
Two New and One First Recorded Species of Xylaria Isolated from Fallen Leaves in Hainan Tropical Rainforest National Park in China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Source
2.2. Morphological Investigation
2.3. Molecular Research
2.4. Phylogenetic Analyses
3. Results
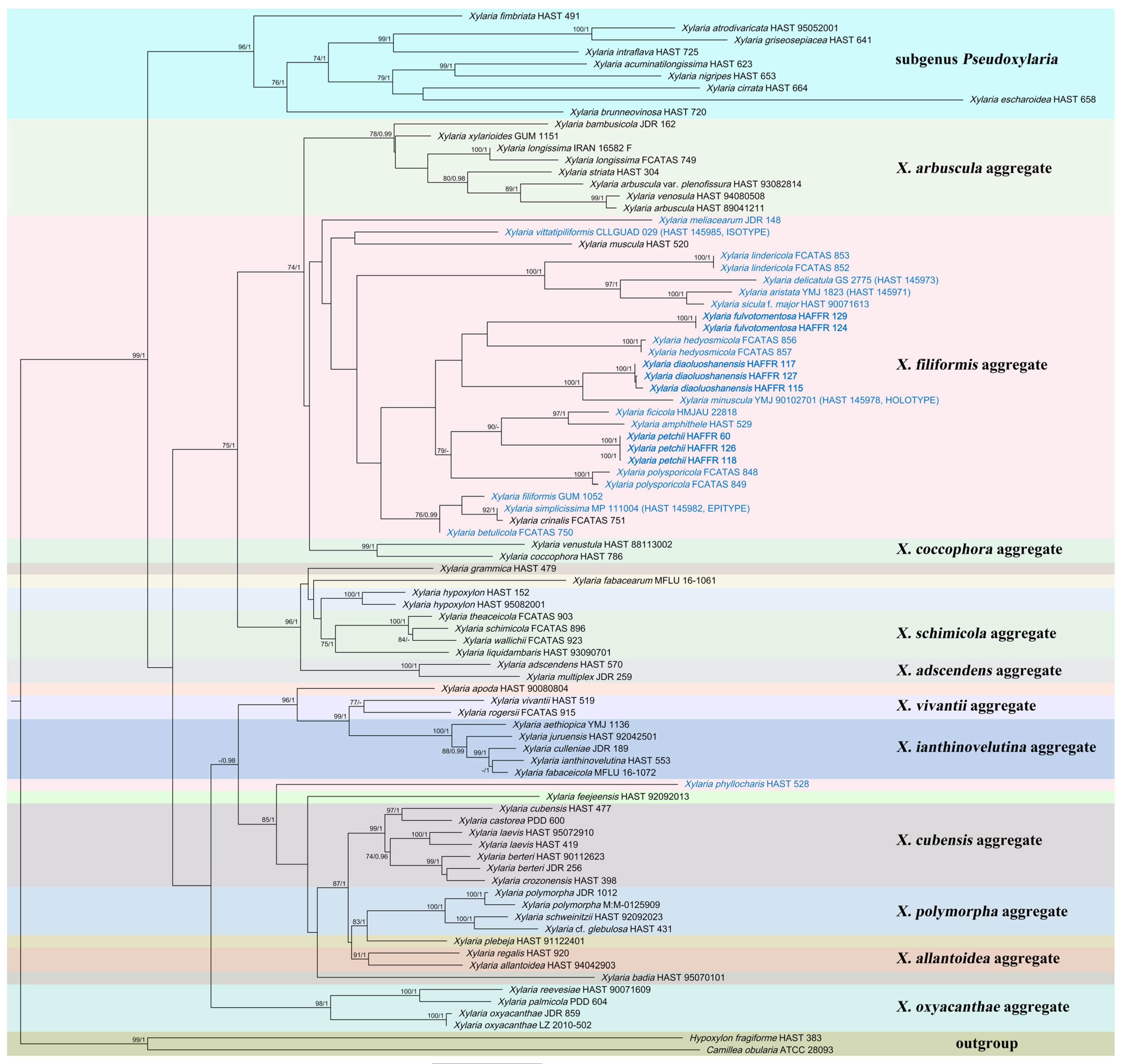
3.1. Molecular Phylogeny
3.2. Taxonomy
4. Discussion
- 1 Stromata branched, long stipes that bear one to three clavae on each terminal branch
- .....................................................................................................................................X. luxurians
- 1 Stromata unbranched to occasionally branched.....................................................................2
- 2 Stipes tomentose or glabrous to tomentose.............................................................................3
- 2 Stipes glabrous...........................................................................................................................15
- 3 Fertile parts filiform.....................................................................................................X. duranii
- 3 Fertile parts not filiform.............................................................................................................4
- 4 Fertile parts cylindrical...............................................................................................................5
- 4 Fertile parts capitate..................................................................................................................11
- 5 Fertile parts overlain by dark long spikes...............................................................X. asperata
- 5 Fertile parts without dark long spikes.....................................................................................6
- 6 Fertile parts glabrous.........................................................................................X. appendiculata
- 6 Fertile parts overlain by tomentum..........................................................................................7
- 7 Fertile parts densely covered by yellow tomentum...................................X. fulvotomentosa
- 7 Fertile parts with non-yellow tomentum.................................................................................8
- 8 Ascospores with non-cellular appendages..............................................................................9
- 8 Ascospores without non-cellular appendages......................................................................10
- 9 Ascospores (14.5–)15–16.5(–17) × (8–)8.5–9.5(–10) µm..............................................X. allima
- 9 Ascospores (10–)10.5–12(–14) × (5–)6–7(–7.5) µm.........................................................X. lima
- 10 Surface of fertile parts with half-exposed to fully exposed perithecial contours...............
- ...........................................................................................................................................X. castilloi
- 10 Surface of fertile parts lacking perithecial mounds or with slight perithecial mounds
- .......................................................................................................................................X. maitlandii
- 11 Stromata with an acute apex.................................................................................................12
- 11 Stromata with a rounded apex..............................................................................................14
