
Soil-Microbial CNP Content and Ecological Stoichiometry Characteristics of Typical Broad-Leaved Tree Communities in Fanjing Mountain in Spring
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Natural Overview of the Study Area
2.2. Soil Sample Collection
2.3. Determination of Soil-Microbial Carbon, Nitrogen, and Phosphorus Concentrations
2.4. Data Processing
3. Results and Analysis
3.1. Effects of Forest Stand Type and Soil Depth on Soil-Microbial Carbon, Nitrogen, and Phosphorus Concentrations and Stoichiometry Characteristics
3.2. Characteristics of Soil Carbon, Nitrogen, Phosphorus and Stoichiometric Ratios
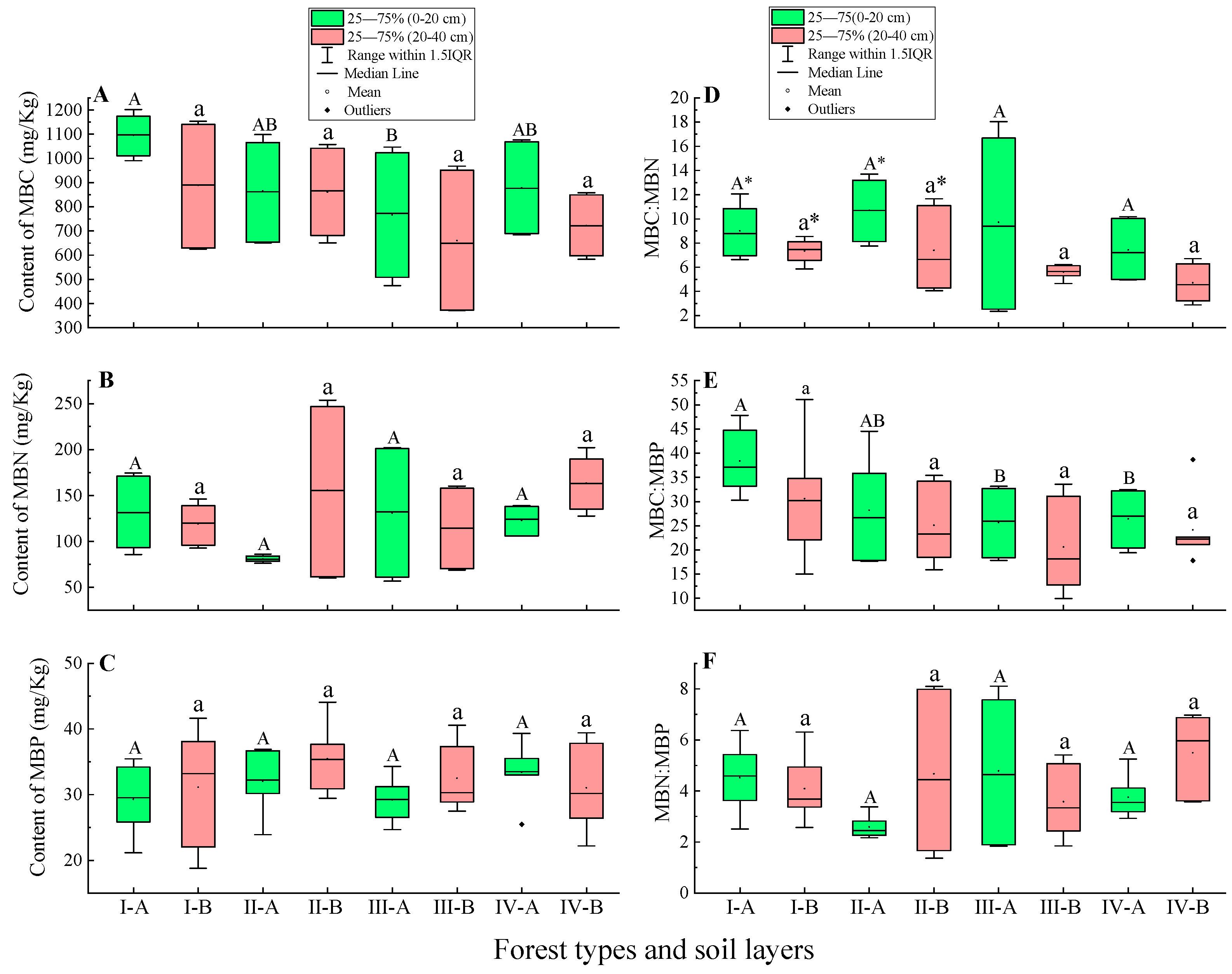
3.3. Soil Microbial Biomass Carbon, Nitrogen, Phosphorus Content and Stoichiometry Characteristics
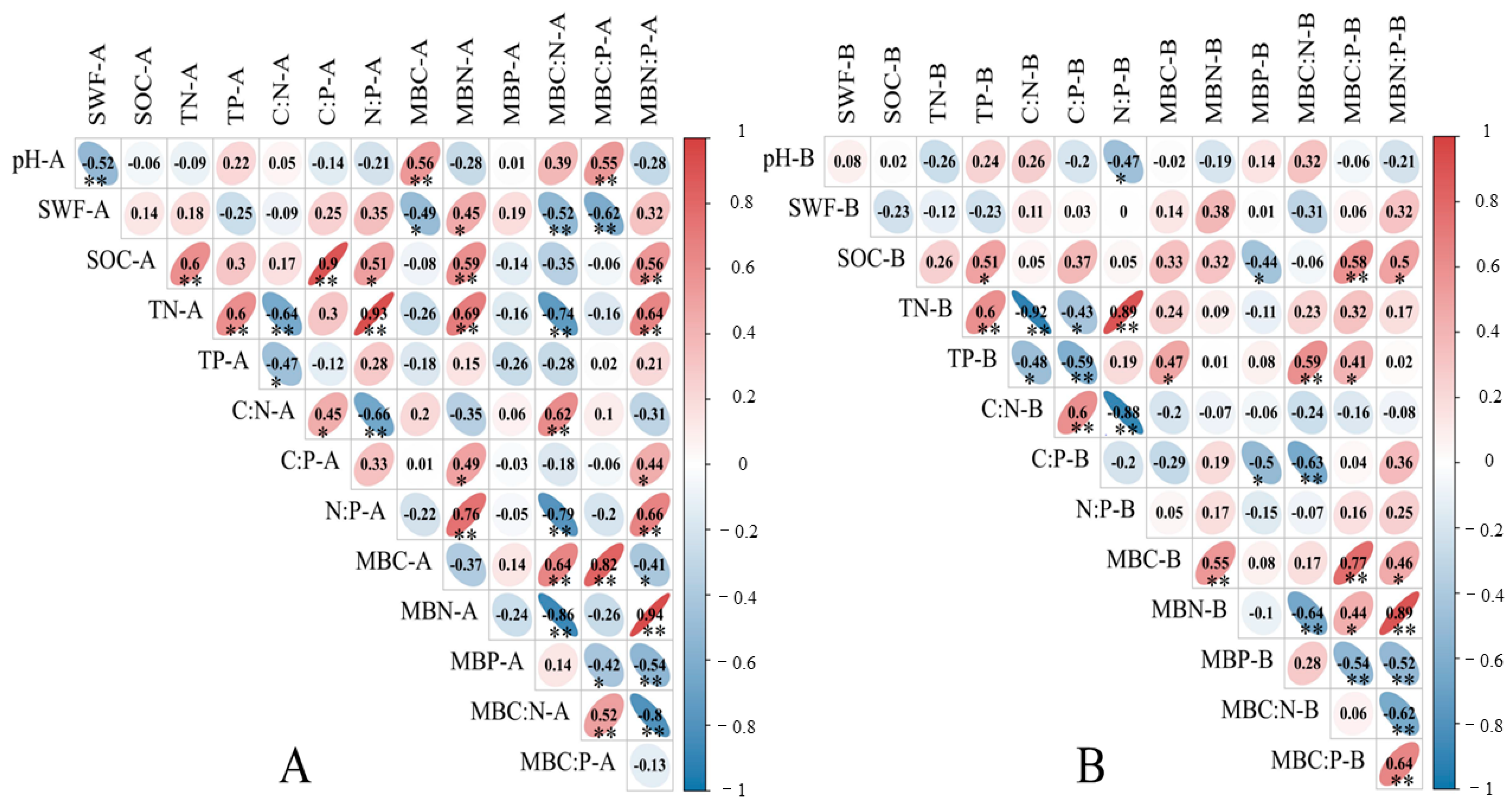
3.4. Correlation between Soil-Microbial Biomass Carbon, Nitrogen, Phosphorus and Stoichiometric Ratio
3.5. Redundancy Analysis
4. Discussion
4.1. Soil Carbon, Nitrogen, and Phosphorus of Different Forest Stand Types and Their Ecological Stoichiometry Characteristics
4.2. Soil Microbial Biomass Carbon, Nitrogen, and Phosphorus Contents in Different Forest Stand Types and Their Ecological Stoichiometry Characteristics
4.3. Soil-Microorganism Carbon, Nitrogen, Phosphorus Relationship
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, Y.; Wang, J.; Dang, X.; Han, Y.; Gao, Y.; Duan, X.; Jin, S. Soil Carbon, Nitrogen and Phosphorus Densities and Ecological Stoichiometry Characteristics of Haloxylon ammodendron Plantations in Arid Desert Area. J. Soil Water Conserv. 2022, 36, 259–266. (In Chinese) [Google Scholar]
- Zhang, Z.M.; Huang, X.F.; Zhou, Y.C. Factors influencing the evolution of human-driven rocky desertification in karst areas. Land Degrad. Dev. 2021, 32, 817–829. [Google Scholar] [CrossRef]
- Bertrand, I.; Viaud, V.; Daufresne, T.; Pellerin, S.; Recous, S. Stoichiometry constraints challenge the potential of agroecological practices for the soil C storage. A review. Agron. Sustain. Dev. 2019, 6, 54. [Google Scholar] [CrossRef]
- Liu, H.; Qi, J.C.; Liu, D.Q.; Yang, J.W.; Chen, M.W.; Li, S.P.; Li, C.J.; Li, C.Z. Effects of Different Forest Types On Stoichiometric Characteristics of Carbon, Nitrogen, and Phosphorus in Tropical Soils, China. Sustainability 2024, 16, 480. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.L.; Qiu, Q.Y.; Zhou, Y.; You, W.B. Changes in Soil Properties and Enzyme Stoichiometry in Three Different Forest Types Changed to Tea Plantations. Forests 2023, 14, 2043. [Google Scholar] [CrossRef]
