
Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax


1
Co-Innovation Center for Sustainable Forestry in Southern China, College of Ecology and the Environment, Nanjing Forestry University, Nanjing 210037, China
2
Laboratory of Animal Behaviour and Conservation, College of Life Sciences, Nanjing Forestry University, Nanjing 210037, China
3
Amphibian Specialist Group, Species Survival Commission, International Union for the Conservation of Species, Toronto, ON M8W 1P5, Canada
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(5), 259; https://doi.org/10.3390/d16050259
Submission received: 15 March 2024
/
Revised: 12 April 2024
/
Accepted: 15 April 2024
/
Published: 25 April 2024
(This article belongs to the Special Issue Ectotherms in a Dynamic Environment: Understanding Patterns of Ecology, Distribution, Evolution, and Threats to Species)
Abstract
:Understanding species from an ecological and phylogenetic perspective facilitates an understanding of their conservation status in relation to the changing world. The frog genus Pelophylax is among the largest in terms of amphibian biomass in the Palearctic, but species have not been thoroughly studied at the Asian continental scale. The phylogeographic relationship, behavioural ecology, and ecological requirements within the genus need clarification, despite generally good local coverage. Here, for the first time, we conducted a literature review focused on phylogeography and behavioural ecology, supported by ecological niche modelling of twelve Asian Pelophylax lineages. Finally, we compiled the known threats for each of the lineages. We first determined the presence of twelve species and species-candidate lineages. In terms of behavioural ecology, the main difference among lineages is the time to metamorphosis and the time to reach sexual maturity. The ecological models highlighted a match between the known presence of each clade and their suitable habitat and highlighted the Syr Darya drainage on the eastern shore of the Caspian Sea for the non-described Pelophylax “Syr Darya lineage”. Finally, we highlighted the greatest variation among lineages in terms of threats, as some lineages are threatened by numerous factors, whereas others are expanding.
1. Introduction
Delineating the boundaries of species distribution is a preliminary requisite to the determination of presence, phylogenetic patterns, behavioural ecology, and the threat levels that may require conservation actions [1,2]. Habitat suitability models are broadly used to determine suitable habitats for species based on environmental variables [3]. Thus, the study of climatic variables can shed light on the climatic tolerance of a species, thereby helping to understand the limits of its spatial distribution [4]. In turn, these results may facilitate fieldwork by predicting suitable habitats and potential distribution [5]. Such models are particularly useful for species that do not disperse over long distances, such as amphibians, which generally have continuous distribution patterns [6]. To determine ranges, presence points can be used to build ecological niche models and determine the suitable habitat for a species [7,8]. Such an integrated analysis is especially valuable for genera with widespread distributions, as surveying the whole range of the species is not always possible. Here, we follow this modelling approach for the widespread frog genus Pelophylax, focusing on all Asian lineages.
The Pelophylax genus is a widespread group of water frogs that has been intensively studied in specific localities since Carl von Linné described the first species in the genus: Pelophylax esculentus (Linnaeus, 1758). Currently, there are 22 species and three hybrid asexual forms in the genus, which are distributed throughout the Palaearctic [9]. The data for Pelophylax are characterised by in-depth studies on both extreme longitudes of the Palearctic [10,11], but not as much information is available for the dry landscapes of central Asia. The genus Pelophylax is generally present in mixed and deciduous forests, inhabiting a wide variety of flowing and stagnant water habitats from shallow puddles and streams to large lakes and rivers [12]. In general, the species of this genus prefer open and warm areas with abundant herbaceous vegetation [13].
General information on the distribution and occurrence of species has become increasingly accessible thanks to molecular barcoding and studies on the cryptic morphological characters of the genus [14]. In addition, knowledge is also available from combined regional and national distribution accounts through citizen science occurrence databases (e.g., iNaturalist) and other data collection platforms (e.g., GBIF, AmphibiaChina). This exciting momentum provides an opportune framework for re-examination of the Pelophylax lineages, focusing on Asia.
Here, our goal is to enhance the understanding of species richness patterns and refine biogeographic boundaries by cohesively delineating all lineages of Pelophylax occurring in Asia. We do so by reviewing the available phylogeographic literature and illustrating the relationships using models that integrate the ecological requirements of each lineage. Finally, we identify the threats to these lineages.
2. Materials and Methods
The Pelophylax genus is widespread across most landscapes in Asia (Figure 1), as the species have generally broad ecological requirements and there is at least one species present in each type of wetland [15]. The distribution of the species in the genus is likely regulated through niche segregation and habitat use between species complexes [16,17,18,19]. Recently, population expansions were reported outward of all range boundaries, following human-driven range extensions, such as for P. nigromaculatus [20,21] and P. ridibundus [22].
For each species, our review first follows logical assessments for phylogeographic and phylogenetic updates, followed by summaries of the species’ behavioural ecology. Next, we address ecological boundaries using environmental niche models. We modelled the habitat suitability of the following 12 Pelophylax species: P. plancyi, P. porosus, P. chosenicus, P. hubeiensis, P. fukienensis, P. mongolius, P. ridibundus, P. bedriagae, P. terentievi, and three candidate species, including the Pelophylax “Syr Darya” lineage in central Asia [23], and we follow the candidate species taxonomy segregation for P. nigromaculatus “reinhardtii” and P. nigromaculatus ”nigromaculatus” [12].