- 12 Ascospores with non-cellular appendages..............................................................X. axifera
- 12 Ascospores without non-cellular appendages....................................................................13
- 13 Consistency fragile, ascospores (10–)10.5–12.5(–14) × (5.5–)6–7(–7.5) µm............................
- .......................................................................................................................X. aristata var. aristata
- 13 Consistency soft, ascospores (15–)15.5–17(–18) × (6.5–)7.5–9(–9.5) µm..........X. hispidipes
- 14 Ascospores (13.5–)14–16(–17) × (5.5–)6–7(–7.5) µm...........................X. aristata var. hirsuta
- 14 Ascospores 8–9(–9.5) × 4–4.5(–6.6) µm................................................................X. imminuta
- 15 Fertile parts cylindrical or conical to subglobose, most perithecia gather near the top of
- the stromata, with several occasionally scattered below.............................................X. petchii
- 15 Fertile parts with uniform morphology, perithecia lacking the above cluster patterns
- .......................................................................................................................................................16
- 16 Fertile parts cylindrical...........................................................................................................17
- 16 Fertile parts not cylindrical....................................................................................................30
- 17 Ascospores with non-cellular appendages..........................................................................18
- 17 Ascospores without non-cellular appendages....................................................................25
- 18 Stromata with an outer peeling layer split into band-like stripes, perithecia 150–200 µm
- .......................................................................................................................................X. vittiformis
- 18 Stromata without an outer peeling layer or the outer peeling layer without band-like
- stripes, perithecia greater than 250 µm.....................................................................................19
- 19 Surface of fertile parts lacking perithecial mounds, ascospores (22–)23.5–27(–28) × (8.5–)
- 9–10.5(–11) µm................................................................................................X. spiculaticlavata
- 19 Surface of fertile parts with conspicuous perithecial mounds, ascospores length less
- than 19 µm and width nearly less than 9 µm.......................................................................20
- 20 Stromata with an outer peeling layer split into narrow or thread-like stripes, ascospores
- (13.5–)14–15(–17) × (4.5–)5–6(–7) µm........................................................................X. minuscula
- 20 Stromata without an outer peeling layer or the outer peeling layer without narrow or
- thread-like stripes.........................................................................................................................21
- 21 Stromata with a blunt apex, ascospores 8–10 × 4–6 μm........................................X. kamatii
- 21 Stromata with an acute apex, ascospores length greater than 10.3 μm..........................22
- 22 Ostioles conic-papillate, tilting upwards, 120–150 μm broad at base............................
