HPV16-Related Cervical Cancers and Precancers Have Increased Levels of Host Cell DNA Methylation in Women Living with HIV

 ,
,
Abstract
:1. Introduction
2. Results
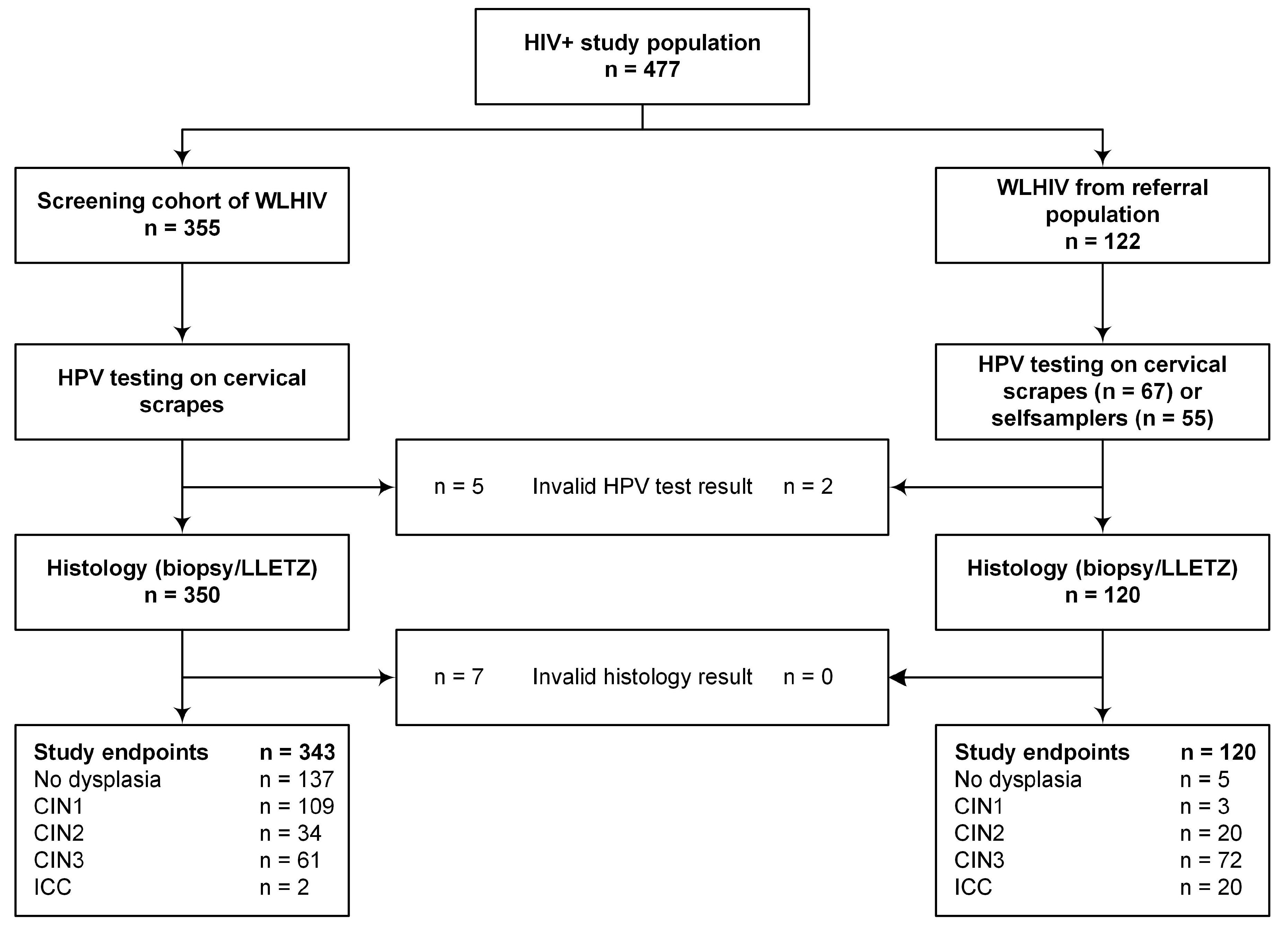
2.1. Study Population
2.2. HPV Prevalence: Overall and Type-Specific
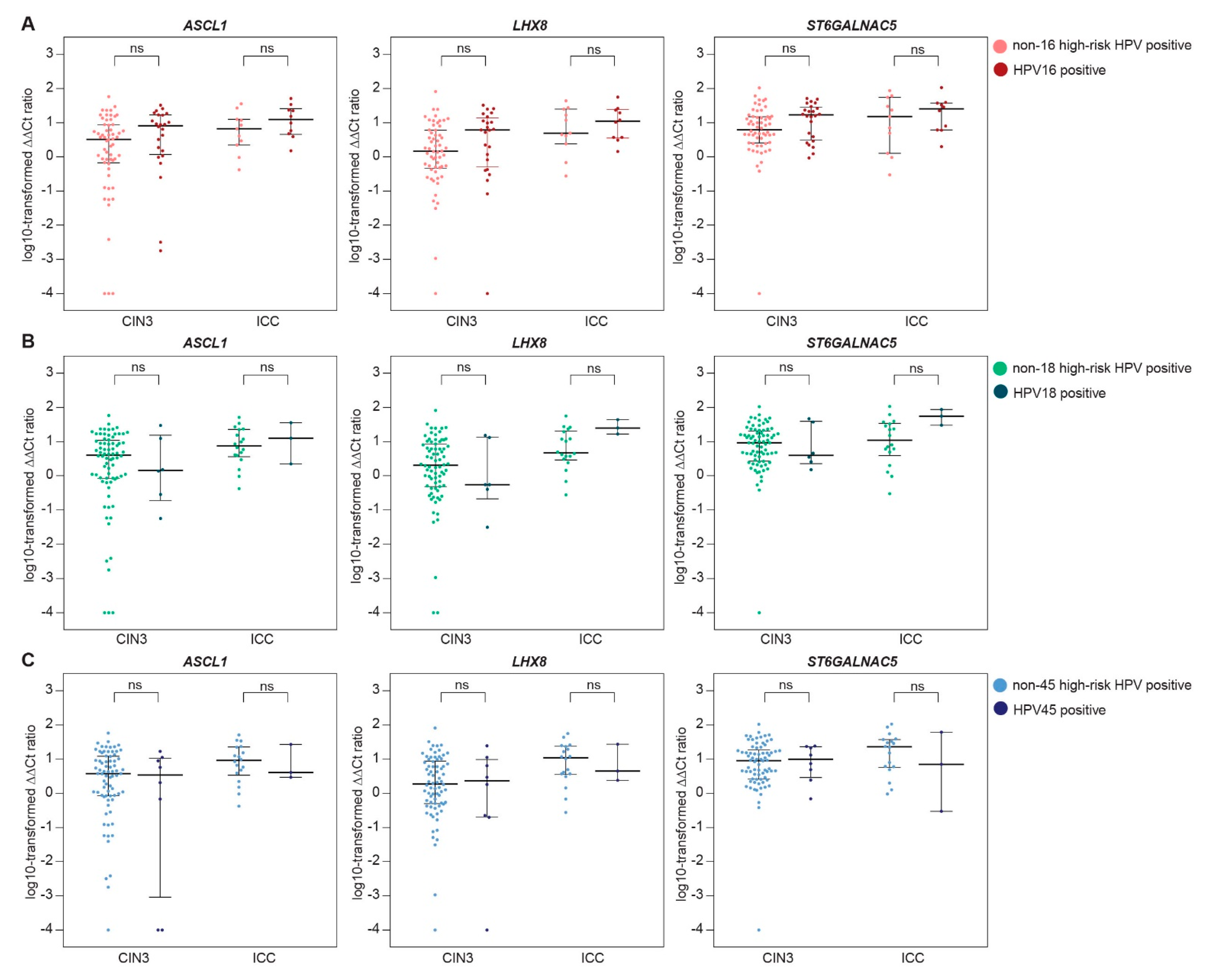
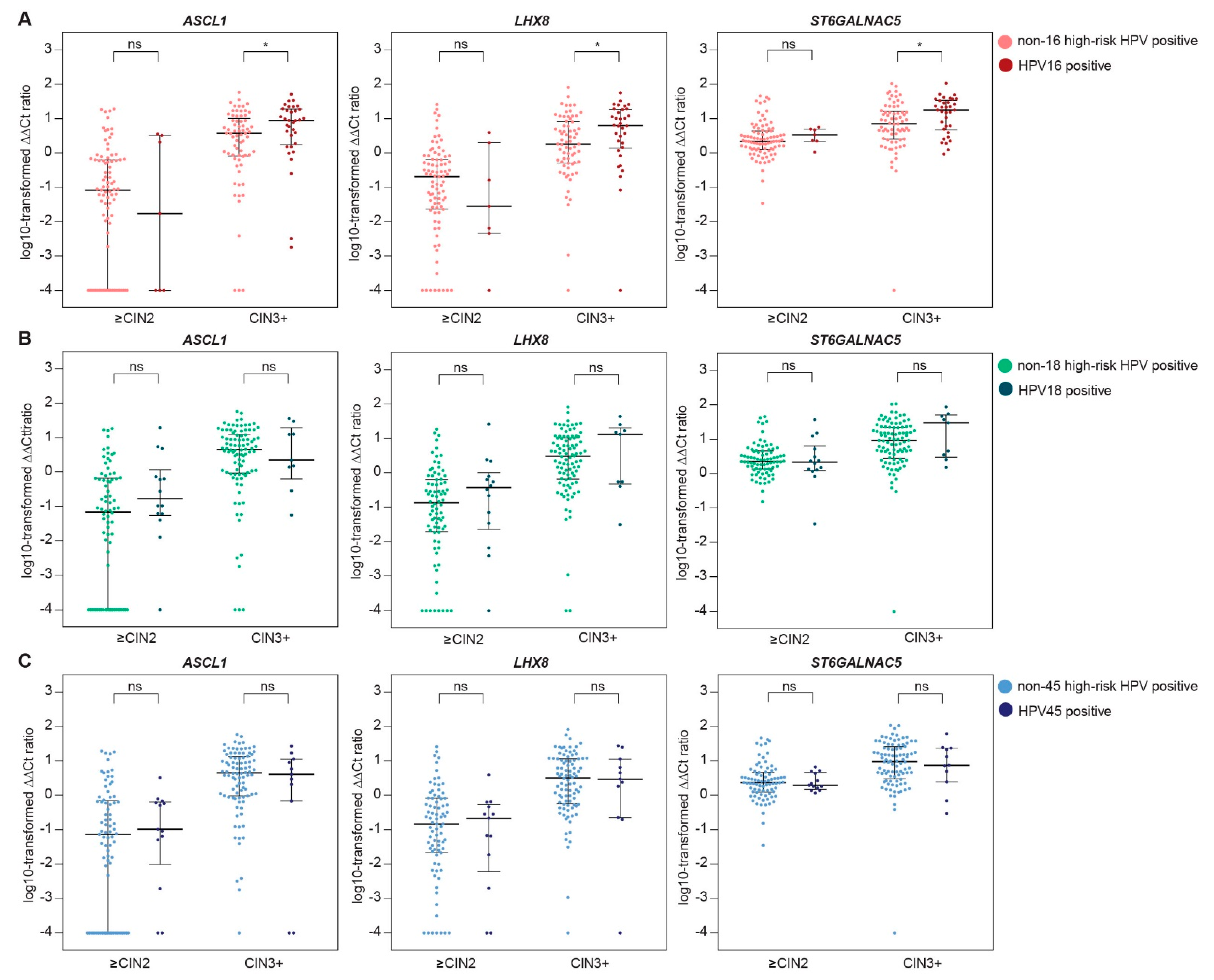
2.3. Methylation Levels Across HPV Type-Specific Disease Categories
3. Discussion
4. Methods
4.1. Study Population and Specimen Collection
4.2. High-Risk HPV Testing
4.3. Methylation Analysis
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Sanjose, S.; Diaz, M.; Castellsague, X.; Clifford, G.; Bruni, L.; Munoz, N.; Bosch, F.X. Worldwide prevalence and genotype distribution of cervical human papillomavirus DNA in women with normal cytology: A meta-analysis. Lancet Infect. Dis. 2007, 7, 453–459. [Google Scholar] [CrossRef]
- Munoz, N.; Bosch, F.X.; de Sanjose, S.; Tafur, L.; Izarzugaza, I.; Gili, M.; Viladiu, P.; Navarro, C.; Martos, C.; Ascunce, N.; et al. The causal link between human papillomavirus and invasive cervical cancer: A population-based case-control study in Colombia and Spain. Int. J. Cancer 1992, 52, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.; Peto, J.; Meijer, C.J.; Munoz, N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Bosch, F.X.; Lorincz, A.; Munoz, N.; Meijer, C.J.; Shah, K.V. The causal relation between human papillomavirus and cervical cancer. J. Clin. Pathol. 2002, 55, 244–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Biological agents. Volume 100 B. A review of human carcinogens. IARC Monogr. Eval. Carcinog. Risks Hum. 2012, 100, 1–441. [Google Scholar]
- De Sanjose, S.; Quint, W.G.V.; Alemany, L.; Geraets, D.T.; Klaustermeier, J.E.; Lloveras, B.; Tous, S.; Felix, A.; Bravo, L.E.; Shin, H.-R.; et al. Human papillomavirus genotype attribution in invasive cervical cancer: A retrospective cross-sectional worldwide study. Lancet Oncol. 2010, 11, 1048–1056. [Google Scholar] [CrossRef]
- Guan, P.; Howell-Jones, R.; Li, N.; Bruni, L.; de Sanjose, S.; Franceschi, S.; Clifford, G.M. Human papillomavirus types in 115,789 HPV-positive women: A meta-analysis from cervical infection to cancer. Int. J. Cancer 2012, 131, 2349–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifford, G.M.; Goncalves, M.A.; Franceschi, S.; HPV and HIV Study Group. Human papillomavirus types among women infected with HIV: A meta-analysis. AIDS 2006, 20, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
- Strickler, H.D.; Palefsky, J.M.; Shah, K.V.; Anastos, K.; Klein, R.S.; Minkoff, H.; Duerr, A.; Massad, L.S.; Celentano, D.D.; Hall, C.; et al. Human papillomavirus type 16 and immune status in human immunodeficiency virus-seropositive women. J. Natl. Cancer Inst. 2003, 95, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Anastos, K.; Hoover, D.R.; Burk, R.D.; Cajigas, A.; Shi, Q.; Singh, D.K.; Cohen, M.H.; Mutimura, E.; Sturgis, C.; Banzhaf, W.C.; et al. Risk factors for cervical precancer and cancer in HIV-infected, HPV-positive Rwandan women. PLoS ONE 2010, 5, e13525. [Google Scholar] [CrossRef] [PubMed]
