.; Low, W.C. Human bone marrow stem cells exhibit neural phenotypes and ameliorate neurological deficits after grafting into the ischemic brain of rats. Exp. Neurol 2002, 174, 11–20. [Google Scholar]
- Munoz-Elias, G.; Marcus, A.J.; Coyne, T.M.; Woodbury, D.; Black, I.B. Adult bone marrow stromal cells in the embryonic brain: Engraftment, migration, differentiation, and long-term survival. J. Neurosci 2004, 24, 4585–4595. [Google Scholar]
- Lepski, G.; Jannes, C.E.; Strauss, B.; Marie, S.K.; Nikkhah, G. Survival and neuronal differentiation of mesenchymal stem cells transplanted into the rodent brain are dependent upon microenvironment. Tissue Eng. Part A 2010, 16, 2769–2782. [Google Scholar]
- Park, S.; Kim, E.; Koh, S.E.; Maeng, S.; Lee, W.D.; Lim, J.; Shim, I.; Lee, Y.J. Dopaminergic differentiation of neural progenitors derived from placental mesenchymal stem cells in the brains of Parkinson’s disease model rats and alleviation of asymmetric rotational behavior. Brain Res 2012, 1466, 158–166. [Google Scholar]
- Zhang, J.; Li, Y.; Chen, J.; Cui, Y.; Lu, M.; Elias, S.B.; Mitchell, J.B.; Hammill, L.; Vanguri, P.; Chopp, M. Human bone marrow stromal cell treatment improves neurological functional recovery in EAE mice. Exp. Neurol 2005, 195, 16–26. [Google Scholar]
- Zhang, J.; Li, Y.; Lu, M.; Cui, Y.; Chen, J.; Noffsinger, L.; Elias, S.B.; Chopp, M. Bone marrow stromal cells reduce axonal loss in experimental autoimmune encephalomyelitis mice. J. Neurosci. Res 2006, 84, 587–595. [Google Scholar]
- Liu, X.J.; Zhang, J.F.; Sun, B.; Peng, H.S.; Kong, Q.F.; Bai, S.S.; Liu, Y.M.; Wang, G.Y.; Wang, J.H.; Li, H.L. Reciprocal effect of mesenchymal stem cell on experimental autoimmune encephalomyelitis is mediated by transforming growth factor-beta and interleukin-6. Clin. Exp. Immunol 2009, 158, 37–44. [Google Scholar]
- Peron, J.P.; Jazedje, T.; Brandao, W.N.; Perin, P.M.; Maluf, M.; Evangelista, L.P.; Halpern, S.; Nisenbaum, M.G.; Czeresnia, C.E.; Zatz, M.; et al. Human endometrial-derived mesenchymal stem cells suppress inflammation in the central nervous system of EAE mice. Stem. Cell Rev 2011. [Google Scholar] [CrossRef]
- Barhum, Y.; Gai-Castro, S.; Bahat-Stromza, M.; Barzilay, R.; Melamed, E.; Offen, D. Intracerebroventricular transplantation of human mesenchymal stem cells induced to secrete neurotrophic factors attenuates clinical symptoms in a mouse model of multiple sclerosis. J. Mol. Neurosci 2010, 41, 129–137. [Google Scholar]
- Gordon, D.; Pavlovska, G.; Uney, J.B.; Wraith, D.C.; Scolding, N.J. Human mesenchymal stem cells infiltrate the spinal cord, reduce demyelination, and localize to white matter lesions in experimental autoimmune encephalomyelitis. J. Neuropathol. Exp. Neurol 2010, 69, 1087–1095. [Google Scholar]
- Fisher-Shoval, Y.; Barhum, Y.; Sadan, O.; Yust-Katz, S.; Ben-Zur, T.; Lev, N.; Benkler, C.; Hod, M.; Melamed, E.; Offen, D. Transplantation of placenta-derived mesenchymal stem cells in the EAE mouse model of MS. J. Mol. Neurosci 2012. [Google Scholar] [CrossRef]
- Bai, L.; Lennon, D.P.; Caplan, A.I.; Dechant, A.; Hecker, J.; Kranso, J.; Zaremba, A.; Miller, R.H. Hepatocyte growth factor mediates mesenchymal stem cell-induced recovery in multiple sclerosis models. Nat. Neurosci 2012. [Google Scholar] [CrossRef]
- Neuhuber, B.; Timothy Himes, B.; Shumsky, J.S.; Gallo, G.; Fischer, I. Axon growth and recovery of function supported by human bone marrow stromal cells in the injured spinal cord exhibit donor variations. Brain Res 2005, 1035, 73–85. [Google Scholar]
- Rivera, F.J.; Couillard-Despres, S.; Pedre, X.; Ploetz, S.; Caioni, M.; Lois, C.; Bogdahn, U.; Aigner, L. Mesenchymal stem cells instruct oligodendrogenic fate decision on adult neural stem cells. Stem. Cells 2006, 24, 2209–2219. [Google Scholar]
