
Effect of Photon Hormesis on Dose Responses to Alpha Particles in Zebrafish Embryos

Abstract
:1. Introduction
2. Results
2.1. Part A: Effect of Alpha-Particle Dose with or without Additional Photon Dose
- (1)
- A1 group: embryos received ~1.1 mGy alpha-particle irradiation at 5 hpf;
- (2)
- A2 group: embryos received ~2.2 mGy alpha-particle irradiation at 5 hpf;
- (3)
- A4 group: embryos received ~4.4 mGy alpha-particle irradiation at 5 hpf;
- (4)
- A8 group: embryos received ~8.8 mGy alpha-particle irradiation at 5 hpf; and
- (5)
- AC group: embryos were sham irradiated with alpha particles at 5 hpf.
- (1)
- AX1 group: embryos received ~1.1 mGy alpha-particle irradiation and 10 mGy X-ray irradiation at 5 hpf;
- (2)
- AX2 group: embryos received ~2.2 mGy alpha-particle irradiation and 10 mGy X-ray irradiation at 5 hpf;
- (3)
- AX4 group: embryos received ~4.4 mGy alpha-particle irradiation and 10 mGy X-ray irradiation at 5 hpf;
- (4)
- AX8 group: embryos received ~8.8 mGy alpha-particle irradiation and 10 mGy X-ray irradiation at 5 hpf; and
- (5)
- AXC group: embryos were sham irradiated with alpha particles and X-rays at 5 hpf.
- (i)
- NA1+ < NAX1+, photon hormesis was absent, p = 3.44 × 10−5
- (ii)
- NA2+ < NAX2+, photon hormesis was absent, p = 6.75 × 10−10
- (iii)
- NA4+ > NAX4+, photon hormesis was present, p = 0.0185
- (iv)
- NA8+ > NAX8+, photon hormesis was present, p = 1.19 × 10−8
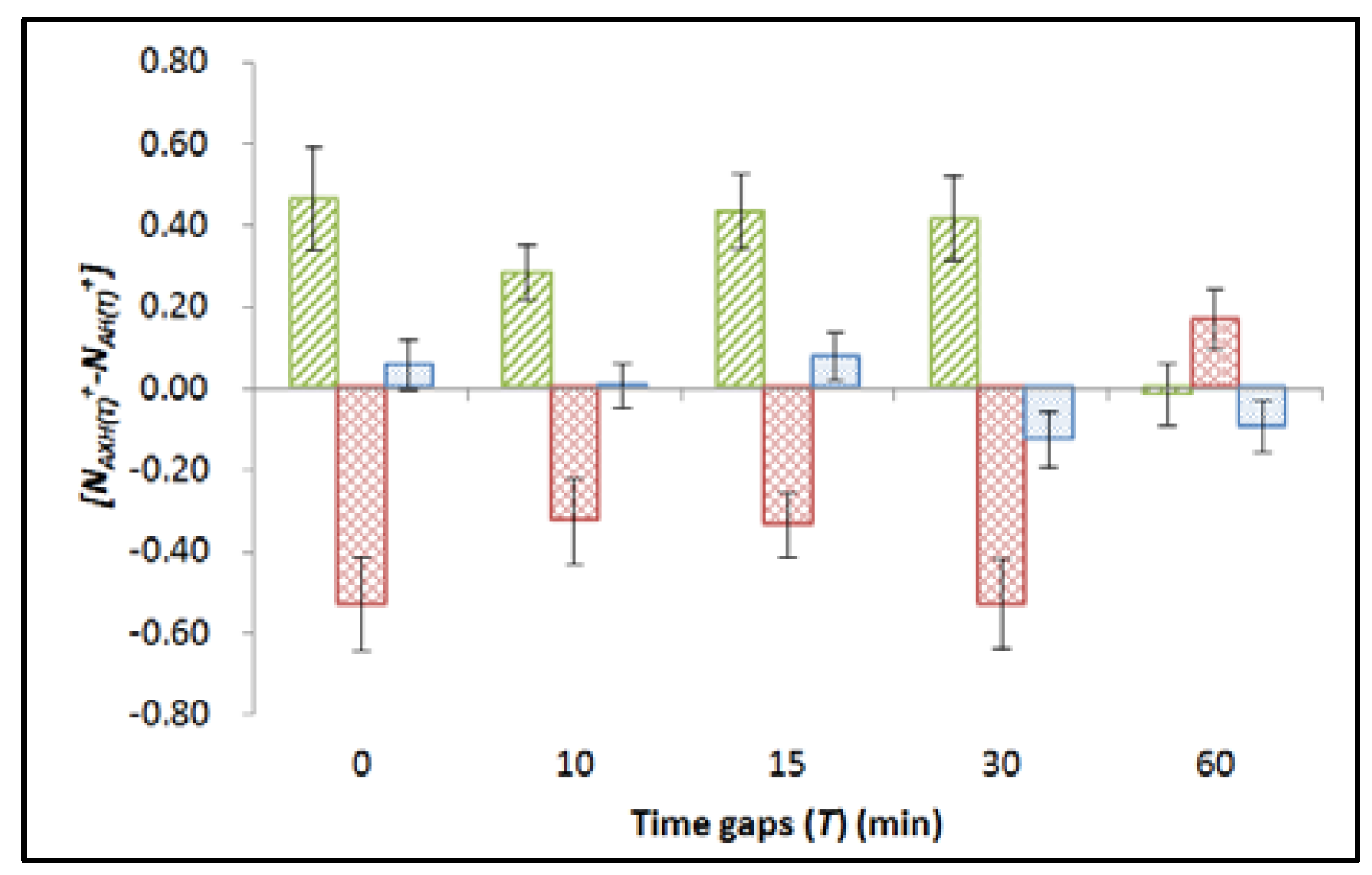
2.2. Part B: Effect of Time Gap between Alpha-Particle and X-ray Irradiations
- (1)
- AXH(T) group: embryos were irradiated with 4.4 mGy alpha-particle dose and then received 10 mGy X-ray dose with H = 24 (hpf) and T = 0, 10, 15, 30, or 60 (min);
- (2)
- AH(T) group: embryos were irradiated with 4.4 mGy alpha-particle dose and then sham irradiated with X-rays, with H = 24 (hpf) and T = 0, 10, 15, 30, or 60 (min); and
- (3)
- CH(T) group: embryos were sham irradiated with alpha particles and X-rays, with H = 24 (hpf) and T = 0, 10, 15, 30, or 60 (min).
2.3. Part C: Effect of Time Point at Which Apoptotic Counts Were Determined
3. Discussion
4. Materials and Methods
4.1. Zebrafish Embryos
4.2. Alpha-Particle Irradiation
4.3. X-Ray Irradiation
4.4. Experimental Protocols
4.4.1. Part A: Effect of Alpha-Particle Dose with or without Additional Photon Dose
4.4.2. Part B: Effect of Time Gap between Alpha-Particle and X-ray Irradiations
4.4.3. Part C: Effect of Time Point at Which Apoptotic Counts Were Determined
4.5. Quantification of Apoptosis by Vital Dye Staining
4.6. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Geetanjali, P.; Sarma, A.; Chatterjee, A. The influence of reduced glutathione on chromosome damage induced by X-rays or heavy ion beams of different LETs and on the interaction of DNA lesions induced by radiations and bleomycin. Mutat. Res. 2010, 696, 154–159. [Google Scholar]
