GIS-modeling of the Range Shifts of the Sub-fossil and Extant European Pond Turtle (Emys orbicularis) in Eastern Europe in Holocene



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. GIS Modelling
- BIO1 = Annual Mean Temperature
- BIO2 = Mean Diurnal Range (Mean of monthly (max temp - min temp))
- BIO3 = Isothermality (BIO2/BIO7) (* 100)
- BIO4 = Temperature Seasonality (standard deviation *100)
- BIO5 = Max Temperature of Warmest Month
- BIO6 = Min Temperature of Coldest Month
- BIO7 = Temperature Annual Range (BIO5-BIO6)
- BIO8 = Mean Temperature of Wettest Quarter
- BIO9 = Mean Temperature of Driest Quarter
- BIO10 = Mean Temperature of Warmest Quarter
- BIO11 = Mean Temperature of Coldest Quarter
- BIO12 = Annual Precipitation
- BIO13 = Precipitation of Wettest Month
- BIO14 = Precipitation of Driest Month
- BIO15 = Precipitation Seasonality (Coefficient of Variation)
- BIO16 = Precipitation of Wettest Quarter
- BIO17 = Precipitation of Driest Quarter
- BIO18 = Precipitation of Warmest Quarter
- BIO19 = Precipitation of Coldest Quarter
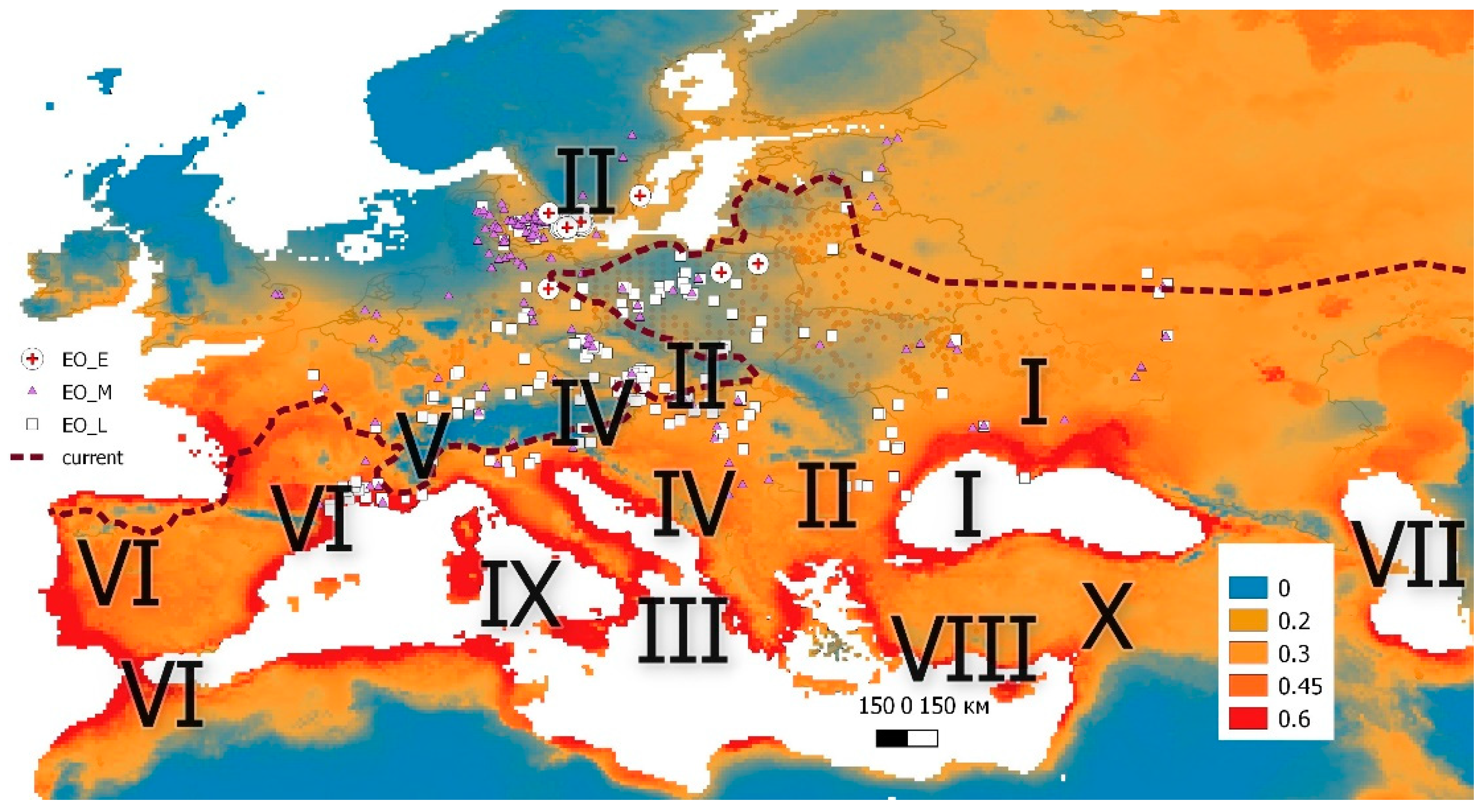
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fritz, U. Zur innerartlichen variabilität von Emys orbicularis (Linnaeus, 1758). 5b. Intraspezifische Hierarchie und Zoogeographie (Reptilia: Testudines: Emydidae). Zool. Abh. Staatl. Mus. Tierkd. Dresd. 1996, 49, 31–71. [Google Scholar]
- Fritz, U. Emys orbicularis (Linnaeus, 1758)—Europäische Sumpfschildkröte. In Handbuch der Reptilien und Amphibien Europas; Fritz, U., Schildkröten, I., Eds.; Aula-Verlag: Wiebelsheim, Germany, 2001; pp. 343–515. [Google Scholar]
- Fritz, U.; Laufer, H. Europaische Sumpfschildkrote, Emys orbicularis (Linnaeus, 1758). In Die Amphibien und Reptilien Baden-Wurttembergs; Laufer, H., Fritz, C., Sowig, P., Eds.; Ulmer: Stuttgart, Germany, 2007; pp. 511–524. [Google Scholar]
- Sommer, R.S.; Fritz, U.; Seppä, H.; Ekström, J.; Persson, A.; Liljegren, R. When the pond turtle followed the reindeer: Effect of the last extreme global warming event on the timing of faunal change in Northern Europe. Glob. Chang. Biol. 2011, 17, 2049–2053. [Google Scholar] [CrossRef]