- Qin, J.; Liu, Y.; Bi, Q.K.; Chen, Z.Q.; Zhang, B.W. Response of Leaf and Soil C, N and P Stoichiometry in Different Pinus massoniana Forest Types to Slope Aspect in the Dabie Mountains Region of North Subtropical, China. Front. Environ. Sci. 2023, 11, 1148986. [Google Scholar] [CrossRef]
- Deng, X.; Zhu, L.; Song, X.C.; Tang, J.; Tan, Y.B.; Deng, N.N.; Zheng, W.; Cao, J.Z. Soil Ecological Stoichiometry Characteristics of Different Stand Types in Maoershan Nature Reserve. Soil Bull. 2022, 53, 366–373. (In Chinese) [Google Scholar]
- Liu, Y.Y.; Luo, W.M.; Mu, G.T.; Wu, X.L.; Su, S.C.; Zhang, Z.M. C:N:P stoichiometric characteristics of the soil-vegetation system of three rare tree species growing on Mount Fanjing in Southwest China. Glob. Ecol. Conserv. 2021, 32, e01893. [Google Scholar] [CrossRef]
- Fiore-Donno, A.M.; Human, Z.R.; Štursová, M.; Mundra, S.; Morgado, L.; Kauserud, H.; Baldrian, P.; Bonkowski, M. Soil compartments (bulk soil, litter, root and rhizosphere) as main drivers of soil protistan communities distribution in forests with different nitrogen deposition. Soil Biol. Biochem. 2022, 168, 108628. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Wu, X.L.; Zhang, J.C.; Huang, X.F. Distribution and migration characteristics of microplastics in farmland soils, surface water and sediments in Caohai Lake, southwestern plateau of China. J. Clean. Prod. 2022, 366, 132912. [Google Scholar] [CrossRef]
- Gong, Z.J.; Sheng, M.Y.; Zheng, X.J.; Zhang, Y.; Wang, L.J. Ecological Stoichiometry of C, N, P and Si of Karst Masson Pine Forests: Insights for the Forest Management in Southern China. Sci. Total Environ. 2024, 912, 169490. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, Y.; Cui, X. Response of soil C. N, and P ecological stoichiometry to specific forest types and altitude in cold-temperateforests. J. Cent. South Univ. For. Technol. 2022, 42, 141–147. (In Chinese) [Google Scholar]
- Yang, X.; Wang, P.; Gao, D.; Gao, N.; Li, L.; Yang, X.; Zhong, Q. Ecological stoichiometry of Form Quercus pannosa and Form Pinus squamata in the Yaoshan Nature Reserve, Yunnan. Acta Ecol. Sin. 2019, 39, 4021–4028. (In Chinese) [Google Scholar]
