

Microbial Diversity in Bee Species and Bee Products: Pseudomonads Contribution to Bee Well-Being and the Biological Activity Exerted by Honey Bee Products: A Narrative Review

Abstract
:1. Introduction
2. Bee Species and Their Products
2.1. Bee Species
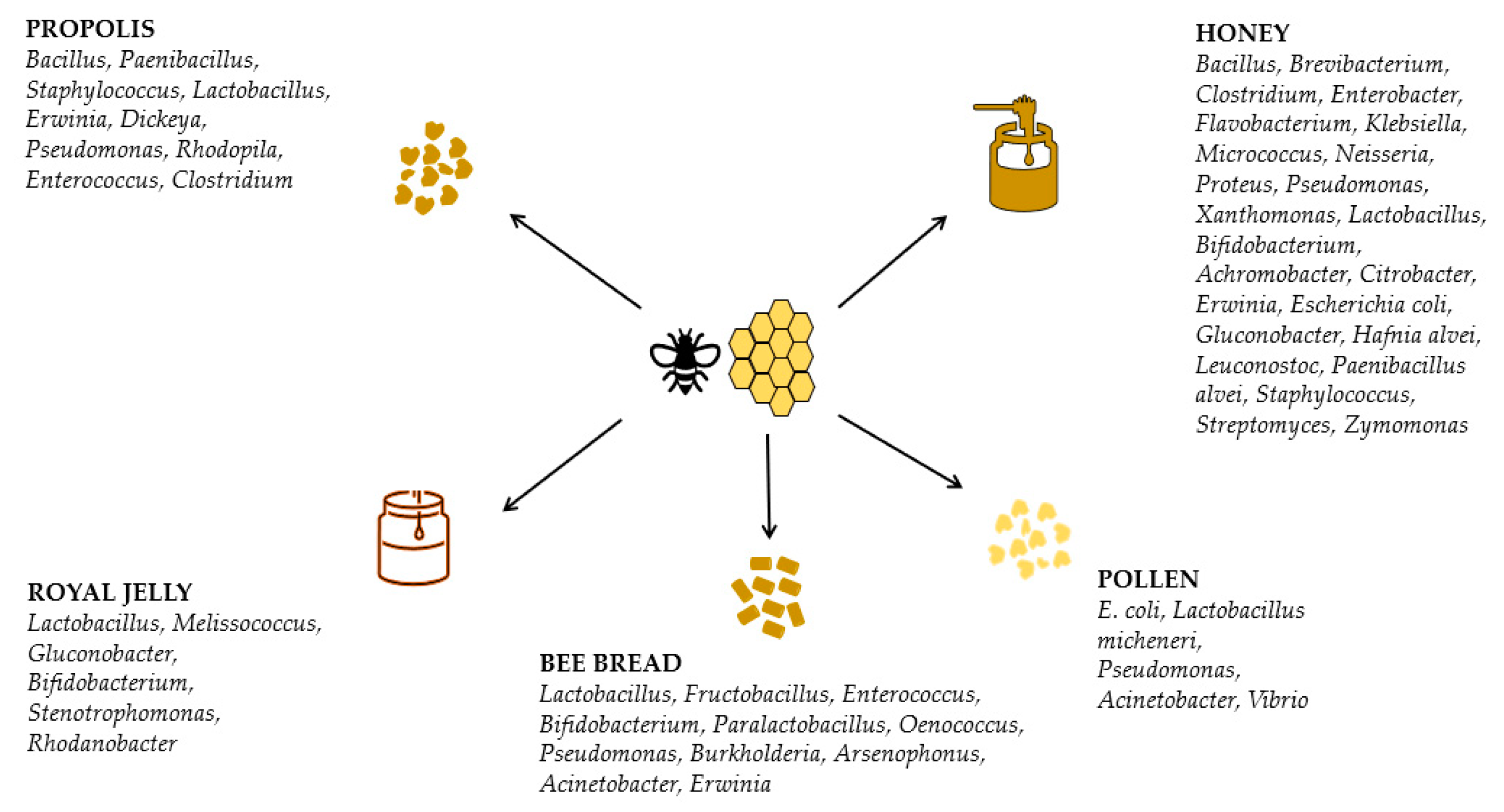
2.2. Honey
2.3. Bee-Collected Pollen and Bee Bread
2.4. Propolis
2.5. Royal Jelly
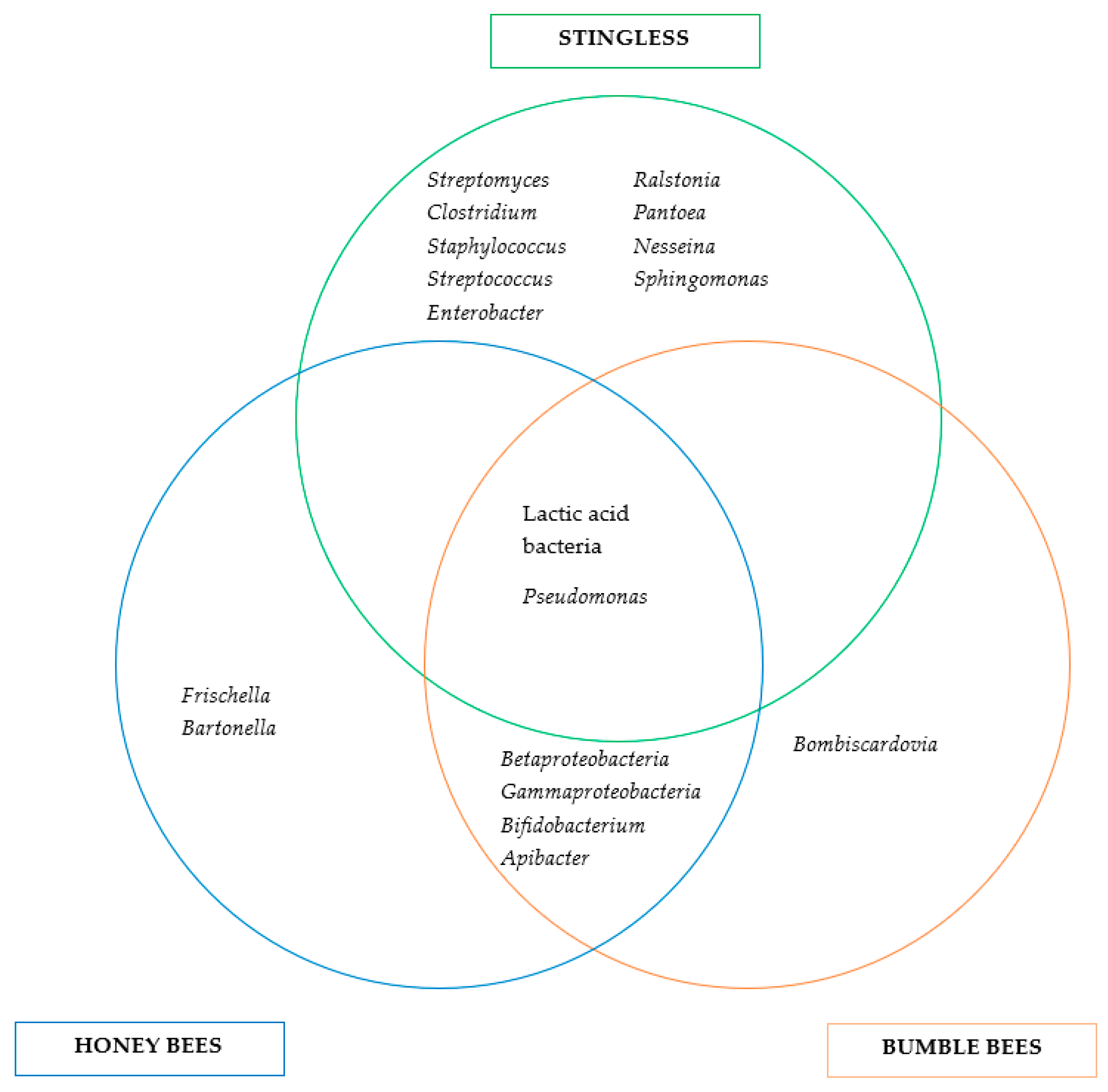
3. Microbiomes of Bee Species and Bee Products
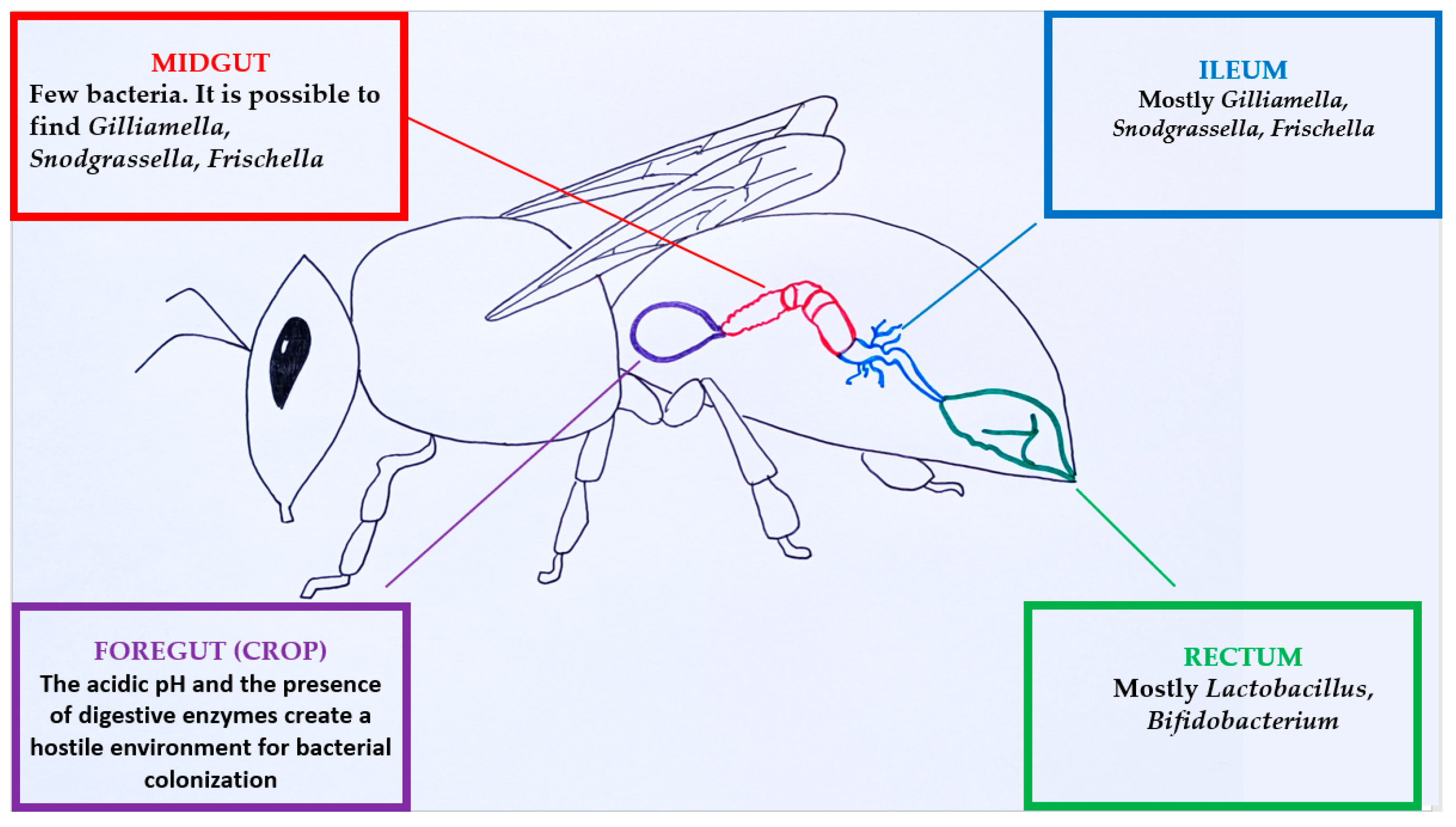
3.1. Bee Microbiome
3.2. Honey Microbiome
3.3. Bee-Collected Pollen and Bee Bread Microbiome
3.4. Propolis Microbiome
3.5. Royal Jelly Microbiome
4. Contribution of Pseudomonads to Bee Well-Being and the Biological Activity Exerted by Honey Bee Products

{kind=link}
{kind=link}
{kind=link}
| Pseudomonas Function in Bees and Honey Bee Products | Authors | |
|---|---|---|
| Social bees | Catabolize glyphosate and use it as an additional carbon source | Zhao et al., 2015 [141] |
| Contributing to resistance against insecticides such as flumethrin | Yu et al., 2021 [130] | |
| Fight pathogens by producing secondary metabolites | Khan et al., 2020 [30] | |
| Bumble bee queens | Assist queens in enduring harsh environmental conditions and prepare them for hibernation, thus allowing them to reproduce | Wang et al., 2019 [41] |
| Honey bee products | Preserve honey bee products against spoilage microorganisms | Tsadila et al., 2021 [57] |
| Pseudomonads are implicated in the conversion of bee pollen to bee bread | Barta et al., 2022 [148] Disayathanoowat et al., 2020 [26] | |
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crotti, E.; Sansonno, L.; Prosdocimi, E.M.; Vacchini, V.; Hamdi, C.; Cherif, A.; Gonella, E.; Marzorati, M.; Balloi, A. Microbial Symbionts of Honeybees: A Promising Tool to Improve Honeybee Health. N. Biotechnol. 2013, 30, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Ribière, C.; Hegarty, C.; Stephenson, H.; Whelan, P.; O’Toole, P.W. Gut and Whole-Body Microbiota of the Honey Bee Separate Thriving and Non-Thriving Hives. Microb. Ecol. 2019, 78, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Castelli, L.; Branchiccela, B.; Romero, H.; Zunino, P.; Antúnez, K. Seasonal Dynamics of the Honey Bee Gut Microbiota in Colonies Under Subtropical Climate: Seasonal Dynamics of Honey Bee Gut Microbiota. Microb. Ecol. 2022, 83, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial Ecology of the Hive and Pollination Landscape: Bacterial Associates from Floral Nectar, the Alimentary Tract and Stored Food of Honey Bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.J.; Rusch, D.B.; Stewart, F.J.; Mattila, H.R.; Newton, I.L.G. Saccharide Breakdown and Fermentation by the Honey Bee Gut Microbiome. Environ. Microbiol. 2015, 17, 796–815. [Google Scholar] [CrossRef]
- Donkersley, P.; Rhodes, G.; Pickup, R.W.; Jones, K.C.; Wilson, K. Bacterial Communities Associated with Honeybee Food Stores Are Correlated with Land Use. Ecol. Evol. 2018, 8, 4743–4756. [Google Scholar] [CrossRef]
