
Phylogenetic Trends in the Dissymmetrisation of Genitalia in Hadenini (Lepidoptera, Noctuidae)

1
Department Evolutionary Zoology, University of Debrecen, Egyetem tér 1., H-4032 Debrecen, Hungary
2
Heterocera Press Ltd., Szent István körút 4., H-1137 Budapest, Hungary
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(4), 248; https://doi.org/10.3390/d16040248
Submission received: 18 March 2024
/
Revised: 6 April 2024
/
Accepted: 11 April 2024
/
Published: 22 April 2024
(This article belongs to the Special Issue Speciation, Phylogenetics and Taxonomy of Lepidoptera)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The external genital appendages of Noctuidae represent correlated–coevolved elements of a complex structure. The pathways of changes are delimited by some constraints, and they are parallelly evolved in different phyletic lines. Asymmetrical male external genitalia were found in Hadenini, in Polia and its closely related genera, but also in Anarta (s. l.). In Poliina, the asymmetry of the genital capsule is mostly expressed in the Holarctic Polia nebulosa species group, forming the basal split within Polia (s. str.). Higher species diversity, as a consequence of asymmetry connected with the split of functions between the right and left sides, was observed in Polia (s. str.) and in Anarta (s. l.). The highest level of expansivity was observed in the Holarctic Anarta, where some species are migrating and becoming invasive ones, while a bulk of species populates the steppic and mountainous habitats of North America and Central Asia. Parallelly, while some genera/subgenera of the subtribe Poliina are oligotypic and/or consist of strictly localised species, the sensu stricto Polia species with asymmetrical genital capsules are widely distributed. The diversification of the Hadulina phylogenetic line should have been initiated by the Messinian aridisation crisis. The core area of diversification of Poliina was connected with the Sino-Himalayan region, followed by expansion into the Holarctic boreo-nemoral zone.
1. Introduction
The evolution of asymmetry in insect genitalia belongs to the most fascinating aspects of insect diversity. Since asymmetrical genitalia are described in nearly all orders, several hypotheses were formulated on their evolutionary origins. Most authors hypothesised that the changes in details of the genital capsule appear as results of sexual selection for the optimisation of reproductive success [1,2,3,4,5,6,7,8,9]. The changes in the co-adapted and co-evolved traits of genitalia of both sexes, however, consist of indispensable pre-conditions of any selective processes.
We simply hypothesised, as an updated re-formulation of the “classical” lock-and-key principle, that the structural elements of male and female genitalia in insects are co-evolved under some general constraints. More precisely, the external genital appendages of Noctuidae have been considered as correlated–coevolved elements of a phylogenetically inherited complex structure, “Bauplan” [10]. It was shown that the pathways of changes are delimited by some constraints, which were parallelly evolved in different phyletic lines of Noctuidae. We found that these constraints maintain the symmetry of sclerotised structures which have their own musculature (harpe/clasper). Moreover, we found that the asymmetric configuration of some sclerotised parts without their own muscles can evolve more rapidly and divergently. In such cases, the divergent allocation of different functions seems to be associated with (i) some selective advantages due to more effective stimulation (e.g., the specialised bristles on the saccular processes in Hadeninae) and (ii) by the mechanic fixation of certain parts of the genitalia during elongate copulation, which is accomplished by specialised terminal appendages (digitus, pollex, pseudo-pollex, etc.) of the valvae.
It was also shown that the elongated process of copulation is supported by the three-point fixation (the “seat belt” principle) of the sclerotised parts. This basic principle was recognised in “nested” structures of larger (uncus, valvae with harpes/claspers) and smaller (terminal clasping structures of valvae) parts of the genital capsule. It was shown that dissymmetrisation can effectively enhance the variations in the spatial geometry of these “spandrels” [11] but without the change in the “Bauplan”, which is preserved in different subtribes of Hadenini. We suggested, in terms of the study by Gregory [12], that the originally symmetrical “Bauplan” with its homologous structures represents a phyletic “heritage”, while functional dissymmetrisation driven by selective optimization is the “habitus” in which some homoplasies often occur.
To sum up, several homologous structures were recognised in the genital capsule of Noctuidae and focused onto the following structures and processes. The most general trends were described by Hacker et al. [13] as follows:
- In the male copulatory organ, the endophallus/vesica (the term “vesica” is generally used in the Noctuidae taxonomy and morphology) is long and tubular, with a subbasal diverticulum armed by a single small, spine-like cornutus, and with a stripe of fasciculate cornuti distally; the ductus ejaculatorius is positioned terminally (Poliina, Mamestrina, Hadenina) as opposed to the short, globular endophallus, without fasciculate cornuti, elongated diverticulum armed with terminal cornutus, and basally positioned ductus ejaculatorius (Discestrina);
- In the female genitalia, the appendix bursae is well differentiated, elongated or laterally positioned, with ductus seminalis originating at the posterior end of the appendix bursae (Poliina, Mamestrina, Hadenina) versus the completely reduced appendix bursae and the ductus seminalis positioned near the end of the ductus bursae (Discestrina).
Based on these general morphological traits, we formulated the following phylogenetic hypotheses:
- The secondary dissymmetrisation of homologous structures is most often connected with functional changes in which the clasping and stimulating functions of saccular extensions are bilaterally differentiated.
- Dissymmetrisation should enhance species diversity. This will be demonstrated in the phylogenetic and biogeographical patterns in the genera of subtribes Poliina and Discestrina of the subfamily Hadeninae.
- Furthermore, we have to demonstrate that the characteristics of the genital capsule and the traits connected with sperm transfer and their changes appear highly correlated and co-evolved.
2. Material and Methods
The material of our surveys is based on the taxonomic revisions of Hadeninae. During the past thirty years or so, the authors prepared more than 20.000 permanent genitalia slides, including thousands of photos and drawings documenting the genitalia of the studied specimens; most of them are also available in digitalised form. Morpho-taxonomic details of these surveys were already published in numerous, partly monographic papers and books [14,15,16,17,18].
The terminology of the parts of the copulatory organs follows the standard works on Lepidoptera [19,20], the recent publication on Cordero and Baixeras [4], and the publications on Noctuidae [21,22,23,24,25], respectively. The subdivision of subfamilies, tribes, and subtribes generally follows that outlined by Hacker et al. [13] with modifications by Lafontaine and Schmidt [26]. Nomenclature of genera and subgenera generally follows the checklist of the Palaearctic Noctuidae [27] with the exception of a few personal decisions.
As the characteristics of the clasping apparatus and the traits connected with sperm transfer and their changes are not independent but highly correlated, we neither constructed a data matrix nor attempted to conduct a strict cladistic analysis based on several independent characteristics.