- Berdnikovs, S. The Holocene history of the European pond tortoise (Emys orbicularis L.) in the East Baltic Area. Proc. Latv. Acad. Sci. 1999, 53, 87–92. [Google Scholar]
- Sommer, R.S.; Persson, A.; Wiesecke, N.; Fritz, U. Holocene recolonization and extinction history of the pond turtle, Emys orbicularis (L., 1758), in Europe. Quat. Sci. Rev. 2007, 26, 3099–3107. [Google Scholar] [CrossRef]
- Sommer, R.S.; Lindqvist, C.; Persson, A.; Bringsøe, H.; Rhodin, A.G.J.; Schneeweiss, N.; Široký, P.; Bachmann, L.; Fritz, U. Unexpected early extinction of the European pond turtle (Emys orbicularis) in Sweden and climatic impact on its Holocene range. Mol. Ecol. 2009, 18, 1252–1262. [Google Scholar] [CrossRef] [PubMed]
- Bakharev, V.A. The history of the area Emys orbicularis (Testudines, Emydidae) and the current state of its borders within Belarus. Praci Ukrainskogo Gerpetologichnogo Tovaristva 2014, 5, 12–19. (In Russian) [Google Scholar]
- Rosengren, E. The colonisation history of the Scandinavian fauna presented through subfossil finds along one of Its major Immigration Routes; Scania, Southern Sweden. Environ. Ecol. Res. 2015, 3, 143–149. [Google Scholar] [CrossRef]
- Fritz, U. Die Europaische Sumpfschildkrote (Emys orbicularis); Laurenti Verlag: Bielefeld, Germany, 2003. [Google Scholar]
- Kleewein, A.; Wöss, G. Status of the European pond turtle, Emys orbicularis (Reptilia: Testudines: Emydidae) in Vorarlberg, Austria. Acta Herpetol. 2013, 8, 65–67. [Google Scholar]
- Vamberger, M.; Fritz, U. Big data can cause big mistakes: Using the Societas Europaea Herpetologica atlas by Sillero et al. (2014), the distribution of Emys orbicularis will be misunderstood. Biologia 2018, 73, 281–283. [Google Scholar] [CrossRef]
- Tatarinov, K.A. Pliocene-anthropogenic and prescription fauna of amphibians of reptiles of Volyn Polesie, Podolia and eastern Carpathians. In Voprosy Gerpetologii; Nauka: Moscow, Russia, 1973; pp. 176–177. (In Russian) [Google Scholar]