- ...................................................................................................................X. appendiculatoides
- 22 Ostioles papillate or slightly papillate, less than 80 μm broad at base...........................23
- 23 Stromata surface blackish brown, ascospores (15–)16.5–18(–19) × (7.5–)8–9(–9.5) µm
- .............................................................................................................................X. phyllophila
- 23 Stromata surface black, ascospores length less than 16.5 µm and width nearly less than
- 7.5 µm...........................................................................................................................................24
- 24 Stromata filiform, with long sterile filiform apexes up to 10–35 mm, ascospores
- ellipsoid to fusiform, (10.3–)11.5–14(–16.5) × (4.1–)4.6–5.7(–6.8) μm........X. diaoluoshanensis
- 24 Stromata cylindrical, with a mucronate apex 2 mm, ascospores ellipsoid, (11.5–)12.5–
- 14.5(–15) × 5.5–8 μm............................................................................................X. polysporicola
- 25 Stromata surface dark vinaceous brown, ascospores strongly inequilateral...............
- .................................................................................................................................X. phyllocharis
- 25 Stromata surface not dark vinaceous brown, ascospores inequilateral..........................26
- 26 Ascospores light brown to brown, (5.5–)6–7 × 3–3.5(–4) µm............................X. diminuta
- 26 Ascospores brown to blackish brown, length greater than 8.5 µm and width larger than
- 4 µm...........................................................................................................................................27
- 27 Stromata without an outer layer.........................................................................X. neblinensis
- 27 Stromata with an outer layer.................................................................................................28
- 28 Stromata with an acuminate or mucronate apex, ascospores (9–)9.5–10.5(–11) × (5.5–)6–
- 6.5(–7) µm............................................................................................................X. noduliformis
- 28 Stromata with a long acicular apex......................................................................................29
- 29 Stromata 23–35 mm total length, overlain by an outer peeling layer split into narrow or
- thread-like stripes, ascospores (8.5–)9–11 × 4–6 μm.................................................X. foliicola
- 29 Stromata 61–78 mm total length, overlain by an outer peeling layer split into band-like
- stripes, ascospores (10–)11–12(–12.5) × (5.5–)6–7(–7.5) µm........X. vittatipiliformis
- 30 Fertile parts filiform................................................................................................................31
- 30 Fertile parts not filiform.........................................................................................................36
- 31 Ascospores with spiral germ slit.....................................................................X. meliacearum
- 31 Ascospores with straight germ slit.......................................................................................32
- 32 Consistency fragile, ascospores (15–)16.5–19(–21.5) × (5–)5.5–6.5(–7.5) µm.....................
- ..................................................................................................................................X. simplicissima
- 32 Consistency soft, ascospores length less than 14.5 µm.......................................................33