- Palefsky, J. Human papillomavirus-related disease in people with HIV. Curr. Opin. HIV AIDS 2009, 4, 52–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.W.; Kuhn, L.; Ellerbrock, T.V.; Chiasson, M.A.; Bush, T.J.; Wright, T.C., Jr. Human papillomavirus infection in women infected with the human immunodeficiency virus. N. Engl. J. Med. 1997, 337, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Massad, L.S.; Xie, X.; D’Souza, G.; Darragh, T.M.; Minkoff, H.; Wright, R.; Colie, C.; Sanchez-Keeland, L.; Strickler, H.D. Incidence of cervical precancers among HIV-seropositive women. Am. J. Obstet. Gynecol. 2015, 212, 606.e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Clifford, G.M.; Franceschi, S.; Keiser, O.; Schoni-Affolter, F.; Lise, M.; Dehler, S.; Levi, F.; Mousavi, M.; Bouchardy, C.; Wolfensberger, A.; et al. Immunodeficiency and the risk of cervical intraepithelial neoplasia 2/3 and cervical cancer: A nested case-control study in the Swiss HIV cohort study. Int. J. Cancer 2016, 138, 1732–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denslow, S.A.; Rositch, A.F.; Firnhaber, C.; Ting, J.; Smith, J.S. Incidence and progression of cervical lesions in women with HIV: A systematic global review. Int. J. STD AIDS 2014, 25, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Madeleine, M.M.; Biggar, R.J.; Engels, E.A. Risk of human papillomavirus-associated cancers among persons with AIDS. J. Natl. Cancer Inst. 2009, 101, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Grulich, A.E.; van Leeuwen, M.T.; Falster, M.O.; Vajdic, C.M. Incidence of cancers in people with HIV/AIDS compared with immunosuppressed transplant recipients: A meta-analysis. Lancet 2007, 370, 59–67. [Google Scholar] [CrossRef]
- Clifford, G.M.; de Vuyst, H.; Tenet, V.; Plummer, M.; Tully, S.; Franceschi, S. Effect of HIV Infection on Human Papillomavirus Types Causing Invasive Cervical Cancer in Africa. J. Acquir. Immune Defic. Syndr. 2016, 73, 332–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, H.; Alemany, L.; Lacey, C.; Chibwesha, C.J.; Sahasrabuddhe, V.; Banura, C.; Denny, L.; Parham, G.P. The burden of human papillomavirus infections and related diseases in sub-saharan Africa. Vaccine 2013, 31 (Suppl. 5), F32–F46. [Google Scholar] [CrossRef] [PubMed]