- Barendsen, G.N.; Beusker, T.L.J.; Vergroesen, A.J.; Budke, L. Effects of different ionizing radiations on human cells in tissue culture. II. Biological experiments. Radiat. Res. 1960, 13, 841–849. [Google Scholar] [CrossRef]
- Geard, C.R. Chromosomal aberration production by “track segment” charged particles as a function of linear energy transfer. Radiat. Prot. Dosim. 1985, 13, 1–4. [Google Scholar]
- Hendry, J.H.; Simon, S.L.; Wojcik, A.; Sohrabi, M.; Burkart, W.; Cardis, E.; Laurier, D.; Tirmarche, M.; Hayata, I. Human exposure to high natural background radiation: What can it teach us about radiation risks? J. Radiol. Prot. 2009, 29, A29–A42. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.N.; Guan, Z.J.; Stokes, M.J.; Young, E.C.M. The assessment of the natural radiation dose committed to the Hong Kong people. J. Environ. Radioact. 1992, 17, 31–48. [Google Scholar] [CrossRef]
- Yu, K.N.; Chan, T.F.; Young, E.C.M. The variation of radon exhalation rates from building surfaces of different ages. Health Phys. 1995, 68, 716–718. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.N.; Young, E.C.M.; Li, K.C. A study of factors affecting indoor radon properties. Health Phys. 1996, 71, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.N.; Young, E.C.M.; Stokes, M.J.; Guan, Z.J.; Cho, K.W. A survey of radon and thoron progeny for dwellings in Hong Kong. Health Phys. 1997, 73, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.N.; Nikezic, D.; Ng, F.M.F.; Leung, J.K.C. Long-term measurements of radon progeny concentrations with solid-state nuclear track detectors. Radiat. Meas. 2005, 40, 560–568. [Google Scholar] [CrossRef]
- Yu, K.N.; Lau, B.M.F.; Nikezic, D. Assessment of environmental radon hazard using human respiratory tract models. J. Hazard. Mater. 2006, 132, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Cucinotta, F.A. Heavy ion carcinogenesis and human space exploration. Nat. Rev. Cancer 2008, 8, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonsen, L.C.; Wilson, J.W.; Kim, M.H.; Cucinotta, F.A. Radiation exposure for human mars exploration. Health Phys. 2000, 79, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Coderre, J.A.; Morris, G.M. The radiation biology of boron neutron capture therapy. Radiat. Res. 1999, 151, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Durand, R.E.; Olive, P.L. Irradiation of multi-cell spheroids with fast neutrons versus X-rays: A qualitative difference in sub-lethal damage repair capacity or kinetics. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1976, 30, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Ngo, F.Q.H.; Elkind, M.M. Damage interaction in V79 Chinese hamster cells resulting from the combined exposure to X-rays and fast neutrons (Abstract Gg-5). Radiat. Res. 1977, 70, 706–707. [Google Scholar]