- 33 Ascospores with non-cellular appendages..........................................................................34
- 33 Ascospores without non-cellular appendages....................................................................35
- 34 Ascospores light brown, short fusoid....................................................................X. fliformis
- 34 Ascospores brown to dark brown, ellipsoid...........................................................X. vagans
- 35 Stromata 15–20 mm total length, ascospores ellipsoid, 12–13.5 × 4–5 μm.......X. eugeniae
- 35 Stromata 35–83 mm total length, ascospores ellipsoid to shortly fusoid, (9–)9.5–10.5(–
- 11) × (3.5–)4–4.5(–5) µm..................................................................X. vermiformis
- 36 Ascospores with non-cellular appendages..........................................................................37
- 36 Ascospores without non-cellular appendages....................................................................43
- 37 Surface of fertile parts with conspicuous perithecial mounds.........................................38
- 37 Surface of fertile parts lacking perithecial mounds...........................................................42
- 38 Ascospores with cellular appendage on one end...............................................................39
- 38 Ascospores without cellular appendage..............................................................................40
- 39 Stromata 50 mm total length, without a long apex, ostioles slightly papillate, ascospores
- (12–)12.5–15.5(–17) × (5–)6–7.5(–8) µm...............................................................X. amphithele
- 39 Stromata 91–147 mm total length, with a long apex, ostioles coarsely papillate,
- ascospores (10.5–)11.5–13.5(–15) × (5–)5.5–6.5(–7.5) µm...................................X. nainitalensis
- 40 Stromata without an acute apex, ascospores (14.5–)15.5–18(–19) × (5–)5.5–6.5(–7) μm
- ...........................................................................................................................................X. heloidea
- 40 Stromata with an acute apex.................................................................................................41
- 41 Stromata with a mucronate apex, ascospores (10–)10.5–12(–12.5) × (5–)5.5–6(–6.5) µm
- .....................................................................................................................................X. pisoniae
- 41 Stromata with a long acicular apex, much longer than the fertile part, ascospores (8.5–)
- 9.5–11(–12) × (4–)4.5–6(–6.5) µm........................................................................................X. sicula
- 42 Stromata surface dull grayish brown, overlain with a thin pellicle cracked reticulately
- into plaques 100–200 µm broad, fertile parts capitate...........................................X. hypsipoda
- 42 Stromata surface dark brown to blackish, without the above plaques, fertile parts peltate
- .............................................................................................................................................X. memecyli
- 43 Ascospores dark brown, nearly semicircular to broadly ellipsoid, (10–)10.5–12.5(–14) ×
- (5.5–)6–7(–7.5) µm........................................................................................................X. delicatula
- 43 Ascospores dark brown to blackish brown, ellipsoid, (12.5–)13–15(–16) × (7.5–)8–9(–10) µm
- ..................................................................................................................................................X. clusiae
