Abnormal Hippocampal Melatoninergic System: A Potential Link between Absence Epilepsy and Depression-Like Behavior in WAG/Rij Rats?

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Melatonin Had No Role in the Circadian Rhythm of Absence Seizures and No Impact on their Incidence in WAG/Rij Rats
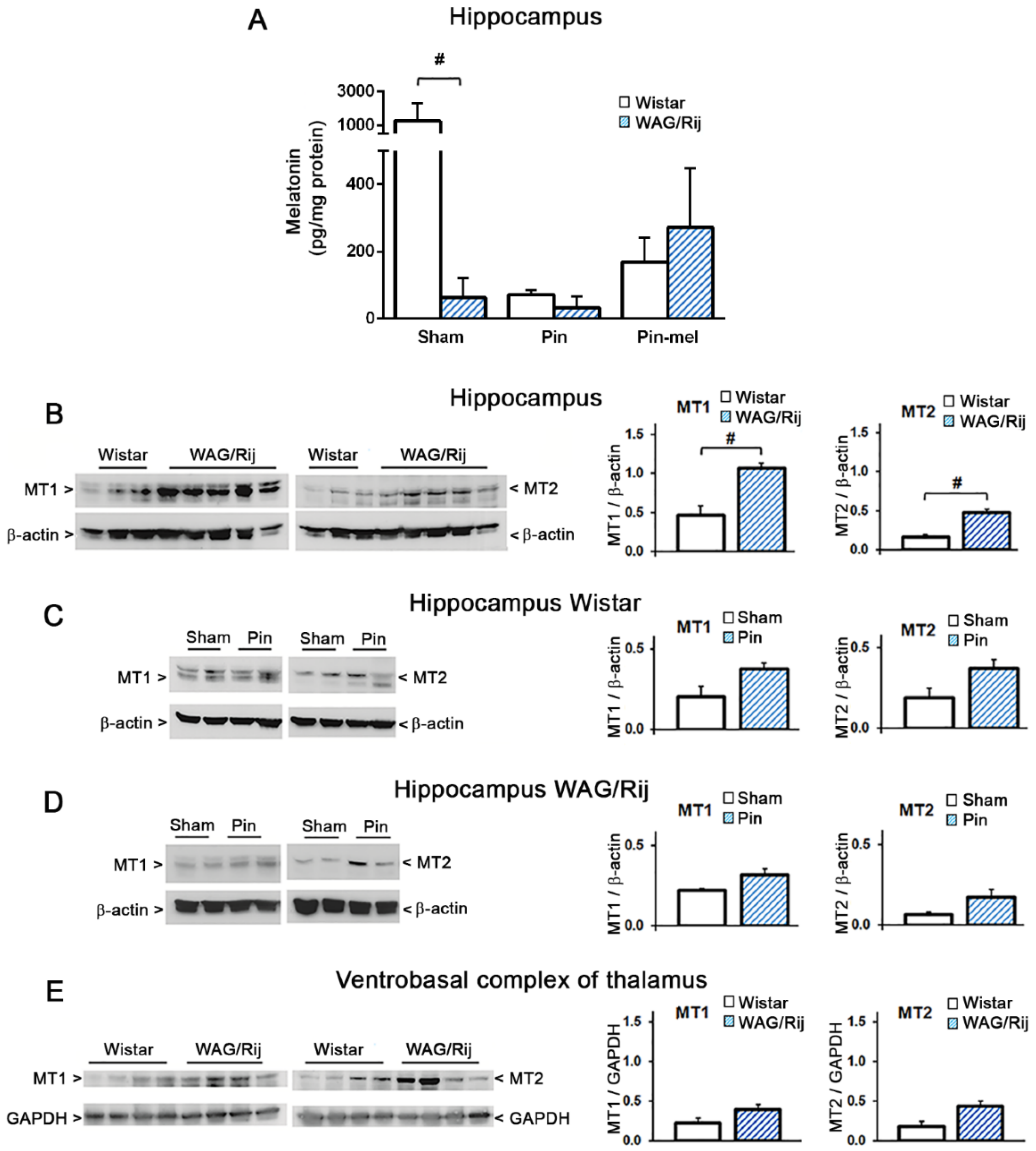
2.2. Abnormalities in Hippocampal Melatonergic System in WAG/Rij Rats
2.3. Reduced Hippocampal Cell Density in WAG/Rij Rats
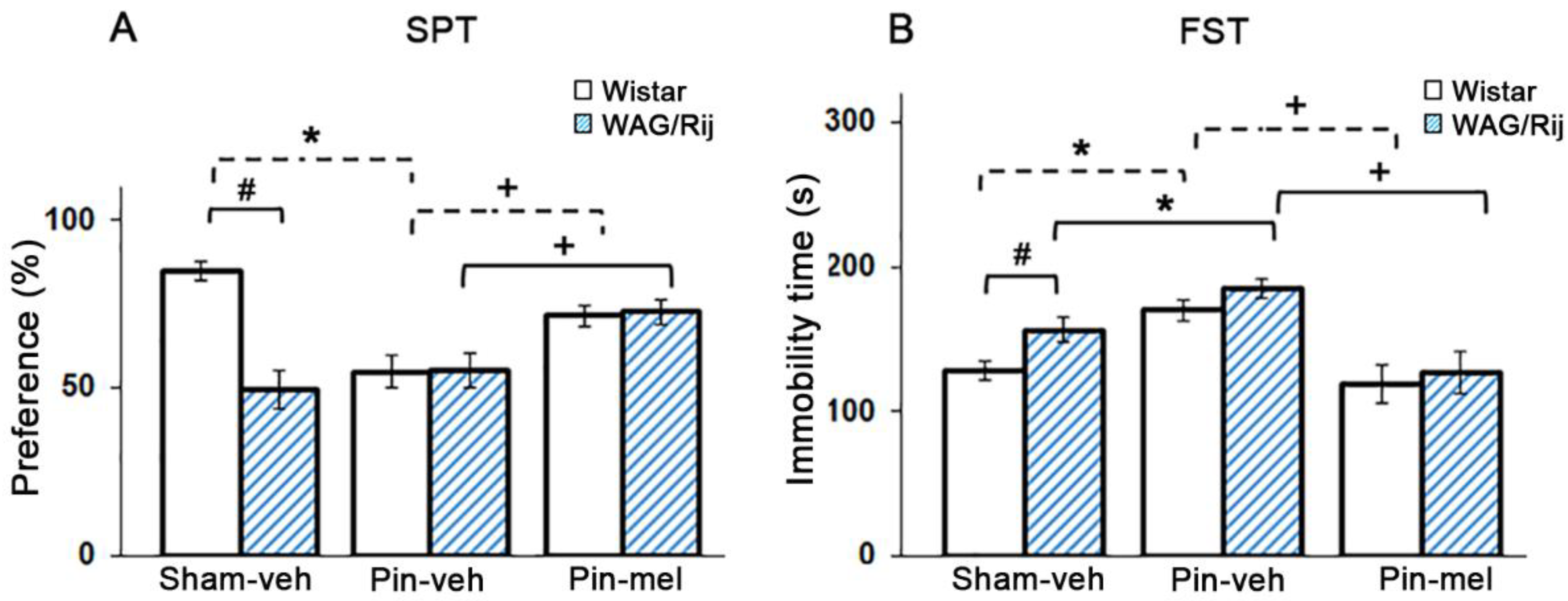
2.4. Influence of Melatonin on Depression-Like Behavior in WAG/Rij and Non-Epileptic Wistar Rats
3. Discussion
4. Materials and Methods
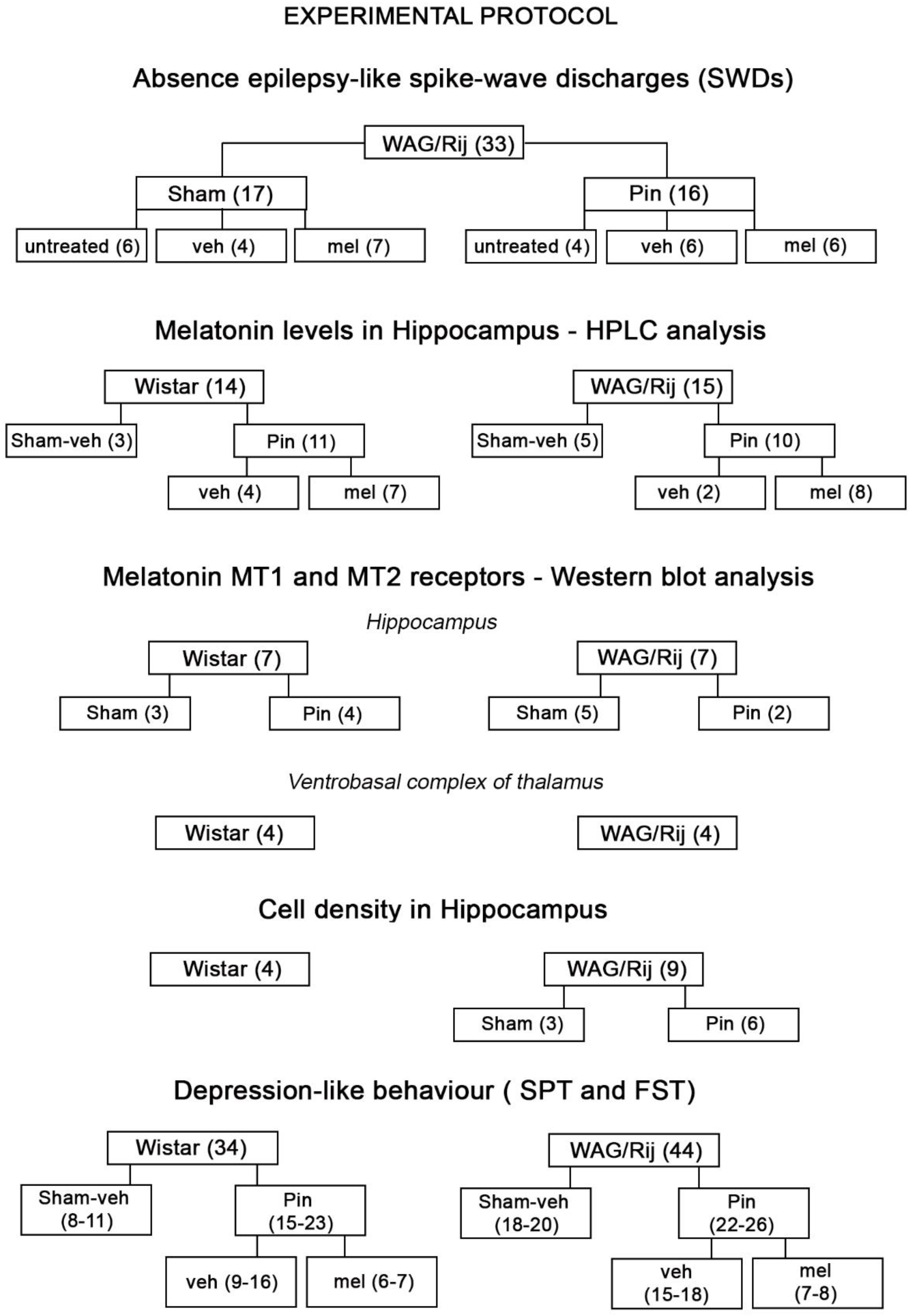
4.1. Animals
4.2. Drugs
4.3. Pinealectomy
4.4. Implantation of Electrodes and Recording and Analysis of SWDs
4.5. Measurements of Melatonin
4.6. Western Blotting of Melatonin Receptors
4.7. Histology
4.8. Behavioral Tests
4.8.1. Saccharine Preference Test (SPT)
4.8.2. Forced Swim Test (FST)
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| AUC | Area under the curve |
| CNS | Central nervous system |
| EEG | Electroencephalogram |
| ESCoG | Electrosubcorticogram |
| FST | Forced swim test |
| HPA | Hypothalamic-pituitary-adrenal axis |
| HPLC | High performance liquid chromatography |
| KA | Kainic acid (kainate) |
| LSD | Least significant difference |
| Mel | Melatonin |
| MT | Melatonin receptor |
| Pin | Pinealectomy (rats with removed pineal gland) |
| PT in SWDs | Percent time in SWDs |
| rANOVA | Repeated measure analysis of variance |
| OL | Percent overlap |
| s.c. | Subcutaneous |
| SE | Status epilepticus |
| S.E.M. | Standard error of mean |
| Sham | Sham-operated (rats without removed pineal gland) |
| SPT | Saccharine preference test |
| SWD | Spike-wave discharge |
| WAG/Rij | Wistar Albino Glaxo from Rijswijk rats |
References
- Mohanan, P.V.; Yamamoto, H.A. Preventive effect of melatonin against brain mitochondria DNA damage, lipid peroxidation and seizures induced by kainic acid. Toxicol. Lett. 2002, 129, 99–105. [Google Scholar] [CrossRef]
- Guisti, P.; Lipartiti, M.; Franceschini, D.; Schiavo, N.; Floream, M.; Manev, H. Neuroprotection by melatonin from kainate-induced excitotoxicity in rats. FASEB J. 1996, 10, 891–896. [Google Scholar] [CrossRef]