The taxonomic list of the species and all details mentioned in this survey are provided in the Supplementary Materials.
3. Results
3.1. Morphological Trends in the Asymmetry of the Genital Capsule: Poliina
The plesiomorphic bilateral symmetry of the complex male genital capsule consists of several elementary traits. It includes the symmetrical basic configuration of the sclerotised parts of the valvae such as the dilated clavus, the shape of the saccular processes, and the bundles of bristles on the saccular processes (Figure 1). The secondary dissymmetrisation of these structures partly implies the extension and shape of the processes but, most conspicuously, the bilateral differentiation of the bundles of fixed bristles. The dissymmetrisation in the tufts of bristles on the saccular extensions is shown as a general trend connected with a subdivision of the mechanic versus sexually stimulating function of bristles (macrotricha).
In the subtribe Poliina (Hadenini), the species of certain genera/subgenera (Haderonia, Tricheurois, Protopolia, and Metallopolia) display the plesiomorphic bilaterally symmetrical genital capsule (Figure 2). The latter two subgenera of the Polia species also have nearly symmetrical basic structures, but with differentiation in the brushes of setae on the saccular processes. These genera/subgenera are restricted to the Sino-Himalayan core area, including the adjacent part of the Tibetan Plateau, and are usually represented by pairs or groups of allopatric species or subspecies (Figure 3).
Oppositely, the bulk of the much more diverse genus Polia is subdivided into several species groups with different trends of changes in the configuration of the genital capsule. They represent considerable diversity in the combination of the differentiation in the bristles and of the asymmetrical morphology of saccular processes. The differentiation of the brushes of the fixed bristles usually emerges on the left side, while the dilatation or bifurcation (e.g., P. vesperugo–P. propodea species pair (Figure 4), P. serratilinea) and the reduction in the distal part of the saccular processes (P. goliath species complex, P. piniae) typically appear on the right side (Figure 5).
The taxonomically isolated species (Polia albirena, P. subcontigua, P. griseifusa, P. hepatica, and P. malchani) only show a slight differentiation in the tufts of the bristles combined with a nearly completely symmetrical genital capsule (Figure 6). More expressed asymmetry was observed in different groups of species, especially in the P. nebulosa species group (comprising four Palearctic and four Nearctic species), which represents the basal split within Polia s. str. (see the Discussion Section). These species and the more isolated P. serratilinea show the most extreme cases of an asymmetrical shape of the saccular processes (Figure 7).
Three groups of species (the P. nebulosa, P. bombycina, and P. lamuta species groups and the P. vesperugo–P. propodea sister species pair) have a Holarctic range (see below). The species of the Southern Siberian–Western Chinese group (P. tiefi, P. vespertilio, and P. atrax) are closely allied to the P. vesperugo–P. propodea pair, and they have more or less asymmetrical saccular processes with a compact stripe of strong bristles on the left side only. The Western Chinese P. atrax, with two allopatric subspecies, is possibly the most ancient species of this group.
3.2. Morphological Trends in the Asymmetry of the Genital Capsule: Discestrina
The ancestral configuration of the genital capsule is originally bilaterally symmetrical in the subtribe Discestrina, too. We can distinguish two phyletic lineages of dissymmetrisation (Figure 8, Figure 9, Figure 10 and Figure 11):
- Odontelia–Thargelia lineage: Only the right saccular process has become elongated, often digitiform; however, the whole capsule is changed to be distorted and asymmetrical in the Thargelia species (Figure 8). The taxa of the entire lineage are geographically restricted to the eremic belts of West and Central Asia.
- Anarta (s. l.) (including Hadula, Trichoclea, and Calocestra) lineage: The saccular processes have become asymmetrically elongated and differentiated on both sides; in Hadula, it is relatively simple on the left side, and in Trichoclea and Calocestra, there is a high diversity of variations on both sides (Figure 9). The species of these genera show a considerable ecological divergence from the eremic belts (Hadula, Trichoclea) to multizonal ranges (Calocestra).
The originally symmetrical configuration appears in both major clades in the moderately diverse genera (Odontelia vs. Cardepia and Cardiestra). With the exceptions of a few Mediterranean species of the genus Cardepia, nearly all species of these groups are connected with the West and Central Asiatic eremic, often halophytic habitats (Figure 10). The symmetrical configuration of the genital capsule parallelly turned into a secondarily asymmetrical one, i.e., the dissymmetrical structures of valvae in the species-rich Palearctic Thargelia (with endophagous larvae feeding in subterraneous parts of desert plants) vs. Anarta (including the multi-diverse subgenera Hadula, Trichoclea, and Calocestra). In both phyletic lines, most derived asymmetrical valvae are associated, as a rule, with the dissymmetrisation of the juxta as well (Figure 11).
3.3. Phylogenetic Diversification as Consequence of Dissymmetry: “Lock-and-Key” Structures in Poliina vs. Discestrina—Trends and Trade-Offs
The basic divergence between Poliina versus Discestrina appears in the configuration of the female genitalia (“lock”) as well, together with associated changes in the male “key” structures. The phylogenetically most significant characteristics of the female genitalia are the positions of the appendix bursae and ductus seminalis; these correspond to the configuration of the male endophallus being responsible for spermatophore transfer. In Poliina, the appendix bursae is strongly differentiated, mostly saccate in Haderonia, Protopolia, Metallopolia, and Polia (Figure 12), while it is tubular and often rugose in Tricheurois and Ctenoceratoda (Figure 13). In all species of these genera, the endophallus is long, tubular, and usually armed distally with a long fascia of cornuti terminated close to the ductus ejaculatorius. Oppositely, in all species of the Discestrina phyletic line, the appendix bursae is extremely reduced and closely allied to the distal edge of the ductus bursae (Figure 14). In these genera, the endophallus is strongly simplified, and its favourable nesting in the bursa copulatrix is ensured by the single elongate diverticulum ending in a small cornutus only.
The corresponding traits in the male sperm transfer apparatus and female genitalia can be summarised as follows:
- The elongate tubular endophallus with a terminal ductus ejaculatorius is associated with a strongly differentiated appendix bursae in Poliina;
- The simplified endophallus with a basally located ductus ejaculatorius is associated with an extremely reduced appendix bursae in the Discestrina.
In Poliina, the configuration of the female genitalia is basically uniform. The whole antrum–ductus bursae–corpus bursae + appendix bursae apparatus is co-adapted to the elongate aedeagus and the long, tubular endophallus in the most extreme form in the Ctenoceratoda species. Among the Polia species, the ductus bursae is the longest, with a mostly extended, funnel-shaped antrum in the basally differentiated P. nebulosa group and a mostly shortened one in the Arctic-Boreal species P. richardsoni, P. lamuta, and P. rogenhoferi.
In Discestrina, the female genitalia are much more uniform, the ductus bursae is short and distally funnel-shaped, and the bursa copulatrix consists of a simple globular and a narrower rugulose part with a reduced appendix bursae.
