Hamster Melatonin Receptors: Cloning and Binding Characterization of MT1 and Attempt to Clone MT2

 and
and
Abstract
:1. Introduction
2. Results
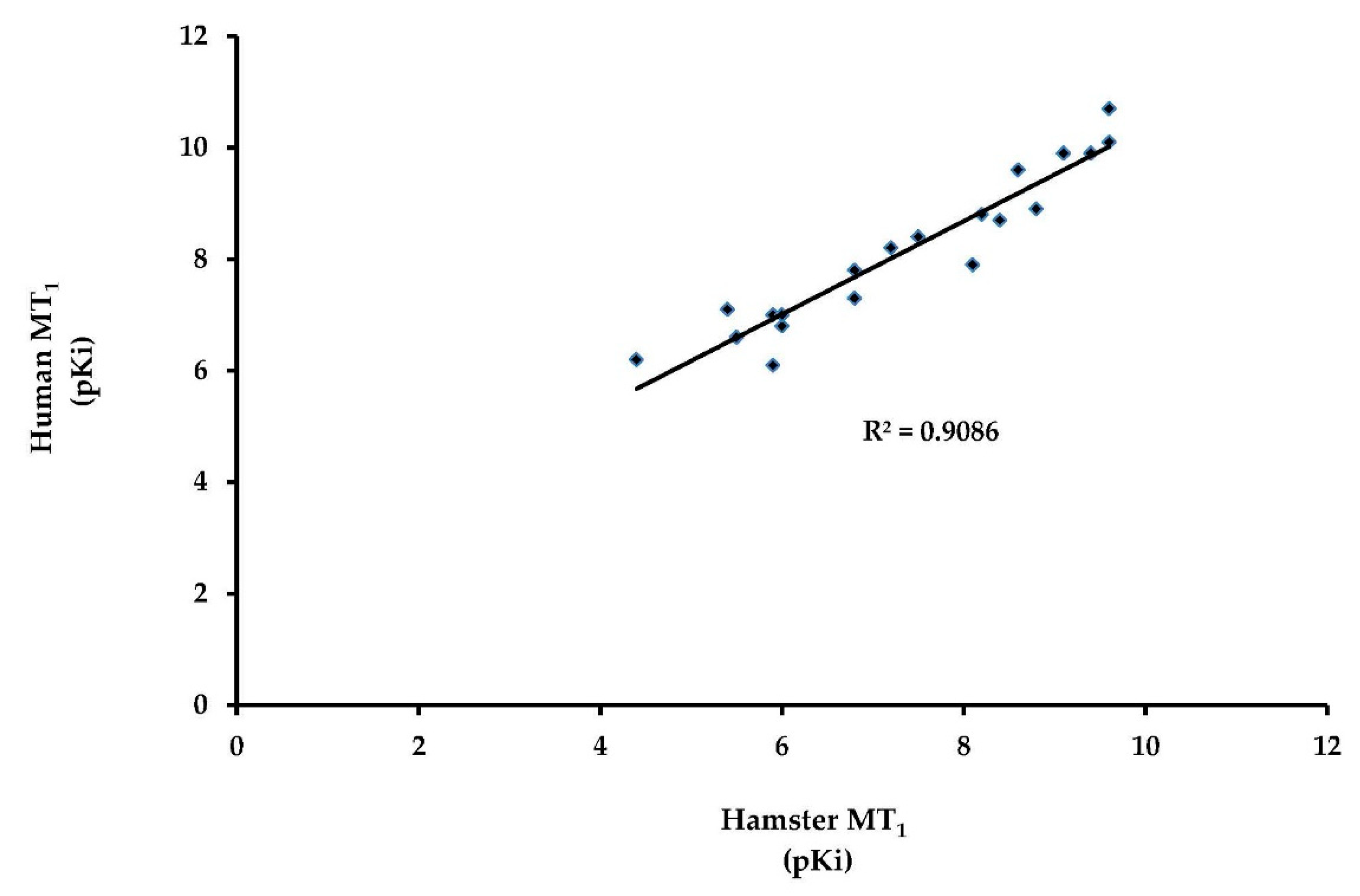
2.1. The European Hamster MT1 Gene Encodes a Functional Receptor
2.2. Cloning of Exon 1 of the Phodopus sungorus MT2 Gene
2.3. The Cricetus cricetus MT2 Receptor Seems to Be Functional
2.3.1. Cloning of Exon 2 of the Cricetus cricetus MT2 Receptor
2.3.2. Cloning of Exon 1 of the Cricetus cricetus MT2 Receptor
2.3.3. Molecular Pharmacology
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Collection
4.1.1. Cricetus cricetus
4.1.2. Phodopus sungorus
4.2. RNA Extraction
4.3. Cloning of the Phodopus sungorus MT2 Gene
4.4. Cloning of the Cricetus cricetus MT1 Gene
4.5. Cloning of the Cricetus cricetus MT2 Gene
4.5.1. Cloning of a Partial Fragment of Exon 1 of the MT2 Receptor from Cricetus cricetus
4.5.2. 5′ RACE of the Cricetus cricetus MT2 Gene
4.5.3. Further Attempts to Clone Exon 1 of the Cricetus cricetus MT2 Receptor
4.5.4. Cloning of Exon 2 and the 3′-UTR of the Cricetus cricetus MT2 Receptor
4.6. Subcloning and Sequencing
4.7. Establishment of Transient CHO-Flp-in MT1 Cell Lines
4.8. Membrane Preparations
4.9. Establishment of Transient COS7/Chimeric MT2 Cell Lines
4.9.1. Design of the Chimeric MT2 Receptor
4.9.2. Establishment of the Transient COS7-Chimeric MT2 Cell Line
4.10. Whole Cell and Membrane 2-[125I]-Iodomelatonin Binding Assay
Author Contributions
Funding
Conflicts of Interest
References
- Klein, D.C. Photoneural regulation of the mammalian pineal gland. Ciba Found. Symp. 1985, 117, 38–56. [Google Scholar] [PubMed]
- Kolář, J.; Macháčková, I.; Eder, J.; Prinsen, E.; van Dongen, W.; van Onckelen, H.; Illnerová, H. Melatonin: Occurrence and daily rhythm in Chenopodium rubrum. Phytochemistry 1997, 44, 1407–1413. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current Status and Future Perspectives in Plant Science. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Coomans, C.P.; Ramkisoensing, A.; Meijer, J.H. The suprachiasmatic nuclei as a seasonal clock. Front. Neuroendocrinol. 2015, 37, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, G.A.; Clarke, I.J.; Hut, R.A.; Hazlerigg, D.G. Characterizing a Mammalian Circannual Pacemaker. Science 2006, 314, 1941–1944. [Google Scholar] [CrossRef] [PubMed]
- De Bodinat, C.; Guardiola-Lemaitre, B.; Mocaër, E.; Renard, P.; Muñoz, C.; Millan, M.J. Agomelatine, the first melatonergic antidepressant: Discovery, characterization and development. Nat. Rev. Drug Discov. 2010, 9, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J.; Gobert, A.; Lejeune, F.; Dekeyne, A.; Newman-Tancredi, A.; Pasteau, V.; Rivet, J.