- Kuss-Duerkop, S.K.; Westrich, J.A.; Pyeon, D. DNA Tumor Virus Regulation of Host DNA Methylation and Its Implications for Immune Evasion and Oncogenesis. Viruses 2018, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, R.D.; Snijders, P.J.; Heideman, D.A.; Meijer, C.J. Clinical implications of (epi)genetic changes in HPV-induced cervical precancerous lesions. Nat. Rev. Cancer 2014, 14, 395–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wentzensen, N.; Sherman, M.E.; Schiffman, M.; Wang, S.S. Utility of methylation markers in cervical cancer early detection: Appraisal of the state-of-the-science. Gynecol. Oncol. 2009, 112, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, A.; Sasai, N.; Nakao, M.; Defossez, P.A. The role of methyl-binding proteins in chromatin organization and epigenome maintenance. Brief Funct. Genom. 2012, 11, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Molano, M.; Moreno-Acosta, P.; Morales, N.; Burgos, M.; Buitrago, L.; Gamboa, O.; Alvarez, R.; Garland, S.M.; Tabrizi, S.N.; Steenbergen, R.D.; et al. Association Between Type-specific HPV Infections and hTERT DNA Methylation in Patients with Invasive Cervical Cancer. Cancer Genom. Proteom. 2016, 13, 483–491. [Google Scholar] [CrossRef]
- Schutze, D.M.; Kooter, J.M.; Wilting, S.M.; Meijer, C.J.; Quint, W.; Snijders, P.J.; Steenbergen, R.D. Longitudinal assessment of DNA methylation changes during HPVE6E7-induced immortalization of primary keratinocytes. Epigenetics 2015, 10, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlaat, W.; Snoek, B.C.; Heideman, D.A.M.; Wilting, S.M.; Snijders, P.J.F.; Novianti, P.W.; van Splunter, A.P.; Peeters, C.F.W.; van Trommel, N.E.; Massuger, L.; et al. Identification and validation of a 3-gene methylation classifier for HPV-based cervical screening on self-samples. Clin. Cancer Res. 2018, 24, 3456–3464. [Google Scholar] [CrossRef] [PubMed]
- Kremer, W.W.; Van Zummeren, M.; Novianti, P.W.; Richter, K.L.; Verlaat, W.; Snijders, P.J.; Heideman, D.A.; Steenbergen, R.D.; Dreyer, G.; Meijer, C.J. Detection of hypermethylated genes as markers for cervical screening in women living with HIV. J. Int. AIDS Soc. 2018, 21, e25165. [Google Scholar] [CrossRef] [PubMed]
- Clifford, G.M.; Tully, S.; Franceschi, S. Carcinogenicity of Human Papillomavirus (HPV) Types in HIV-Positive Women: A Meta-Analysis From HPV Infection to Cervical Cancer. Clin. Infect. Dis. 2017, 64, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Bulk, S.; Berkhof, J.; Bulkmans, N.W.; Zielinski, G.D.; Rozendaal, L.; van Kemenade, F.J.; Snijders, P.J.; Meijer, C.J. Preferential risk of HPV16 for squamous cell carcinoma and of HPV18 for adenocarcinoma of the cervix compared to women with normal cytology in The Netherlands. Br. J. Cancer 2006, 94, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Bierkens, M.; Hesselink, A.T.; Meijer, C.J.; Heideman, D.A.; Wisman, G.B.; van der Zee, A.G.; Snijders, P.J.; Steenbergen, R.D. CADM1 and MAL promoter methylation levels in hrHPV-positive cervical scrapes increase proportional to degree and duration of underlying cervical disease. Int. J. Cancer 2013, 133, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Van Zummeren, M.; Kremer, W.W.; Van Aardt, M.C.; Breytenbach, E.; Richter, K.L.; Rozendaal, L.; Witte, B.I.; De Strooper, L.M.A.; Hesselink, A.T.; Heideman, D.A.M.; et al. Selection of women at risk for cervical cancer in an HIV-infected South African population. AIDS 2017, 31, 1945–1953. