1. Introduction
Ultraviolet (UV) rays are capable of inducing melanin production in melanocytes and promoting melanin transportation to the outermost layer of the skin—the keratinocytes. These melanins form a cap over the nucleus of both cell types and protect DNA from direct energy destruction [
1,
2]. On the other hand, UV rays are also able to initiate nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) dominated reactive oxygen species (ROS) production and chemiexcitation of melanin fragments that affect DNA stability in melanocytes [
3,
4,
5]. The oncogenic characteristics of UV-induced ROS signaling have not yet been fully elucidated, particularly in the transformation of melanocytes to melanomas.
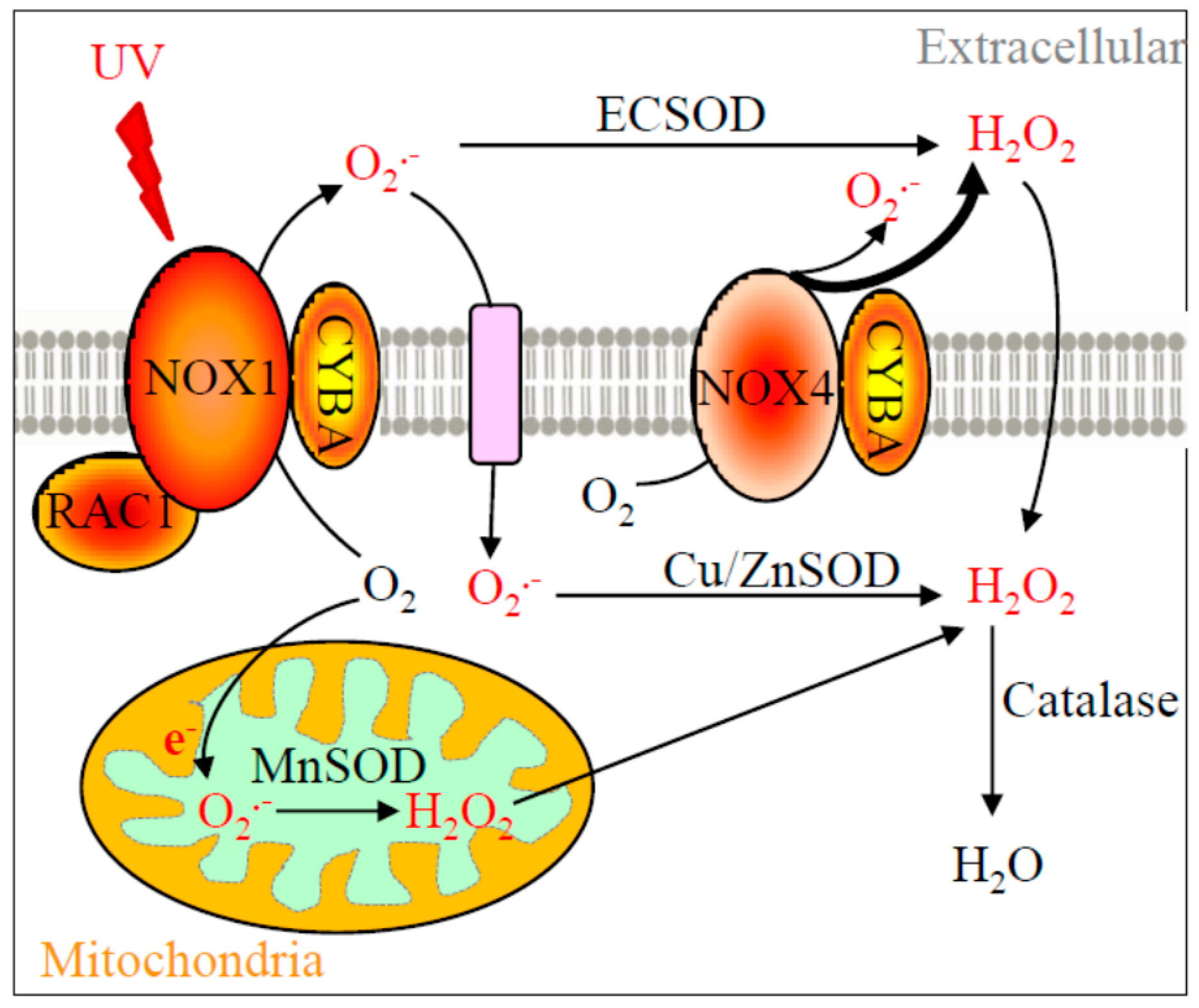
Recent understanding of melanoma photobiology has implied the etiological role of NOX enzymes, particularly NOX1 and NOX4 [
6,
7,
8]. NOX enzymes produce superoxide and/or hydrogen peroxide when coupled with CYBA (p22phox) membrane protein [
9]. RAC1, a newly defined melanoma oncogene [
10], is shown to enhance NOX1 activity [
11]. The downstream ROS metabolizing enzymes, e.g., copper-zinc superoxide dismutase (Cu-ZnSOD,
SOD1), manganese superoxide dismutase (MnSOD,
SOD2), and extracellular superoxide dismutase (ECSOD,
SOD3), convert superoxide to hydrogen peroxide. Catalase then transforms hydrogen peroxide to water molecules (
Figure 1). The cellular locations of NOX1, RAC1, NOX4, CYBA, and SOD enzymes, and their functions in ROS production and metabolism are illustrated in
Figure 1. Little is known about the comprehensive role of this entire pathway in melanoma formation. However, risk associated with these genes has been reported in various health conditions. For example, V16A variant in
SOD2 (rs4880) showed an impaired mitochondrial importing function and was associated with prostate cancer risk [
12]. The rs7277748 and rs4998557 variants in
SOD1 were found to be associated with amyotrophic lateral sclerosis [
13]. Variants rs2536512 and rs699473 in
SOD3 were linked to cerebral infarction [
14] and brain tumor [
15].
Although the causal network of melanoma has not yet been fully elucidated [
17], UV exposure is the most tangible environmental risk factor that can be readily modified by behavioral precautions [