- Corby-Harris, V.; Maes, P.; Anderson, K.E. The Bacterial Communities Associated with Honey Bee (Apis mellifera) Foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef]
- Anderson, K.E.; Rodrigues, P.A.P.; Mott, B.M.; Maes, P.; Corby-Harris, V. Ecological Succession in the Honey Bee Gut: Shift in Lactobacillus Strain Dominance during Early Adult Development. Microb. Ecol. 2016, 71, 1008–1019. [Google Scholar] [CrossRef]
- Papp, M.; Békési, L.; Farkas, R.; Makrai, L.; Judge, M.F.; Maróti, G.; Tozsér, D.; Solymosi, N. Natural Diversity of the Honey Bee (Apis mellifera) Gut Bacteriome in Various Climatic and Seasonal States. PLoS ONE 2022, 17, e0273844. [Google Scholar] [CrossRef]
- Romero, S.; Nastasa, A.; Chapman, A.; Kwong, W.K.; Foster, L.J. The Honey Bee Gut Microbiota: Strategies for Study and Characterization. Insect Mol. Biol. 2019, 28, 455–472. [Google Scholar] [CrossRef]
- Dai, P.; Yan, Z.; Ma, S.; Yang, Y.; Wang, Q.; Hou, C.; Wu, Y.; Liu, Y.; Diao, Q. The Herbicide Glyphosate Negatively Affects Midgut Bacterial Communities and Survival of Honey Bee during Larvae Reared in Vitro. J. Agric. Food Chem. 2018, 66, 7786–7793. [Google Scholar] [CrossRef]
- Ricigliano, V.A.; Williams, S.T.; Oliver, R. Effects of Different Artificial Diets on Commercial Honey Bee Colony Performance, Health Biomarkers, and Gut Microbiota. BMC Vet. Res. 2022, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.R.; Truong, A.T.; Choi, Y.S.; Lee, M.Y.; Kim, B.Y.; Seo, M.; Yoon, S.S.; Yoo, M.S.; Van Quyen, D.; Cho, Y.S. Comparison of the Gut Microbiome of Sacbrood Virus-Resistant and -Susceptible Apis cerana from South Korea. Sci. Rep. 2022, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Dong, Y.; Gu, C.; Zhang, X.; Ma, H. Processing Technologies for Bee Products: An Overview of Recent Developments and Perspectives. Front. Nutr. 2021, 8, 834. [Google Scholar] [CrossRef] [PubMed]
- Kafantaris, I.; Amoutzias, G.D.; Mossialos, D. Foodomics in Bee Product Research: A Systematic Literature Review. Eur. Food Res. Technol. 2020, 247, 309–331. [Google Scholar] [CrossRef]
- Brudzynski, K. Honey as an Ecological Reservoir of Antibacterial Compounds Produced by Antagonistic Microbial Interactions in Plant Nectars, Honey and Honey Bee. Antibiotics 2021, 10, 551. [Google Scholar] [CrossRef]
- Saccà, M.L.; Bianchi, G.; Scalzo, R. Lo Biosynthesis of 2-Heptanone, a Volatile Organic Compound with a Protective Role against Honey Bee Pathogens, by Hive Associated Bacteria. Microorganisms 2021, 9, 2218. [Google Scholar] [CrossRef]
- Bonilla-Rosso, G.; Engel, P. Functional Roles and Metabolic Niches in the Honey Bee Gut Microbiota. Curr. Opin. Microbiol. 2018, 43, 69–76. [Google Scholar] [CrossRef]
- de Sousa, L.P. Bacterial Communities of Indoor Surface of Stingless Bee Nests. PLoS ONE 2021, 16, e0252933. [Google Scholar] [CrossRef]
