The Interplay between Glucose-Regulated Protein 78 (GRP78) and Steroids in the Reproductive System

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
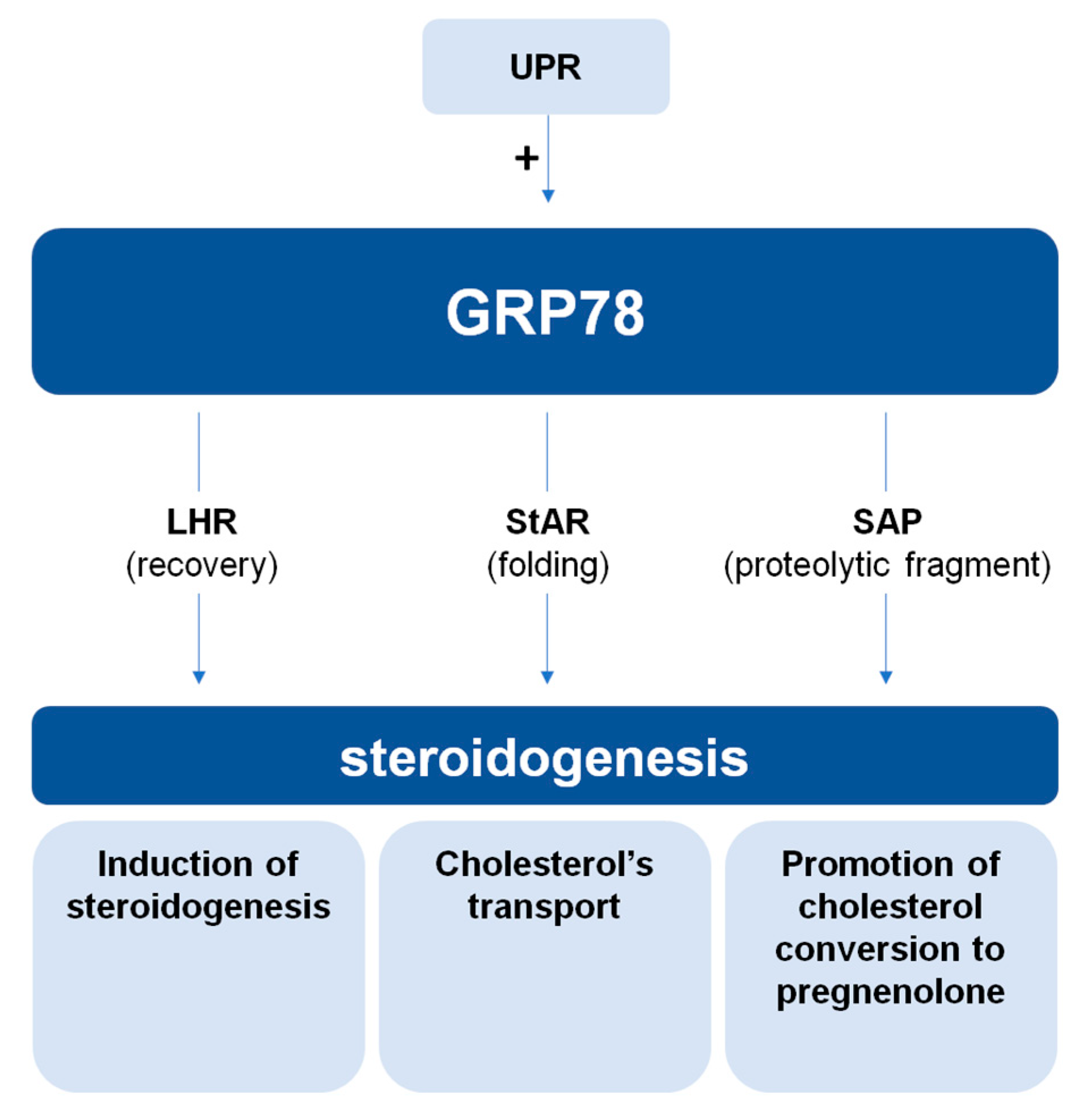
2. The Role of GRP78 in Steroidogenesis
3. The Role of Sex Steroids on GRP78 Expression and Function
3.1. Estrogen
3.1.1. Female Reproduction: Uterus and Early Pregnancy
3.1.2. Tumorigenesis: Endometrial, Ovary and Breast Cancer
3.2. Testosterone
3.2.1. Male Reproduction: Testis
3.2.2. Tumorigenesis: Prostate Cancer
4. Conclusions
Funding
Conflicts of Interest
Abbreviations
| AR | Androgen Receptor |
| ATF6 | Activating Transcription Factor 6 |
| BPA | Bisphenol A |
| CHOP | C/EBO Homologous Protein |
| E2 | Estradiol |
| ER | Endoplasmic Reticulum |
| ESR | Estrogens Receptor |
| FSH | Follicule-Stimulating Hormone |
| FSHR | Follicule-Stimulating Hormone Receptor |
| GRP78 | Glucose-Regulated Protein 78 |
| hCG | Human Chorionic Gonadotropin |
| HSP | Heat Shock Protein |
| IRE1 | Inositol-Requiring Enzyme 1 |
| LDL | Low-Density Lipoprotein |
| LH | Luteinizing Hormone |
| LHR | Luteinizing Hormone receptor |
| PERK | Protein Kinase R-like Endoplasmic Reticulum Kinase |
| SAP | Steroidogenesis-Activator Polypeptide |
| StAR | Steroidogenic Acute Regulatory Protein |
| T | Testosterone |
| UPR | Unfolded Protein Response |
| 3βHSD2 | 3β-Hydroxysteroid dehydrogenase type 2 |
| 17βHSD | 17β-Hydroxysteroid dehydrogenase |
References
- Guzel, E.; Basar, M.; Ocak, N.; Arici, A.; Kayisli, U.A. Bidirectional interaction between unfolded-protein-response key protein HSPA5 and estrogen signaling in human endometrium. Biol. Reprod. 2011, 85, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods 2005, 35, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Stetler, R.A.; Gan, Y.; Zhang, W.; Liou, A.K.; Gao, Y.; Cao, G.; Chen, J. Heat shock proteins: Cellular and molecular mechanisms in the central nervous system. Prog. Neurobiol. 2010, 92, 184–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, M.; Zhang, Y.; Lee, A.S. Beyond the endoplasmic reticulum: Atypical GRP78 in cell viability, signalling and therapeutic targeting. Biochem. J. 2011, 434, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, R.J. Stress signaling from the lumen of the endoplasmic reticulum: Coordination of gene transcriptional and translational controls. Genes Dev. 1999, 13, 1211–1233. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Schroder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat. Res. 2005, 569, 29–63. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chen, X.; Hendershot, L.; Prywes, R. ER stress regulation of ATF6 localization by dissociation of BiP/GRP78 binding and unmasking of Golgi localization signals. Dev. Cell 2002, 3, 99–111. [Google Scholar] [CrossRef]