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, L.M.; Meijer, C.J.; Berkhof, J.; Hesselink, A.T.; Snijders, P.J.; Steenbergen, R.D.; Heideman, D.A. Methylation analysis of the FAM19A4 gene in cervical scrapes is highly efficient in detecting cervical carcinomas and advanced CIN2/3 lesions. Cancer Prev. Res. (Phila.) 2014, 7, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, L.M.; van Zummeren, M.; Steenbergen, R.D.; Bleeker, M.C.; Hesselink, A.T.; Wisman, G.B.; Snijders, P.J.; Heideman, D.A.; Meijer, C.J. CADM1, MAL and miR124-2 methylation analysis in cervical scrapes to detect cervical and endometrial cancer. J. Clin. Pathol. 2014, 67, 1067–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boers, A.; Wang, R.; van Leeuwen, R.W.; Klip, H.G.; de Bock, G.H.; Hollema, H.; van Criekinge, W.; de Meyer, T.; Denil, S.; van der Zee, A.G.; et al. Discovery of new methylation markers to improve screening for cervical intraepithelial neoplasia grade 2/3. Clin. Epigenet. 2016, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Eijsink, J.J.; Lendvai, A.; Volders, H.H.; Klip, H.; Buikema, H.J.; van Hemel, B.M.; Schuuring, E.; van der Zee, A.G.; Wisman, G.B. Methylation markers for CCNA1 and C13ORF18 are strongly associated with high-grade cervical intraepithelial neoplasia and cervical cancer in cervical scrapings. Cancer Epidemiol. Biomark. Prev. 2009, 18, 3000–3007. [Google Scholar] [CrossRef] [PubMed]
- Kelly, H.A.; Chikandiwa, A.; Warman, R.; Segondy, M.; Sawadogo, B.; Vasiljevic, N.; Didelot, M.N.; Meda, N.; Weiss, H.A.; Delany-Moretlwe, S.; et al. Associations of human gene EPB41L3 DNA methylation and cervical intraepithelial neoplasia in women living with HIV-1 in Africa. AIDS 2018, 32, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, H.; Franceschi, S.; Plummer, M.; Mugo, N.R.; Sakr, S.R.; Meijer, C.J.; Heideman, D.A.; Tenet, V.; Snijders, P.J.; Hesselink, A.T.; et al. Methylation Levels of CADM1, MAL, and MIR124-2 in Cervical Scrapes for Triage of HIV-Infected, High-Risk HPV-Positive Women in Kenya. J. Acquir. Immune Defic. Syndr. 2015, 70, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nye, M.D.; Hoyo, C.; Huang, Z.; Vidal, A.C.; Wang, F.; Overcash, F.; Smith, J.S.; Vasquez, B.; Hernandez, B.; Swai, B.; et al. Associations between methylation of paternally expressed gene 3 (PEG3), cervical intraepithelial neoplasia and invasive cervical cancer. PLoS ONE 2013, 8, e56325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, A.C.; Henry, N.M.; Murphy, S.K.; Oneko, O.; Nye, M.; Bartlett, J.A.; Overcash, F.; Huang, Z.; Wang, F.; Mlay, P.; et al. PEG1/MEST and IGF2 DNA methylation in CIN and in cervical cancer. Clin. Transl. Oncol. 