- Banach, M.; Gurdziel, E.; Jêdrych, M.; Borowicz, K. Melatonin in experimental seizures and epilepsy. Pharmacol. Rep. 2011, 63, 1–11. [Google Scholar] [CrossRef]
- Kopp, C.; Vogel, E.; Rettori, M.; Delagrange, P.; Misslin, R. Anxiolytic-like properties of melatonin receptor agonists in mice: Involvement of MT1 and/or MT2 receptors in the regulation of emotional responsiveness. Neuropharmacology 2000, 39, 1865–1871. [Google Scholar] [CrossRef]
- Loiseau, F.; Le Bihan, C.; Hamon, M.; Thiébot, M.H. Effects of melatonin and agomelatine in anxiety-related procedures in rats: Interaction with diazepam. Eur. Neuropsychopharmacol. 2006, 16, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.A.; Ahmed, H.I.; El-Morsy, E.M. Melatonin protects against diazinon-induced neurobehavioral changes in rats. Neurochem. Res. 2013, 38, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Kilic, E.; Kilic, U.; Bacigaluppi, M.; Guo, Z.; Abdallah, N.B.; Wolfer, D.P.; Reiter, R.J.; Hermann, D.M.; Bassetti, C.L. Delayed melatonin administration promotes neuronal survival, neurogenesis and motor recovery, and attenuates hyperactivity and anxiety after mild focal cerebral ischemia in mice. J. Pineal Res. 2008, 45, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nava, F.; Carta, G. Melatonin reduces anxiety induced by lipopolysaccharide in the rat. Neurosci. Lett. 2001, 307, 57–60. [Google Scholar] [CrossRef]
- Atanasova, M.; Petkova, Z.; Pechlivanova, D.; Dragomirova, P.; Blazhev, A.; Tchekalarova, J. Strain dependent effects of long-term treatment with melatonin on kainic acid-induced status epilepticus, oxidative stress and the expression of heat shock proteins. Pharmacol. Biochem. Behav. 2013, 111, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.; Petkova, Z.; Pechlivanova, D.; Moyanova, S.; Kortenska, L.; Mitreva, R.; Lozanov, V.; Atanasova, D.; Lazarov, N.; Stoynev, A. Prophylactic treatment with melatonin after status epilepticus: Effects on epileptogenesis, neuronal damage, and behavioral changes in a kainite model of temporal lobe epilepsy. Epilepsy Behav. 