- Gardner, B.M.; Pincus, D.; Gotthardt, K.; Gallagher, C.M.; Walter, P. Endoplasmic reticulum stress sensing in the unfolded protein response. Cold Spring Harb. Perspect. Biol. 2013, 5, a013169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.H.; Zhang, X. Roles of GRP78 in physiology and cancer. J. Cell. Biochem. 2010, 110, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Casas, C. GRP78 at the Centre of the Stage in Cancer and Neuroprotection. Front. Neurosci. 2017, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Kania, E.; Pajak, B.; Orzechowski, A. Calcium homeostasis and ER stress in control of autophagy in cancer cells. BioMed Res. Int. 2015, 2015, 352794. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. Glucose-regulated proteins in cancer: Molecular mechanisms and therapeutic potential. Nat. Rev. Cancer 2014, 14, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Ogino, Y.; Tohyama, S.; Kohno, S.; Toyota, K.; Yamada, G.; Yatsu, R.; Kobayashi, T.; Tatarazako, N.; Sato, T.; Matsubara, H.; et al. Functional distinctions associated with the diversity of sex steroid hormone receptors ESR and AR. J. Steroid Biochem. Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, X.A.; Warren, D.W.; Gregoire, J.; Pedersen, R.C.; Lee, A.S. The rat 78,000 dalton glucose-regulated protein (GRP78) as a precursor for the rat steroidogenesis-activator polypeptide (SAP): The SAP coding sequence is homologous with the terminal end of GRP78. Mol. Endocrinol. 1989, 3, 1944–1952. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Molecular biology of steroid hormone synthesis. Endocr. Rev. 1988, 9, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Pelham, H.R. A C-terminal signal prevents secretion of luminal ER proteins. Cell 1987, 48, 899–907. [Google Scholar] [CrossRef]
- Pedersen, R.C.; Brownie, A.C. Cholesterol side-chain cleavage in the rat adrenal cortex: Isolation of a cycloheximide-sensitive activator peptide. Proc. Natl. Acad. Sci. USA 1983, 80, 1882–1886. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.; Pawlak, K.J.; Burak, W.E.; Perry, E.E.; Marshall, B.; Whittal, R.M.; Bose, H.S. Mitochondrial metabolic regulation by GRP78. Sci. Adv. 2017, 3, e1602038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizrachi, D.; Segaloff, D.L. Intracellularly located misfolded glycoprotein hormone receptors associate with different chaperone proteins than their cognate wild-type receptors. Mol. Endocrinol. 2004, 18, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Menon, K.M.; Gunaga, K.P. Role of cyclic AMP in reproductive processes. Fertil. Steril. 1974, 25, 732–750. [Google Scholar] [CrossRef]
- Hsueh, A.J.; Adashi, E.Y.; Jones, P.B.; Welsh, T.H., Jr. Hormonal regulation of the differentiation of cultured ovarian granulosa cells. Endocr. Rev. 1984, 5, 76–127. [Google Scholar] [CrossRef] [PubMed]
- Kogure, K.; Nakamura, K.; Ikeda, S.; Kitahara, Y.; Nishimura, T.; Iwamune, M.; Minegishi, T. Glucose-regulated protein, 78-kilodalton is a modulator of luteinizing hormone receptor expression in luteinizing granulosa cells in rats. Biol. Reprod. 2013, 88, 8. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, T.S.; Park, C.K.; Lee, S.H.; Kim, J.M.; Lee, K.S.; Lee, I.K.; Park, J.W.; Lawson, M.A.; Lee, D.S. hCG-induced endoplasmic reticulum stress triggers apoptosis and reduces steroidogenic enzyme expression through activating transcription factor 6 in Leydig cells of the testis. J. Mol. Endocrinol. 