- McNally, N.J.; Deronde, J.; Hinchliffe, M. The effect of sequential irradiation with X-rays and fast neutrons on the survival of V79 Chinese hamster cells. Int. J. Radiat. Biol. 1984, 45, 301–310. [Google Scholar] [CrossRef]
- Ngo, F.Q.H.; Blakely, E.A.; Tobias, C.A. Sequential exposures of mammalian cells to low- and high-LET radiation, I. Lethal effects following X-ray and neon-ion irradiation. Radiat. Res. 1981, 87, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Murthy, M.S.S.; Madhvanath, U.; Subrahmanyam, P.; Rao, B.S.; Reddy, N.M.S. Synergistic effect of simultaneous exposure to 60Co gamma rays and 210Po alpha rays in diploid yeast. Radiat. Res. 1975, 63, 185–190. [Google Scholar] [CrossRef] [PubMed]
- McNally, N.J.; Deronde, J.; Folkard, M. Interaction between X-rays and alpha-particle damage in V-79 cells. Int. J. Radiat. Biol. 1988, 53, 917–920. [Google Scholar] [CrossRef]
- Brooks, A.L.; Newton, G.J.; Shyr, L.J.; Seiler, F.A.; Scott, B.R. The combined effects of alpha-particles and X-rays on cell killing and micronuclei induction in lung epithelial cells. Int. J. Radiat. Biol. 1990, 58, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Rithidech, K.N.; Scott, B.R. Evidence for radiation hormesis after in vitro exposure of human lymphocytes to low doses of ionizing radiation. Dose Response 2008, 6, 252–271. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormetic mechanisms. Crit. Rev. Toxicol. 2013, 43, 580–606. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormesis: Why it is important to toxicology and toxicologists. Environ. Toxicol. Chem. 2008, 27, 1452–1474. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Linda, A.B. Toxicology rethinks its central belief. Nature 2003, 421, 691–692. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Defining hormesis. Hum. Exp. Toxicol. 2002, 21, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. The emergence of the dose–response concept in biology and medicine. Int. J. Mol. Sci. 2016, 17, 2034. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.P.; Kong, E.Y.; Konishi, T.; Kobayashi, A.; Suya, N.; Cheng, S.H.; Yu, K.N. Low-dose neutron dose response of zebrafish embryos obtained from the Neutron exposure Accelerator System for Biological Effect Experiments (NASBEE) facility. Radiat. Phys. Chem. 2015, 114, 12–17. [Google Scholar] [CrossRef]
- Scott, B.R. Low-dose-radiation stimulated natural chemical and biological protection against lung cancer. Dose Response 2008, 6, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.R.; Sanders, C.L.; Mitchel, R.E.J.; Boreham, D.R. CT scans may reduce rather than increase the risk of cancer. J. Am. Phys. Surg. 2008, 13, 8–11. [Google Scholar]