2013, 27, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Petkova, Z.; Tchekalarova, J.; Pechlivanova, D.; Moyanova, S.; Kortenska, L.; Mitreva, R.; Popov, D.; Markova, P.; Lozanov, V.; Atanasova, D.; et al. Treatment with melatonin after status epilepticus attenuates seizure activity and neuronal damage but does not prevent the disturbance in diurnal rhythms and behavioral alterations in spontaneously hypertensive rats in kainate model of temporal lobe epilepsy. Epilepsy Behav. 2014, 31, 198–208. [Google Scholar] [CrossRef] [PubMed]
- De Lima, E.; Soares, J.M., Jr.; del Carmen, S.G.Y.; Valente, S.G.; Priel, M.R.; Baracat, E.C.; Cavalheiro, E.A.; da Graça Naffah-Mazzacoratti, M.; Amado, D. Effects of pinealectomy and the treatment with melatonin on the temporal lobe epilepsy in rats. Brain Res. 2005, 1043, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Janjoppi, L.; Silva de Lacerda, A.F.; Scorza, F.A.; Amado, D.; Cavalheiro, E.A.; Arida, R.M. Influence of pinealectomy on the amygdala kindling development in rats. Neurosci. Lett. 2006, 392, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, M.; Aydin-Abidin, S.; Abidin, I.; Akca, M.; Canpolat, S.; Cansu, A. Evaluation of the role of chronic daily melatonin administration and pinealectomy on penicillin-induced focal epileptiform activity and spectral analysis of ECoG in rats: An in vivo electrophysiological study. Neurochem. Res. 2013, 38, 1672–1685. [Google Scholar] [CrossRef] [PubMed]
- Sandyk, R.; Tsagas, N.; Anninos, P.A. Melatonin as a proconvulsant hormone in humans. Int. J. Neorosci. 1992, 63, 125–135. [Google Scholar] [CrossRef]
- Peled, N.; Shorer, Z.; Peled, E.; Pillar, G. Melatonin effect on seizures in children with severe neurologic deficit disorders. Epilepsia 2001, 42, 1208–1210. [Google Scholar] [CrossRef] [PubMed]
- Aygün, H.; Aydin, D.; Inanir, S.; Ekici, F.; Ayyildiz, M.; Ağar, E. The effects of agomelatine and melatonin on ECoG activity of absence epilepsy model in WAG/Rij rats. Turk. J. Biol. 2015, 904–910. [Google Scholar] [CrossRef]
- Kldiashvili, S.; Melechov, M.; Morenkov, E.; Coenen, A.M.L.; van Luijtelaar, G.; Chepurnov, S.A. Effects of the sleep-wake rhythm modulating hormone melatonin on absence seizures in the EEG of rats. Sleep Wake 2001, 12, 58–61. [Google Scholar]