The changes in the correlated characteristics of male and female genitalia show some trade-offs both in Poliina and Discestrina:
- The simplified genital capsule is associated with a sophisticated “lock-and-key” structure in the diverse genus Ctenoceratoda. The “loop” of the endophallus interlocks with the globular corpus bursae, and the long tubular section of the endophallus is armed by the fasciculate cornute, which fits to the elongated, tubular appendix bursae (Figure 15).
- The sophisticated genital capsule is associated with a somewhat simplified “lock-and-key”; in the multi-diverse Polia, even the fasciculate cornuti are sometimes reduced.
- Independently, a similar trend was found in both phyletic lines of the Discestrina clade as well, in Hadula, Trichoclea, and Calocestra.
Interestingly, we could not find any phylogenetic trend in the number and configuration of the signa in the corpus bursae, since the presence of versus reduction in this trait was rather eclectic in the different species groups and in closely related species.
4. Discussion
Asymmetrical male external genitalia have been regularly observed in several generic groups of Hadenini, e.g., in Polia and its closely related genera, but also in Anarta (including several subgenera, e.g., Hadula and Calocestra). In both large clades, the bilateral saccular processes became asymmetrical. However, these two large suprageneric groups display completely different organisations of the sperm transfer system. Therefore, they were ascribed to different subtribes, Poliina vs. Discestrina [13,26], and as a consequence, they cannot be considered closely related groups within the tribe Hadenini.
The phylogenetic significance of asymmetry was shown in both target groups (subtribes). In Poliina, the asymmetry of the genital capsule was observed in different groups of species; however, it is mostly expressed in the Polia nebulosa species group. This group was already shown to be monophyletic based on the genital morphological characteristics of both sexes and prominent according to the CO1 sequence [18]. This group was already predicted [28] and recently demonstrated [29] to be the sister group of all other North American Polia. This group of species forms the basal split within Polia s. str. and is characterised by a relatively high species diversity, consisting of four Palearctic and four Nearctic species. McCabe [28] already confirmed that the four North American Polia species are closely related to the Eurasiatic P. nebulosa, and this Nearctic group of species is possibly a descendent of the “nebulosa” lineage. The with P. nebulosa most closely related P. piniae–P. discalis species-pair occurs near to the Pacific Coast, while the more derived P. nimbosa and P. imbrifera are widely distributed in the boreo-temperate zone.
In the genera/subgenera with symmetrical genital capsule, e.g., in Haderonia, Metallopolia, and Protopolia, we only observed vicarious species pairs, mostly with a South-West China vs. Himalaya disjunct range, which are evidently originated by allopatric differentiation. Oppositely, the evolved asymmetry is followed by enhanced species diversity in phylogenetic lines within Polia s. str. It emerged in a limited manner in the vicarious species of the P. bombycina group and P. propodea–P. vesperugo pair [18], and also in the circumpolar Arctic belt (the “triplet” P. lamuta, P. richardsoni, and P. rogenhoferi), but mostly in the Sino-Himalayan-Siberian region, which has phylogenetically less-known species groups (P. atrax, P. tiefi, and P. vespertilio). Unfortunately, in some cases, the sister species relations are actually only supported by genital morphological traits due to the lack of barcode data for most East Palearctic species. We experienced essentially similar hiatuses, but also some contradicting data (e.g., in the Odontelia and Thargelia species) of barcode information in the case of Hadulina moths as well.
Based on these morphological and biogeographical patterns of two specious groups of Noctuidae moths, we have to re-consider the main hypotheses formulated regarding the possible explanations of asymmetrical genitalia. Sexual selection was often hypothesised to act on the male mating structures through intrasexual competition [3,6,7,8]. For example, selection occurs in the case of polyandry when certain characteristics of male reproductive structures result in the enhanced success of spermatophore transfer. According to another hypothesis regarding female choice, if male quality varies, then females should be able to choose the sperm of higher quality males. Models were developed to determine how the males could manipulate female behaviour to their advantage. It was also hypothesised that selection also should act on females to avoid this manipulation [30,31,32,33], leading to an intersexual arms race involving the mating structures of males and females.
We think that, exclusively, neither the intrasexual nor the intersexual selectionist hypotheses are suitable to explain the trends of changes in external genitalia. It is evident that the coupling of genitalia can only follow the “trial and error/success” of pre-copulatory games (pheromones, “courtship” rituals, etc.) in which a female choice mechanism can be assumed. However, the process of sexual selection is more complex, because spermatophore transfer is a long, multi-step process [34]. In this phase of copulation, a real male–male interaction has never been observed in Noctuidae. The survey of female genitalia demonstrated that the removal of spermatophores from the bursa copulatrix by competing males can also be fully excluded. However, sperm competition between different males can be strongly supposed [35,36], as similarly positioned spermatophores of different males in the bursa copulatrix were often observed by other researchers [21,37] and in our study in many different species of the “pest clade” of Noctuids (Varga and Szanyi mscr.). Therefore, the firm clasping of genitalia during copulation functions as an elementary pre-condition of successful spermatophore transfer. Consequently, only those males will be able to optimally allocate spermatophore which (i) have successfully stimulated the female in the first phase of the copula and (ii) possesses optimally fitted genital structures for a firm clasping of genital parts during the whole spermatophore transfer process in the next phase of the copula. This means that the differentiation in the originally bilaterally symmetrical structures should have some selective advantages according to the different allocation of the main functions (i.e., dissymmetrisation); thus, fitted stimulation and clasping may enhance the optimal interlocking of genitalia. We consider that the individual (accidental and fluctuating) and geographical variations of asymmetric parts supply raw material for optimising selection, probably combined with genetic drift in allopatric, isolated populations. We hypothesise that this selection pressure should be much more expressed on the internal genitalia, as it is directly responsible for spermatophore transfer.
Stekolnikov and Kusnetzov [38] showed that the muscles attached to the genital capsule are only exceptionally asymmetrical. This fact suggests that the twisting and pitching of genitalia should only involve the terminal parts of the phallus, the endophallus, and the bursa–appendix complex, respectively, i.e., the inner genitalia. The “lock-and-key” (LKM) structures of the inner genitalia are much more strictly correlated and co-evolved than those of the external structures since “in most cases the LKMs seem to reinforce the results of the pre-copulatory isolation mechanisms (phaenology, behaviour, pheromones, mechanical devices etc.” [9]. This means that some trade-offs should evolve between external vs. internal genitalia. We showed in many cases, both in Poliina and Discestrina, that sophisticated external genitalia are combined with relatively simplified internal genitalia. The general validity of this principle was already demonstrated in some lineages of Apameini [39] and in species groups of Chersotis (Noctuinae) [40] and Ctenoceratoda (Hadenini: Poliina) [41,42].