- Bauer, G. Low dose radiation and intercellular induction of apoptosis: Potential implications for the control of oncogenesis. Int. J. Radiat. Biol. 2007, 83, 873–888. [Google Scholar] [CrossRef] [PubMed]
- Portess, D.I.; Bauer, G.; Hill, M.A.; O’Neill, P. Low-dose irradiation of nontransformed cells stimulates the selective removal of precancerous cells via intercellular induction of apoptosis. Cancer Res. 2007, 67, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.R.; Di Palma, J. Sparsely ionizing diagnostic and natural background radiation are likely preventing cancer and other genomic-instability associated diseases. Dose Response 2006, 5, 230–255. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.P.; Kong, E.Y.; Kobayashi, A.; Suya, N.; Uchihori, Y.; Cheng, S.H.; Konishi, T.; Yu, K.N. Neutron induced bystander effect among zebrafish embryos. Radiat. Phys. Chem. 2015, 117, 153–159. [Google Scholar] [CrossRef]
- Bladen, C.L.; Lam, W.K.; Dynan, W.S.; Kozlowski, D.J. DNA damage response and Ku80 function in the vertebrate embryo. Nucleic Acids Res. 2005, 33, 3002–3010. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Konishi, T.; Oikawa, M.; Cheng, S.H.; Yu, K.N. Threshold number of protons for inducing adaptive response in zebrafish embryos. J. Radiol. Prot. 2013, 33, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Ng, C.Y.P.; Kobayashi, A.; Konishi, T.; Suya, N.; Ishikawa, T.; Cheng, S.H.; Yu, K.N. Bystander effect between zebrafish embryos in vivo induced by high-dose X-rays. Environ. Sci. Technol. 2013, 47, 6368–6376. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Ng, C.Y.P.; Kong, M.K.Y.; Cheng, S.H.; Yu, K.N. Adaptive response to ionizing radiation induced by cadmium in zebrafish embryos. J. Radiol. Prot. 2013, 33, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Ng, C.Y.P.; Kobayashi, A.; Konishi, T.; Oikawa, M.; Cheng, S.H.; Yu, P.K.N. Exogenous carbon monoxide suppresses adaptive response induced in zebrafish embryos in vivo by microbeam protons. J. Radiat. Res. 2014, 55, i115. [Google Scholar] [CrossRef]
- Choi, V.W.Y.; Yu, K.N. Embryos of the zebrafish Danio rerio in studies of non-targeted effects of ionizing radiation. Cancer Lett. 2015, 356, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Daroczi, B.; Kari, G.; McAleer, M.F.; Wolf, J.C.; Rodeck, U.; Dicker, A.P. In vivo radioprotection by the fullerene nanoparticle DF-1 as assessed in a zebrafish model. Clin. Cancer Res. 2006, 12, 7086–7091. [Google Scholar] [CrossRef] [PubMed]
- Geiger, G.A.; Parker, S.E.; Beothy, A.P.; Tucker, J.A.; Mullins, M.C.; Kao, G.D. Zebrafish as a “Biosensor”? Effects of ionizing radiation and amifostine on embryonic viability and development. Cancer Res. 2006, 66, 8172–8181. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.Y.; Cheng, S.H.; Yu, K.N. Biphasic and triphasic dose responses in zebrafish embryos to low-dose 150 kV X-rays with different hardness. J. Radiat. Res. 2016, 57, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.Y.; Yeung, W.K.; Chan, T.K.Y.; Cheng, S.H.; Yu, K.N. Exogenous nitric oxide suppresses in vivo X-ray-induced targeted and non-targeted effects in zebrafish embryos. Int. J. Mol. Sci. 2016, 17, 1321. [Google Scholar] [CrossRef] [PubMed]