- Van Luijtelaar, G.; Coenen, A.M.L. Two types of electrocortical paroxysms in an inbred strain of rats. Neurosci. Lett. 1986, 70, 393–397. [Google Scholar] [CrossRef]
- Coenen, A.M.; Van Luijtelaar, E.L. Genetic animal models for absence epilepsy: A review of the WAG/Rij strain of rats. Behav. Genet. 2003, 33, 635–655. [Google Scholar] [CrossRef] [PubMed]
- Sarkisova, K.Y.; Kuznetsova, G.D.; Kulikov, M.A.; van Luijtelaar, G. Spike-wave discharges are necessary for the expression of behavioral depression-like symptoms. Epilepsia 2010, 51, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, A.B.; Weisbrot, D.M.; Nolan, E.E.; Gadow, K.D.; Vitale, S.A.; Andriola, M.R.; Lenn, N.J.; Novak, G.P.; Hermann, B.P. Symptoms of depression and anxiety in pediatric epilepsy patients. Epilepsia 1998, 39, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Vega, C.; Guo, J.; Killory, B.; Danielson, N.; Vestal, M.; Berman, R.; Martin, L.; Gonzalez, J.L.; Blumenfeld, H.; Spann, M. Symptoms of Anxiety and Depression in Childhood Absence Epilepsy. Epilepsia 2011, 52, e70–e74. [Google Scholar] [CrossRef] [PubMed]
- Zakzanis, K.K. Statistics to tell the truth, the whole truth, and nothing but the truth: Formulae, illustrative numerical examples, and heuristic interpretation of effect size analysis for neurophysiological researchers. Arch. Clin. Neuropsychol. 2001, 16, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Blumenfeld, H. Cellular and network mechanisms of spike-wave seizures. Epilepsia 2005, 9, 21–33. [Google Scholar] [CrossRef] [PubMed]
- De Butte, E.M.; Pappas, B.A. Pinealectomy causes hippocampal CA1 and CA3 cell loss: Reversal by melatonin supplementation. Neurobiol. Aging 2007, 28, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.; Nenchovska, Z.; Atanasova, D.; Lazarov, N.; Kortenska, L.; Stefanova, M.; Alova, L.; Atanasova, M. Consequences of long-term treatment with agomelatine on depressive-like behavior and neurobiological abnormalities in pinealectomized rats. Behav. Brain Res. 2016, 302, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Kanner, A.M.; Balabanov, A. Depression and epilepsy: How closely related are they? Neurology 2002, 58, S27–S39. [Google Scholar] [CrossRef] [PubMed]