Therefore, the most probable explanation is that higher diversity and reproductive success are consequences of asymmetry. This pathway seems to be supported by the higher species diversity connected with the “split of functions between right and left sides” [43], which was observed in Polia (s. str.) vs. Metallopolia, Protopolia, Haderonia and Tricheurois, and in Anarta s.l. (see the different subgenera), but also in Thargelia, compared with Cardiestra and Odontelia, which have symmetrical genital capsules. Similar cases of secondary asymmetry were shown, however, in phylogenetically distant groups of Lepidoptera (e.g., Hesperidae: Erynnis perseus) [44], which can be considered analogous configurations of the saccular processes of Polia.
We think that higher species diversity and expansion capacity are parallelly evolved in both the Poliina and Discestrina phyletic lineages. The highest level of expansivity was observed in the Holarctic Anarta (subgenera Trichoclea/Calocestra) clade, where some species were described as migrating and becoming invasive ones, such as Anarta (Trichoclea) trifolii, while a considerable bulk of species populates the arid steppic and mountainous habitats of North America and Central Asia. Parallelly, some genera/subgenera of the subtribe Poliina are oligotypic and/or consist of strictly localised species (Haderonia, Tricheurois, Metallopolia, and Protopolia), while the s. str. Polia species with asymmetrical genital capsule are generally widely distributed, e.g., the Euro-Siberian nemoral P. hepatica, P. nebulosa, P. bombycina, the Euro-Siberian Boreal P. vesperugo, the Trans-Palearctic steppic-montane P. serratilinea, and the circumpolar Arctic P. richardsoni. Other species expanded from the Inner Asiatic area and Southern Siberian mountains to the Urals, such as P. subcontigua, P. malchani, and P. vespertilio. Furthermore, there are some vicarious sister species or closely related species groups in North America and Eurasia, such as the P. nebulosa group, which constitutes the basal split within Polia s.str., the P. bombycina vs. P. purpurissata + P. nugatis “triplets”, the P. vesperugo–P. propodea pair, etc. [18].
Hypothetically, the core area of ancestral diversification of the Hadulina phylogenetic line should have been connected with the ancient Mediterranean area, possibly initiated by the Messinian aridisation crisis, as it was shown in different other groups such as cicadas, sandflies, scorpions, reptiles, etc. [45,46,47,48,49]. Oppositely, the ancient core area of diversification of Poliina was connected with the tectonic events in the Sino-Himalayan (“Ancient Kathaysian”) region, which was suggested earlier by Varga [50,51],followed by the expansion in repeated waves into the Holarctic boreo-nemoral zone, (i) first following the “Rhododendron-corridor” and then (ii) crossing an aridity filter into Central and Inner Asia.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/d16040248/s1, The full names of the genus and species-level taxa mentioned in the article.
Author Contributions
Conceptualization, Z.V. and L.R.; investigation, Z.V., G.R. and L.R.; resources, Z.V., G.R. and L.R.; data curation; Z.V., G.R. and L.R.; writing—original draft preparation, Z.V.; writing—review and editing, Z.V., G.R. and L.R.; visualization, G.R. and L.R.; project administration, Z.V.; funding acquisition, Z.V., G.R. and L.R. All authors have read and agreed to the published version of the manuscript.
Funding
Z. Varga was supported by the Hungarian National Research Foundation OTKA, Grant No. T73597, and by the SYNTHESYS Project, Grants No. DE-TAF-6708; G. Ronkay was supported by the DE-TAF-3514 and AT-TAF-5609 grants; and L. Ronkay was supported by the FR-TAF-562 and SE-TAF-6919 grants.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article and in the Supplementary Material, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors are grateful to Wolfram Mey and Théo Léger (Museum für Naturkunde, Berlin), Martin Honey and Alberto Zilli (Natural History Museum, London), Sabine Gaal-Haszler and Martin Lödl (Naturhistorisches Museum, Vienna), Robert Trusch and Michael Falkenberg (Landessammlungen für Naturkunde, Karlsruhe), Bernard Landry (Museum d’Histoire Naturelle, Genève), and Dieter Stüning (Zoological Research Museum Alexander Koenig, Bonn) for providing us with the possibility of studying valuable collection materials, including types.
Conflicts of Interest
The authors and the Heterocera Press Ltd. declare no conflicts of interest.
References
- Arnqvist, G. Comparative evidence for the evolution of genitalia by sexual selection. Nature 1998, 393, 784–786. [Google Scholar] [CrossRef]
- Arnqvist, G. Antagonistic coevolution between the sexes in a group of insects. Nature 2002, 415, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Arnqvist, G.; Edvardsson, M.; Friberg, U.; Nilsson, T. Sexual conflict promotes speciation in insects. Proc. Natl. Acad. Sci. USA 2000, 97, 10460–10464. [Google Scholar] [CrossRef] [PubMed]
- Cordero, C.; Baixeras, J. Sexual Selection Within the Female Genitalia in Lepidoptera. In Cryptic Female Choice in Arthropods; Springer: New York, NY, USA, 2015; pp. 325–350. [Google Scholar]
- Eberhard, W.G. Sexual Selection and Animal Genitalia; Harvard University Press: Cambridge, MA, USA, 1985; 244p. [Google Scholar]
- Eberhard, W.G. Evaluating models of sexual selection: Genitalia as a test case. Am. Nat. 1993, 142, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, W.G.; Lehmann, G.U.C. Demonstrating sexual selection by cryptic female choice on male genitalia: What is enough? Evolution 2019, 73, 2415–2435. [Google Scholar] [CrossRef] [PubMed]
- Hosken, D.J.; Stockley, P. Sexual selection and genital evolution. Trends Ecol. Evol. 2004, 19, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, K. The lock-and-key mechanisms of the internal genitalia of the Noctuidae (Lepidoptera): How are they selected for? Eur. J. Entomol. 2008, 105, 13–25. [Google Scholar] [CrossRef]
- Varga, Z.; Ronkay, L. Structural constraints of secondary asymmetry in male external genitalia of Noctuidae. Insect Syst. Evol. 2013, 44, 349–372. [Google Scholar] [CrossRef]
- Gould, S.J. The exaptive excellence of spandrels as a term and prototype. Proc. Natl. Acad. Sci. USA 1997, 94, 10750–10755. [Google Scholar] [CrossRef] [PubMed]
- Gregory, W.K. Habitus factors in the skeleton of fossil and recent mammals. Proc. Am. Philos. Soc. 1936, 76, 429–444. [Google Scholar]
- Hacker, H.; Ronkay, L.; Hreblay, M. Hadeninae I. Noctuidae Europaeae Volume 4; Entomological Press: Sorø, Denmark, 2002; 419p. [Google Scholar]
- Ronkay, L.; Varga, Z. On the taxonomy of the genera Odontelia Hampson, 1905, and Thargelia Püngeler, 1900 (Noctuidae, Hadeninae). Ann. Hist.-Nat. Musei Natl. Hung. 1998, 90, 175–193. [Google Scholar]
- Varga, Z.; Ronkay, L.; Ronkay, G. Metallopolia, a new subgenus of Polia, with the description of two new species and a new subspecies (Noctuidae, Noctuinae, Hadenini). J. Asia-Pac. Entomol. 2017, 21, 217–232. [Google Scholar] [CrossRef]
- Varga, Z.; Ronkay, L.; Ronkay, G. Revised taxonomic check list of the Eurasiatic species of the subtribe Poliina (Noctuidae, Noctuinae, Hadenini). Dtsch. Entomol. Z. 2017, 64, 133–160. [Google Scholar] [CrossRef]
- Varga, Z.; Gyulai, P.; Ronkay, G.; Ronkay, L. Review of the species groups of the genus Ctenoceratoda Varga, 1992 with description of four new species and a new subspecies (Lepidoptera, Noctuidae). Acta Zool. Acad. Sci. Hung. 2018, 64, 51–74. [Google Scholar] [CrossRef]