2014, 16, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Westrich, J.A.; Warren, C.J.; Pyeon, D. Evasion of host immune defenses by human papillomavirus. Virus Res. 2017, 231, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Cicchini, L.; Blumhagen, R.Z.; Westrich, J.A.; Myers, M.E.; Warren, C.J.; Siska, C.; Raben, D.; Kechris, K.J.; Pyeon, D. High-Risk Human Papillomavirus E7 Alters Host DNA Methylome and Represses HLA-E Expression in Human Keratinocytes. Sci. Rep. 2017, 7, 3633. [Google Scholar] [CrossRef] [PubMed]
- Cicchini, L.; Westrich, J.A.; Xu, T.; Vermeer, D.W.; Berger, J.N.; Clambey, E.T.; Lee, D.; Song, J.I.; Lambert, P.F.; Greer, R.O.; et al. Suppression of Antitumor Immune Responses by Human Papillomavirus through Epigenetic Downregulation of CXCL14. MBio 2016, 7, e00270-16. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Hsiao, Y.H.; Lai, Y.H.; Chen, Y.C.; Chen, Y.J.; Chou, J.L.; Chan, M.W.; Lin, Y.H.; Tsou, Y.A.; Tsai, M.-H.; et al. DNA methylation profiles and biomarkers of oral squamous cell carcinoma. Epigenetics 2015, 10, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, B.; Yao, B.; Li, J.L.; Fields, C.R.; Delmas, A.L.; Liu, C.; Robertson, K.D. DNMT1 and DNMT3B modulate distinct polycomb-mediated histone modifications in colon cancer. Cancer Res. 2009, 69, 7412–7421. [Google Scholar] [CrossRef] [PubMed]
- Oster, B.; Thorsen, K.; Lamy, P.; Wojdacz, T.K.; Hansen, L.L.; Birkenkamp-Demtroder, K.; Sorensen, K.D.; Laurberg, S.; Orntoft, T.F.; Andersen, C.L. Identification and validation of highly frequent CpG island hypermethylation in colorectal adenomas and carcinomas. Int. J. Cancer 2011, 129, 2855–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tommasi, S.; Karm, D.L.; Wu, X.; Yen, Y.; Pfeifer, G.P. Methylation of homeobox genes is a frequent and early epigenetic event in breast cancer. Breast Cancer Res. 2009, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Bos, P.D.; Zhang, X.H.; Nadal, C.; Shu, W.; Gomis, R.R.; Nguyen, D.X.; Minn, A.J.; van de Vijver, M.J.; Gerald, W.L.; Foekens, J.A.; et al. Genes that mediate breast cancer metastasis to the brain. Nature 2009, 459, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Park, N.I.; Guilhamon, P.; Desai, K.; McAdam, R.F.; Langille, E.; O’Connor, M.; Lan, X.; Whetstone, H.; Coutinho, F.J.; Vanner, R.J.; et al. ASCL1 Reorganizes Chromatin to Direct Neuronal Fate and Suppress Tumorigenicity of Glioblastoma Stem Cells. Cell Stem Cell 2017, 21, 411. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Du, X.; Zhang, S.; Wang, Q.; Yin, Y.; Qiu, X.; Da, P.; Yue, H.; Wu, H.; Xu, F. Achaete-scute complex homologue-1 promotes development of laryngocarcinoma via facilitating the epithelial-mesenchymal transformation. Tumour Biol. 2017, 39, 1010428317705752. [Google Scholar] [CrossRef] [PubMed]