- Trinka, E.; Kienpointner, G.; Unterberger, I.; Luef, G.; Bauer, G.; Doering, L.B.; Doering, S. Psychiatric comorbidity in juvenile myoclonic epilepsy. Epilepsia 2006, 47, 2086–2091. [Google Scholar] [CrossRef] [PubMed]
- Mula, M.; Schmitz, B. Depression in epilepsy: Mechanisms and therapeutic approach. Ther. Adv. Neurol. Disord. 2009, 2, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Sankar, R.; Mazarati, A. Neurobiology of depression as a comorbidity of epilepsy. In Jasper’s Basic Mechanisms of the Epilepsies, 4th ed.; Noebels, J.L., Avoli, M., Rogawski, M.A., Olsen, R.W., Delgado-Escueta, A.V., Eds.; Oxford University Press: Bethesda, MD, USA, 2012; pp. 1399–1416. [Google Scholar]
- Terra, V.C.; de Paola, L.; Silvado, C.E. Are children affected by epileptic neuropsychiatric comorbidities? Epilepsy Behav. 2014, 38, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Sarkisova, K.; van Luijtelaar, G. The WAG/Rij strain: A genetic animal model of absence epilepsy with comorbidity of depression. Progr. Neuro Psychopharmacol. Biol. Psychiatry 2011, 35, 854–876. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Citraro, R.; Davoli, A.; Gallelli, L.; Di Paola, E.D.; De Sarro, G. Ameliorating effects of aripiprazole on cognitive functions and depressive-like behavior in a genetic rat model of absence epilepsy and mild-depression comorbidity. Neuropharmacology 2013, 64, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Van Luijtelaar, E.L.J.M.; Coenen, A.M.L. Circadian rhythmicity in absence epilepsy in rats. Epilepsy Res. 1988, 2, 331–336. [Google Scholar] [CrossRef]
- Quigg, M.D.; Straume, M.; Menaker, M.; Bertam, E.H., III. Temporal distribution of partial seizures: Comparison of an animal model with human partial epilepsy. Ann. Neurol. 1998, 43, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.; Pechlivanova, D.; Itzev, D.; Lazarov, N.; Markova, P.; Stoynev, A. Diurnal rhythms of spontaneous recurrent seizures and behavioural alterations of Wistar and spontaneously hypertensive rats in kainate model of epilepsy. Epilepsy Behav. 2010, 17, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Liu, X.; Manchester, L.C.; Rosales-Corral, S.; Tan, D.