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Husbands, D.R.; Urbina, H.; Lewis, S.M.; Aime, M.C. Xylaria karyophthora: A new seed-inhabiting fungus of Greenheart from Guyana. Mycologia 2018, 110, 434–447. [Google Scholar] [CrossRef]
- Wangsawat, N.; Ju, Y.M.; Phosri, C.; Whalley, A.J.S.; Suwannasai, N. Twelve new taxa of Xylaria associated with termite nests and soil from Northeast Thailand. Biology 2021, 10, 575. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 771. [Google Scholar]
- Ma, H.X.; Song, Z.K.; Pan, X.Y.; Li, Y.; Yang, Z.N.; Qu, Z. Multi-gene phylogeny and taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China. Diversity 2022, 14, 37. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Lin, C.R.; Fang, M.J.; Rogers, J.D.; Fournier, J.; Lechat, C.; Ju, Y.M. Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioideae (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol. Phylogenet. Evol. 2010, 54, 957–969. [Google Scholar] [CrossRef]
- Kim, C.S.; Jo, G.W.; Kwag, Y.N.; Oh, S.O.; Lee, S.G.; Sung, G.H.; Oh, G.; Shrestha, B.; Kim, S.Y.; Shin, C.H.; et al. New records of Xylaria species in Korea: X. ripicola sp. nov. and X. tentaculata. Mycobiology 2016, 44, 21–28. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D.; Hsieh, H.M. Xylaria species associated with fallen fruits and seeds. Mycologia 2018, 110, 726–749. [Google Scholar] [CrossRef]
- Rogers, J.D.; Ju, Y.M.; Lehmann, J. Some Xylaria species on termite nests. Mycologia 2005, 97, 914–923. [Google Scholar] [CrossRef]
- Ju, Y.M.; Hsieh, H.M. Xylaria species associated with nests of Odontotermes formosanus in Taiwan. Mycologia 2007, 99, 936–957. [Google Scholar] [CrossRef]
- Fournier, J.; Lechat, C.; Courtecuisse, R. The genus Xylaria sensu lato (Xylariaceae) in Guadeloupe and Martinique (French West Indies) III. Taxa with slender upright stromata. Ascomycete.org 2020, 12, 81–164. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Ju, Y.M.; Lechat, C.; Fournier, J.; Huart, D. New ecological, morphological, cultural and molecular phylogenetic insights into Xylaria guepini (Xylariaceae). Ascomycete.org 2022, 14, 177–184. [Google Scholar]
- Ju, Y.M.; Hsieh, H.M.; He, X.S. Wulingshen, the massive Xylaria sclerotia used as traditional Chinese medicine, is produced by multiple species. Mycologia 2022, 114, 175–189. [Google Scholar] [CrossRef]
- Pan, X.Y.; Song, Z.K.; Qu, Z.; Liu, T.D.; Ma, H.X. Three new Xylaria species (Xylariaceae, Xylariales) on fallen leaves from Hainan Tropical Rainforest National Park. MycoKeys 2022, 86, 47–63. [Google Scholar] [CrossRef]
- Ju, Y.M.; Hsieh, H.M. Xylaria species associated with fallen leaves and petioles. Bot. Stud. 2023, 64, 19. [Google Scholar] [CrossRef]
- Ma, H.X.; Li, Y. Xylaria crinalis and X. betulicola from China—Two new species with thread-like stromata. Sydowia 2018, 70, 37–49. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Fournier, J.; Ju, Y.M.; Hsieh, H.M.; Lindermann, U. Xylaria aethiopica sp. nov.—A new pod-inhabiting species of Xylaria (Xylariaceae) from Ethiopia. Ascomycete.org 2018, 10, 209–215. [Google Scholar]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Hashemi, S.A.; Zare, R.; Khodaparast, S.A.; Elahinia, S.A. A new Xylaria species from Iran. Mycol. Iran. 2015, 2, 1–10. [Google Scholar]
- Roensch, P.; Roensch, S.; Reiher, A.; Otto, P. Investigations on the fructicolous Xylaria delitschii and Xylaria Oxyacanthae. Boletus 2010, 32, 106–122. [Google Scholar]
- Persoh, D.; Melcher, M.; Graf, K.; Fournier, J.; Stadler, M.; Rambold, G. Molecular and morphological evidence for the delimitation of Xylaria hypoxylon. Mycologia 2009, 101, 256–268. [Google Scholar] [CrossRef]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsaard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from amultigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef]