- Degrandi-Hoff Man, G.; Eckholm, B.; Anderson, K.E. Honey Bee Health: The Potential Role of Microbes. In Honey Bee Colony Health: Challenges and Sustainable Solutions; CRC Press: Boca Raton, FL, USA, 2011; pp. 1–12. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Kaltenpoth, M. Microbial Communities of Three Sympatric Australian Stingless Bee Species. PLoS ONE 2014, 9, e105718. [Google Scholar] [CrossRef]
- Khan, S.; Somerville, D.; Frese, M.; Nayudu, M. Environmental Gut Bacteria in European Honey Bees (Apis mellifera) from Australia and Their Relationship to the Chalkbrood Disease. PLoS ONE 2020, 15, e0238252. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Ricigliano, V.A.; Copeland, D.C.; Mott, B.M.; Maes, P. Social Interaction Is Unnecessary for Hindgut Microbiome Transmission in Honey Bees: The Effect of Diet and Social Exposure on Tissue-Specific Microbiome Assembly. Invertebr. Microbiol. 2023, 85, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Casalone, E.; Cavalieri, D.; Daly, G.; Vitali, F.; Perito, B. Propolis Hosts a Diverse Microbial Community. World J. Microbiol. Biotechnol. 2020, 36, 50. [Google Scholar] [CrossRef] [PubMed]
- Pełka, K.; Worobo, R.W.; Walkusz, J.; Szweda, P. Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials. Antibiotics 2021, 10, 713. [Google Scholar] [CrossRef]
- Disayathanoowat, T.; Li, H.; Supapimon, N.; Suwannarach, N.; Lumyong, S.; Chantawannakul, P.; Guo, J. Different Dynamics of Bacterial and Fungal Communities in Hive-Stored Bee Bread and Their Possible Roles: A Case Study from Two Commercial Honey Bees in China. Microorganisms 2020, 8, 264. [Google Scholar] [CrossRef]
- Jones, J.C.; Fruciano, C.; Marchant, J.; Hildebrand, F.; Forslund, S.; Bork, P.; Engel, P.; Hughes, W.O.H. The Gut Microbiome Is Associated with Behavioural Task in Honey Bees. Insectes Soc. 2018, 65, 419–429. [Google Scholar] [CrossRef]
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Di Gioia, D. Beneficial Microorganisms for Honey Bees: Problems and Progresses. Appl. Microbiol. Biotechnol. 2016, 100, 9469–9482. [Google Scholar] [CrossRef]
- Arredondo, D.; Castelli, L.; Porrini, M.P.; Garrido, P.M.; Eguaras, M.J.; Zunino, P.; Antúnez, K. Lactobacillus kunkeei Strains Decreased the Infection by Honey Bee Pathogens Paenibacillus larvae and Nosema Ceranae. Benef. Microbes 2018, 9, 279–290. [Google Scholar] [CrossRef]
- Khan, K.A.; Al-Ghamdi, A.A.; Ghramh, H.A.; Ansari, M.J.; Ali, H.; Alamri, S.A.; Al-Kahtani, S.N.; Adgaba, N.; Qasim, M.; Hafeez, M. Structural Diversity and Functional Variability of Gut Microbial Communities Associated with Honey Bees. Microb. Pathog. 2020, 138, 103793. [Google Scholar] [CrossRef]
- Subotic, S.; Boddicker, A.M.; Nguyen, V.M.; Rivers, J.; Briles, C.E.; Mosier, A.C. Honey Bee Microbiome Associated with Different Hive and Sample Types over a Honey Production Season. PLoS ONE 2019, 14, e0223834. [Google Scholar] [CrossRef]
- EU Overview. Detailed Information on Honey Bees. Available online: https://food.ec.europa.eu/animals/live-animal-movements/honey-bees_en (accessed on 9 August 2023).