- Li, Y.Y.; Fu, F.W.; Li, J.R.; Chen, W.S.; Ding, H.H.; Xiao, S.Y. Stoichiometric Characteristics of Abies georgei var smithii Plants in Southeast Tibet. Sustainability 2023, 15, 8458. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C. Responses and regulation mechanisms of microbial decomposers to substrate carbon, nitrogen, and phosphorus stoichiometry. Chin. J. Plant Ecol. 2016, 40, 620–630. [Google Scholar]
- Heuck, C.; Weig, A.; Spohn, M. Soil microbial biomass C:N:P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef]
- Fan, H.B.; Wu, J.P.; Liu, W.F.; Yuan, Y.H.; Hu, L.; Cai, Q.K. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Li, W.N.; Ali, I.; Han, X.M.; Ullah, S.; Yang, M. Soil C, N, P, K and Enzymes Stoichiometry of an Endangered Tree Species, Parashorea chinensis of Different Stand Ages Unveiled Soil Nutrient Limitation Factors. Forests 2023, 14, 624. [Google Scholar] [CrossRef]
- Luo, H.Q.; Yu, J.L.; Li, R.X.; Gu, J.D.; Luo, L.; Zhang, Y.Y.; He, Y.; Xiao, Y.L.; Deng, S.H.; Zhang, Y.Z.; et al. Microbial biomass C:N:P as a better indicator than soil and ecoenzymatic C: N:P for microbial nutrient limitation and C dynamics in Zoige Plateau peatland soils. Int. Biodeterior. Biodegrad. 2022, 175, 105492. [Google Scholar] [CrossRef]
- Chen, X.; Feng, J.G.; Ding, Z.J.; Tang, M.; Zhu, B. Changes in soil total, microbial and enzymatic C-N-P contents and stoichiometry with depth and latitude in forest ecosystems. Sci. Total Environ. 2022, 816, 151583. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.D.; He, M.; Jiang, C.L.; Liu, F. Soil microbial stoichiometry and community structure responses to long-term natural forest conversion to plantations in a subtropical region. Environ. Sci. Pollut. R. 2022, 29, 27560–27570. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.D.; Feng, F.J.; Lu, C.; Fu, Y.M. C:N:P stoichiometry of different soil components after the transition of temperate primary coniferous and broad-leaved mixed forests to secondary forests. Soil Tillage Res. 2022, 216, 105260. [Google Scholar] [CrossRef]
- Zhou, Z. Research on the Fanjing Mountain; Guizhou People’s Publishing House: Guiyang, China, 1990. [Google Scholar]
- Han, W.; Luo, L.; Zhang, S. Adsorption of tetrabromobisphenol A on soils: Contribution of soil components and influence of soil properties. Colloids Surf. A Physicochem. Eng. Asp. 2013, 428, 60–64. [Google Scholar] [CrossRef]