18]. Therefore, the purpose of this study was to explore the relationship between the hypothesized photobiological pathway and risk of melanoma. Specifically, our aim was to use the candidate gene approach to discover the association of variations in the genetic profile of the redox enzymes with melanoma (
Figure 1). Building upon this rationale, functional genetic variants, namely single nucleotide polymorphisms (SNPs), were identified in this study with a priori chance of being associated with the risk of melanoma based on the following criteria: (1) not a well-known somatic mutation found in tumors with an established causality; (2) presented strong associations with many other health conditions in humans; and (3) with a potential to alter normal protein function based on the nucleotide substitution. For instance, variant rs8031 in
SOD2 was found to be associated with kidney complications in subjects with Type 1 diabetes [
19]. Variant rs10951982 in
RAC1 has been implied in the increased risk of hypertension [
20]. Even though rs10951982 in
RAC1 has not yet been reported in ROS-related malignancies, somatic mutations of
RAC1 (e.g.,
RAC1P29S) were found in 9.2% of sun-exposed melanoma tumors [
21,
22].
With this genetic profiling information in hand, we hope to lay a foundation to identify those individuals predisposed to UV exposure and risk of melanoma. This in turn will contribute to a better primary prevention strategy, such as earlier-life behavioral precautions. To the best of our knowledge, our work was the first to use a hypothesis-driven and pathway-based approach to study the association between genetic variations in the ROS pathway and risk of melanoma.
2. Results
2.1. Study Participants
Gender and age distributions of melanoma patients and healthy controls are listed in
Table 1. In total, 177 retrieved cases and 172 recruited controls were approximately matched for age groups and gender. Overall, there are higher percentages of female patients aged 19–39 (55.4%) and 40–59 (26.5%), while, at age 60 and older, there is a higher percentage of male patients (47.9%). This may reflect the actual sex ratios of melanoma incidence at different age groups [
23]. Of particular note, cases were retrieved from the international Genes, Environment, and Melanoma (GEM) study, which may not be strictly generalizable to a broader melanoma patient population.