- Askeev, I.V.; Askeev, O.V.; Galimova, D.N. Vertebrates, the natural environment and man in the Holocene of the Volga-Kama region. In Proceedings of the Materials of the Second Scientific Conference Dynamics of Ecosystems in the Holocene, Chelyabinsk, Russia, 12–14 October 2010; Smirnov, N.G., Ed.; Rifey: Yekaterinburg, Russia, 2010; pp. 20–24. (In Russian). [Google Scholar]
- Meeske, A.C.M.; Pupins, M.; Rybczynski, K. Erste Ergebnisse zur Verbreitung und zum Status der Europäischen Sumpfschildkröte (Emys orbicularis) am nördlichen Rand ihrer Verbreitung in Litauen und Lettland. Zeitschr. Feldherpetol. 2006, 13, 71–99. [Google Scholar]
- Pupins, M.; Pupina, A. Distribution of European pond turtle Emys orbicularis (LINNAEUS, 1758) on the northern edge of its area in Latvia. Rev. Esp. Herpetol. 2008, 22, 149–157. [Google Scholar]
- Pupins, M.; Pupina, A. The data on the observations of the European pond turtle (Emys orbicularis L.) at the northern edge of its area in Latvia. Acta Biol. Univ. Daugavp. 2008, 8, 35–46. [Google Scholar]
- Pupins, M.; Pupina, A.; Bakharev, V.A. The spread of the European pond turtle (Emys orbicularis L., 1758) in Latvia and in the territories bordering Belarus. Vesn. Mazyrsk. dzyarzh. ped. ўnіv. IP Shamyakіna 2010, 1, 35–38. (In Russian) [Google Scholar]
- Pupins, M.; Pupina, A. Updated distribution of the European Pond Turtle, Emys orbicularis (L., 1758) (Emydidae) on the extreme northern border of its European range in Latvia. Acta Zool. Bulg. 2017, 10, 133–137. [Google Scholar]
- Široký, P.; Stuchlík, S.; Fritz, U.; Moravec, J. Basic morphological data of native Czech Emys orbicularis revealed by subfossil finds. Biologia 2009, 64, 795–797. [Google Scholar] [CrossRef]
- GBIF.org Emys Orbicularis (Linnaeus, 1758) in GBIF Secretariat (2017). GBIF Backbone Taxonomy. Checklist Dataset. Available online: https://doi.org/10.15468/dl.62qcmk (accessed on 9 March 2019).