- Jing, Z.; Cheng, J.; Su, J.; Bai, Y.; Jin, J. Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland. Ecol. Eng. 2014, 64, 171–178. [Google Scholar] [CrossRef]
- Tang, L.L.; Wan, K.Y.; Cheng, C.P.; Li, R.H.; Wang, D.Z.; Pan, J.F.; Tao, Y.; Xie, J.; Chen, F. Effect of fertilization patterns on the assemblage of weed communities in an upland winter wheat field. J. Plant Ecol. 2014, 7, 39–50. [Google Scholar] [CrossRef]
- Yin, S.; Wang, C.; Jin, Y.; Zhou, Z. Changes in soil-microbe-exoenzyme C:N:P stoichiometry along an altitudinal gradient in Mt. Datudingzi, Northeast China. Chin. J. Plant Ecol. 2019, 43, 999–1009. (In Chinese) [Google Scholar] [CrossRef]
- Chen, F.; Liu, F.; Bai, X.; Wu, L.; Chen, Z. Spatial heterogeneity and ecological stoichiometry characteristics of soil carbon, nitrogen and phosphorus under different micro-geomorphology in karst mountains. Acta Ecol. Sin. 2022, 42, 10201–10213. (In Chinese) [Google Scholar]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Black, C.; Goring, C. Organic Phosohorus in Soils. Agronomy 1953, 1, 123–152. [Google Scholar]
- Mazzarino, M.J.; Szott, L.; Jimenez, M. Dynamics of soil total C and total N microbial biomass, and water-soluble C in tropical agroecosystems. Soil Biol. Biochem. 1993, 25, 205–214. [Google Scholar] [CrossRef]
- Makino, W.; Cotner, J.B.; Sterner, R.W.; Elser, J.J. Are Bacteria More Like Plants or Animals? Growth Rate and Resource Dependence of Bacterial C:N:P Stoichiometry. Funct. Ecol. 2003, 17, 121–130. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Xue, H.L.; Lan, X.; Liang, H.G.; Zhang, Q. Characteristics and Environmental Factors of Stoichiometric Homeostasis of Soil Microbial Biomass Carbon, Nitrogen and Phosphorus in China. Sustainability 2019, 11, 2804. [Google Scholar] [CrossRef]
- Xiang, Y.; Chen, S.; Pan, P.; Ouyang, X.; Ning, J.; Li, Q. Stoichiometric traits of C, N and P of leaf-litter-soil system of Pinus massoniana forest. J. For. Environ. 2019, 39, 120–126. (In Chinese) [Google Scholar]
- Beugnon, R.; Bu, W.S.; Bruelheide, H.; Davrinche, A.; Du, J.; Haider, S.; Kunz, M.; Oheimb, G.V.; Garcia, M.D.P.; Saadani, M.; et al. Abiotic and Biotic Drivers of Tree Trait Effects On Soil Microbial Biomass and Soil Carbon Concentration. Ecol. Monogr. 2023, 93, e1563. [Google Scholar] [CrossRef]
- Kumar, U.; Kaviraj, M.; Panneerselvam, P.; Nayak, A.K. Conversion of Mangroves into Rice Cultivation Alters Functional Soil Microbial Community in Sub-humid Tropical Paddy Soil. Front. Environ. Sci. 2022, 10, 858028. [Google Scholar] [CrossRef]