-M.; Cussac, D. The Novel Melatonin Agonist Agomelatine (S20098) Is an Antagonist at 5-Hydroxytryptamine2C Receptors, Blockade of Which Enhances the Activity of Frontocortical Dopaminergic and Adrenergic Pathways. J. Pharmacol. Exp. Ther. 2003, 306, 954–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audinot, V.; Bonnaud, A.; Grandcolas, L.; Rodriguez, M.; Nagel, N.; Galizzi, J.-P.; Balik, A.; Messager, S.; Hazlerigg, D.G.; Barrett, P.; et al. Molecular cloning and pharmacological characterization of rat melatonin MT1 and MT2 receptors. Biochem. Pharmacol. 2008, 75, 2007–2019. [Google Scholar] [CrossRef] [PubMed]
- Cogé, F.; Guenin, S.; Fery, I.; Migaud, M.; Devavry, S.; Slugocki, C.; Legros, C.; Ouvry, C.; Cohen, W.; Renault, N.; et al. The end of a myth: Cloning and characterization of the ovine melatonin MT2 receptor. Br. J. Pharmacol. 2009, 158, 1248–1262. [Google Scholar] [CrossRef] [PubMed]
- Devavry, S.; Legros, C.; Brasseur, C.; Cohen, W.; Guenin, S.-P.; Delagrange, P.; Malpaux, B.; Ouvry, C.; Cogé, F.; Nosjean, O.; Boutin, J.A. Molecular pharmacology of the mouse melatonin receptors MT1 and MT2. Eur. J. Pharmacol. 2012, 677, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Piketty, V.; Pelletier, J. Melatonin receptors in the lamb pars tuberalis/median eminence throughout the day. Neuroendocrinology 1993, 58, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Becker-André, M.; Wiesenberg, I.; Schaeren-Wiemers, N.; André, E.; Missbach, M.; Saurat, J.H.; Carlberg, C. Pineal gland hormone melatonin binds and activates an orphan of the nuclear receptor superfamily. J. Biol. Chem. 1994, 269, 28531–28534. [Google Scholar] [CrossRef] [PubMed]
- Becker-André, M.; Wiesenberg, I.; Schaeren-Wiemers, N.; André, E.; Missbach, M.; Saurat, J.-H.; Carlberg, C. Pineal gland hormone melatonin binds and activates an orphan of the nuclear receptor superfamily. J. Biol. Chem. 1997, 272, 16707. [Google Scholar] [CrossRef]
- Jan, J.E.; Reiter, R.J.; Wong, P.K.H.; Bax, M.C.O.; Ribary, U.; Wasdell, M.B. Melatonin has membrane receptor-independent hypnotic action on neurons: An hypothesis. J. Pineal Res. 2011, 50, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Nosjean, O.; Ferro, M.; Cogé, F.; Beauverger, P.; Henlin, J.-M.; Lefoulon, F.; Fauchère, J.-L.; Delagrange, P.; Canet, E.; Boutin, J.A. Identification of the melatonin-binding site MT3 as the quinone reductase 2. J. Biol. Chem. 2000, 275, 31311–31317. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT1 and MT2 Melatonin Receptors: A Therapeutic Perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, C.; Lovenberg, T.W. Molecular and pharmacological characterization of the mouse histamine H3 receptor. Eur. J. Pharmacol. 2003, 467, 57–65. [Google Scholar] [CrossRef]