SNP candidates and their currently known disease associations are listed in
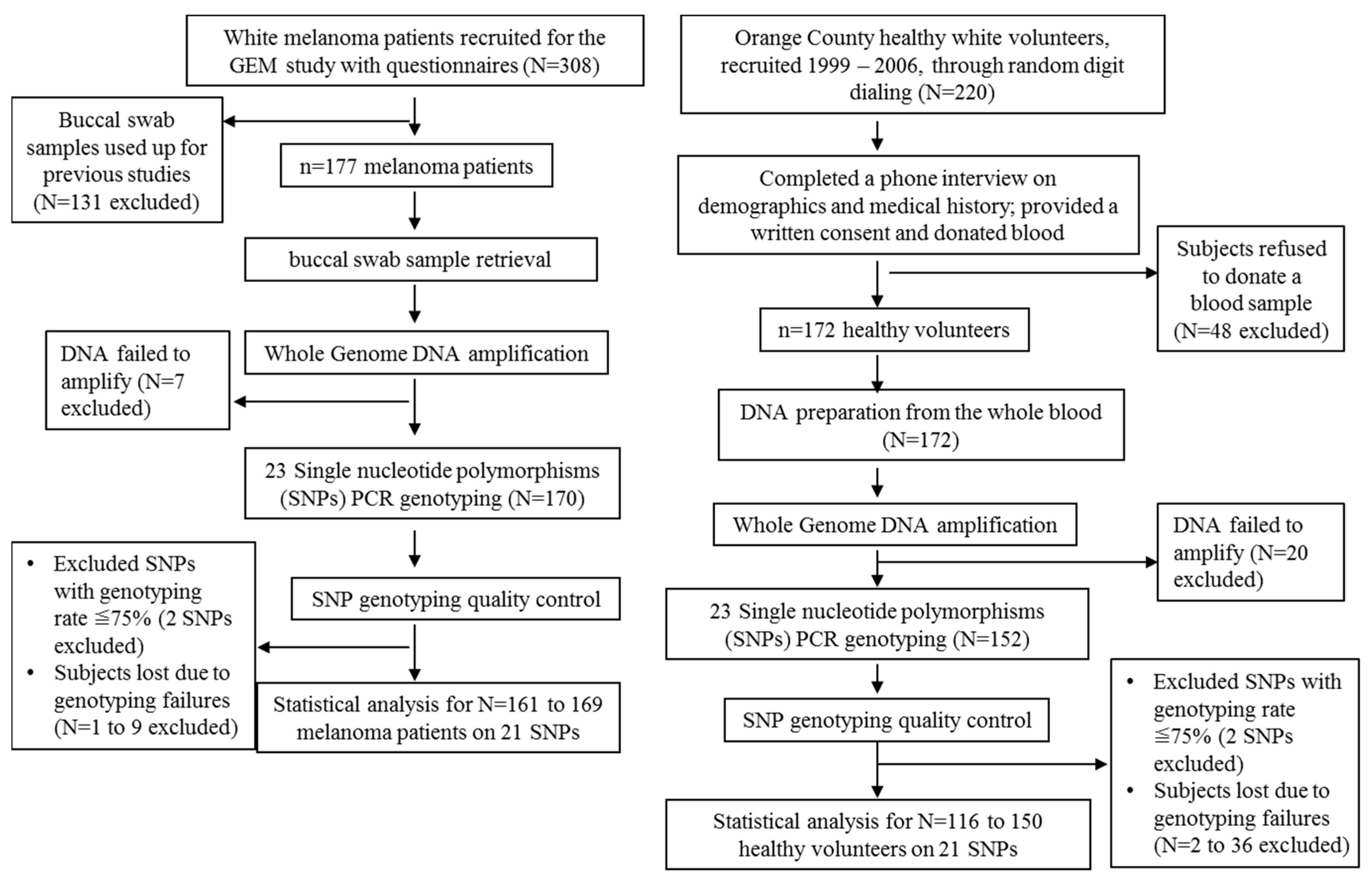
Table 2. Whole genome DNA amplification was successfully carried out in 322 study participants including 170 (96%) melanoma patients and 152 (88.4%) healthy controls (
Figure 2). However, for each SNP, there were different number of failed genotyping samples due to poor PCR reaction, and the overall successful genotyping rates were between 66.4% and 98.7% in the controls, and between 78.8% and 99.4% in the cases. SNPs with genotyping rate less than 75% on either arm (case or control group) of the participants were thus excluded from further analyses (SNPs rs13306296 and rs585197 were excluded,
Table 3). Ultimately, 161–169 melanoma patients, and 116–150 healthy controls remained to be further analyzed (
Figure 2).