- Donakonda, S.; Sinha, S.; Dighe, S.N.; Rao, M.R.S. System analysis identifies distinct and common functional networks governed by transcription factor ASCL1, in glioma and small cell lung cancer. Mol. Biosyst. 2017, 13, 1481–1494. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.C.; Ronnett, B.M.; Kurman, R.J.; Ferenczy, A. Precancerous Lesions of the Cervix. In Blaustein’s Pathology of the Female Genital Tract, 6th ed.; Kurman, R.J., Hedrick Ellenson, L., Ronnett, B.M., Eds.; Springer: New York, NY, USA, 2011; pp. 194–252. [Google Scholar]
- Van den Brule, A.J.; Pol, R.; Fransen-Daalmeijer, N.; Schouls, L.M.; Meijer, C.J.; Snijders, P.J. GP5+/6+ PCR followed by reverse line blot analysis enables rapid and high-throughput identification of human papillomavirus genotypes. J. Clin. Microbiol. 2002, 40, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Polman, N.J.; Ostrbenk, A.; Xu, L.; Snijders, P.J.F.; Meijer, C.; Poljak, M.; Heideman, D.A.M.; Arbyn, M. Evaluation of the Clinical Performance of the HPV-Risk Assay Using the VALGENT-3 Panel. J. Clin. Microbiol. 2017, 55, 3544–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesselink, A.T.; Berkhof, J.; van der Salm, M.L.; van Splunter, A.P.; Geelen, T.H.; van Kemenade, F.J.; Bleeker, M.G.; Heideman, D.A. Clinical validation of the HPV-risk assay, a novel real-time PCR assay for detection of high-risk human papillomavirus DNA by targeting the E7 region. J. Clin. Microbiol. 2014, 52, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Bravo, I.G.; Snijders, P.J.; Gissmann, L.; Pawlita, M.; Waterboer, T. Bead-based multiplex genotyping of human papillomaviruses. J. Clin. Microbiol. 2006, 44, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| HPV-Negative | HPV-Positive | |||||||
|---|---|---|---|---|---|---|---|---|
| Any * | Single | Multiple | ||||||
| n | % | n | % | n | % | n | % | |
| ≤CIN1 (n = 254) | 183 | 72.0% | 71 | 28.0% | 55 | 77.5% | 16 | 22.5% |
| CIN2 (n = 54) | 12 | 22.2% | 42 | 77.8% | 23 | 54.8% | 19 | 45.2% |
| CIN3 (n = 133) | 10 | 7.5% | 123 | 92.5% | 83 | 67.5% | 40 | 32.5% |
| ICC (n = 22) | 0 | 0.0% | 22 | 100% | 19 | 86.4% | 3 | 13.6% |
| ≤CIN1 n = 71 | CIN2 n = 42 | CIN3 n = 123 | ICC n = 22 | Total n = 258 | CIN3:≤CIN1 | ICC:≤CIN1 | ICC:CIN3 | CIN3 vs. ≤CIN1 | ICC vs. ≤CIN1 | ICC vs. CIN3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type | n | % | n | % | n | % | n | % | N | % | PR | PR | PR | OR (95% CI) | OR (95% CI) | OR (95% CI) | |||
| Overall * | |||||||||||||||||||
| 16 | 7 | 9.9% | 6 | 14.3% | 39 | 31.7% | 10 | 45.5% | 62 | 24.0% | 3.22 | 4.61 | 1.43 | 4.37 | (1.81–10.58) | 7.36 | (2.33–23.21) | 1.62 | (0.62–4.27) |
| 18 | 8 | 11.3% | 8 | 19.0% | 11 | 8.9% | 3 | 13.6% | 30 | 11.6% | 0.79 | 1.21 | 1.52 | - | 1.10 | (0.25–4.79) | 1.83 | (0.38–8.75) | |
| 31 | 7 | 9.9% | 7 | 16.7% | 11 | 8.9% | 2 | 9.1% | 27 | 10.5% | 0.91 | 0.92 | 1.02 | - | 1.04 | (0.19–5.59) | 1.37 | (0.26–7.15) | |