- Alnouri, M.W.; Jepards, S.; Casari, A.; Schiedel, A.C.; Hinz, S.; Müller, C.E. Selectivity is species-dependent: Characterization of standard agonists and antagonists at human, rat, and mouse adenosine receptors. Purinergic Signal. 2015, 11, 389–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logez, C.; Berger, S.; Legros, C.; Banères, J.-L.; Cohen, W.; Delagrange, P.; Nosjean, O.; Boutin, J.A.; Ferry, G.; Simonin, F.; Wagner, R. Recombinant Human Melatonin Receptor MT1 Isolated in Mixed Detergents Shows Pharmacology Similar to That in Mammalian Cell Membranes. PLoS ONE 2014, 9, e100616. [Google Scholar] [CrossRef] [PubMed]
- Logez, C.; Damian, M.; Legros, C.; Dupré, C.; Guéry, M.; Mary, S.; Wagner, R.; M’Kadmi, C.; Nosjean, O.; Fould, B.; et al. Detergent-free Isolation of Functional G Protein-Coupled Receptors into Nanometric Lipid Particles. Biochemistry 2016, 55, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.-T.; Chu, M.-X.; Fu, Y.; Fang, L.; Ye, S.-C. Analysis of polymorphism, structure and function of exon 2 of ovine melatonin receptor 1b gene: A clue as to why it lacks expression in sheep. J. Pineal Res. 2007, 42, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Neumann, K.; Michaux, J.; Lebedev, V.; Yigit, N.; Colak, E.; Ivanova, N.; Poltoraus, A.; Surov, A.; Markov, G.; Maak, S.; et al. Molecular phylogeny of the Cricetinae subfamily based on the mitochondrial cytochrome b and 12S rRNA genes and the nuclear vWF gene. Mol. Phylogenet. Evol. 2006, 39, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, D.R.; Liu, C.; Reppert, S.M. Nature’s knockout: The Mel1b receptor is not necessary for reproductive and circadian responses to melatonin in Siberian hamsters. Mol. Endocrinol. 1996, 10, 1478–1487. [Google Scholar] [PubMed]
- Canguilhem, B.; Marx, C. Regulation of the body weight of the European Hamster during the annual cycle. Pflugers Arch. 1973, 338, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Gubitz, A.K.; Reppert, S.M. Assignment of the melatonin-related receptor to human chromosome X (GPR50) and mouse chromosome X (Gpr50). Genomics 1999, 55, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Clement, N.; Renault, N.; Guillaume, J.-L.; Cecon, E.; Journé, A.-S.; Laurent, X.; Tadagaki, K.; Cogé, F.; Gohier, A.; Delagrange, P.; et al. Importance of the second extracellular loop for melatonin MT1 receptor function and absence of melatonin binding in GPR50. Br. J. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Audinot, V.; Mailliet, F.; Lahaye-Brasseur, C.; Bonnaud, A.; Gall, A.L.; Amossé, C.; Dromaint, S.; Rodriguez, M.; Nagel, N.; Galizzi, J.-P.; et al. New selective ligands of human cloned melatonin MT1 and MT2 receptors. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 367, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Legros, C.; Devavry, S.; Caignard, S.; Tessier, C.; Delagrange, P.; Ouvry, C.; Boutin, J.A.; Nosjean, O. Melatonin MT1 and MT2 receptors display different molecular pharmacologies only in the G-protein coupled state. Br. J. Pharmacol. 2014, 171, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Yasuo, S.; Yoshimura, T.; Ebihara, S.; Korf, H.