- McAleer, M.F.; Davidson, C.; Davidson, W.R.; Yentzer, B.Y.; Farber, S.A.; Rodeck, U.; Dicker, A.P. Novel use of zebrafish as a vertebrate model to screen radiation protectors and sensitizers. Int. J. Radiat. Oncol. Biol. Phys. 2005, 61, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.P.; Pereira, S.; Cheng, S.H.; Adam-Guillermin, C.; Garnier-Laplace, J.; Yu, K.N. Combined effects of depleted uranium and ionising radiation on zebrafish embryos. Radiat. Prot. Dosim. 2015, 167, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.P.; Cheng, S.H.; Yu, K.N. Hormetic effect induced by depleted uranium in zebrafish embryos. Aquat. Toxicol. 2016, 175, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.P.; Kong, E.Y.; Kobayashi, A.; Suya, N.; Uchihori, Y.; Cheng, S.H.; Konishi, T.; Yu, K.N. Non-induction of radioadaptive response in zebrafish embryos by neutrons. J. Radiat. Res. 2016, 57, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.P.; Pereira, S.; Cheng, S.H.; Adam-Guillermin, C.; Garnier-Laplace, J.; Yu, K.N. Combined effects of alpha particles and depleted uranium on zebrafish (Danio rerio) embryos. J. Radiat. Res. 2016, 57, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Yum, E.H.W.; Ng, C.K.M.; Lin, A.C.C.; Cheng, S.H.; Yu, K.N. Effects of alpha particles on zebrafish embryos. Appl. Radiat. Isot. 2010, 68, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.Y.; Cheng, S.H.; Yu, K.N. Zebrafish as an in vivo model to assess epigenetic effects of ionizing radiation. Int. J. Mol. Sci. 2016, 17, 2108. [Google Scholar] [CrossRef] [PubMed]
- Barbazuk, W.B.; Korf, I.; Kadavi, C.; Heyen, J.; Tate, S.; Wun, E.; Bedell, J.A.; McPherson, J.D.; Johnson, S.L. The syntenic relationship of the zebrafish and human genomes. Genome Res. 2000, 10, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; von Hofsten, J.; Olsson, P.E. Generating transparent zebrafish: A refined method to improve detection of gene expression during embryonic development. Mar. Biotechnol. 2001, 10, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.C.; McNally, N.J. Cell survival and DNA double-strand break repair following X-ray or neutron irradiation of V79 cells. Int. J. Radiat. Biol. 1988, 54, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Ngo, F.Q.H.; Han, A.; Elkind, M.M. On the repair of sub-lethal damage in V79 Chinese hamster cells resulting from irradiation with fast neutrons or fast neutrons combined with X-rays. Int. J. Radiat. Biol. 1977, 32, 507–511. [Google Scholar] [CrossRef]