- Varga, Z.; Ronkay, G.; Ronkay, L. Taxonomic survey of the Polia (Polia) nebulosa species complex (Noctuidae, Noctuinae, Hadenini) with the description of two new subspecies. Acta Zool. Acad. Sci. Hung. 2019, 65, 107–122. [Google Scholar] [CrossRef]
- Hodges, R.W. Sphingoidea. In The Moths of America North of Mexico Volume 21; Wedge Entomological Research Foundation: Washington, DC, USA, 1971; pp. 1–158, pl. 1–14. [Google Scholar]
- Heppner, J.B. Butterflies and Moths (Lepidoptera). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: New York, NY, USA, 2008; pp. 627–675. [Google Scholar]
- Lafontaine, J.D. Noctuoidea, Noctuidae (Part) Noctuinae (Part-Euxoa). In The Moths of America North of Mexico, Fascicle 27.2; Dominick, R.B., Ed.; Wedge Entomological Research Foundation: Washington, DC, USA, 1987; 238p. [Google Scholar]
- Lafontaine, J.D. Noctuoidea, Noctuidae (part Noctuinae II). In The Moths of America North of Mexico, Fascicle 27.3; Dominick, R.B., Ed.; Wedge Entomological Research Foundation: Washington, DC, USA, 1998; 348p. [Google Scholar]
- Fibiger, M. Noctuidae Europaeae Volume 3. Noctuinae III; Entomological Press: Sorø, Denmark, 1997; 418p. [Google Scholar]
- Fibiger, M.; Lafontaine, J.D. A review of the higher classification of Noctuoidea (Lepidoptera) with special reference to the Holarctic fauna. Esperiana 2005, 11, 7–92. [Google Scholar]
- Mikkola, K.; Lafontaine, J.D.; Gill, J. Noctuoidea: Noctuidae (part): Xyleninae (part): Apameini (part-Apamea group of genera). In The Moths of America North of Mexico. Fascicle 26.9; Hodges, R.W., Ed.; The Wedge Entomological Research Foundation: Washington, DC, USA, 2009; 192p. [Google Scholar]
- Lafontaine, D.; Schmidt, B.C. Annotated check list of the Noctuoidea (Insecta, Lepidoptera) of North America North of Mexico. ZooKeys 2010, 40, 40. [Google Scholar] [CrossRef]
- Fibiger, M.; Hacker, H.H. Systematic List of the Noctuoidea of Europe (Notodontidae, Nolidae, Arctiidae, Lymantriidae, Erebidae, Micronoctuidae, and Noctuidae). Esperiana 2005, 11, 93–182. [Google Scholar]
- McCabe, T.M. A reclassification of the Polia complex for North America (Lepidoptera: Noctuidae). In New York State Museum, Bulletin; The University of the State of New York: Albany, NY, USA, 1980; 432, i–vi, pp. 1–141. [Google Scholar]
- Zahiri, R.; Lafontaine, J.D.; Schmidt, B.C.; deWaard, J.R.; Zakharov, E.V.; Hebert, P.D.N. A transcontinental challenge—A test of DNA barcode performance for 1,541 species of Canadian Noctuoidea (Lepidoptera). PLoS ONE 2014, 9, e92797. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.D.; Marshall, D.C.; Cooley, J.R. Evolutionary perspectives on insect mating. In The Evolution of Mating Systems in Insects and Arachnids; Cambridge University Press: Cambridge, UK, 2010; pp. 4–31. [Google Scholar] [CrossRef]
- Holland, B.; Rice, W.R. Experimental removal of sexual selection reverses intersexual antagonistic coevolution and removes a reproductive load. Proc. Natl. Acad. Sci. USA 1999, 96, 5083–5088. [Google Scholar] [CrossRef] [PubMed]
- Gavrilets, S.; Arnqvist, G.; Friberg, U. The evolution of female mate choice by sexual conflict. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Cordero, C.; Eberhard, W.G. Female choice of sexually antagonistic male adaptations: Critical review of some recent research. J. Evol. Biol. 2003, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Callahan, P.S.; Chapin, J.B. Morphology of the reproductive system and mating in two representative members of the family Noctuidae, Pseudaletia unipuncta and Peridroma margaritosa with comparison to Heliothis zea. Ann. Entomol. Soc. Am. 1960, 53, 763–782. [Google Scholar] [CrossRef]
- Drummond, B.A. Multiple mating and sperm competition in the Lepidoptera. In Sperm Competition and the Evolution of Animal Mating Systems; Smith, R.L., Ed.; Academic Press: Orlando, FL, USA, 1984; pp. 291–371. [Google Scholar]
- Birkhead, T.; Møller, A.P. (Eds.) Sperm Competition and Sexual Selection; Academic Press: San Diego, CA, USA, 1998; 826p. [Google Scholar]
- Mikkola, K. Evidence for lock-and-key mechanisms in the internal genitalia of the Apamea moths (Lep.: Noctuidae). Syst. Entomol. 1992, 17, 145–153. [Google Scholar] [CrossRef]
- Stekolnikov, A.A.; Kuznetzov, V.I. Functional morphology of the male genitalia and a new tribal division of the Ennominae (Lepidoptera, Geometridae). Entomol. Rev. 1982, 61, 92–122. [Google Scholar]
- Zilli, A.; Varga, Z.; Ronkay, G.; Ronkay, L. Apameini I. Taxonomic Atlas of the Eurasian and North African Noctuoidea, Heterocera Press: Budapest, Hungary, 2009; Volume 3, 393p.
- Varga, Z.; Ronkay, L. New and revised taxa of the genera Chersotis Boisduval, 1840 and Dichagyris Lederer, 1857 from Central Asia (Lep.: Noctuidae). Esperiana 1996, 5, 175–214. [Google Scholar]
- Varga, Z. Taxonomic notes on the Genus Haderonia Staudinger, 1896 with the description of a new Genus Ctenoceratoda and four new species (Lep.: Noctuidae). Acta Zool. Hung. Acad. Sci. Hung. 1992, 38, 95–112. [Google Scholar]
- Varga, Z.; Gyulai, P. Taxonomy of the genus Ctenoceratoda Varga, 1992 (Lep.: Noctuidae, Hadeninae) with the description of seven new species. Acta Zool. Hung. Acad. Sci. Hung. 1999, 45, 169–197. [Google Scholar]
- Huber, B.A.; Sinclair, B.I.; Schmitt, M. The evolution of asymmetric genitalia in spiders and insects. Biol. Rev. 2007, 82, 647–698. [Google Scholar] [CrossRef]
- Scott, J.A. Mating of butterflies. J. Res. Lepid. 1973, 11, 99–127. [Google Scholar] [CrossRef]
- Husemann, M.; Schmitt, T.; Zachos, F.E.; Ulrich, W.; Habel, J.C. Palaearctic biogeography revisited: Evidence for the existence of a north African refugium for Western Palaearctic biota. J. Biogeogr. 2014, 41, 81–94. [Google Scholar] [CrossRef]
- Podnar, M.; Grbac, I.; Tvrtkovic, N.; Hörweg, C.; Haring, E. Hidden diversity, ancient divergences, and tentative Pleistocene microrefugia of European scorpions (Euscorpiidae: Euscorpiinae) in the eastern Adriatic region. J. Zool. Syst. Evol. Res. 2021, 59, 1824–1849. [Google Scholar] [CrossRef]
- Salvi, D.; Pinho, C.; Mendes, J.; Harris, D.J. Fossil-calibrated time tree of Podarcis wall lizards provides limited support for biogeographic calibration models. Mol. Phylogenet. Evol. 2021, 161, 107169. [Google Scholar] [CrossRef] [PubMed]
- Trajer, A.J.; Sebestyén, V.; Padisák, J. The impacts of the Messinian salinity crisis on the biogeography of three Mediterranean sandfly (Diptera: Psychodidae) species. Geobios 2021, 65, 51–66. [Google Scholar] [CrossRef]
- Costa, G.J.; Nunes, V.L.; Marabuto, E.; Mendes, R.; Silva, D.N.; Pons, P.; Bas, J.M.; Hertach, T.; Paulo, O.S.; Simões, P.C. The effect of the Messinian salinity crisis on the early diversification of the Tettigettalna cicadas. Zool. Scr. 2023, 52, 100–116. [Google Scholar] [CrossRef]
- Varga, Z. Biogeography and Evolution of the oreal Lepidoptera in the Palearctic. Acta Zool. Hung. Acad. Sci. Hung. 1997, 42, 289–330. [Google Scholar]
- Varga, Z. Centres of endemism of Noctuidae (Lepidoptera) in the Palaearctic arid mountains: Biogeographical and phylogenetic implications. Beiträge Entomol.–Contrib. Entomol. 2022, 72, 1–35. [Google Scholar] [CrossRef]
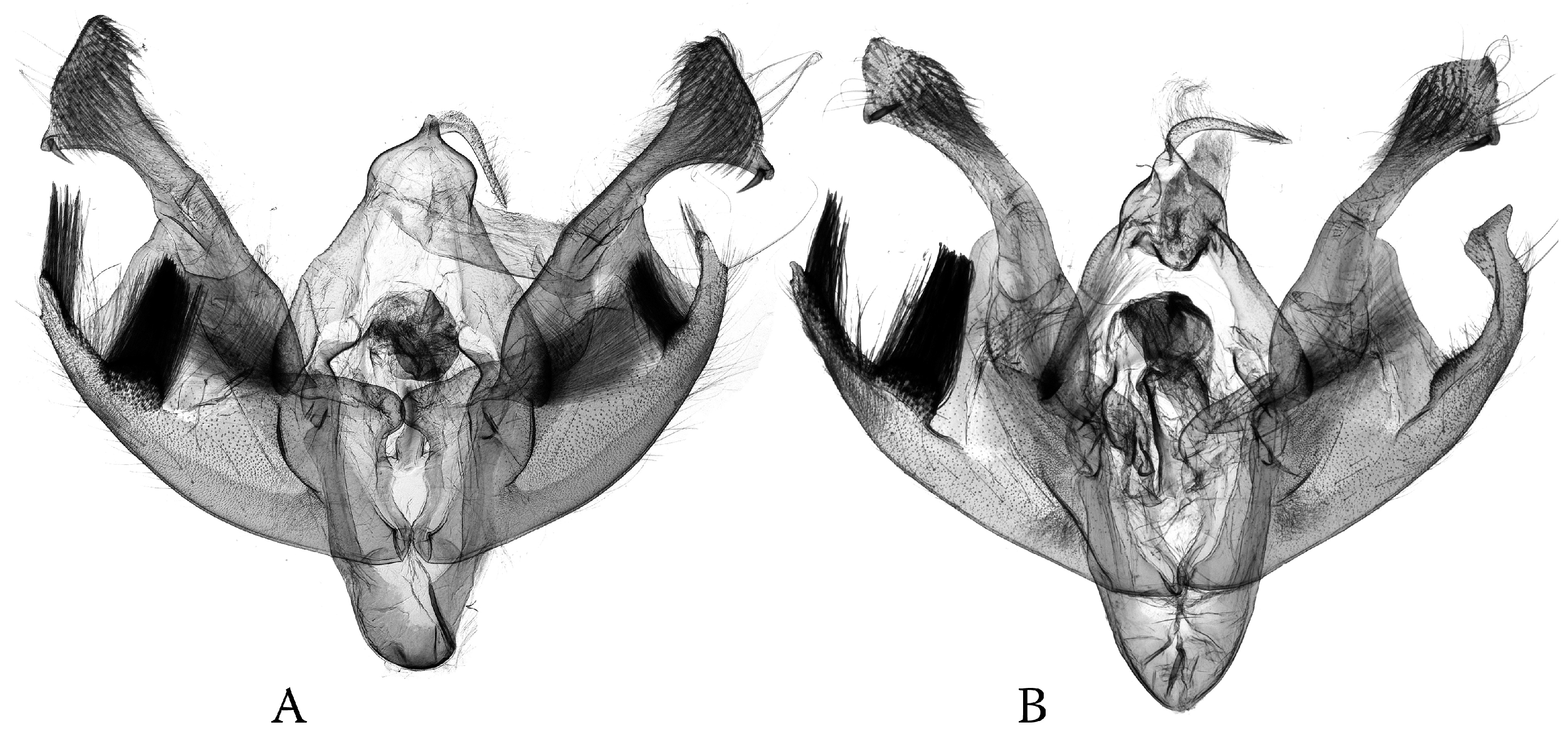
Figure 1.
Examples of symmetrical vs. asymmetrical genital capsules. (A) Haderonia arschanica with reduced saccular processes; (B) Polia (Metallopolia) kalikotei with short, symmetrical saccular processes and slightly asymmetrical setae; (C) Polia (Polia) nebulosa with strongly asymmetrical saccular processes and differentiated groups of setae (males).
Figure 1.
Examples of symmetrical vs. asymmetrical genital capsules. (A) Haderonia arschanica with reduced saccular processes; (B) Polia (Metallopolia) kalikotei with short, symmetrical saccular processes and slightly asymmetrical setae; (C) Polia (Polia) nebulosa with strongly asymmetrical saccular processes and differentiated groups of setae (males).