-W. Melatonin Transmits Photoperiodic Signals through the MT1 Melatonin Receptor. J. Neurosci. 2009, 29, 2885–2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jockers, R.; Maurice, P.; Boutin, J.A.; Delagrange, P. Melatonin receptors, heterodimerization, signal transduction and binding sites: What’s new? Br. J. Pharmacol. 2008, 154, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Barrett, P.; Conway, S.; Morgan, P.J. Digging deep-structure-function relationships in the melatonin receptor family. J. Pineal Res. 2003, 35, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.; Bothorel, B.; Ciocca, D.; Valour, D.; Gaudeau, A.; Dupré, C.; Lizzo, G.; Brasseur, C.; Riest-Fery, I.; Stephan, J.-P.; et al. Gene expression profiling during European hamster’s hibernation. Sci. Rep. 2018, in press. [Google Scholar]
- Sano, Y.; Shiina, T.; Naitou, K.; Nakamori, H.; Shimizu, Y. Hibernation-specific alternative splicing of the mRNA encoding cold-inducible RNA-binding protein in the hearts of hamsters. Biochem. Biophys. Res. Commun. 2015, 462, 322–325. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Animal Name | Protein Names |
|---|---|---|
| MG598322 | European hamster (Cricetus cricetus) | Melatonin receptor type 1A (MT1) mRNA, complete cds |
| MG598323 | European hamster (Cricetus cricetus) | Melatonin receptor type 1B (MT2) mRNA, partial cds |
| MG784343 | Platypus (Ornithorhynchus anatinus) | Melatonin receptor type 1C (Mel1c) mRNA, complete cds |
| KF774288 | Short-nosed fruit bat (Cynopterus titthaecheilus) | Melatonin receptor type 1B (MT2), mRNA, complete cds |
| KC238664 | Free-tailed bat (Tadarida brasiliensis) | Melatonin receptor type 1B (MT2), mRNA, complete cds |
| KF486508.1 | Sparrow hawk (Accipiter nisus) | Melatonin receptor type 1B variant (MT2), mRNA, complete cds |
| KF486510 | Imperial eagle (Aquila heliacal) | Melatonin receptor type 1B variant (MT2) mRNA, complete cds |
| KF486512.1 | King penguin (Aptenodytes patagonicus) | Melatonin receptor type 1B variant (MT2) mRNA, complete cds |
| KF486516.1 | Humboldt penguin (Spheniscus humboldti) | Melatonin receptor type 1B variant (MT2) mRNA, complete cds |
| KF486517 | Ural owl (Strix uralensis) | Melatonin receptor type 1B variant (MT2) mRNA, complete cds |
| KF486518 | Clawed frog (Xenopus laevis) | Melatonin receptor type 1C (Mel1c) mRNA, complete cds |
| EU679365.1 | Common sheep (Ovis aries) | Melatonin receptor type 1B (MT2) mRNA, complete cds |
| Hamster MT1 | Human MT1 | Human MT2 | |
|---|---|---|---|
| MLT | 8.6 | 9.6 | 9.3 |
| 2I-MLT | 9.6 | 10.7 | 9.8 |
| 6 Chloro MLT | 8.4 | 8.7 | 9.6 |
| Luzindole | 5.5 | 6.6 | 7.6 |
| 4PPDOT | 5.9 | 6.8 | 8.9 |
| S 20098 | 9.1 | 9.9 | 9.9 |
| FLN68/ramelteon | 9.6 | 10.1 | 10.3 |
| D600 (+/−) | 5.9 | 7 | <5 |
| S20928 | 5.4 | 7.1 | 7 |
| S21278 | 4.4 | 6.2 | 6.2 |
| S22153 | 7.2 | 8.2 | 8 |
| S70254 | 5.9 | 7 | 9 |
| S73893 | 7.5 | 8.4 | 8.1 |
| S75436 | 8.0 | 7.9 | 8.9 |