- Rivera, F.J.; Siebzehnrubl, F.A.; Kandasamy, M.; Couillard-Despres, S.; Caioni, M.; Poehler, A.M.; Berninger, B.; Sandner, B.; Bogdahn, U.; Goetz, M.; et al. Mesenchymal stem cells promote oligodendroglial differentiation in hippocampal slice cultures. Cell Physiol. Biochem 2009, 24, 317–324. [Google Scholar]
- Phinney, D.G.; Prockop, D.J. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair–current views. Stem. Cells 2007, 25, 2896–2902. [Google Scholar]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar]
- Parekkadan, B.; van Poll, D.; Suganuma, K.; Carter, E.A.; Berthiaume, F.; Tilles, A.W.; Yarmush, M.L. Mesenchymal stem cell-derived molecules reverse fulminant hepatic failure. PLoS One 2007, 2. [Google Scholar] [CrossRef]
- Scuteri, A.; Cassetti, A.; Tredici, G. Adult mesenchymal stem cells rescue dorsal root ganglia neurons from dying. Brain Res 2006, 1116, 75–81. [Google Scholar]
- Crigler, L.; Robey, R.C.; Asawachaicharn, A.; Gaupp, D.; Phinney, D.G. Human mesenchymal stem cell subpopulations express a variety of neuro-regulatory molecules and promote neuronal cell survival and neuritogenesis. Exp. Neurol 2006, 198, 54–64. [Google Scholar]
- Wilkins, A.; Kemp, K.; Ginty, M.; Hares, K.; Mallam, E.; Scolding, N. Human bone marrow-derived mesenchymal stem cells secrete brain-derived neurotrophic factor which promotes neuronal survival in vitro. Stem Cell Res. 2009, 3, 63–70. [Google Scholar]
- Bai, L.; Caplan, A.; Lennon, D.; Miller, R.H. Human mesenchymal stem cells signals regulate neural stem cell fate. Neurochem. Res 2007, 32, 353–362. [Google Scholar]
- Akiyama, Y.; Radtke, C.; Honmou, O.; Kocsis, J.D. Remyelination of the spinal cord following intravenous delivery of bone marrow cells. Glia 2002, 39, 229–236. [Google Scholar]
- Satake, K.; Lou, J.; Lenke, L.G. Migration of mesenchymal stem cells through cerebrospinal fluid into injured spinal cord tissue. Spine (Phila Pa 1976) 2004, 29, 1971–1979. [Google Scholar]
- Vercelli, A.; Mereuta, O.M.; Garbossa, D.; Muraca, G.; Mareschi, K.; Rustichelli, D.; Ferrero, I.; Mazzini, L.; Madon, E.; Fagioli, F. Human mesenchymal stem cell transplantation extends survival, improves motor performance and decreases neuroinflammation in mouse model of amyotrophic lateral sclerosis. Neurobiol. Dis 2008, 31, 395–405. [Google Scholar]
- Chen, J.; Li, Y.; Katakowski, M.; Chen, X.; Wang, L.; Lu, D.; Lu, M.; Gautam, S.C.; Chopp, M. Intravenous bone marrow stromal cell therapy reduces apoptosis and promotes endogenous cell proliferation after stroke in female rat. J. Neurosci. Res 2003, 73, 778–786. [Google Scholar]
- Li, Y.; Chen, J.; Chen, X.G.; Wang, L.; Gautam, S.C.; Xu, Y.X.; Katakowski, M.; Zhang, L.J.; Lu, M.; Janakiraman, N.; et al. Human marrow stromal cell therapy for stroke in rat: Neurotrophins and functional recovery. Neurology 2002, 59, 514–523. [Google Scholar]
- Munoz, J.R.; Stoutenger, B.R.; Robinson, A.P.; Spees, J.L.; Prockop, D.J. Human stem/progenitor cells from bone marrow promote neurogenesis of endogenous neural stem cells in the hippocampus of mice. Proc. Natl. Acad. Sci. USA 2005, 102, 18171–18176. [Google Scholar]
- Ohtaki, H.; Ylostalo, J.H.; Foraker, J.E.; Robinson, A.P.; Reger, R.L.; Shioda, S.; Prockop, D.J. Stem/progenitor cells from bone marrow decrease neuronal death in global ischemia by modulation of inflammatory/immune responses. Proc. Natl. Acad. Sci. USA 2008, 105, 14638–14643. [Google Scholar]
- Zhou, C.; Zhang, C.; Chi, S.; Xu, Y.; Teng, J.; Wang, H.; Song, Y.; Zhao, R. Effects of human marrow stromal cells on activation of microglial cells and production of inflammatory factors induced by lipopolysaccharide. Brain Res 2009, 1269, 23–30. [Google Scholar]
- Kim, Y.J.; Park, H.J.; Lee, G.; Bang, O.Y.; Ahn, Y.H.; Joe, E.; Kim, H.O.; Lee, P.H. Neuroprotective effects of human mesenchymal stem cells on dopaminergic neurons through anti-inflammatory action. Glia 2009, 57, 13–23. [Google Scholar]