- Gkantiragas, A.G.; Gabrielli, J. A Meta-Analysis of the 16S-RRNA Gut Microbiome Data in Honeybees (Apis mellifera). bioRxiv 2021. [Google Scholar] [CrossRef]
- Kwong, W.K.; Medina, L.A.; Koch, H.; Sing, K.W.; Soh, E.J.Y.; Ascher, J.S.; Jaffé, R.; Moran, N.A. Dynamic Microbiome Evolution in Social Bees. Sci. Adv. 2017, 3, e1600513. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Moran, N.A. The Role of the Gut Microbiome in Health and Disease of Adult Honey Bee Workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey Bee Gut Microbiome Is Altered by In-Hive Pesticide Exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef]
- Kešnerová, L.; Emery, O.; Troilo, M.; Liberti, J.; Erkosar, B.; Engel, P. Gut Microbiota Structure Differs between Honeybees in Winter and Summer. ISME J. 2020, 14, 801–814. [Google Scholar] [CrossRef]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of Acquisition of the Gut Microbiota of the Honey Bee Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef]
- Kwong, W.K.; Moran, N.A. Gut Microbial Communities of Social Bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef]
- Deng, Y.; Yang, S.; Zhao, H.; Luo, J.; Yang, W.; Hou, C. Antibiotics-Induced Changes in Intestinal Bacteria Result in the Sensitivity of Honey Bee to Virus. Environ. Pollut. 2022, 314, 120278. [Google Scholar] [CrossRef]
- Wang, L.; Wu, J.; Li, K.; Sadd, B.M.; Guo, Y.; Zhuang, D.; Zhang, Z.; Chen, Y.; Evans, J.D.; Guo, J.; et al. Dynamic Changes of Gut Microbial Communities of Bumble Bee Queens through Important Life Stages. mSystems 2019, 4, 10–1128. [Google Scholar] [CrossRef]
- Rothman, J.A.; Leger, L.; Graystock, P.; Russell, K.; McFrederick, Q.S. The Bumble Bee Microbiome Increases Survival of Bees Exposed to Selenate Toxicity. Environ. Microbiol. 2019, 21, 3417–3429. [Google Scholar] [CrossRef]
- Cameron, S.A.; Sadd, B.M. Annual Review of Entomology Global Trends in Bumble Bee Health. Annu. Rev. Entomol. 2020, 65, 209–232. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.H.; Miao, C.H.; Chen, Y.F.; Dong, Z.X.; Cao, Z.; Liao, S.Q.; Wang, J.X.; Wang, Z.W.; Guo, J. The Composition of Bacteria in Gut and Beebread of Stingless Bees (Apidae: Meliponini) from Tropics Yunnan, China. Antonie Van Leeuwenhoek Int. J. General. Mol. Microbiol. 2021, 114, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.M.; Tran, L.; McKee, C.G.; Ortega Polo, R.; Newman, T.; Lansing, L.; Griffiths, J.S.; Bilodeau, G.J.; Rott, M.; Marta Guarna, M. Honey Bees as Biomonitors of Environmental Contaminants, Pathogens, and Climate Change. Ecol. Indic. 2022, 134, 108457. [Google Scholar] [CrossRef]
- Ghosh, S.; Namin, S.M.; Jung, C. Differential Bacterial Community of Bee Bread and Bee Pollen Revealed by 16s RRNA High-Throughput Sequencing. Insects 2022, 13, 863. [Google Scholar] [CrossRef]
- Powell, J.E.; Lau, P.; Rangel, J.; Arnott, R.; De Jong, T.; Moran, N.A. The Microbiome and Gene Expression of Honey Bee Workers Are Affected by a Diet Containing Pollen Substitutes. PLoS ONE 2023, 18, e0286070. [Google Scholar] [CrossRef]
- Almasri, H.; Liberti, J.; Brunet, J.L.; Engel, P.; Belzunces, L.P. Mild Chronic Exposure to Pesticides Alters Physiological Markers of Honey Bee Health without Perturbing the Core Gut Microbiota. Sci. Rep. 2022, 12, 4281. [Google Scholar] [CrossRef]
- Laconi, A.; Tolosi, R.; Mughini-Gras, L.; Mazzucato, M.; Ferrè, N.; Carraro, L.; Cardazzo, B.; Capolongo, F.; Merlanti, R.; Piccirillo, A. Beehive Products as Bioindicators of Antimicrobial Resistance Contamination in the Environment. Sci. Total Environ. 2022, 823, 151131. [Google Scholar] [CrossRef]
- Erban, T.; Ledvinka, O.; Kamler, M.; Hortova, B.; Nesvorna, M.; Tyl, J.; Titera, D.; Markovic, M.; Hubert, J. Bacterial Community Associated with Worker Honeybees (Apis mellifera) Affected by European Foulbrood. PeerJ 2017, 5, e3816. [Google Scholar] [CrossRef]
- Miller, D.L.; Smith, E.A.; Newton, I.L.G. A Bacterial Symbiont Protects Honey Bees from Fungal Disease. ASM J. 2021, 12, 3. [Google Scholar] [CrossRef]
- Dalenberg, H.; Maes, P.; Mott, B.; Anderson, K.E.; Spivak, M. Propolis Envelope Promotes Beneficial Bacteria in the Honey Bee (Apis mellifera) Mouthpart Microbiome. Insects 2020, 11, 453. [Google Scholar] [CrossRef]
- De Paula, J.C.; Doello, K.; Mesas, C.; Kapravelou, G.; Cornet-Gómez, A.; Orantes, F.J.; Martínez, R.; Linares, F.; Prados, J.C.; Porres, J.M.; et al. Exploring Honeybee Abdominal Anatomy through Micro-CT and Novel Multi-Staining Approaches. Insects 2022, 13, 556. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Remmas, N.; Voidarou, C.; Vrioni, G.; Konstantinidis, T.; Ntougias, S.; Tsakris, A. Microbial Community Structure among Honey Samples of Different Pollen Origin. Antibiotics 2023, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Wang, L.; Jin, Y.; Zhang, J.; Su, L.; Zhang, X.; Zhou, J.; Li, Y. The Microbial Community Dynamics during the Vitex Honey Ripening Process in the Honeycomb. Front. Microbiol. 2017, 8, 1649. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, P.B.; Adeleke, O.E.; OOla, I. Honey: A Reservoir for Microorganisms and an Inhibitory Agent for Microbes. Afr. Health Sci. 2007, 7, 159–165. [Google Scholar] [PubMed]
- Tsadila, C.; Nikolaidis, M.; Dimitriou, T.G.; Kafantaris, I.; Amoutzias, G.D.; Pournaras, S.; Mossialos, D. Antibacterial Activity and Characterization of Bacteria Isolated from Diverse Types of Greek Honey against Nosocomial and Foodborne Pathogens. Appl. Sci. 2021, 11, 5801. [Google Scholar] [CrossRef]
- Kňazovická, V.; Gábor, M.; Miluchová, M.; Bobko, M.; Medo, J. Diversity of bacteria in slovak and foreign honey, with assessment of its physico-chemical quality and counts of cultivable microorganisms. J. Microbiol. Biotechnol. Food Sci. 2019, 9, 414–421. [Google Scholar] [CrossRef]
- Lee, H.; Churey, J.J.; Worobo, R.W. Antimicrobial Activity of Bacterial Isolates from Different Floral Sources of Honey. Int. J. Food Microbiol. 2008, 126, 240–244. [Google Scholar] [CrossRef]