| 33 | 5 | 7.0% | 3 | 7.1% | 14 | 11.4% | 1 | 4.5% | 23 | 8.9% | 1.62 | 0.65 | 0.40 | 1.94 | (0.34–5.89) | - | - | ||
| 35 | 6 | 8.5% | 9 | 21.4% | 14 | 11.4% | 1 | 4.5% | 30 | 11.6% | 1.35 | 0.54 | 0.40 | 1.83 | (0.65–5.15) | - | - | ||
| 39 | 2 | 2.8% | 1 | 2.4% | 2 | 1.6% | - | - | 5 | 1.9% | 0.58 | - | - | - | - | - | |||
| 45 | 10 | 14.1% | 6 | 14.3% | 14 | 11.4% | 3 | 13.6% | 33 | 12.8% | 0.81 | 0.97 | 1.20 | - | - | 1.25 | (0.33–4.78) | ||
| 51 | 1 | 1.4% | 5 | 11.9% | 5 | 4.1% | - | - | 11 | 4.3% | 2.89 | - | - | 2.88 | (0.31–27.06) | - | - | ||
| 52 | 12 | 16.9% | 5 | 11.9% | 17 | 13.8% | 2 | 9.1% | 36 | 14.0% | 0.82 | 0.54 | 0.66 | - | - | - | |||
| 56 | 10 | 14.1% | 4 | 9.5% | 11 | 8.9% | 1 | 4.5% | 26 | 10.1% | 0.63 | 0.32 | 0.51 | - | - | - | |||
| 58 | 3 | 4.2% | 4 | 9.5% | 14 | 11.4% | - | - | 21 | 8.1% | 2.69 | - | - | 2.73 | (0.75–9.95) | - | - | ||
| 59 | 4 | 5.6% | 3 | 7.1% | 5 | 4.1% | - | - | 12 | 4.7% | 0.72 | - | - | - | - | - | |||
| 66 | 4 | 5.6% | 3 | 7.1% | 14 | 11.4% | 2 | 9.1% | 23 | 8.9% | 2.02 | 1.61 | 0.80 | 1.96 | (0.61–6.35) | 1.63 | (0.24–11.07) | - | |
| 68 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |||
| X | 9 | 12.7% | 4 | 9.5% | 11 | 8.9% | - | - | 24 | 9.3% | 0.71 | - | - | - | - | - | |||
| Single | n = 55 | n = 23 | n = 83 | n = 19 | n = 180 | ||||||||||||||
| 16 | 4 | 7.3% | 1 | 4.3% | 20 | 24.1% | 10 | 52.6% | 35 | 7.6% | 3.31 | 7.24 | 2.18 | 3.79 | (1.19–12.09) | 11.4 | (3.10–41.90) | 3.01 | (1.06–8.55) |
| 18 | 7 | 12.7% | 3 | 13.0% | 4 | 4.8% | 3 | 15.8% | 17 | 3.7% | 0.38 | 1.24 | 3.28 | - | 1.02 | (0.21–4.90) | 4.36 | (0.56–34.0) | |
| 31 | 4 | 7.3% | 2 | 8.7% | 4 | 4.8% | 1 | 5.3% | 11 | 2.4% | 0.66 | 0.72 | 1.09 | - | - | 1.03 | (0.09–12.31) | ||
| 33 | 2 | 3.6% | - | - | 7 | 8.4% | 1 | 5.3% | 10 | 2.2% | 2.32 | 1.45 | 0.62 | 2.85 | (0.54–15.08) | 1.25 | (0.09–17.45) | - | |
| 35 | 2 | 3.6% | 6 | 26.1% | 8 | 9.6% | 0 | - | 16 | 3.5% | 2.65 | - | - | 3.78 | (0.74–19.32) | - | - | ||
| 39 | 1 | 1.8% | - | - | 1 | 1.2% | 0 | - | 2 | 0.4% | 0.66 | - | - | - | - | - | |||
| 45 | 7 | 12.7% | 1 | 4.3% | 5 | 6.0% | 3 | 15.8% | 16 | 3.5% | 0.47 | 1.24 | 2.62 | - | 1.11 | (0.26–4.65) | 2.58 | (0.54–12.42) | |
| 51 | 1 | 1.8% | 1 | 4.3% | 2 | 2.4% | 0 | - | 4 | 0.9% | 1.33 | - | - | 2.00 | (0.17–23.43) | - | - | ||
| 52 | 6 | 10.9% | 2 | 8.7% | 7 | 8.4% | 0 | - | 15 | 3.2% | 0.77 | - | - | - | - | - | |||
| 56 | 6 | 10.9% | - | - | 5 | 6.0% | 1 | 5.3% | 12 | 2.6% | 0.55 | 0.48 | 0.87 | - | - | 1.56 | (0.17–14.15) | ||
| 58 | 1 | 1.8% | 1 | 4.3% | 6 | 7.2% | 0 | - | 8 | 1.7% | 3.98 | - | - | 4.30 | (0.50–36.68) | - | - | ||
| 59 | 2 | 3.6% | 1 | 4.3% | 1 | 1.2% | 0 | - | 4 | 0.9% | 0.33 | - | - | - | - | - | |||
| 66 | 3 | 5.5% | 1 | 4.3% | 3 | 3.6% | 0 | - | 7 | 1.5% | 0.66 | - | - | - | - | - | |||
| A. | ASCL1 | LHX8 | ST6GALNAC5 | ||||||