2013, 50, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, S.J.; Koo, D.B.; Lee, S.R.; Kong, I.K.; Ryoo, J.W.; Park, Y.I.; Chang, K.T.; Lee, D.S. Progesterone production is affected by unfolded protein response (UPR) signaling during the luteal phase in mice. Life Sci. 2014, 113, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, S.J.; Kim, T.S.; Kim, J.M.; Lee, D.S. Testosterone production by a Leydig tumor cell line is suppressed by hyperthermia-induced endoplasmic reticulum stress in mice. Life Sci. 2016, 146, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, H.; Lin, P.; Wang, A.; Wang, L.; Jin, Y. ATF6 knockdown decreases apoptosis, arrests the S phase of the cell cycle, and increases steroid hormone production in mouse granulosa cells. Am. J. Physiol. Cell Physiol. 2017, 312, C341–C353. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Harada, M.; Hirota, Y.; Zhao, L.; Azhary, J.M.; Yoshino, O.; Izumi, G.; Hirata, T.; Koga, K.; Wada-Hiraike, O.; et al. A Potential Role for Endoplasmic Reticulum Stress in Progesterone Deficiency in Obese Women. Endocrinology 2017, 158, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Lachance, C.; Bailey, J.L.; Leclerc, P. Expression of Hsp60 and Grp78 in the human endometrium and oviduct, and their effect on sperm functions. Hum. Reprod. 2007, 22, 2606–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnaudeau, S.; Arboit, P.; Bischof, P.; Shin-ya, K.; Tomida, A.; Tsuruo, T.; Irion, O.; Cohen, M. Glucose-regulated protein 78: A new partner of p53 in trophoblast. Proteomics 2009, 9, 5316–5327. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Jin, Y.; Lan, X.; Yang, Y.; Chen, F.; Wang, N.; Li, X.; Sun, Y.; Wang, A. GRP78 expression and regulation in the mouse uterus during embryo implantation. J. Mol. Histol. 2014, 45, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Herington, J.; Galindo, C.L.; Ding, T.; Brown, N.; Reese, J.; Paria, B.C. Cross-species transcriptomic approach reveals genes in hamster implantation sites. Reproduction 2014, 148, 607–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.; Hou, X.; Zhou, H.E.; Wang, H.; Das, S.K. BiP is a molecular link between the phase I and phase II estrogenic responses in uterus. Mol. Endocrinol. 2006, 20, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Xu, F.; Li, P.; Sanchez, N.S.; Wang, H.; Das, S.K. Increased level of cellular BiP critically determines estrogenic potency for a xenoestrogen kepone in the mouse uterus. Endocrinology 2007, 148, 4774–4785. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Chen, F.; Yang, Y.; Song, Y.; Li, X.; Lan, X.; Jin, Y.; Wang, A. GRP78 expression and immunohistochemical localization in the female reproductive tract of mice. Theriogenology 2012, 78, 1824–1829. [Google Scholar] [CrossRef] [PubMed]