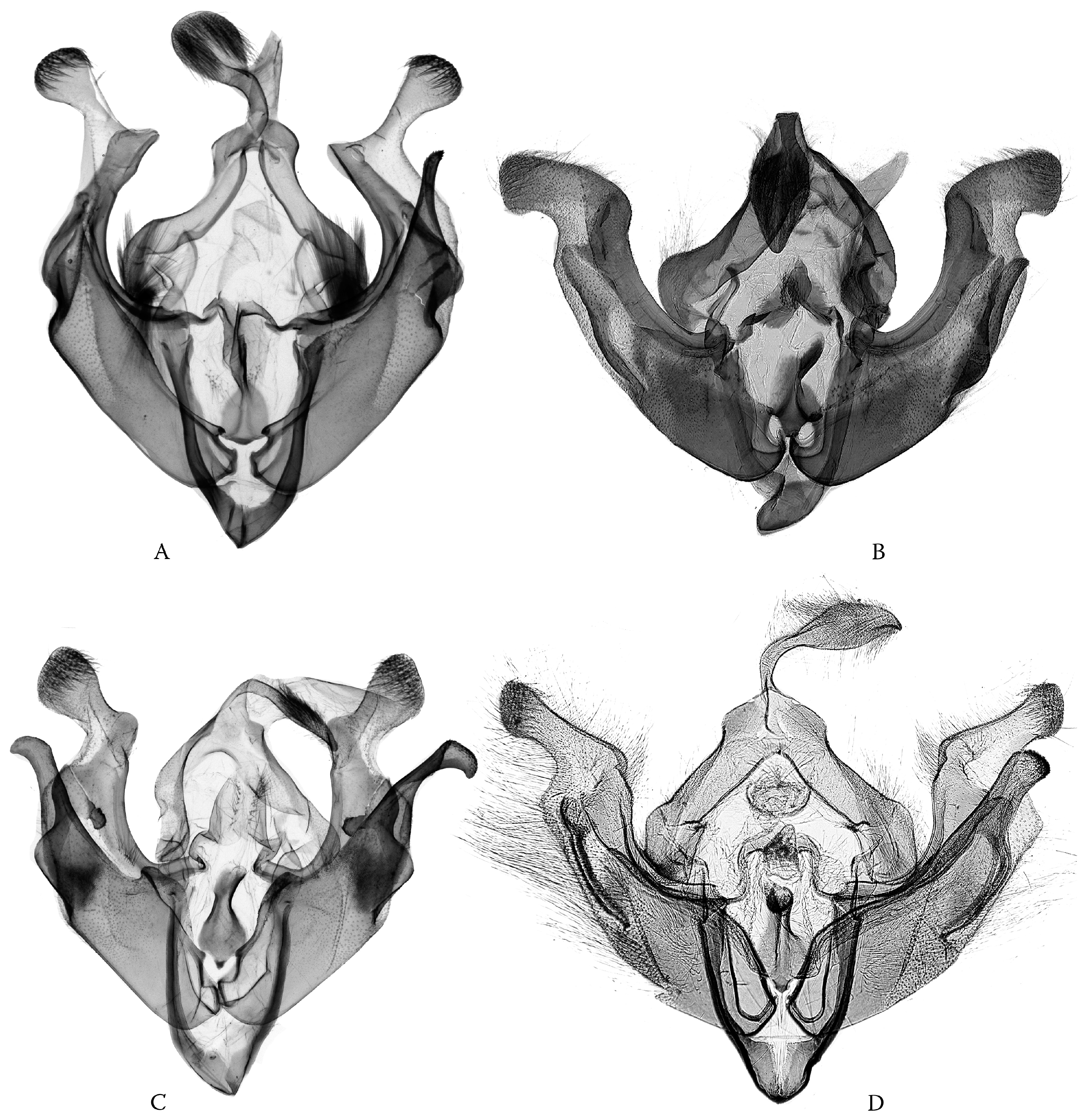
Figure 2.
Four different genera/subgenera of Poliina with nearly completely symmetrical genital capsules: (A) Haderonia miserabilis; (B) Ctenoceratoda sukharevae; (C) Tricheurois retrusa; and (D) Polia (Protopolia) praecipua (males).
Figure 2.
Four different genera/subgenera of Poliina with nearly completely symmetrical genital capsules: (A) Haderonia miserabilis; (B) Ctenoceratoda sukharevae; (C) Tricheurois retrusa; and (D) Polia (Protopolia) praecipua (males).