| N | Median | p-Value | Median | p-Value | Median | p-Value | |||
| HPV16 vs. other types | |||||||||
| CIN3 | other | 58 | 3.21 | 1.46 | 6.27 | ||||
| HPV16 | 24 | 8.14 | 0.09 | 6.16 | 0.08 | 16.68 | 0.05 | ||
| ICC | other | 11 | 6.54 | 4.86 | 15.16 | ||||
| HPV16 | 10 | 12.69 | 0.26 | 11.05 | 0.73 | 25.48 | 0.53 | ||
| HPV18 vs. other types * | |||||||||
| CIN3 | other | 76 | 4.03 | 2.04 | 9.09 | ||||
| HPV18 | 6 | 1.45 | 0.71 | 0.55 | 0.55 | 4.02 | 0.78 | ||
| ICC | other | 18 | 7.58 | 4.68 | 11.55 | ||||
| HPV18 | 3 | 12.56 | 0.62 | 24.84 | 0.07 | 54.85 | 0.06 | ||
| HPV45 vs. other types * | |||||||||
| CIN3 | other | 74 | 3.82 | 1.90 | 8.92 | ||||
| HPV45 | 8 | 3.91 | 0.60 | 2.39 | 0.76 | 10.32 | 0.90 | ||
| ICC | other | 18 | 9.22 | 10.98 | 22.86 | ||||
| HPV45 | 3 | 4.07 | 0.76 | 4.49 | 0.92 | 7.11 | 0.62 | ||
| B. | ASCL1 | LHX8 | ST6GALNAC5 | ||||||
| N | Median | p-Value | Median | p-Value | Median | p-Value | |||
| HPV16 vs. other types | |||||||||
| ≤CIN2 | other | 87 | 0.08 | - | 0.20 | - | 2.17 | - | |
| HPV16 | 7 | 0.02 | 0.90 | 0.03 | 0.50 | 3.29 | 0.34 | ||
| CIN3+ | other | 69 | 3.75 | - | 1.83 | - | 7.11 | - | |
| HPV16 | 34 | 8.83 | 0.017 | 6.34 | 0.019 | 17.79 | 0.026 | ||
| HPV18 vs. other types * | |||||||||
| ≤CIN2 | other | 80 | 0.07 | - | 0.13 | - | 2.26 | - | |
| HPV18 | 14 | 0.18 | 0.10 | 0.37 | 0.34 | 2.17 | 0.87 | ||
| CIN3+ | other | 94 | 4.48 | - | 3.03 | - | 9.09 | - | |
| HPV18 | 9 | 2.24 | 0.88 | 13.08 | 0.48 | 29.92 | 0.27 | ||
| HPV45 vs. other types * | |||||||||
| ≤CIN2 | other | 81 | 0.07 | - | 0.14 | - | 2.37 | - | |
| HPV45 | 13 | 0.10 | 0.58 | 0.21 | 0.67 | 1.94 | 0.93 | ||
| CIN3+ | other | 92 | 4.48 | - | 3.20 | - | 9.56 | - | |
| HPV45 | 11 | 4.07 | 0.69 | 2.97 | 0.94 | 7.49 | 0.81 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kremer, W.W.; Van Zummeren, M.; Heideman, D.A.M.; Lissenberg-Witte, B.I.; Snijders, P.J.F.; Steenbergen, R.D.M.; Dreyer, G.; Meijer, C.J.L.M. HPV16-Related Cervical Cancers and Precancers Have Increased Levels of Host Cell DNA Methylation in Women Living with HIV. Int. J. Mol. Sci. 2018, 19, 3297. https://doi.org/10.3390/ijms19113297
Kremer WW, Van Zummeren M, Heideman DAM, Lissenberg-Witte BI, Snijders PJF, Steenbergen RDM, Dreyer G, Meijer CJLM. HPV16-Related Cervical Cancers and Precancers Have Increased Levels of Host Cell DNA Methylation in Women Living with HIV. International Journal of Molecular Sciences. 2018; 19(11):3297. https://doi.org/10.3390/ijms19113297
Chicago/Turabian StyleKremer, Wieke W., Marjolein Van Zummeren, Daniëlle A. M. Heideman, Birgit I. Lissenberg-Witte, Peter J. F. Snijders, Renske D. M. Steenbergen, Greta Dreyer, and Chris J. L. M. Meijer. 2018. "HPV16-Related Cervical Cancers and Precancers Have Increased Levels of Host Cell DNA Methylation in Women Living with HIV" International Journal of Molecular Sciences 19, no. 11: 3297. https://doi.org/10.3390/ijms19113297