- Drobenkov, S.M. Population Ecology of the European Pond Turtle (Emys orbicularis) in Belarus; Belarusskaja Nauka: Minsk, Belarus, 2012. (In Russian) [Google Scholar]
- Drobenkov, S.M. Current state, anthropogenic threats and conservation of the European pond turtle (Emys orbiclularis) in Belarus. Acta Biol. Univ. Daugavp. 2014, 14, 29–36. [Google Scholar]
- Bakiev, A.G. Emys orbicularis bog turtle in the Samara region (regional summary). In Actual Problems of Herpetology and Toxicology (Collection of Scientific Papers); IEVB RAN: Tolyatti, Russia, 2001; Volume 5, pp. 3–18. (In Russian) [Google Scholar]
- Zavialov, E.V.; Tabachishin, V.G.; Shlyakhtin, G.V. Recent distribution habitat of reptiles in the North of Low Volga region. Sovr. gerpetol. 2004, 2, 54. (In Russian) [Google Scholar]
- Khabibullin, V. Distribution of Emys orbicularis in the South Urals, Russia. Biol. Bratislava. 2004, 59, 27–32. [Google Scholar]
- Borisova, N.; Urykina, L. About new findings of the European magnetic turtle—Emys orbicularis (Linnaeus, 1758) (Reptilia, Chelonii, Emydidae)—In the Chuvash republic. In Natural Science Research in Chuvashia: Materials of Reports of the Regional Scientific-Practical Conference (Cheboksary, 21 November 2017); Plakat: Cheboksary, Russia, 2017; Volume 4, p. 80. (In Russian) [Google Scholar]
- Dujsebayeva, T.; Brushko, Z.; Saraev, F.; Bidashko, F. Distribution of the marsh turtle Emys orbicularis (Linnaeus, 1758) on the eastern periphery of the species (Kazakhstan) Spatial Time Dynamics Biota and Ecosystems Aralo-Caspian Pool. In Proceedings of the II International Conference, Orenburg, Russia, 9–13 October 2017; pp. 174–183. (In Russian). [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; En Chee, Y.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Brown, J.L.; Hill, D.J.; Dolan, A.M.; Carnaval, A.C.; Haywood, A.M. PaleoClim, high spatial resolution paleoclimate surfaces for global land areas. Sci. Data 2018, 5, 180254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekrasova, O.D.; Oskikro, O.S. Significant areas of the Steppe zone of Ukraine and their importance in preservation of the turtle Emys orbicularis (Linnaeus, 1758). In Proceedings of the Scientific-Practical Conference Protected Case in the Steppe Zone of Ukraine (for the 90th Anniversary of Creation of the Admiralty Reserve), Donetsk Region, Ukraine, 14–15 March 2017; Volume 2, pp. 264–269. [Google Scholar]
- Varushchenko, S.I. Analysis of the Late Pleistocene and Holocene history of the development of the natural environment of the northwestern shelf of the Black Sea. In The Fluctuations of the World Ocean Level and Issues of Marine Geomorphology; Nauka: Moscow, Russia, 1975; pp. 50–62. (In Russian) [Google Scholar]