Figure 3.
(A) Polia (Metallopolia) culta; (C) Polia (Metallopolia) dysgnorima; (B) Polia (Metallopolia) kisculta; and (D) Polia (Metallopolia) metagnorima (males). Himalayan (A,B) vs. West China (C,D) allopatric pairs of species.
Figure 3.
(A) Polia (Metallopolia) culta; (C) Polia (Metallopolia) dysgnorima; (B) Polia (Metallopolia) kisculta; and (D) Polia (Metallopolia) metagnorima (males). Himalayan (A,B) vs. West China (C,D) allopatric pairs of species.

Figure 4.
(A) Polia (Polia) vesperugo; (B) Polia (Polia) propodea. Genital capsules of Palearctic vs. Nearctic Boreal sister species.
Figure 4.
(A) Polia (Polia) vesperugo; (B) Polia (Polia) propodea. Genital capsules of Palearctic vs. Nearctic Boreal sister species.

Figure 5.
(A) Polia (Polia) gigantea; (B) Polia (Polia) piniae. Genital capsules displaying extreme dissymmetry in Polia nebulosa group.
Figure 5.
(A) Polia (Polia) gigantea; (B) Polia (Polia) piniae. Genital capsules displaying extreme dissymmetry in Polia nebulosa group.

Figure 6.
(A) Polia (Polia) griseifusa; (B) Polia (Leuconephropolia) albirena. Genital capsules illustrating taxonomically isolated species with symmetrical structures.
Figure 6.
(A) Polia (Polia) griseifusa; (B) Polia (Leuconephropolia) albirena. Genital capsules illustrating taxonomically isolated species with symmetrical structures.