- Lanza, C.; Morando, S.; Voci, A.; Canesi, L.; Principato, M.C.; Serpero, L.D.; Mancardi, G.; Uccelli, A.; Vergani, L. Neuroprotective mesenchymal stem cells are endowed with a potent antioxidant effect in vivo. J. Neurochem 2009, 110, 1674–1684. [Google Scholar]
- Bork, S.; Pfister, S.; Witt, H.; Horn, P.; Korn, B.; Ho, A.D.; Wagner, W. DNA methylation pattern changes upon long-term culture and aging of human mesenchymal stromal cells. Aging Cell 2010, 9, 54–63. [Google Scholar]
- Bonab, M.M.; Alimoghaddam, K.; Talebian, F.; Ghaffari, S.H.; Ghavamzadeh, A.; Nikbin, B. Aging of mesenchymal stem cell in vitro. BMC Cell Biol. 2006, 7. [Google Scholar] [CrossRef]
- Kassem, M.; Marie, P.J. Senescence-associated intrinsic mechanisms of osteoblast dysfunctions. Aging Cell 2011, 10, 191–197. [Google Scholar]
- Mareschi, K.; Ferrero, I.; Rustichelli, D.; Aschero, S.; Gammaitoni, L.; Aglietta, M.; Madon, E.; Fagioli, F. Expansion of mesenchymal stem cells isolated from pediatric and adult donor bone marrow. J. Cell Biochem 2006, 97, 744–754. [Google Scholar]
- Moerman, E.J.; Teng, K.; Lipschitz, D.A.; Lecka-Czernik, B. Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: The role of PPAR-γ2 transcription factor and TGF-β/BMP signaling pathways. Aging Cell 2004, 3, 379–389. [Google Scholar]
- Schallmoser, K.; Bartmann, C.; Rohde, E.; Bork, S.; Guelly, C.; Obenauf, A.C.; Reinisch, A.; Horn, P.; Ho, A.D.; Strunk, D.; et al. Replicative senescence-associated gene expression changes in mesenchymal stromal cells are similar under different culture conditions. Haematologica 2010, 95, 867–874. [Google Scholar]
- Stolzing, A.; Scutt, A. Age-related impairment of mesenchymal progenitor cell function. Aging Cell 2006, 5, 213–224. [Google Scholar] [Green Version]
- Tokalov, S.V.; Gruener, S.; Schindler, S.; Iagunov, A.S.; Baumann, M.; Abolmaali, N.D. A number of bone marrow mesenchymal stem cells but neither phenotype nor differentiation capacities changes with age of rats. Mol. Cells 2007, 24, 255–260. [Google Scholar]
- Wagner, W.; Ho, A.D.; Zenke, M. Different facets of aging in human mesenchymal stem cells. Tissue Eng. Part B Rev 2010, 16, 445–453. [Google Scholar]
- Wilson, A.; Shehadeh, L.A.; Yu, H.; Webster, K.A. Age-related molecular genetic changes of murine bone marrow mesenchymal stem cells. BMC Genomics 2010, 11. [Google Scholar] [CrossRef]
- Zhou, S.; Greenberger, J.S.; Epperly, M.W.; Goff, J.P.; Adler, C.; Leboff, M.S.; Glowacki, J. Age-related intrinsic changes in human bone-marrow-derived mesenchymal stem cells and their differentiation to osteoblasts. Aging Cell 2008, 7, 335–343. [Google Scholar]
- Bergman, R.J.; Gazit, D.; Kahn, A.J.; Gruber, H.; McDougall, S.; Hahn, T.J. Age-related changes in osteogenic stem cells in mice. J. Bone Miner. Res 1996, 11, 568–577. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Al Jumah, M.A.; Abumaree, M.H. The Immunomodulatory and Neuroprotective Effects of Mesenchymal Stem Cells (MSCs) in Experimental Autoimmune Encephalomyelitis (EAE): A Model of Multiple Sclerosis (MS). Int. J. Mol. Sci. 2012, 13, 9298-9331. https://doi.org/10.3390/ijms13079298
Al Jumah MA, Abumaree MH. The Immunomodulatory and Neuroprotective Effects of Mesenchymal Stem Cells (MSCs) in Experimental Autoimmune Encephalomyelitis (EAE): A Model of Multiple Sclerosis (MS). International Journal of Molecular Sciences. 2012; 13(7):9298-9331. https://doi.org/10.3390/ijms13079298
Chicago/Turabian StyleAl Jumah, Mohammed A., and Mohamed H. Abumaree. 2012. "The Immunomodulatory and Neuroprotective Effects of Mesenchymal Stem Cells (MSCs) in Experimental Autoimmune Encephalomyelitis (EAE): A Model of Multiple Sclerosis (MS)" International Journal of Molecular Sciences 13, no. 7: 9298-9331. https://doi.org/10.3390/ijms13079298