2.2. SNP Associations
Chi-square or Fisher’s exact test of independence was performed to identify SNP frequency differences between melanoma patients and healthy controls under genotypic, allelic, recessive, and dominant SNP models (
Table 3). An exact test of genotype counts on Hardy–Weinberg equilibrium (HWE) was conducted to identify and exclude SNPs not in genotype balance in our study sample. Under the genotypic model, five SNPs exhibited statistically significant (
p < 0.05) frequency differences between cases and controls: rs10951982 (
RAC1), rs8031 (
SOD2), rs2536512 (
SOD3), rs4673 (
CYBA), and rs1049255 (
CYBA) (
Table 3). The allelic model only determined three of them as being significant: rs10951982 (
RAC1), rs8031 (
SOD2), and rs2536512 (
SOD3). These three alleles exhibited significance in the recessive model as well. In the dominant model, rs10951982 (
RAC1), rs4673 (
CYBA), and rs1049255 (
CYBA) showed significance. The rs1001179 (
CAT) showed a significant difference between cases and controls in the dominant and recessive models but the significance disappeared in the other two models.
2.3. Bivariate Logistic Regression Analyses
The top five SNPs identified from the genotypic model without HWE violations were fitted into bivariate logistic regressions with additive, recessive, and dominant allele models, respectively. The odds ratios of melanoma risk were calculated using the homozygous major allele genotype as the reference (
Table 4). Odds ratios derived from the regression models were compared to a corrected significance level at 0.00238 (0.05/21) to justify for multiple comparisons among the remaining 21 SNP candidates. Odds ratios with
p-values < 0.00238 were considered having statistical significance in the results.
In the additive allele model, carrying one copy of minor allele A in rs10951982 (RAC1) was significantly associated with a higher risk of melanoma (OR 8.98, 95% CI: 5.08, 16.44, p < 0.001), as compared to those who carried homozygous minor alleles AA (OR 8.23, 95% CI: 2.73, 28.39, p < 0.001). Dominant allele model further showed that combined minor allele copies (GA+AA) as compared to homozygous major alleles GG exhibited the highest risk of melanoma (OR 8.91, 95% CI: 5.09, 16.19, p < 0.001). A similar result was observed in rs4673 (CYBA), with one copy of the minor allele A exhibiting a higher risk of melanoma (OR 1.96, 95% CI: 1.23, 3.15, p = 0.005), and further confirmed in a dominant allele model (OR 1.84, 95% CI: 1.16, 2.92, p = 0.010). However, the p-values did not reach the corrected significance level of 0.00238.
The unadjusted odds of melanoma increased with homozygous minor allele T in rs1049255 (CYBA). TT exhibited an OR of 2.44 (95% CI: 1.27, 4.79, p = 0.008) in the additive model and an OR of 1.97 (95% CI: 1.10, 3.61, p = 0.022) in the recessive model. In both scenarios, p-values were greater than 0.00238, thus were non-significant because of the stringent Bonferroni correction for multiple comparison.
In contrast, homozygous minor allele genotypes at both rs8031 (SOD2) and rs2536512 (SOD3) exhibited significant association with a reduced risk of melanoma in the additive allele model, with 84% reduction in odds of melanoma (OR 0.16, 95% CI: 0.06, 0.39, p < 0.001) for rs8031 (SOD2), and 92% reduction in odds of melanoma (OR 0.08, 95% CI: 0.01, 0.31, p = 0.001) for rs2536512 (SOD3). Similar results were also observed in the recessive model, where an 85% reduction in the odds of melanoma (OR 0.15, 95% CI: 0.06, 0.33, p < 0.001) was observed for rs8031 (SOD2) with TT minor alleles, and a 91% reduction (OR 0.09, 95% CI: 0.01, 0.33, p = 0.002 with marginal significance) for rs2536512 (SOD3) with AA minor alleles.
2.4. Multivariate Logistic Regression Analyses
We continued to fit these top five SNPs into multivariate logistic regression under the three SNP models, controlling for major melanoma risk factors including gender, age at diagnosis, family history of melanoma, and lifetime ever-sunburned (
Table 5). After adjusting for these risk factors, rs1049255 (
CYBA), rs4673 (
CYBA), rs8031 (
SOD2), and rs2536512 (
SOD3) were no longer associated with melanoma risk in all three models (
p > 0.00238).
Consistent with what we have found in
Table 4, the most significant genotype was heterozygous GA genotype in rs10951982 (
RAC1), which exhibited an OR of 6.15 (95% CI: 2.98, 13.44,
p < 0.001) after controlling for other risk factors. This minor allele also showed a significant association with melanoma risk in the dominant model (OR 5.79, 95% CI: 2.84, 12.51,
p < 0.001). Similar results were also found for rs4673 (
CYBA) but with only marginal significance. Heterozygous GA genotype was associated with an increased risk of melanoma (OR 2.17, 95% CI: 1.17, 4.07,
p = 0.015), which was further confirmed in the dominant allele model (OR 1.88, 95% CI: 1.03, 3.47,
p = 0.042), although the
p-values did not reach the corrected significance level of 0.00238.