- Pomastowski, P.; Złoch, M.; Rodzik, A.; Ligor, M.; Kostrzewa, M.; Buszewski, B. Analysis of Bacteria Associated with Honeys of Different Geographical and Botanical Origin Using Two Different Identification Approaches: MALDI-TOF MS and 16S RDNA PCR Technique. PLoS ONE 2019, 14, e0217078. [Google Scholar] [CrossRef]
- Kwakman, P.H.S.; Zaat, S.A.J. Antibacterial Components of Honey. IUBMB Life 2012, 64, 48–55. [Google Scholar] [CrossRef]
- Majtan, J.; Bucekova, M.; Kafantaris, I.; Szweda, P.; Hammer, K.; Mossialos, D. Honey Antibacterial Activity: A Neglected Aspect of Honey Quality Assurance as Functional Food. Trends Food Sci. Technol. 2021, 118, 870–886. [Google Scholar] [CrossRef]
- Olivieri, C.; Marota, I.; Rollo, F.; Luciani, S. Tracking Plant, Fungal, and Bacterial DNA in Honey Specimens*. J. Forensic Sci. 2012, 57, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Schuh, C.M.A.P.; Aguayo, S.; Zavala, G.; Khoury, M. Exosome-like Vesicles in Apis mellifera Bee Pollen, Honey and Royal Jelly Contribute to Their Antibacterial and pro-Regenerative Activity. J. Exp. Biol. 2019, 222, jeb208702. [Google Scholar] [CrossRef]
- Lee, H.; Churey, J.J.; Worobo, R.W. Purification and Structural Characterization of Bacillomycin F Produced by a Bacterial Honey Isolate Active against Byssochlamys fulva H25. J. Appl. Microbiol. 2008, 105, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Ngalimat, M.S.; Raja Abd. Rahman, R.N.Z.; Yusof, M.T.; Syahir, A.; Sabri, S. Characterisation of Bacteria Isolated from the Stingless Bee, Heterotrigona itama, Honey, Bee Bread and Propolis. PeerJ 2019, 7, e7478. [Google Scholar] [CrossRef]
- Pajor, M.; Worobo, R.W.; Milewski, S.; Szweda, P. The Antimicrobial Potential of Bacteria Isolated from Honey Samples Produced in the Apiaries Located in Pomeranian Voivodeship in Northern Poland. Int. J. Environ. Res. Public. Health 2018, 15, 2002. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Quiñones, C.R.; Moreno-Terrazas, R.; Natividad-Bonifacio, I.; Quiñones-Ramírez, E.I.; Vázquez-Salinas, C. Microbiological Assessment of Honey in México. Rev. Argent. Microbiol. 2018, 50, 75–80. [Google Scholar] [CrossRef] [PubMed]
- D’alvise, P.; Böhme, F.; Codrea, M.C.; Seitz, A.; Nahnsen, S.; Binzer, M.; Rosenkranz, P.; Hasselmann, M. The Impact of Winter Feed Type on Intestinal Microbiota and Parasites in Honey Bees. Apidologie 2018, 49, 252–264. [Google Scholar] [CrossRef]
- Didaras, N.A.; Kafantaris, I.; Dimitriou, T.G.; Mitsagga, C.; Karatasou, K.; Giavasis, I.; Stagos, D.; Amoutzias, G.D.; Hatjina, F.; Mossialos, D. Biological Properties of Bee Bread Collected from Apiaries Located across Greece. Antibiotics 2021, 10, 555. [Google Scholar] [CrossRef]
- Giampieri, F.; Quiles, J.L.; Cianciosi, D.; Forbes-Hernández, T.Y.; Orantes-Bermejo, F.J.; Alvarez-Suarez, J.M.; Battino, M. Bee Products: An Emblematic Example of Underutilized Sources of Bioactive Compounds. J. Agric. Food Chem. 2022, 70, 6833–6848. [Google Scholar] [CrossRef]
- Oladipupo-Alade, E.O.; Lawal, O.A.; Oyewo, I.O.; Odiaka, I.E.; Haastrup, N.O.; Oyedele, M.D.; Ganiyu, O.A. Molecular Characterization of Gut Bacterial Flora of Honeybee (Apis mellifera Adansonii) from Some Selected Apiaries in Ogun State, Nigeria. J. Appl. Sci. Environ. Manag. 2021, 25, 605–608. [Google Scholar] [CrossRef]
- Georgi, I.; Asoutis Didaras, N.A.; Nikolaidis, M.; Dimitriou, T.G.; Charistos, L.; Hatjina, F.; Amoutzias, G.D.; Mossialos, D. The Impact of Vairimorpha (Nosema) Ceranae Natural Infection on Honey Bee (Apis mellifera) and Bee Bread Microbiota. Appl. Sci. 2022, 12, 11476. [Google Scholar] [CrossRef]
- El Ghouizi, A.; Bakour, M.; Laaroussi, H.; Ousaaid, D.; El Menyiy, N.; Hano, C.; Lyoussi, B. Bee Pollen as Functional Food: Insights into Its Composition and Therapeutic Properties. Antioxidants 2023, 12, 557. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, S.M.; Mahmud-Ab-Rashid, N.K.; Zawawi, N. Stingless Bee-Collected Pollen (Bee Bread): Chemical and Microbiology Properties and Health Benefits. Molecules 2021, 26, 957. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Carroll, M.J.; Sheehan, T.; Mott, B.M.; Maes, P.; Corby-Harris, V. Hive-Stored Pollen of Honey Bees: Many Lines of Evidence Are Consistent with Pollen Preservation, Not Nutrient Conversion. Mol. Ecol. 2014, 23, 5904–5917. [Google Scholar] [CrossRef]
- Santos, L.M.; Fonseca, M.S.; Sokolonski, A.R.; Deegan, K.R.; Araújo, R.P.C.; Umsza-Guez, M.A.; Barbosa, J.D.V.; Portela, R.D.; Machado, B.A.S. Propolis: Types, Composition, Biological Activities, and Veterinary Product Patent Prospecting. J. Sci. Food Agric. 2020, 100, 1369–1382. [Google Scholar] [CrossRef]
- Rufatto, L.C.; dos Santos, D.A.; Marinho, F.; Henriques, J.A.P.; Roesch Ely, M.; Moura, S. Red Propolis: Chemical Composition and Pharmacological Activity. Asian Pac. J. Trop. Biomed. 2017, 7, 591–598. [Google Scholar] [CrossRef]
- Kunugi, H.; Ali, A.M. Royal Jelly and Its Components Promote Healthy Aging and Longevity: From Animal Models to Humans. Int. J. Mol. Sci. 2019, 20, 4662. [Google Scholar] [CrossRef]
- Pasupuleti, V.R.; Sammugam, L.; Ramesh, N.; Gan, S.H. Honey, Propolis, and Royal Jelly: A Comprehensive Review of Their Biological Actions and Health Benefits. Oxid. Med. Cell Longev. 2017, 2017, 1259510. [Google Scholar] [CrossRef]
- Anderson, K.E.; Ricigliano, V.A.; Mott, B.M.; Copeland, D.C.; Floyd, A.S.; Maes, P. The Queen’s Gut Refines with Age: Longevity Phenotypes in a Social Insect Model. Microbiome 2018, 6, 108. [Google Scholar] [CrossRef]
- Moran, N.A. Genomics of the Honey Bee Microbiome. Curr. Opin. Insect Sci. 2015, 10, 22–28. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The Gut Microbiota of Insects—Diversity in Structure and Function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Jabal-Uriel, C.; Alba, C.; Higes, M.; Rodríguez, J.M.; Martín-Hernández, R. Effect of Nosema Ceranae Infection and Season on the Gut Bacteriome Composition of the European Honeybee (Apis mellifera). Sci. Rep. 2022, 12, 9326. [Google Scholar] [CrossRef] [PubMed]