Figure 7.
(A) Polia (Polia) serratilinea serratilinea; (B) Polia (Polia) serratilinea spalax. Genital capsules—case of dissymmetry in saccular processes (allopatric subspecies).
Figure 7.
(A) Polia (Polia) serratilinea serratilinea; (B) Polia (Polia) serratilinea spalax. Genital capsules—case of dissymmetry in saccular processes (allopatric subspecies).

Figure 8.
(A) Odontelia arenicola; (B) Thargelia gigantea. Genital capsules—cases of nearly symmetrical vs. strongly dissymmetrical structures in Discestrina.
Figure 8.
(A) Odontelia arenicola; (B) Thargelia gigantea. Genital capsules—cases of nearly symmetrical vs. strongly dissymmetrical structures in Discestrina.

Figure 9.
(A) Anarta (Calocestra) farnhami palaearctica; (B) Anarta (Hadula) sabulorum; (C) Anarta (Calocestra) mendica; and (D) Anarta (Hadula) halodeserti (males). A + C vs. B + D—two phyletic lines of dissymmetrisation in Anarta (s. l.).
Figure 9.
(A) Anarta (Calocestra) farnhami palaearctica; (B) Anarta (Hadula) sabulorum; (C) Anarta (Calocestra) mendica; and (D) Anarta (Hadula) halodeserti (males). A + C vs. B + D—two phyletic lines of dissymmetrisation in Anarta (s. l.).

Figure 10.
(A) Cardiestra gobideserti; (B) Cardepia sociabilis deserticola—cases of nearly symmetrical genital capsules in Discestrina.
Figure 10.
(A) Cardiestra gobideserti; (B) Cardepia sociabilis deserticola—cases of nearly symmetrical genital capsules in Discestrina.

Figure 11.
(A) Thargelia distincta; (B) Thargelia leucostigma—cases of strongly dissymmetrical genital capsules in Discestrina.
Figure 11.
(A) Thargelia distincta; (B) Thargelia leucostigma—cases of strongly dissymmetrical genital capsules in Discestrina.

Figure 12.
(A) Polia (Metallopolia) metagnorima; (B) Polia (Polia) malchani (females)—the tubular shape of appendix bursae (A) corresponds with the elongate tubular endophallus, the short appendix bursae (B) with the simplified endophallus of males.
Figure 12.
(A) Polia (Metallopolia) metagnorima; (B) Polia (Polia) malchani (females)—the tubular shape of appendix bursae (A) corresponds with the elongate tubular endophallus, the short appendix bursae (B) with the simplified endophallus of males.

Figure 13.
(A) Tricheurois landryi; (B) Ctenoceratoda tancrei (females)—the tubular shape of the appendix bursae corresponds with the elongate tubular endophallus with fasciculate cornuti in males.
Figure 13.
(A) Tricheurois landryi; (B) Ctenoceratoda tancrei (females)—the tubular shape of the appendix bursae corresponds with the elongate tubular endophallus with fasciculate cornuti in males.

Figure 14.
(A) Anarta (Hadula) insolita; (B) Anarta (Calocestra) schawyra (females)—the appendix bursae is completely reduced in both phyletic lines of Discestrina.
Figure 14.
(A) Anarta (Hadula) insolita; (B) Anarta (Calocestra) schawyra (females)—the appendix bursae is completely reduced in both phyletic lines of Discestrina.

Figure 15.
The sophisticated “lock-and-key” configuration of the corpus + appendix bursae vs. endophallus is a generic synapomorphy of the Ctenoceratoda species (Ctenoceratoda tancrei).
Figure 15.
The sophisticated “lock-and-key” configuration of the corpus + appendix bursae vs. endophallus is a generic synapomorphy of the Ctenoceratoda species (Ctenoceratoda tancrei).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Varga, Z.; Ronkay, G.; Ronkay, L. Phylogenetic Trends in the Dissymmetrisation of Genitalia in Hadenini (Lepidoptera, Noctuidae). Diversity 2024, 16, 248. https://doi.org/10.3390/d16040248
AMA Style
Varga Z, Ronkay G, Ronkay L. Phylogenetic Trends in the Dissymmetrisation of Genitalia in Hadenini (Lepidoptera, Noctuidae). Diversity. 2024; 16(4):248. https://doi.org/10.3390/d16040248
Chicago/Turabian StyleVarga, Zoltán, Gábor Ronkay, and László Ronkay. 2024. "Phylogenetic Trends in the Dissymmetrisation of Genitalia in Hadenini (Lepidoptera, Noctuidae)" Diversity 16, no. 4: 248. https://doi.org/10.3390/d16040248
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.