- Lenk, P.; Fritz, U.; Joger, U.; Wink, M. Mitochondrial phylogeography of the European pond turtle, Emys orbicularis (Linnaeus 1758). Mol. Ecol. 1999, 8, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Fritz, U.; Ayaz, D.; Hundsdörfer, A.K.; Kotenko, T.; Guicking, D.; Wink, M.; Tok, C.V.; Çiçek, K.; Buschbom, J. Mitochondrial diversity of European pond turtles (Emys orbicularis) in Anatolia and the Ponto-Caspian region: Multiple old refuges, hotspot of extant diversification and critically endangered endemics. Organ. Divers. Evol. 2009, 9, 100–114. [Google Scholar] [CrossRef]
- Stuckas, H.; Velo-Antón, G.; Fahd, S.; Kalboussi, M.; Rouag, R.; Arculeo, M.; Marrone, F.; Sacco, F.; Vamberger, M.; Fritz, U. Where are you from, stranger? The enigmatic biogeography of North African pond turtles (Emys orbicularis). Organ. Divers. Evol. 2014, 14, 295–306. [Google Scholar] [CrossRef]
- Velo-Antón, G.; Wink, M.; Schneeweiss, N.; Fritz, U. Native or not? Tracing the origin of wild-caught and captive freshwater turtles in a threatened and widely distributed species (Emys orbicularis). Conserv. Genet. 2011, 12, 583–588. [Google Scholar] [CrossRef]
- Pedall, I.; Fritz, U.; Stuckas, H.; Valdeón, A.; Wink, M. Gene flow across secondary contact zones of the Emys orbicularis complex in the Western Mediterranean and evidence for extinction and re-introduction of pond turtles on Corsica and Sardinia (Testudines: Emydidae). J. Zool. Syst. Evol. Res. 2011, 49, 44–57. [Google Scholar] [CrossRef]
- Pöschel, J.; Heltai, B.; Graciá, E.; Quintana, M.F.; Velo-Antón, G.; Arribas, O.; Valdeón, O.; Wink, M.; Fritz, U.; Vamberger, M. Complex hybridization patterns in European pond turtles (Emys orbicularis) in the Pyrenean Region. Sci. Rep. 2018, 8, 15925. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Silins, J.; Lamsters, V. Latvian Reptiles and Amphibians; Valters un Rapa: Riga, Latvia, 1934; p. 96. (In Latvian) [Google Scholar]
- Pupins, M. First report on recording of the invasive species Trachemys scripta elegans, a potential competitor of Emys orbicularis in Latvia. Acta Univ. Latv. 2007, 273, 37–46. [Google Scholar]
- Kuybida, V.V.; Nekrasova, O.D.; Kutsokon, Y.K.; Lopatynska, V.V.; Truskavetska, I.Ya. Summer Fish Kills in the Kaniv Reservoir. Hydrobiol. J. 2019, 55, 103–106. [Google Scholar] [CrossRef]
- Pupins, M.; Pupina, A. First records of 5 allochthonous species and subspecies of turtles (Trachemys scripta troostii, Mauremys caspica, Mauremys rivulata, Pelodiscus sinensis, Testudo horsfieldii) and new records of subspecies Trachemys scripta elegans in Latvia. Manag. Biol. Invas. 2011, 2, 69–81. [Google Scholar] [CrossRef]