- Song, Z.K.; Pan, X.Y.; Li, C.T.; Ma, H.X.; Li, Y. Two new species of Hypoxylon (Hypoxylaceae) from China based on morphological and DNA sequence data analyses. Phytotaxa 2022, 538, 213–224. [Google Scholar] [CrossRef]
- Deng, H.; Wang, Y.; Lei, J.R.; Chen, Z.Z.; Liang, Z.Q.; Zeng, N.K. Four New Species of Strobilomyces (Boletaceae, Boletales) from Hainan Island, Tropical China. J. Fungi 2023, 9, 1128. [Google Scholar] [CrossRef]
- San Martín, F.; Rogers, J.D.; Lavín, P. Algunas especies de Xylaria (Pyrenomycetes, Sphaeriales) habitantes en hojarasca de bosques Mexicanos. Rev. Mex. Micol. 1997, 13, 58–69. [Google Scholar]
- Lloyd, C.G. Mycological notes no. 73. Mycol. Writ. 1924, 7, 1301–1332. [Google Scholar]
- Hladki, A.I.; Romero, A.I. A preliminary account of Xylaria in the Tucuman Province, Argentina, with a key to the known species from the Northern Provinces. Fungal Divers. 2010, 42, 79–96. [Google Scholar] [CrossRef]
- Ma, H.X.; Vasilyeva, L.; Li, Y. A new species of Xylaria from China. Mycotaxon 2011, 116, 151–155. [Google Scholar] [CrossRef]
- Samuels, G.J.; Rogerson, C.T. New Ascomycetes from the Guayana Highland. Mem. N. Y. Bot. Garden 1990, 64, 165–183. [Google Scholar]
- Rogers, J.D.; Scott, D.; Ju, Y.M. Xylaria pisoniae sp. nov. from Pisonia leaves in Hawaii. Harv. Pap. Bot. 2001, 6, 189–191. [Google Scholar]
- Pande, A. Contribution to the Xylariaceae of Western India II. Nova Hedwig. 1973, 24, 13–16. [Google Scholar]
- San Martín, F.; Lavín, P.; Rogers, J.D. Some species of Xylaria (Hymenoascomycetes, Xylariaceae) associated with oaks in México. Mycotaxon 2001, 79, 337–360. [Google Scholar]
- Huang, G.; Guo, L.; Liu, N. Two new species of Xylaria and X. diminuta new to China. Mycotaxon 2014, 129, 149–152. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Substrate/Origin | Specimen No. | GenBank No. | Reference | ||
|---|---|---|---|---|---|---|
| ITS | β-Tubulin | RPB2 | ||||
| Xylaria acuminatilongissima | Termite nests/China | HAST 623 | EU178738 | GQ502711 | GQ853028 | [5] |
| X. aethiopica | Pods of Millettia ferruginea/Ethiopia | YMJ 1136 | MH790445 | MH785221 | MH785222 | [26] |
| X. adscendens | Wood/Guadeloupe | HAST 570 | GU300101 | GQ487708 | GQ844817 | [5] |
| X. allantoidea | Trunk/China | HAST 94042903 | GU324743 | GQ502692 | GQ848356 | [5] |
| X. amphithele | Dead leaves/Guadeloupe | HAST 529 | GU300083 | GQ478218 | GQ844796 | [5] |
| X. apoda | Bark/China | HAST 90080804 | GU322437 | GQ495930 | GQ844823 | [5] |
| X. arbuscula | Bark/China | HAST 89041211 | GU300090 | GQ478226 | GQ844805 | [5] |
| X. arbuscula var. plenofissura | Wood/China | HAST 93082814 | GU339495 | GQ478225 | GQ844804 | [5] |
| X. aristata | Petioles/Indonesia | YMJ 1823 (HAST 145971) | OQ883719 | NA | NA | [14] |
| X. atrodivaricata | Termite nests/China | HAST 95052001 | EU178739 | GQ502713 | GQ853030 | [5] |
| X. badia | Bamboo culm/China | HAST 95070101 | GU322446 | GQ495939 | GQ844833 | [5] |
| X. bambusicola | Bamboo culm/Thailand | JDR 162 | GU300088 | GQ478223 | GQ844801 | [5] |
| X. berteri | Bark/USA | JDR 256 | GU324750 | GQ502698 | GQ848363 | [5] |
| X. berteri | Bark/China | HAST 90112623 | GU324749 | AY951763 | GQ848362 | [5] |
| X. betulicola | Leaves of Betula/China | FCATAS 750 | MF774332 | NA | NA | [15] |
| X. brunneovinosa | Termite nests/China | HAST 720 | EU179862 | GQ502706 | GQ853023 | [5] |
| X. castorea | Wood/New Zealand | PDD 600 | GU324751 | GQ502703 | GQ853018 | [5] |
| X. cirrata | Termite nests/China | HAST 664 | EU179863 | GQ502707 | GQ853024 | [5] |
| X. coccophora | Wood/French | HAST 786 | GU300093 | GQ487701 | GQ844809 | [5] |