The homozygous minor allele TT genotype in rs8031 (SOD2) was found associated with a decreased risk of melanoma, with an OR of 0.32 (95% CI: 0.09, 0.94, p = 0.047) in the additive model, and an OR of 0.26 (95% CI: 0.08, 0.70, p = 0.011) in the recessive allele model, which indicated that homozygous minor alleles TT reduced the odds of melanoma by 74%, but neither of these results reached the universal significance level of 0.00238.
3. Discussion
After removal of SNP markers with high error rates during the assessment of genotyping quality, 21 SNP candidates remained to be eligible for the genetic association analysis. Eight SNPs showed significant association with melanoma but three of them were not in Hardy–Weinberg equilibrium, which may suggest that there are multiple alleles in the same locus, and we missed genotyping of other alleles. Therefore, only five SNP candidates showed genotypic significance and were further analyzed in regression models, including rs10951982 (
RAC1), rs1049255 (
CYBA), rs4673 (
CYBA), rs8031 (
SOD2), and rs2536512 (
SOD3). We corrected the universal
p-value to be compared with at 0.00238 (0.05/21, 21 SNPs being tested) to justify the multiple comparison issue in genetic association studies, using a Bonferroni approach [
48,
49]. The rs10951982 (
RAC1) and rs4673 (
CYBA) exhibited the highest increased risk of melanoma when presenting one copy of the minor allele in the unadjusted regression model, but rs4673 did not reach the universal significance level at 0.00238 in the multivariate regression model with adjustments for melanoma risk factors including age, sex, family history of melanoma, and lifetime ever-sunburned. Of particular note, a homozygous minor allele TT genotype in rs8031 (
SOD2) was found to be associated with reduced risk of melanoma in the bivariate regression, however significance was lost in the multivariate regression analyses.
SOD2 is known to be a major superoxide detoxifying enzyme of cells, and therefore an altered function or expression of this enzyme may lead to unbalanced redox homeostasis and thus potentially increase or decrease the risk of melanoma [
40]. Since SOD2 converts superoxide to hydrogen peroxide (
Figure 1), which belongs to a type of ROS, the function of SOD2 is thus double-edged. Our multivariate analysis indicated that homozygous TT allele in rs8031 reduced the risk of melanoma, but little is currently known about the molecular function of this variant. We suggest a lab-based functional molecular biology study to unravel the discrepancy between zygote expression and enzymatic activity in this particular SNP.
SNPs rs1049255 and rs4673 in
CYBA showed genotypic frequency differences between cases and controls in the unadjusted model (
Table 4), with more patients carrying higher copies of minor alleles in rs1049255. Variant rs4673 changes the amino acid at position 72 from a tyrosine to a histidine (Y72H) of the CYBA (p22phox) protein, which is frequently referred to a C242T variant in the literature [
50]. The T allele exhibited decreased dimerization with NOX and therefore may potentially reduce NOX activity and cellular ROS level [
32]. In fact, the CT and TT genotype showed lower NADPH oxidase activity in hypertensive patients as compared with CC genotype [
51]. However, opposite observation was also reported, where the CT genotype and T allele are associated with higher risk of coronary artery disease [
52]. In our study, the CT and TT (GA and AA) showed higher risk for melanoma as compared to CC (GG) allele (the dominant model in
Table 4 and
Table 5). This observation needs further validation. Variant rs1049255 is located in the 3′ untranslated region (3′ UTR) of the
CYBA gene. Although the molecular function of this SNP is unknown, current understanding of 3′ UTR is an important miRNA binding site, and SNPs located in this region might have the potential to regulate mRNA stability and translation efficiency [
53,
54].