-X.; Madrid, J.A. Processes underlying chronodiscription and their proposed association with illness. In Chronobiology and Obesity; Garaulet, M., Ordovás, J.M., Eds.; Springer: New York, NY, USA, 2013; pp. 55–73. ISBN 978-1-4614-5082-5. [Google Scholar]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Oxenkrug, G.F.; McIntyre, I.M.; Gershon, S. Effects of pinealectomy and aging on the serum corticosterone circadian rhythm in rats. J. Pineal Res. 1984, 1, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Vale, M.I.C.; Anhê, G.F.; Borges-Silva, C.N.; Andreotti, S.; Peres, S.B.; Cipolla-Neto, J.; Lima, F.B. Pinealectomy alters adipose tissue adaptability to fasting in rats. Metabolism 2004, 53, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. Prolonged glucocorticoid exposure reduces hippocampal neuron number: Implications for aging. J. Neurosci. 1985, 5, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N.; Paula-Barbosa, M.M.; Almeida, O.F.X. Ligand and subfield specificity of corticoid-induced neuronal loss in the rat hippocampal formation. Neuroscience 1999, 89, 1079–1087. [Google Scholar] [CrossRef]
- Jobe, P.C.; Dailey, J.W.; Wernicke, J.F. A noradrenergic and serotonergic hypothesis of the linkage between epilepsy and affective disorders. Neurobiology 1999, 13, 317–356. [Google Scholar] [CrossRef]
- Kanner, A.M. Can neurobiological pathogenic mechanisms of depression facilitate the development of seizure disorders? Lancet Neurol. 2012, 11, 1093–1102. [Google Scholar] [CrossRef]
- Medel-Matus, J.S.; Shin, D.; Sankar, R.; Mazarati, A. Inherent vulnerabilities in monoaminergic pathways predict the emergence of depressive impairments in an animal model of chronic epilepsy. Epilepsia 2017, 58, e116–e121. [Google Scholar] [CrossRef] [PubMed]
- Anton-Tay, F.; Chou, C.; Anton, S.; Wurtman, R.J. Brain serotonin concentration: Elevation following intraperitoneal administration of melatonin. Science 1968, 162, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Míguez, J.M.; Martín, F.J.; Aldegunde, M. Melatonin effects on serotonin synthesis and metabolism in the striatum, nucleus accumbens, and dorsal and median raphe nuclei of rats. Neurochem. Res. 1997, 22, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-López, S.; Mahar, I.; Bambico, F.R.; Labonté, B.; Ochoa-Sánchez, R.; Leyton, M.; Gobbi, G. Short-term effects of melatonin and pinealectomy on serotonergic neuronal activity across the light–dark cycle. J. Psychopharmacol. 