- DeGrandi-Hoffman, G.; Corby-Harris, V.; DeJong, E.W.; Chambers, M.; Hidalgo, G. Honey Bee Gut Microbial Communities Are Robust to the Fungicide Pristine® Consumed in Pollen. Apidologie 2017, 48, 340–352. [Google Scholar] [CrossRef]
- Ellegaard, K.M.; Engel, P. Genomic Diversity Landscape of the Honey Bee Gut Microbiota. Nat. Commun. 2019, 10, 446. [Google Scholar] [CrossRef]
- Martinson, V.G.; Danforth, B.N.; Minckley, R.L.; Rueppell, O.; Tingek, S.; Moran, N.A. A Simple and Distinctive Microbiota Associated with Honey Bees and Bumble Bees. Mol. Ecol. 2011, 20, 619–628. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef]
- Elijah Powell, J.; Eiri, D.; Moran, N.A.; Rangel, J. Modulation of the Honey Bee Queen Microbiota: Effects of Early Social Contact. PLoS ONE 2018, 13, e0200527. [Google Scholar] [CrossRef]
- Yoshiyama, M.; Kimura, K. Bacteria in the Gut of Japanese Honeybee, Apis cerana Japonica, and Their Antagonistic Effect against Paenibacillus larvae, the Causal Agent of American Foulbrood. J. Invertebr. Pathol. 2009, 102, 91–96. [Google Scholar] [CrossRef]
- Gruneck, L.; Khongphinitbunjong, K.; Popluechai, S. Gut Microbiota Associated with Two Species of Domesticated Honey Bees from Thailand. Symbiosis 2021, 83, 335–345. [Google Scholar] [CrossRef]
- Praet, J.; Parmentier, A.; Schmid-Hempel, R.; Meeus, I.; Smagghe, G.; Vandamme, P. Large-Scale Cultivation of the Bumblebee Gut Microbiota Reveals an Underestimated Bacterial Species Diversity Capable of Pathogen Inhibition. Environ. Microbiol. 2018, 20, 214–227. [Google Scholar] [CrossRef]
- Schmidt, K.; Engel, P. Mechanisms Underlying Gut Microbiota-Host Interactions in Insects. J. Exp. Biol. 2021, 224, jeb207696. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.P.; Perutka, J.; Powell, J.E.; Geng, P.; Richhart, D.D.; Byrom, M.; Kar, S.; Davies, B.W.; Ellington, A.D.; Moran, N.A.; et al. Genetic Engineering of Bee Gut Microbiome Bacteria with a Toolkit for Modular Assembly of Broad-Host-Range Plasmids. ACS Synth. Biol. 2018, 7, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- de Paula, G.T.; Menezes, C.; Pupo, M.T.; Rosa, C.A. Stingless Bees and Microbial Interactions. Curr. Opin. Insect Sci. 2021, 44, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee Gut Microbiota Promotes Host Weight Gain via Bacterial Metabolism and Hormonal Signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, T.C.; Butler, È.; Markowicz, P.; Lindholm, C.; Larsson, L.; Vásquez, A. Lactic Acid Bacterial Symbionts in Honeybees—An Unknown Key to Honey’s Antimicrobial and Therapeutic Activities. Int. Wound J. 2016, 13, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, 33188. [Google Scholar] [CrossRef]
- Dosch, C.; Manigk, A.; Streicher, T.; Tehel, A.; Paxton, R.J.; Tragust, S. The Gut Microbiota Can Provide Viral Tolerance in the Honey Bee. Microorganisms 2021, 9, 871. [Google Scholar] [CrossRef]
- Rothman, J.A.; Leger, L.; Kirkwood, J.S.; Mcfrederick, Q.S.; Stabb, E.V. Cadmium and Selenate Exposure Affects the Honey Bee Microbiome and Metabolome, and Bee-Associated Bacteria Show Potential for Bioaccumulation. Appl. Environ. Microbiol. 2019, 85, e01411-19. [Google Scholar] [CrossRef]
- Sato, A.; Okamura, Y.; Murakami, M. Diversification and Selection Pattern of CYP6B Genes in Japanese Papilio Butterflies and Their Association with Host Plant Spectra. PeerJ 2020, 8, e10625. [Google Scholar] [CrossRef]
- Sinacori, M.; Francesca, N.; Alfonzo, A.; Cruciata, M.; Sannino, C.; Settanni, L.; Moschetti, G. Cultivable Microorganisms Associated with Honeys of Different Geographical and Botanical Origin. Food Microbiol. 2014, 38, 284–294. [Google Scholar] [CrossRef]
- Scepankova, H.; Pinto, C.A.; Paula, V.; Estevinho, L.M.; Saraiva, J.A. Conventional and Emergent Technologies for Honey Processing: A Perspective on Microbiological Safety, Bioactivity, and Quality. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5393–5420. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.T.; Peck, M.W. Genomes, Neurotoxins and Biology of Clostridium Botulinum Group I and Group II. Res. Microbiol. 2015, 166, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, K.; Flick, R. Accumulation of Soluble Menaquinones MK-7 in Honey Coincides with Death of Bacillus Spp. Present in Honey. Food Chem. X 2019, 1, 100008. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Argueso, T.; Rodriguez-Navarro, A. Microbiology of Ripening Honey. Appl. Microbiol. 1975, 30, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Snowdon, J.A.; Cliver, D.O. Microorganisms in Honey. Int. J. Food Microbiol. 1996, 31, 1–26. [Google Scholar] [CrossRef]
- Kacániová, M.; Pavličová, S.; Haščík, P.; Kociubinski, G.; Kńazovická, V.; Sudzina, M.; Sudzinová, J.; Fikselová, M. Microbial Communities in Bees, Pollen and Honey from Slovakia. Acta Microbiol. Immunol. Hung. 2009, 56, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, T.C.; Vásquez, A. Detection and Identification of a Novel Lactic Acid Bacterial Flora within the Honey Stomach of the Honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef]
- Grabowski, N.T.; Klein, G. Microbiology and Food-Borne Pathogens in Honey. Crit. Rev. Food Sci. Nutr. 2015, 57, 1852–1862. [Google Scholar] [CrossRef]
- Jacinto-Castillo, D.F.; Canto, A.; Medina-Medina, L.A.; O’Connor-Sánchez, A. Living in Honey: Bacterial and Fungal Communities in Honey of Sympatric Populations of Apis mellifera and the Stingless Bee Melipona Beecheii, in Yucatan, Mexico. Arch. Microbiol. 2022, 204, 718. [Google Scholar] [CrossRef]
- Ambika Manirajan, B.; Ratering, S.; Rusch, V.; Schwiertz, A.; Geissler-Plaum, R.; Cardinale, M.; Schnell, S. Bacterial Microbiota Associated with Flower Pollen Is Influenced by Pollination Type, and Shows a High Degree of Diversity and Species-Specificity. Environ. Microbiol. 2016, 18, 5161–5174. [Google Scholar] [CrossRef]
- Manirajan, B.A.; Maisinger, C.; Ratering, S.; Rusch, V.; Schwiertz, A.; Cardinale, M.; Schnell, S. Diversity, Specificity, Co-Occurrence and Hub Taxa of the Bacterial–Fungal Pollen Microbiome. FEMS Microbiol. Ecol. 2018, 94, 112. [Google Scholar] [CrossRef] [PubMed]