RAC1-GTPase is an NOX1 activator which promotes binding of NOX1 with its subunits and forms the complete enzyme complex [
55,
56,
57]. NOX1 was one of the first cellular molecules found to be directly regulated by RAC1 in the phagocytic process [
58,
59,
60]. However, SNP rs10951982 in
RAC1 alone has not been reported in any ROS-related activities thus far. Information on the function of this locus and its association with any malignancy is limited in the current literature. Nevertheless, this variant has been reported to be associated with over-reactive immune diseases and an increased risk of hypertension [
20,
35,
36,
61]. Considering that
CYBA variants have been widely studied in cardiovascular diseases, including coronary heart disease [
34] and hypertension [
33], which are tightly associated with increased levels of ROS,
RAC1 rs10951982 may also play a part in inducing oxidative stress. Since rs10951982 is the most significant variant in our current study, and in lieu of its function in immune diseases as well as a potential role in NOX1-induced oxidative stress, our discovery might not only suggest an inflammatory microenvironment created by RAC1 that is in favor of melanoma progression [
62], but also indicate an elevation of ROS level via RAC1 in melanoma etiology. In addition, RAC1 is also a crucial kinase in the NRAS and PI3K pathway [
63], both of which are key melanoma oncogenic pathways. Therefore, it is possible that RAC1 plays a non-ROS role and impacts these other oncogenic pathways.
Overall, of the three significant SNPs after adjustment against age, sex, family history and life time sun burn history, the minor allele of
RAC1 rs10951982 (the A allele) showed a consistent role with an increase ROS and thus increased melanoma risk. The minor allele of rs4673 (the A allele) was reported controversial role in ROS association [
51,
64], it may exhibit certain cell-specific effects. In our study, the minor allele showed higher risk for melanoma in a dominant model. The minor allele of rs8031 (the T allele) exhibited a protective role against melanoma risk in a recessive model. It is unclear how this allele modifies ROS levels. Based on our results, the T allele can be associated with either increased or decreased SOD activities as SOD2 is double-edged and can play dual roles in ROS metabolism.
Of particular note, in our regression models, we applied the most common ways of disease transmission, namely additive, recessive, and dominant modes, in our analyses. This was because we did not want to make any assumptions of the disease transmission modes. According to Sham and Purcell [
49], a test that assumed additive effects would have greater power than a test that also allowed dominance, if the true effects at the locus were indeed additive and did not show dominance. Conversely, if the underlying causal variant was recessive, then power would be lost by carrying out an analysis that assumed additively. If there was uncertainty regarding the true pattern of effects at a locus, then it might be appropriate to use several statistical tests to ensure adequate statistical power for all possible scenarios. We therefore included results from these additional models that may provide more information and maintain statistical power as well. Although the covariates were not presented as part of the results in our tables, family history of melanoma and lifetime ever-sunburned controlled in the multivariate models consistently showed statistical significance, whereas sex and age did not. Family history of melanoma [
23], along with fair skin, light hair and eye color are known melanoma genetic risk factors, whereas the levels of sun exposure including sunburns and moles or freckles are important environmental risk factors for melanoma [
65]. The statistical significance of the covariates might indicate a mediating role in our primary study interest, from the susceptible familial genetic makeup of these participants, as well as the behavior or attitude towards sun exposure that resulted in getting sunburns or freckles.
Our study had a few limitations. First, the small sample size does not always provide sufficient power [
66]. Second, by the experimental design, we could only genotype two alleles. Therefore, loci with multiple alleles may not show HWE and must be excluded for analysis. Third, our study participants included only those white individuals from the southern California area, and therefore a loss of generalizability to the broader white population might be expected. Last, a common limitation of case-control studies is that the results provide only an association with risk, but they are not necessarily connected to causality. Replicating findings from another dataset is a common strategy to validate the results identified in our current study. However, even with the most stringent statistical design, SNP findings are usually hard to replicate [
48,
49]. Multiple reasons are considered, such as there are still unknown and uncontrolled confounders, multiple comparisons only lead to chance findings, the gene and environment interaction is not easy to account for, and the target allele is in linkage disequilibrium with the identified allele but the chance finding failed to locate the target allele and thus make replication difficult to achieve. Nevertheless, we will still validate our findings in a separate dataset in our next study, as our ultimate goal is to develop useful markers in prevention.
To conclude, our initial analyses revealed an increased risk of melanoma associated with rs10951982 (RAC1), and a decreased risk associated with rs8031 (SOD2). Multivariate analyses further confirmed the association of an increased risk of melanoma with rs10951982 (RAC1). Our results highlighted the importance of RAC1 enzyme and cellular oxidation-metabolizing efficiency controlled by SOD2 in association with ROS-mediated risk of melanoma. We suggest that these results shall be further validated with the goal of designing novel screening targets to identify highly UV-susceptible individuals, particularly in the RAC1 and SOD2 genes, in order to take the melanoma primary prevention strategy to a precision level.

 ,
, 


{kind=link}
{kind=link}