- Pupina, A.; Pupins, M. First records of new aquatic predator Pelodiscus sinensis (Wiegmann 1835) in Latvia and preliminary ecological risk assessment of the invasion for autochthonic Emys orbicularis (Linnaeus 1758). Acta Biol. Univ. Daugavp. 2016, 16, 61–76. [Google Scholar]
- Tytar, V.; Nekrasova, O.; Pupina, A.; Pupins, M.; Oskyrko, O. Long-term bioclimatic modelling the distribution of the Fire-Bellied toad, Bombina bombina, under the influence of global climate change. Vest. Zool. 2018, 52, 553–556. [Google Scholar] [CrossRef]
- Nekrasova, O.; Tytar, V.; Pupins, M. Bioclimatic limitations to the geographical distribution of Emys orbicularis (Linnaeus, 1758) (Testudines: Emydidae) in the North of the range. In Book of abstracts. XV Portuguese-Spanish Herpetology Congress and XIX Spanish Congress of Herpetology. Biology and Conservation of Herps in the Anthropocene; Congreso de Herpetologia: Salamanca, Spain, 2018; p. 106. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
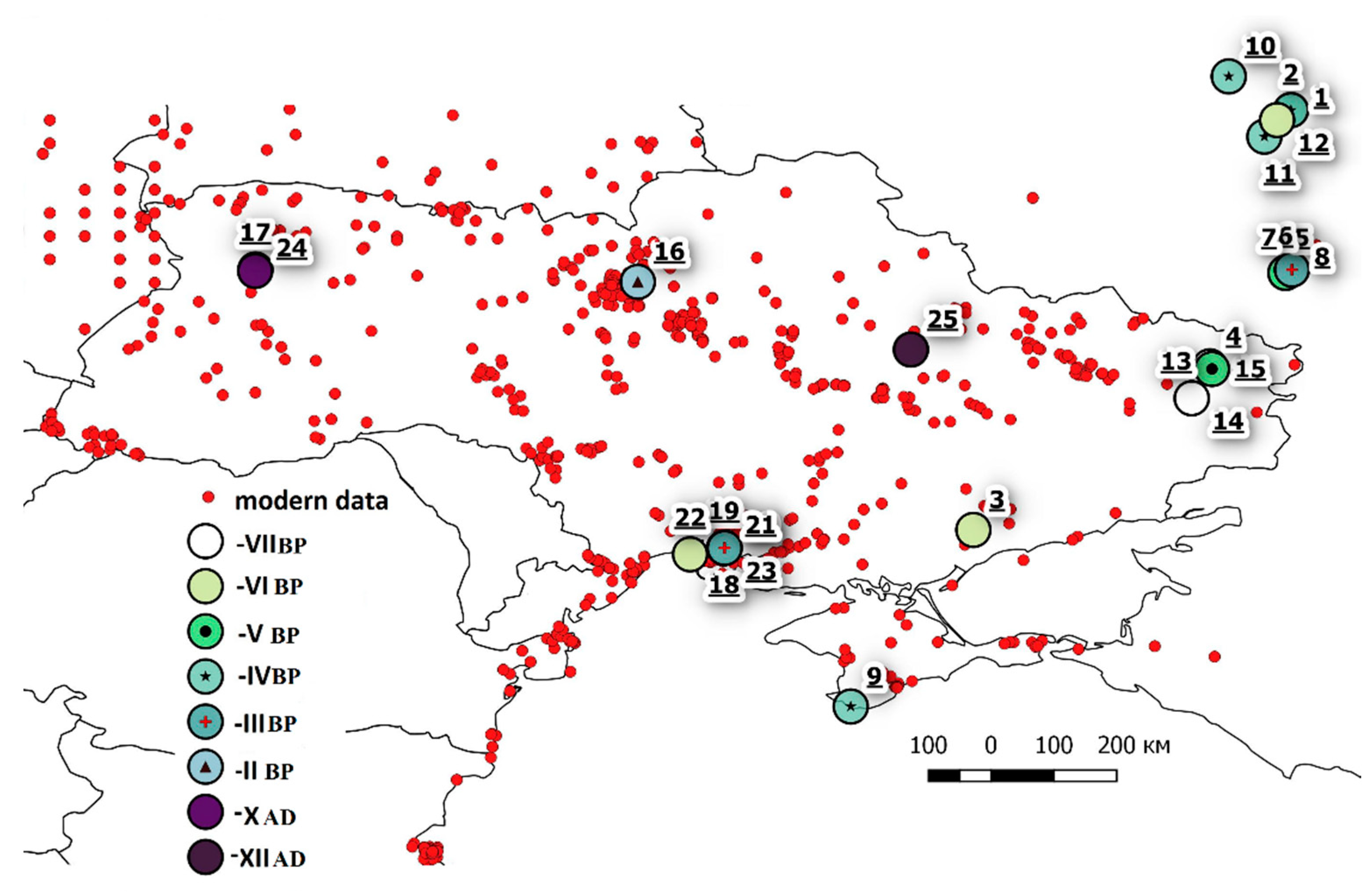
| No | Archaeological Monument | Expeditions No. * | Bones/Individuals | Coordinates | Data | |
|---|---|---|---|---|---|---|
| 1 | Vasil’yevskiy kordon-17, Lipetsk region, Russia | 1, expedition leader R.V. Smoljaninov, A.A. Sviridov | 290/243 | 52.978 | 40.010 | IV cent. BC |
| 2 | Vasil’yevskiy kordon-27, Lipetsk region, Russia | 1, expedition leader R.V. Smoljaninov, S.V.S. Hemenev, A.V. Solovev | 78/70 | 52.977 | 40.010 | IV cent. BC |
| 3 | Kamennaya mogila, Zaporozhskaya region, Ukraine | 2, expedition leader N.S. Kotova | 2 | 46.949 | 35.465 | VI cent. BC |
| 4 | Starobelsk-I, Lugansk region, Ukraine | 3, expedition leader Ju.G.Gurin, K.I. Krasil’nikov, staff Voroshilovgrad Pedagogical Institute T.G. Shevchenko with the participation of O. Shaposhnikov, an employee of the IA of the AS of the USSR | 6/1 | 49.297 | 38.849 | 6954±35 ВР |
| 5 | Cherkassky-3, Voronezh region, Russia | 4, expedition leader A.M. Skorobogatov | 80/80 | 50.6407 | 39.932 | V-VII cent. BC |
| 6 | Cherkassky-3, bottom layer, Voronezh region, Russia | 4, expedition leader A.M. Skorobogatov | 24/24 | 50.640 | 39.932 | 6570-6398 (2σ) to 4710-4540 CalBC |
| 7 | Cherkassky-5, Voronezh region, Russia | 5, expedition leader A.M. Skorobogatov | 28/28 | 50.640 | 39.932 | 6249-5389 CalBC (2σ) |
| 8 | vil. Losevo, Voronezh region, Russia | 4, expedition leader V.D. Berezuckij | 1/1 | 50.686 | 40.024 | III cent. BC |
| 9 | Laspi-I, Crimea, Ukraine | 6, expedition leader D.Ja. Telegin | 4/1 | 44.413 | 33.711 | Middle IV cent. BC |
| 10 | Dolgoe-I, Lipetsk region, Russia | 7, expedition leader V.P. Levenok | 8/8 | 53.461 | 39.120 | Middle IV cent. BC |
| 11 | Lipetsk Lake, Lipetsk region, Russia | 8, A.Yu. Klokov | 2/2 | 52.596 | 39.630 | IV cent. BC |
| 12 | Dobroe-9, Lipetsk region, Russia | 1, expedition leader A.A. Kulichkov | 1/1 | 52.834 | 39.812 | VI cent. BC |