- Papaconstantinou, A.D.; Fisher, B.R.; Umbreit, T.H.; Goering, P.L.; Lappas, N.T.; Brown, K.M. Effects of beta-estradiol and bisphenol A on heat shock protein levels and localization in the mouse uterus are antagonized by the antiestrogen ICI 182,780. Toxicol. Sci. 2001, 63, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Papaconstantinou, A.D.; Goering, P.L.; Umbreit, T.H.; Brown, K.M. Regulation of uterine hsp90α, hsp72 and HSF-1 transcription in B6C3F1 mice by β-estradiol and bisphenol A: Involvement of the estrogen receptor and protein kinase C. Toxicol. Lett. 2003, 144, 257–270. [Google Scholar] [CrossRef]
- Kita, K.; Jin, Y.H.; Sun, Z.; Chen, S.P.; Sumiya, Y.; Hongo, T.; Suzuki, N. Increase in the levels of chaperone proteins by exposure to β-estradiol, bisphenol A and 4-methoxyphenol in human cells transfected with estrogen receptor α cDNA. Toxicol. In Vitro 2009, 23, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Department of Gynecology Obstetrics, Faculty of Medicine, University of Geneva. Human endometrial biopsies study. Unpublished data, 2014. [Google Scholar]
- Cali, G.; Insabato, L.; Conza, D.; Bifulco, G.; Parrillo, L.; Mirra, P.; Fiory, F.; Miele, C.; Raciti, G.A.; di Jeso, B.; et al. GRP78 mediates cell growth and invasiveness in endometrial cancer. J. Cell. Physiol. 2014, 229, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Guzel, E.; Arlier, S.; Guzeloglu-Kayisli, O.; Tabak, M.S.; Ekiz, T.; Semerci, N.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Endoplasmic Reticulum Stress and Homeostasis in Reproductive Physiology and Pathology. Int. J. Mol. Sci. 2017, 18, 792. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, G.; Paridaens, R. Optimal sequence of hormonotherapy in advanced breast cancer. Curr. Opin. Oncol. 2006, 18, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, D.J.; Livezey, M.; Yu, L.; Zheng, X.; Andruska, N. Anticipatory UPR Activation: A Protective Pathway and Target in Cancer. Trends Endocrinol. Metab. 2016, 27, 731–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andruska, N.; Zheng, X.; Yang, X.; Helferich, W.G.; Shapiro, D.J. Anticipatory estrogen activation of the unfolded protein response is linked to cell proliferation and poor survival in estrogen receptor α-positive breast cancer. Oncogene 2015, 34, 3760–3769. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, J.; Lee, A.S. GRP78/BiP inhibits endoplasmic reticulum BIK and protects human breast cancer cells against estrogen starvation-induced apoptosis. Cancer Res. 2007, 67, 3734–3740. [Google Scholar] [CrossRef] [PubMed]
- Cook, K.L.; Shajahan, A.N.; Warri, A.; Jin, L.; Hilakivi-Clarke, L.A.; Clarke, R. Glucose-regulated protein 78 controls cross-talk between apoptosis and autophagy to determine antiestrogen responsiveness. Cancer Res. 2012, 72, 3337–3349. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Zhu, Y.F.; Ma, X.; Lin, M.; Zhou, Z.M.; Sha, J.H. Differential expression of glucose-regulated protein 78 during spermatogenesis. Cell Tissue Res. 2004, 316, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Lachance, C.; Fortier, M.; Thimon, V.; Sullivan, R.; Bailey, J.L.; Leclerc, P. Localization of Hsp60 and Grp78 in the human testis, epididymis and mature spermatozoa. Int. J. Androl. 