| X. crinalis | Wood/China | FCATAS 751 | MF774330 | NA | NA | [15] |
| X. crozonensis | Bark/France | HAST 398 | GU324748 | GQ502697 | GQ848361 | [5] |
| X. cubensis | Log/Russian Far East | HAST 477 | NA | GQ502699 | GQ848364 | [5] |
| X. culleniae | Pod/Thailand | JDR 189 | GU322442 | GQ495935 | GQ844829 | [5] |
| X. delicatula | On decaying leaves/French Guiana | GS 2775 (HAST 145973) | OQ883720 | NA | NA | [14] |
| X. diaoluoshanensis | Fallen leaves/China | HAFFR 115 | OR702611 | OR726655 | NA | This study |
| X. diaoluoshanensis | Fallen leaves/China | HAFFR 117 (HT) | OR702612 | OR726656 | OR757125 | This study |
| X. diaoluoshanensis | Fallen leaves/China | HAFFR 127 | OR702613 | OR726657 | NA | This study |
| X. escharoidea | Termite nests/China | HAST 658 | EU179864 | GQ502709 | GQ853026 | [5] |
| X. fabacearum | Seed pods of Fabaceae/Thailand | MFLU 16-1061 | NR171104 | MT212220 | MT212202 | [27] |
| X. fabaceicola | Seed pods of Fabaceae/Thailand | MFLU 16-1072 | NR171103 | MT212219 | MT212201 | [27] |
| X. feejeensis | Bark/China | HAST 92092013 | GU322454 | GQ495947 | GQ848336 | [5] |
| X. ficicola | Fallen leaves and petioles of Ficus auriculata/China | HMJAU 22818 | MZ351258 | NA | NA | [13] |
| X. filiformis | Herbaceous stem/Iran | GUM 1052 | KP218907 | NA | NA | [28] |
| X. fimbriata | Termite nests/French West Indies | HAST 491 | GU324753 | GQ502705 | GQ853022 | [5] |
| X. fulvotomentosa | Fallen leaves/China | HAFFR 124 (HT) | OR702619 | OR726658 | OR757121 | This study |
| X. fulvotomentosa | Fallen leaves/China | HAFFR 129 | OR702620 | OR726659 | OR757122 | This study |
| X. cf. glebulosa | Fruit/French West Indies | HAST 431 | GU322462 | GQ495956 | GQ848345 | [5] |
| X. grammica | Wood/China | HAST 479 | GU300097 | GQ487704 | GQ844813 | [5] |
| X. griseosepiacea | Termite nests/China | HAST 641 | EU179865 | GQ502714 | GQ853031 | [5] |
| X. hedyosmicola | Fallen leaves of Hedyosmum orientale/China | FCATAS 856 | MZ227121 | MZ221183 | MZ683407 | [13] |
| X. hedyosmicola | Fallen leaves of Hedyosmum orientale/China | FCATAS 857 | MZ227023 | MZ221184 | MZ851780 | [13] |
| X. hypoxylon | Wood/Belgium | HAST 152 | GU300096 | GQ260187 | GQ844812 | [5] |
| X. hypoxylon | Wood/China | HAST 95082001 | GU300095 | GQ487703 | GQ844811 | [5] |
| X. ianthinovelutina | Fruit of Swietenia/Martinique | HAST 553 | GU322441 | GQ495934 | GQ844828 | [5] |
| X. intraflava | Termite nests/China | HAST 725 | EU179866 | GQ502718 | GQ853035 | [5] |
| X. juruensis | Arenga engleri/China | HAST 92042501 | GU322439 | GQ495932 | GQ844825 | [5] |
| X. laevis | Wood/Martinique | HAST 419 | GU324746 | GQ502695 | GQ848359 | [5] |
| X. leavis | Bark/China | HAST 95072910 | GU324747 | GQ502696 | GQ848360 | [5] |
| X. lindericola | Fallen leaves of Lindera robusta/China | FCATAS 852 | MZ005635 | MZ031978 | MZ031982 | [13] |
| X. lindericola | Fallen leaves of Lindera robusta/China | FCATAS 853 | MZ005636 | MZ031979 | MZ048749 | [13] |
| X. liquidambaris | Fruits of Liquidambar formosana/China | HAST 93090701 | GU300094 | GQ487702 | GQ844810 | [5] |
| X. longissima | Wood/China | FCATAS 749 | MF774331 | NA | NA | [15] |
| X. longissima | Wood/Iran | IRAN 16582 F | KP218906 | NA | NA | [28] |
| X. meliacearum | Petioles and infructescence of Guarea guidonia/Puerto Rico | JDR 148 | GU300084 | GQ478219 | GQ844797 | [5] |
| X. minuscula | Fallen leaves of Castanopsis carlesii var. Sessilis/China | YMJ 90102701 (HAST 145978 HOLOTYPE) | OQ883721 | NA | NA | [14] |
| X. multiplex | Wood/USA | JDR 259 | GU300099 | GQ487706 | GQ844815 | [5] |
| X. muscula | Dead branch/French West | HAST 520 | GU300087 | GQ478222 | GQ844800 | [5] |