- McFrederick, Q.S.; Thomas, J.M.; Neff, J.L.; Vuong, H.Q.; Russell, K.A.; Hale, A.R.; Mueller, U.G. Flowers and Wild Megachilid Bees Share Microbes. Microb. Ecol. 2017, 73, 188–200. [Google Scholar] [CrossRef] [PubMed]
- McFrederick, Q.S.; Rehan, S.M. Wild Bee Pollen Usage and Microbial Communities Co-Vary Across Landscapes. Microb. Ecol. 2019, 77, 513–522. [Google Scholar] [CrossRef]
- Mattila, H.R.; Rios, D.; Walker-Sperling, V.E.; Roeselers, G.; Newton, I.L.G. Characterization of the Active Microbiotas Associated with Honey Bees Reveals Healthier and Broader Communities When Colonies Are Genetically Diverse. PLoS ONE 2012, 7, e32962. [Google Scholar] [CrossRef] [PubMed]
- Janashia, I.; Carminati, D.; Rossetti, L.; Zago, M.; Fornasari, M.E.; Haertlé, T.; Chanishvili, N.; Giraffa, G. Characterization of Fructophilic Lactic Microbiota of Apis mellifera from the Caucasus Mountains. Ann. Microbiol. 2016, 66, 1387–1395. [Google Scholar] [CrossRef]
- Grubbs, K.J.; Scott, J.J.; Budsberg, K.J.; Read, H.; Balser, T.C.; Currie, C.R. Unique Honey Bee (Apis mellifera) Hive Component-Based Communities as Detected by a Hybrid of Phospholipid Fatty-Acid and Fatty-Acid Methyl Ester Analyses. PLoS ONE 2015, 10, e0121697. [Google Scholar] [CrossRef]
- Ersoy Omeroglu, E.; Keriman Arserim-Uçar, D.; Yegin, Z.; Çağlayan, N.; Nur Zafer Yurt, M.; Busra Tasbasi, B.; Esma Acar, E.; Ucak, S.; Cengiz Ozalp, V.; Sudagidan, M. Determination of Bacterial Diversity of Propolis Microbiota. Chem. Biodivers. 2023, 20, e202201182. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Kawas, J.R.; Marroquin-Cardona, A.G. Descriptive Bacterial and Fungal Characterization of Propolis Using Ultra-High-Throughput Marker Gene Sequencing. Insects 2019, 10, 402. [Google Scholar] [CrossRef]
- Corby-Harris, V.; Snyder, L.A.; Schwan, M.R.; Maes, P.; McFrederick, Q.S.; Anderson, K.E. Origin and Effect of Alpha 2.2 Acetobacteraceae in Honey Bee Larvae and Description of Parasaccharibacter apium Gen. Nov., Sp. Nov. Appl. Environ. Microbiol. 2014, 80, 7460. [Google Scholar] [CrossRef]
- Asama, T.; Arima, T.H.; Gomi, T.; Keishi, T.; Tani, H.; Kimura, Y.; Tatefuji, T.; Hashimoto, K. Lactobacillus kunkeei YB38 from Honeybee Products Enhances IgA Production in Healthy Adults. J. Appl. Microbiol. 2015, 119, 818–826. [Google Scholar] [CrossRef]
- Evans, J.D.; Armstrong, T.N. Antagonistic Interactions between Honey Bee Bacterial Symbionts and Implications for Disease. BMC Ecol. 2006, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Gomila, M.; Mulet, M.; Zaruma, A.; García-Valdés, E. Past, Present and Future of the Boundaries of the Pseudomonas Genus: Proposal of Stutzerimonas Gen. Nov. Syst. Appl. Microbiol. 2022, 45, 126289. [Google Scholar] [CrossRef] [PubMed]
- Spiers, A.J.; Buckling, A.; Rainey, P.B. The Causes of Pseudomonas Diversity. Microbiology (Reading) 2000, 146 Pt 10, 2345–2350. [Google Scholar] [CrossRef] [PubMed]
- Molina, G.; Pimentel, M.R.; Pastore, G.M. Pseudomonas: A Promising Biocatalyst for the Bioconversion of Terpenes. Appl. Microbiol. Biotechnol. 2013, 97, 1851–1864. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas Genomes: Diverse and Adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef]
- Fernandez De Landa, G.; Alberoni, D.; Baffoni, L.; Fernandez De Landa, M.; Revainera, P.D.; Porrini, L.P.; Brasesco, C.; Quintana, S.; Zumpano, F.; Eguaras, M.J.; et al. The Gut Microbiome of Solitary Bees Is Mainly Affected by Pathogen Assemblage and Partially by Land Use. Environ. Microbiome 2023, 18, 38. [Google Scholar] [CrossRef]
- Anjum, S.I.; Aldakheel, F.; Shah, A.H.; Khan, S.; Ullah, A.; Hussain, R.; Khan, H.; Ansari, M.J.; Mahmoud, A.H.; Mohammed, O.B. Honey Bee Gut an Unexpected Niche of Human Pathogen. J. King Saud. Univ. Sci. 2021, 33, 101247. [Google Scholar] [CrossRef]
- Yu, L.; Yang, H.; Cheng, F.; Wu, Z.; Huang, Q.; He, X.; Yan, W.; Zhang, L.; Wu, X. Honey Bee Apis mellifera Larvae Gut Microbial and Immune, Detoxication Responses towards Flumethrin Stress. Environ. Pollut. 2021, 290, 118107. [Google Scholar] [CrossRef]
- Bosmans, L.; Pozo, M.I.; Verreth, C.; Crauwels, S.; Wilberts, L.; Sobhy, I.S.; Wäckers, F.; Jacquemyn, H.; Lievens, B. Habitat-Specific Variation in Gut Microbial Communities and Pathogen Prevalence in Bumblebee Queens (Bombus terrestris). PLoS ONE 2018, 13, e0204612. [Google Scholar] [CrossRef]
- Tola, Y.H.; Waweru, J.W.; Hurst, G.D.D.; Slippers, B.; Paredes, J.C. Characterization of the Kenyan Honey Bee (Apis mellifera) Gut Microbiota: A First Look at Tropical and Sub-Saharan African Bee Associated Microbiomes. Microorganisms 2020, 8, 1721. [Google Scholar] [CrossRef]
- Ahn, J.H.; Hong, I.P.; Bok, J.I.; Kim, B.Y.; Song, J.; Weon, H.Y. Pyrosequencing Analysis of the Bacterial Communities in the Guts of Honey Bees Apis cerana and Apis mellifera in Korea. J. Microbiol. 2012, 50, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Yang, Y.; Fu, Z.; Diao, Q.; Wang, M.; Luo, Q.; Wang, X.; Dai, P. A Combination of Tropilaelaps Mercedesae and Imidacloprid Negatively Affects Survival, Pollen Consumption and Midgut Bacterial Composition of Honey Bee. Chemosphere 2021, 268, 129368. [Google Scholar] [CrossRef] [PubMed]
- Gaggìa, F.; Jakobsen, R.R.; Alberoni, D.; Baffoni, L.; Cutajar, S.; Mifsud, D.; Nielsen, D.S.; Di Gioia, D. Environment or Genetic Isolation? An Atypical Intestinal Microbiota in the Maltese Honey Bee Apis mellifera Spp. Ruttneri. Front. Microbiol. 2023, 14, 1127717. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Žiarovská, J.; Kowalczewski, P.Ł. In Vitro Antagonistic Effect of Gut Bacteriota Isolated from Indigenous Honey Bees and Essential Oils against Paenibacillus larvae. Int. J. Mol. Sci. 2020, 21, 6736. [Google Scholar] [CrossRef] [PubMed]
- Topal, E.; Ceylan; Tunca, R.; Bay, V.; Aldemir, S.; İnci, H.; Topçuoğlu, U.; Kösoğlu, M. The Effect of Different Feeding Strategies on Honey Bee Gut Microbiota and the Presence of Nosema. S. Afr. J. Anim. Sci. 2022, 52, 577–590. [Google Scholar] [CrossRef]