2010, 33, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Day, A.R.; Lee, A.S. Transcriptional regulation of the gene encoding the 78-kD glucose-regulated protein GRP78 in mouse sertoli cells: Binding of specific factor(s) to the GRP78 promoter. DNA 1989, 8, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, Y.; Takasaki, I.; Kondo, T. Identification of genetic networks involved in the cell injury accompanying endoplasmic reticulum stress induced by bisphenol A in testicular Sertoli cells. Biochem. Biophys. Res. Commun. 2006, 345, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Storm, M.; Sheng, X.; Arnoldussen, Y.J.; Saatcioglu, F. Prostate cancer and the unfolded protein response. Oncotarget 2016, 7, 54051–54066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, X.; Arnoldussen, Y.J.; Storm, M.; Tesikova, M.; Nenseth, H.Z.; Zhao, S.; Fazli, L.; Rennie, P.; Risberg, B.; Waehre, H.; et al. Divergent androgen regulation of unfolded protein response pathways drives prostate cancer. EMBO Mol. Med. 2015, 7, 788–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.S.; Ahmad, I.; Bennett, H.L.; Singh, L.; Nixon, C.; Seywright, M.; Barnetson, R.J.; Edwards, J.; Leung, H.Y. GRP78 up-regulation is associated with androgen receptor status, Hsp70-Hsp90 client proteins and castrate-resistant prostate cancer. J. Pathol. 2011, 223, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Department of Gynecology Obstetrics, Faculty of Medicine, University of Geneva. Prostate cancer cell lines study (PNT1B, PNT2C2, LNCaP, DU145, PC3). Unpublished data, 2011. [Google Scholar]
- Bennett, H.L.; Fleming, J.T.; O’Prey, J.; Ryan, K.M.; Leung, H.Y. Androgens modulate autophagy and cell death via regulation of the endoplasmic reticulum chaperone glucose-regulated protein 78/BiP in prostate cancer cells. Cell Death Dis. 2010, 1, e72. [Google Scholar] [CrossRef] [PubMed]
- Dhamad, A.E.; Zhou, Z.; Zhou, J.; Du, Y. Systematic Proteomic Identification of the Heat Shock Proteins (Hsp) that Interact with Estrogen Receptor α (ERα) and Biochemical Characterization of the ERα–Hsp70 Interaction. PLoS ONE 2016, 11, e0160312. [Google Scholar] [CrossRef] [PubMed]
- Urra, H.; Dufey, E.; Avril, T.; Chevet, E.; Hetz, C. Endoplasmic Reticulum Stress and the Hallmarks of Cancer. Trends Cancer 2016, 2, 252–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hebert-Schuster, M.; Rotta, B.E.; Kirkpatrick, B.; Guibourdenche, J.; Cohen, M. The Interplay between Glucose-Regulated Protein 78 (GRP78) and Steroids in the Reproductive System. Int. J. Mol. Sci. 2018, 19, 1842. https://doi.org/10.3390/ijms19071842
Hebert-Schuster M, Rotta BE, Kirkpatrick B, Guibourdenche J, Cohen M. The Interplay between Glucose-Regulated Protein 78 (GRP78) and Steroids in the Reproductive System. International Journal of Molecular Sciences. 2018; 19(7):1842. https://doi.org/10.3390/ijms19071842
Chicago/Turabian StyleHebert-Schuster, Marylise, Belinda Elisabeth Rotta, Brenna Kirkpatrick, Jean Guibourdenche, and Marie Cohen. 2018. "The Interplay between Glucose-Regulated Protein 78 (GRP78) and Steroids in the Reproductive System" International Journal of Molecular Sciences 19, no. 7: 1842. https://doi.org/10.3390/ijms19071842