- Li, S.; Cui, Y.; Sun, Z.; Liu, H.; Ye, H. Effect of grazing exclusion on soil organic carbon and stoichiometry characteristics of soil microbiall biomass in sagebrush desert. Acta Prataculturae Sin. 2023, 32, 58–70. (In Chinese) [Google Scholar]
- Feng, E.P.; Zhang, L.W.; Kong, Y.H.; Xu, X.K.; Wang, T.; Wang, C.F. Distribution Characteristics of Active Soil Substances along Elevation Gradients in the Southern of Taihang Mountain, China. Forests 2023, 14, 370. [Google Scholar] [CrossRef]
- Ren, C.J.; Zhao, F.Z.; Kang, D.; Yang, G.H.; Han, X.H.; Tong, X.G.; Feng, Y.Z.; Ren, G.X. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. Forest Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sample Size | Altitude | Slope Aspect | Soil Type | Constructive Species |
|---|---|---|---|---|---|
| I | 30 m × 30 m (n = 3) | 634–637 | E86° | Cab Udi Orthic Entisols | Castanopsis fargesii; Castanopsis carlesii (Hemsl.)Hay. |
| 30 m × 30 m (n = 3) | 812–816 | SE125° | Cab Udi Orthic Entisols | Castanopsis fargesii | |
| II | 30 m × 30 m (n = 3) | 1652–1654 | N340° | Cab Udi Orthic Entisols | Cyclobalanopsis multinervis W. C. Cheng et T. Hong; Quercus engleriana Seem. |
| 30 m × 30 m (n = 3) | 1817–1824 | NW303° | Cab Udi Orthic Entisols | Cyclobalanopsis multinervis W. C. Cheng et T. Hong; Betula austrosinensis Chun ex P. C. Li | |
| III | 30 m × 30 m (n = 3) | 1804–1807 | SW221° | Cab Udi Orthic Entisols | Acer sinense Pax |
| 30 m × 30 m (n = 3) | 2061–2063 | E111° | Fec Hydragric Anthrosols | Acer sinense; Pax Cyclobalanopsis multinervis W. C. Cheng et T. Hong | |
| IV | 30 m × 30 m (n = 3) | 2336–2339 | NW312° | Fec Hydragric Anthrosols | Rhododendron argyrophyllum Franch. |
| 30 m × 30 m (n = 3) | 2350–2352 | W266° | Fec Hydragric Anthrosols | Rhododendron leishanicum Fang et X. S. Zhang ex Chamb. |
| Forest Type | Soillayer | Forest Type * Soillayer | ||
|---|---|---|---|---|
| Soil properties | SOC | 5.204 ** | 44.135 ** | 5.387 ** |
| TN | 5.816 ** | 31.429 ** | 1.435 | |
| TP | 20.421 ** | 40.09 ** | 0.944 | |
| C:N | 26.503 ** | 2.251 | 1.09 | |
| C:P | 11.043 ** | 22.919 ** | 4.795 ** | |
| N:P | 9.877 ** | 20.951 ** | 1.976 | |
| WAF | 3.019 * | 0.673 | 4.74 * | |
| ph | 22.593 ** | 22.151 ** | 1.916 | |
| Microbial biomass | MBC | 3.12 * | 3.206 | 0.427 |