- Oladipupo-Alade, E.O.; Lawal, O.A.; Owolola, O.I.; Emmanuel, I.B.; Oyedele, M.D.; Odiaka, I.E.; Haastrup, N.O. Phylogenetic Relationship Between The Gut Bacterial Flora of Honeybee (Apis mellifera) from Apiary in Ogun State, Nigeria. J. Res. For. Wildl. Environ. 2021, 13, 123–134. [Google Scholar]
- Santos, A.C.C.; Borges, L.D.F.; Rocha, N.D.C.; de Carvalho Azevedo, V.A.; Bonetti, A.M.; dos Santos, A.R.; da Rocha Fernandes, G.; Dantas, R.C.C.; Ueira-Vieira, C. Bacteria, Yeasts, and Fungi Associated with Larval Food of Brazilian Native Stingless Bees. Sci. Rep. 2023, 13, 5147. [Google Scholar] [CrossRef]
- Praet, J. The Gut Microbiota of Bumblebees: A Treasure Chest of Biodiversity and Functionality. Ph.D. Dissertation, Ghent University, Ghent, Belgium, 2017. [Google Scholar]
- Zhao, H.; Tao, K.; Zhu, J.; Liu, S.; Gao, H.; Zhou, X. Bioremediation Potential of Glyphosate-Degrading Pseudomonas Spp. Strains Isolated from Contaminated Soil. J. Gen. Appl. Microbiol. 2015, 61, 165–170. [Google Scholar] [CrossRef]
- Wang, G.H.; Berdy, B.M.; Velasquez, O.; Jovanovic, N.; Alkhalifa, S.; Minbiole, K.P.C.; Brucker, R.M. Changes in Microbiome Confer Multigenerational Host Resistance after Sub-Toxic Pesticide Exposure. Cell Host Microbe 2020, 27, 213–224.e7. [Google Scholar] [CrossRef]
- De Souza, M.L.; Wackett, L.P.; Sadowsky, M.J. The atzABC genes encoding atrazine catabolism are located on a self-transmissible plasmid in Pseudomonas sp. strain ADP. Appl. Environ. Microbiol. 1998, 64, 2323–2326. [Google Scholar] [CrossRef]
- Indiragandhi, P.; Anandham, R.; Madhaiyan, M.; Poonguzhali, S.; Kim, G.H.; Saravanan, V.S.; Sa, T. Cultivable Bacteria Associated with Larval Gut of Prothiofos-Resistant, Prothiofos-Susceptible and Field-Caught Populations of Diamondback Moth, Plutella Xylostella and Their Potential for, Antagonism towards Entomopathogenic Fungi and Host Insect Nutrition. J. Appl. Microbiol. 2007, 103, 2664–2675. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Perraud, Q. Pseudomonas Aeruginosa and Its Multiple Strategies to Access Iron. Environ. Microbiol. 2023, 25, 811–831. [Google Scholar] [CrossRef] [PubMed]
- Mossialos, D.; Amoutzias, G.D. Role of Siderophores in Cystic Fibrosis Pathogenesis: Foes or Friends? Int. J. Med. Microbiol. 2009, 299, 87–98. [Google Scholar] [CrossRef]
- Mossialos, D.; Amoutzias, G.D. Siderophores in Fluorescent Pseudomonads: New Tricks from an Old Dog. Future Microbiol. 2007, 2, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Barta, D.G.; Cornea-Cipcigan, M.; Margaoan, R.; Vodnar, D.C. Biotechnological Processes Simulating the Natural Fermentation Process of Bee Bread and Therapeutic Properties—An Overview. Front. Nutr. 2022, 9, 871896. [Google Scholar] [CrossRef] [PubMed]
- Didaras, N.A.; Karatasou, K.; Dimitriou, T.G.; Amoutzias, G.D.; Mossialos, D. Antimicrobial Activity of Bee-Collected Pollen and Beebread: State of the Art and Future Perspectives. Antibiotics 2020, 9, 811. [Google Scholar] [CrossRef] [PubMed]



| Detection and Relative Abundance of Pseudomonads | Reported Pseudomonas Species | Authors | ||
|---|---|---|---|---|
| Honey bees | Apis mellifera (larvae) | 7.73% | - | Yu et al., 2021 [130] |
| Apis mellifera Summer samples | (0.002–18.0%) Mean:0.01% | - | D’Alvise et al., 2017 [69] | |
| Apis mellifera winter samples | 0–0.19% Mean: 0.002% | - | ||
| Apis dorsata (giant honey bee) | Detected | - | Gruneck et al., 2021 [91] | |
| Apis mellifera scutellata | 2.3% | - | Yosef Hamba Tola et al., 2020 [132] | |
| Apis mellifera (adult) | 0.6% | - | Jae-Hyung Ahn et al., 2012 [133] | |
| Apis mellifera | 2.27% | - | Shilong Ma et al., 2021 [134] | |
| Apis mellifera ruttneri | 1–3% | - | Gaggìa et al., 2023 [135] | |
| Apis mellifera | - | Pseudomonas marginalis | Kačániová et al., 2020 [136] | |
| - | Pseudomonas oryzihabitans | |||
| - | Pseudomonas putida | |||
| Apis mellifera anatoliaca | - | Pseudomonas luteola | Topal et al., 2022 [137] | |
| - | Pseudomonas alcaligenes | |||
| Apismellifera adansonii | - | Pseudomonas aeruginosa | Oladipupo-Alade et al., 2021 [138] | |
| - | Pseudomonas plecoglossicida | |||
| Stingless bee | Frieseomelitta varia | Detected | Pseudomonas syringae | De Sousa 2021 [19] |
| Melipona quadrifasciata | Pseudomonas putida | |||
| Lepidotrigonaterminata | 3.35% | - | Qi-Hi Tang et al., 2021 [44] | |
| Lepidotrigona ventralis | 3.18% | - | ||
| Tetragnula pagdeni | 4.63% | - | ||
| Melipona quadrifasciata | Detected | Santos et al., 2023 [139] | ||
| Bumble bees | Bombus lapidarius | 5.98% | - | Jessy Praet 2017 [140] |
| Bombuslucorum | 0.48% | - | ||
| Bombusterrestris | 0.24% | - | ||
| Honey bee products | Bee pollen | Detected | - | Disayathanoowat et al., 2020 [26] Manirajan et al., 2018 [113] Laconi et al., 2022 [49] Harris et al., 2014 [7] |
| Bee bread | Detected | - | Donkersley et al., 2018 [6] Anderson et al., 2014 [76] | |
| Honey | - | Pseudomonas sp. Pseudomonas coleopterorum Pseudomonas fulva Pseudomonas stutzeri | Tsadila et al., 2021 [57] | |
| Detected | - | Stavropoulou et al., 2023 [54] | ||
| - | Pseudomonas sp. | Olivieri et al., 2012 [63] | ||
| - | Pseudomonas migulae | |||
| - | Pseudomonas flectens | |||
| Propolis | Detected | - | Casalone et al., 2020 [24] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsadila, C.; Amoroso, C.; Mossialos, D. Microbial Diversity in Bee Species and Bee Products: Pseudomonads Contribution to Bee Well-Being and the Biological Activity Exerted by Honey Bee Products: A Narrative Review. Diversity 2023, 15, 1088. https://doi.org/10.3390/d15101088
Tsadila C, Amoroso C, Mossialos D. Microbial Diversity in Bee Species and Bee Products: Pseudomonads Contribution to Bee Well-Being and the Biological Activity Exerted by Honey Bee Products: A Narrative Review. Diversity. 2023; 15(10):1088. https://doi.org/10.3390/d15101088
Chicago/Turabian StyleTsadila, Christina, Chiara Amoroso, and Dimitris Mossialos. 2023. "Microbial Diversity in Bee Species and Bee Products: Pseudomonads Contribution to Bee Well-Being and the Biological Activity Exerted by Honey Bee Products: A Narrative Review" Diversity 15, no. 10: 1088. https://doi.org/10.3390/d15101088