| X. nigripes | Termite nests/China | HAST 653 | GU324755 | GQ502710 | GQ853027 | [5] |
| X. oxyacanthae | Fallen seeds/USA | JDR 859 | GU322434 | GQ495927 | GQ844820 | [5] |
| X. oxyacanthae | Fruits/Germany | LZ 2010-502 | HQ414587 | NA | NA | [29] |
| X. palmicola | Fruits/New Zealand | PDD 604 | GU322436 | GQ495929 | GQ844822 | [5] |
| X. petchii | Fallen branches/China | HAFFR 60 | OR702616 | OR735171 | NA | This study |
| X. petchii | Fallen leaves of Daphniphyllum paxianum/China | HAFFR 118 | OR702617 | OR735172 | OR757123 | This study |
| X. petchii | Fallen leaves of Daphniphyllum paxianum/China | HAFFR 126 | OR702618 | OR735173 | OR757124 | This study |
| X. phyllocharis | Dead leaves/French West | HAST 528 | GU322445 | GQ495938 | GQ844832 | [5] |
| X. plebeja | Trunk/China | HAST 91122401 | GU324740 | GQ502689 | GQ848353 | [5] |
| X. polymorpha | Wood/USA | JDR 1012 | GU322460 | GQ495954 | GQ848343 | [5] |
| X. polymorpha | Stump/Germany | M:M-0125909 | FM164944 | NA | NA | [30] |
| X. polysporicola | Fallen leaves of Polyspora hainanensis/China | FCATAS 848 | MZ005592 | MZ031976 | MZ031980 | [13] |
| X. polysporicola | Fallen leaves of Polyspora hainanensis/China | FCATAS 849 | MZ005591 | MZ031977 | MZ031981 | [13] |
| X. reevesiae | Fruits of Reevesia formosana/China | HAST 90071609 | GU322435 | GQ495928 | GQ844821 | [5] |
| X. regalis | Log of Ficus racemose/India | HAST 920 | GU324745 | GQ502694 | GQ848358 | [5] |
| X. rogersii | Fruits of Magnolia sp./China | FCATAS 915 | MZ648827 | NA | MZ707121 | [4] |
| X. schimicola | Fruits of Schima noronhae/China | FCATAS 896 | MZ648850 | MZ695787 | MZ707114 | [4] |
| X. schweinitzii | Bark/China | HAST 92092023 | GU322463 | GQ495957 | GQ848346 | [5] |
| X. sicula f. major | Fallen leaves/China | HAST 90071613 | GU300081 | GQ478216 | GQ844794 | [5] |
| X. simplicissima | On herbaceous stems/Finland | MP 111004 (HAST 145982 EPITYPE) | OQ883722 | NA | NA | [14] |
| X. striata | Branch/China | HAST 304 | GU300089 | GQ478224 | GQ844803 | [5] |
| X. theaceicola | Fruits of Schima villosa/ | FCATAS 903 | MZ648848 | MZ695788 | MZ707115 | [4] |
| X. venosula | Twigs/USA | HAST 94080508 | EF026149 | EF025617 | GQ844806 | [5] |
| X. venustula | Bark/China | HAST 88113002 | GU300091 | GQ487699 | GQ844807 | [5] |
| X. vittatipiliformis | Dead leave/Guadeloupe | CLLGUAD 029 (HAST 145985 ISOTYPE) | OQ883723 | NA | NA | [14] |
| X. vivantii | Fruits of Magnolia sp./Martinique | HAST 519 | GU322438 | GQ495931 | GQ844824 | [5] |
| X. wallichii | Fruits of Schima wallichii/China | FCATAS 923 | MZ648861 | MZ695793 | MZ707118 | [4] |
| X. xylarioides | Wood/Iran | GUM 1151 | KP218909 | NA | NA | [28] |
| Hypoxylon fragiforme | Bark/France | HAST 383 | JN979420 | AY951720 | NA | [19] |
| Camillea obularia | –/Puerto Rico | ATCC 28093 | KY610384 | KX271243 | NA | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, X.; Chen, Z.; Lei, J.; Chen, X.; Wu, T.; Li, Y.; Chen, Y. Two New and One First Recorded Species of Xylaria Isolated from Fallen Leaves in Hainan Tropical Rainforest National Park in China. Diversity 2024, 16, 179. https://doi.org/10.3390/d16030179
Pan X, Chen Z, Lei J, Chen X, Wu T, Li Y, Chen Y. Two New and One First Recorded Species of Xylaria Isolated from Fallen Leaves in Hainan Tropical Rainforest National Park in China. Diversity. 2024; 16(3):179. https://doi.org/10.3390/d16030179
Chicago/Turabian StylePan, Xiaoyan, Zongzhu Chen, Jinrui Lei, Xiaohua Chen, Tingtian Wu, Yuanling Li, and Yiqing Chen. 2024. "Two New and One First Recorded Species of Xylaria Isolated from Fallen Leaves in Hainan Tropical Rainforest National Park in China" Diversity 16, no. 3: 179. https://doi.org/10.3390/d16030179