| MBN | 0.531 | 2.066 | 2.13 | |
| MBP | 0.844 | 0.835 | 0.625 | |
| MBC:MBN | 1.489 | 8.306 | 0.251 | |
| MBC:MBP | 3.743 * | 3.186 | 0.231 | |
| MBP:MBN | 0.56 | 0.954 | 2.092 |
| Item | Soil Layer | Axis 1 | Axis 2 | Axis 3 | Axis 4 | Total Variance |
|---|---|---|---|---|---|---|
| Eigenvalues | A | 0.408 | 0.173 | 0.071 | 0.019 | 1 |
| B | 0.375 | 0.144 | 0.076 | 0.005 | 1 | |
| Species–environment correlations | A | 0.889 | 0.797 | 0.775 | 0.620 | |
| B | 0.909 | 0.702 | 0.711 | 0.356 | ||
| CV of species data (%) | A | 40.8 | 58.1 | 65.2 | 67.2 | |
| B | 37.5 | 51.9 | 59.5 | 60.0 | ||
| CV of species–environment relationship (%) | A | 60.6 | 86.3 | 96.9 | 99.7 | |
| B | 62.1 | 86.0 | 98.6 | 99.4 | ||
| Sum of all eigenvalues | A/B | 1 | ||||
| Variance explained by all variables | A | 0.69 | ||||
| B | 0.62 | |||||
| Variance explained by selected variables | A | 0.66 | ||||
| B | 0.60 | |||||
| Forward selection of variables | RDA 1 | RDA 2 | F value | p value | Extra fit | |
| Aspect | A | −0.1644 | −0.6130 | 3.65 | 0.022 * | 0.14 |
| B | −0.3547 | 0.1657 | 5.31 | 0.004 ** | 0.14 | |
| Altitude | A | 0.1800 | −0.1081 | 4.20 | 0.029 * | 0.14 |
| B | 0.2640 | 0.0017 | 4.71 | 0.011 * | 0.15 | |
| SWF | A | −0.4283 | 0.1617 | 11.63 | 0.001 ** | 0.26 |
| B | 0.0466 | 0.3425 | 3.10 | 0.041 * | 0.08 | |
| Slope | A | −0.3397 | 0.4885 | 3.76 | 0.003 ** | 0.07 |
| B | 0.5033 | 0.3664 | 4.19 | 0.007 ** | 0.16 | |
| Forest type | A | 0.0241 | −0.0111 | 2.80 | 0.024 * | 0.05 |
| pH | B | 0.2809 | −0.1311 | 3.17 | 0.039 * | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, W.; Wu, X.; Chen, S.; Mu, G.; Liu, Y. Soil-Microbial CNP Content and Ecological Stoichiometry Characteristics of Typical Broad-Leaved Tree Communities in Fanjing Mountain in Spring. Diversity 2024, 16, 249. https://doi.org/10.3390/d16040249
Luo W, Wu X, Chen S, Mu G, Liu Y. Soil-Microbial CNP Content and Ecological Stoichiometry Characteristics of Typical Broad-Leaved Tree Communities in Fanjing Mountain in Spring. Diversity. 2024; 16(4):249. https://doi.org/10.3390/d16040249
Chicago/Turabian StyleLuo, Wenmin, Xianliang Wu, Sheng Chen, Guiting Mu, and Yingying Liu. 2024. "Soil-Microbial CNP Content and Ecological Stoichiometry Characteristics of Typical Broad-Leaved Tree Communities in Fanjing Mountain in Spring" Diversity 16, no. 4: 249. https://doi.org/10.3390/d16040249