| 13 | Tuba-V, Lugansk region, Ukraine | 9, expedition leader S.A. Telizhenko | 748/748 | 48.840 | 38.590 | 2nd half VII cent. BC |
| 14 | Tuba-Vb, Lugansk region, Ukraine | 9, S.A. Telizhenko | 272/272 | 48.839 | 38.590 | 2nd half VII cent. BC |
| 15 | Novoselovka-3, Lugansk region, Ukraine | 9, expedition leader S.A. Telizhenko | 1/1 | 49.260 | 38.881 | 5200-5050 BC |
| 16 | Troeshchina-6, Kiev region, Ukraine | 10, V. Kozyuba | 16/1 | 50.511 | 30.663 | II cent. BC |
| 17 | Kolyada, Volyn region, Ukraine | 11, expedition leader S.A. Telizhenko | 3/3 | 50.6892 | 25.191 | III-II cent. BC |
| 18 | Olviya, 2016, semi-dugout 16, Nikolaev region, Ukraine | 12, expedition leader from Ukraine A.V. Bujskix, from Germany J. Fornassier | 10/1 | 46.691 | 31.899 | VI-V cent. BC |
| 19 | Olviya, 2015, Necropolis-4, Nikolaev region, Ukraine | 12, expedition leader from Ukraine—A.V. Bujskix, from Germany J. Fornassier | 1/1 | 46.691 | 31.899 | 1st quarter V cent. BC |
| 20 | Olvia, 2015, cistern, Nikolaev region, Ukraine | 12, expedition leader A.V. Bujskix | 1/1 | 46.688 | 31.905 | V cent. BC |
| 21 | Olvia, 2016, pit 22, Nikolaev region, Ukraine | 12, expedition leader from Ukraine A.V. Bujskix, from Germany J. Fornassier | 1/1 | 46.691 | 31.899 | 2nd half V–III cent. BC |
| 22 | Berezan Island, Nikolaev region, Ukraine | 12, expedition leader A.V. Bujskix, V.V. Krutilov | 8/8 | 46.598 | 31.412 | VII-VI cent. BC–V cent. BC |
| 23 | Olvia, 2018, R-23, Nikolaev region, Ukraine | 12, expedition leader from Ukraine—A.V. Bujskix, from Poland A. Tvardeczkyj | 2/2 | 46.689 | 31.906 | III-IV cent. BC |
| 24 | Ratnov, Volyn region, Ukraine | 11, expedition leader S.A. Telizhenko | 3/3 | 50.668 | 25.188 | X cent. AD |
| 25 | Glinskoe, Poltava region, Ukraine | 13, expedition leader Yu.O. Pugolovok | 33/1 | 49.538 | 34.571 | X-XIV cent. AD |
| Total | 1623/1504 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekrasova, O.; Yanish, Y.; Tytar, V.; Pupins, M. GIS-modeling of the Range Shifts of the Sub-fossil and Extant European Pond Turtle (Emys orbicularis) in Eastern Europe in Holocene. Diversity 2019, 11, 121. https://doi.org/10.3390/d11080121
Nekrasova O, Yanish Y, Tytar V, Pupins M. GIS-modeling of the Range Shifts of the Sub-fossil and Extant European Pond Turtle (Emys orbicularis) in Eastern Europe in Holocene. Diversity. 2019; 11(8):121. https://doi.org/10.3390/d11080121
Chicago/Turabian StyleNekrasova, Oksana, Yevheniia Yanish, Volodymyr Tytar, and Mihails Pupins. 2019. "GIS-modeling of the Range Shifts of the Sub-fossil and Extant European Pond Turtle (Emys orbicularis) in Eastern Europe in Holocene" Diversity 11, no. 8: 121. https://doi.org/10.3390/d11080121