- Ng, C.Y.P.; Cheng, S.H.; Yu, K.N. Photon hormesis deactivates alpha-particle induced bystander effects between zebrafish embryos. Radiat. Phys. Chem. 2017, 133, 72–80. [Google Scholar] [CrossRef]
- Yum, E.H.W.; Ng, C.K.M.; Lin, A.C.C.; Cheng, S.H.; Yu, K.N. Experimental setup for studying the effects of alpha particles on zebrafish embryos. Nucl. Instrum. Methods B 2007, 264, 171–176. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Sshilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Milos, N.; Dingle, A.D. Dynamics of pigment pattern formation in the zebrafish, BrachyDanio rerio. I. Establishment and regulation of the lateral line melanophore stripe during the first eight days of development. J. Exp. Zool. A 1978, 205, 205–216. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T (min) | ANOVA a | Post-Hoc t-Tests | Photon Hormesis | |
|---|---|---|---|---|
| NAX24(T)+ vs. NC24(T)+ b | NAX24(T)+ vs. NA24(T)+ c | |||
| 0 | p = 1.02 × 10−11 * | p = 1.37 × 10−6 ** | p = 2.58 × 10−5 ** | Yes (NAX24(0)+ < NA24(0)+) |
| 10 | p = 6.74 × 10−11 * | p = 1.17 × 10−5 ** | p = 1.50 × 10−3 ** | Yes (NAX24(10)+ < NA24(10)+) |
| 15 | p = 2.34 × 10−13 * | p = 3.10 × 10−6 ** | p = 4.40 × 10−5 ** | Yes (NAX24(15)+ < NA24(15)+) |
| 30 | p = 4.38 × 10−17 * | p = 1.28 × 10−7 ** | p = 6.04 × 10−6 ** | Yes (NAX24(30)+ < NA24(30)+) |
| 60 | p = 1.71 × 10−12 * | p = 1.09 × 10−10 ** | p = 0.0113 ** | No (NAX24(60)+ > NA24(60)+) |
| T (min) | ANOVA a | Post-Hoc t-Tests | Photon Hormesis | |
|---|---|---|---|---|
| NAX20(T)+ vs. NC20(T)+ b | NAX20(T)+ vs. NA20(T)+ c | |||
| 0 | p = 1.21 × 10−15 * | p = 1.04 × 10−11 ** | p = 2.94 × 10−4 ** | No (NAX20(0)+ > NA20(0)+) |
| 10 | p = 3.22 × 10−15 * | p = 2.40 × 10−13 ** | p = 6.49 × 10−5 ** | No (NAX20(10)+ > NA20(10)+) |
| 15 | p = 9.23 × 10−17 * | p = 9.88 × 10−15 ** | p = 5.01 × 10−6 ** | No (NAX20(15)+ > NA20(15)+) |
| 30 | p = 5.58 × 10−9 * | p = 8.26 × 10−9 ** | p = 1.00 × 10−4 ** | No (NAX20(30)+ > NA20(30)+) |
| 60 | p = 2.98 × 10−6 * | p = 2.28 × 10−7 ** | p = 0.414 | No (NAX20(60)+ ~ NA20(60)+) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, C.Y.P.; Cheng, S.H.; Yu, K.N. Effect of Photon Hormesis on Dose Responses to Alpha Particles in Zebrafish Embryos. Int. J. Mol. Sci. 2017, 18, 385. https://doi.org/10.3390/ijms18020385
Ng CYP, Cheng SH, Yu KN. Effect of Photon Hormesis on Dose Responses to Alpha Particles in Zebrafish Embryos. International Journal of Molecular Sciences. 2017; 18(2):385. https://doi.org/10.3390/ijms18020385
Chicago/Turabian StyleNg, Candy Yuen Ping, Shuk Han Cheng, and Kwan Ngok Yu. 2017. "Effect of Photon Hormesis on Dose Responses to Alpha Particles in Zebrafish Embryos" International Journal of Molecular Sciences 18, no. 2: 385. https://doi.org/10.3390/ijms18020385