2012, 26, 830–844. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.; Moyanova, S.; Fusco, A.; Ngomba, R.T. The role of the melatoninergic system in epilepsy and comorbid psychiatric disorders. Brain Res. Bull. 2015, 119, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, R.A.; Reiter, R.J. Rapid pinealectomy in hamsters and other small rodents. Anat. Record. 1965, 24, 83–89. [Google Scholar] [CrossRef]
- Mazarati, A.; Siddarth, P.; Baldwin, R.A.; Shin, D.; Caplan, R.; Sankar, R. Depression after status epilepticus: Behavioural and biochemical deficits and effects of fluoxetine. Brain 2008, 131, 2071–2083. [Google Scholar] [CrossRef] [PubMed]
- Becker, L.A. Effect Size (ES). Available online: https://www.uccs.edu/lbecker/effect-size (assessed on 24 June 2018).





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyanova, S.; De Fusco, A.; Santolini, I.; Celli, R.; Bucci, D.; Mastroiacovo, F.; Battaglia, G.; Nicoletti, F.; Tchekalarova, J. Abnormal Hippocampal Melatoninergic System: A Potential Link between Absence Epilepsy and Depression-Like Behavior in WAG/Rij Rats? Int. J. Mol. Sci. 2018, 19, 1973. https://doi.org/10.3390/ijms19071973
Moyanova S, De Fusco A, Santolini I, Celli R, Bucci D, Mastroiacovo F, Battaglia G, Nicoletti F, Tchekalarova J. Abnormal Hippocampal Melatoninergic System: A Potential Link between Absence Epilepsy and Depression-Like Behavior in WAG/Rij Rats? International Journal of Molecular Sciences. 2018; 19(7):1973. https://doi.org/10.3390/ijms19071973
Chicago/Turabian StyleMoyanova, Slavianka, Antonio De Fusco, Ines Santolini, Roberta Celli, Domenico Bucci, Federica Mastroiacovo, Giuseppe Battaglia, Ferdinando Nicoletti, and Jane Tchekalarova. 2018. "Abnormal Hippocampal Melatoninergic System: A Potential Link between Absence Epilepsy and Depression-Like Behavior in WAG/Rij Rats?" International Journal of Molecular Sciences 19, no. 7: 1973. https://doi.org/10.3390/ijms19071973