| S27128 | 8.7 | 8.9 | 9.2 |
| Div 880 | 5.9 | 6.1 | 8 |
| SD6 | 9.4 | 9.9 | 9.9 |
| SD1881 | 8.2 | 8.8 | 8.6 |
| SD1882 | 6.8 | 7.8 | 7.9 |
| SD1918 | 6.8 | 7.3 | 7.3 |
| A. Primers used for the amplification of the Cricetus cricetus MT1 gene | |||
| Forward Primers | Reverse Primers | ||
| TGCGCTGCGGTGAGACACCCAGGGGACC | GCGTTCCTGAGCTTCTTGTTGC | ||
| ATGAAGGGCAATGGTAGCACTCTGCTCAATGCC | GCGTTCCTGAGCTTCTTGTTGC | ||
| ATGAAGGGCAATGGTAGCACTCTGCTCAATGCC | CCGTATATAATTGCATTGAGGCAGCTG | ||
| CCGCTACTGCTACATTTGCCACAGTCTC | CCGTATATAATTGCATTGAGGCAGCTG | ||
| CAGGAAATATATTTGTGGTGAG | TTAAACAGAGTCCACCTTTA | ||
| GTACTTTTTGCTATTTGCTGGGC | AAGACCCCAACCAGTGTGGATAATC | ||
| B. Primers used to amplify exon 1 of Cricetus cricetus MT2 | |||
| Forward Primers | Reverse Primers | ||
| ATGCCTGAGAACAGTTCTGTGCCAATTGCTGCG (F0) | CGCAGCTTGCGGTTCCTGAGCAC | ||
| CGCGTTCCGCAGCTTGCGGTTCC | |||
| GCTGCGAGGCTGGTGGGCTGGCAGTGCGCT (F3) | CGCAGCTTGCGGTTCCTGAGCAC | ||
| CGCGTTCCGCAGCTTGCGGTTCC | |||
| Note: The reverse primers are located in exon 1 | |||
| C. Primers designed based on the CHO melatonin receptor 2 sequence to amplify exon 1 of the Cricetus cricetus MT2 gene | |||
| Forward Primers | Reverse primer | ||
| ACCCCCACAGTTGCAAACAGAATGCACATCCG | ATTAGAGGATCCTCTTGTACTCCCTGCGGAAGTTCTGGTTC | ||
| CAGAATGCACATCCGATGCCCTGAGAACA | |||
| TCCGATGCCCTGAGAACAGCTCTGT | |||
| ATGCCTGAGAACAGCTCTGTCTCCAATTG | |||
| D. Primers sequences used for exon 1 amplification using genomic DNA as the template | |||
| Forward Sequences | Reverse Sequences | Size of Amplicons | |
| GCCTCTTCCTAGCACTTCGCTAG | GTGGGATGTGAAAGGATCTAAG | 411 bp | |
| CTGCGCGGTGAGGGGGCAGCGGG | GTGGGATGTGAAAGGATCTAAG | 389 bp | |
| GCCCCAAGCAGTACTCACCTTG | GTGGGATGTGAAAGGATCTAAG | 363 bp | |
| GCTTAGTCCCGATGCCCTGAG | GTGGGATGTGAAAGGATCTAAG | 321 bp | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautier, C.; Dufour, E.; Dupré, C.; Lizzo, G.; Caignard, S.; Riest-Fery, I.; Brasseur, C.; Legros, C.; Delagrange, P.; Nosjean, O.; et al. Hamster Melatonin Receptors: Cloning and Binding Characterization of MT1 and Attempt to Clone MT2. Int. J. Mol. Sci. 2018, 19, 1957. https://doi.org/10.3390/ijms19071957
Gautier C, Dufour E, Dupré C, Lizzo G, Caignard S, Riest-Fery I, Brasseur C, Legros C, Delagrange P, Nosjean O, et al. Hamster Melatonin Receptors: Cloning and Binding Characterization of MT1 and Attempt to Clone MT2. International Journal of Molecular Sciences. 2018; 19(7):1957. https://doi.org/10.3390/ijms19071957
Chicago/Turabian StyleGautier, Célia, Emilie Dufour, Clémence Dupré, Giulia Lizzo, Sarah Caignard, Isabelle Riest-Fery, Chantal Brasseur, Céline Legros, Philippe Delagrange, Olivier Nosjean, and et al. 2018. "Hamster Melatonin Receptors: Cloning and Binding Characterization of MT1 and Attempt to Clone MT2" International Journal of Molecular Sciences 19, no. 7: 1957. https://doi.org/10.3390/ijms19071957