We also reviewed the literature for threat assessments for all of these lineages, based on the criteria of The IUCN Red List of Threatened Species [12]. To collect the data needed for each of the four lines of analysis, we searched the literature using the name of each species listed above independently with quotation marks as a keyword (e.g., “Pelophylax xxx”), where xxx represents the name of the species or lineage. We used the global citation database Google Scholar on 12 December 2023 for the search (Figure 2).
For modelling, we downloaded the occurrence data derived from the Global Biodiversity Information Facility database (GBIF.org: DOI: 10.15468/dl.8af3yw; 10.15468/dl.mjstaa; 10.15468/dl.5u7ky4; 10.15468/dl. wsee82; 10.15468/dl.w63w3q; 10.15468/dl.4qbf6e; 10.15468/dl. hcf9pa; 10.15468/dl. dpu5j7; 10.15468/dl.cu533d; 10.15468/dl.qg4sky) and from iNaturalist (all data are compiled in Supplementary Material Table S1). Based on these data, we built niche predictions using the 19 bioclimatic variables extracted from WorldClim v.1.4. [24] and an altitude layer. We used layers with a spatial resolution of 30 arc seconds under WGS 84 projection with a species-specific mask covering the area of occurrence of the focal Pelophylax lineage in ArcGIS 10.6 (ESRI, 2012). To avoid correlation between climatic variables and a loss in the prediction power owing to over-parametrisation, we spatially thinned all occurrence records at a distance of 10 km using Wallace v. 2.0.6 [25,26,27]. Our habitat suitability models used the following numbers of records for each species after adjustment and thinning: P. ridibundus (n = 5720); P. bedriagae (n = 1402); P. terentievi (n = 265); Pelophylax “Syr Darya lineage” (n = 202); P. nigromaculatus “nigromaculatus” (n = 381); P. nigromaculatus “reinhardtii”: (n = 2783); P. porosus (n = 67); P. mongolius (n = 42); P. plancyi (n = 82); P. chosenicus (n = 270); P. hubeiensis (n = 40); and P. fukienensis (n = 2923). We used the maximum training sensitivity plus specificity threshold (MTSS) generated by Maxent for each species as the threshold for moderate suitability [28]. Before generating the models, we calculated Pearson’s correlation coefficient for all 19 bioclimatic variables.
We used the GRASS plug-in to calculate Pearson’s correlation coefficients [26] to eliminate predictor collinearity in ArcGIS 10.6 (ESRI, 2012). For all pairs with high correlation factors (|r| > 0.8), we excluded the variable that was the least biologically important for Pelophylax lineages, as determined by the literature [29]. The resulting dataset contained 13 bioclimatic variables (Table 1): Elevation, Bio 1 (annual mean temperature), Bio 2 (mean diurnal temperature range), Bio 3 (isothermality), Bio 5 (maximum temperature of the warmest month), Bio 8 (mean temperature of the wettest quarter), Bio 9 (mean temperature of the driest quarter), Bio 10 (mean temperature of the warmest quarter), Bio 11 (mean temperature of the coldest quarter), Bio 12 (annual precipitation), Bio 14 (precipitation of the driest month), Bio 15 (precipitation seasonality), Bio 18 (precipitation of the warmest quarter), and Bio 19 (precipitation of the coldest quarter).
Using the reduced sets of environmental and occurrence data, we used MaxEnt v.3.4.4 [30]. For each lineage, we implemented MaxEnt using random seed, auto features, the default regularisation multiplier (=1), 10,000 random background points within a 20-km radius buffer around each occurrence point, a test data percentage of 20, a maximum iteration of 500, and the applied Minimum Training Presence threshold rule [31]. We also used a jackknife analysis to estimate the relative contributions of each variable (Table 1). The model performance was evaluated using the Area Under the Curve (AUC) [32].
3. Results
From the surveyed articles, we selected 72 articles only as only these provided relevant information for the 12 described Asian Pelophylax lineages. Using these references, we reviewed the information pertaining to the phylogeography, behavioural ecology, and conservation of our focal lineages.
All 12 models had moderate or high AUC values, which is considered an excellent fit [33]. The ecological models produced moderately to highly valid results, indicating segregated suitable habitats and the probability of occurrence for all lineages. The AUC values for each were as follows: Pelophylax ridibundus: 0.992; P. bedriagae: 0.985; P. terentievi: 0.994; Pelophylax “Syr Darya” lineage: 0.976; P. nigromaculatus “nigromaculatus”: 0.895; P. nigromaculatus “reinhardtii”: 0.953; P. porosus: 0.997; P. mongolius: 0.980; P. plancyi: 0.990; P. chosenicus: 0.997; P. hubeiensis: 0.981; and P. fukienensis: 0.997. All models generally fitted the known distribution of the lineages they represented when compared with the data available from the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
3.1. Pelophylax nigromaculatus “Nigromaculatus”
3.1.1. Distribution
All Pelophylax nigromaculatus populations are currently assigned to a single species, but two diverging clades are identified as candidate species [12], and we follow this nomenclature. The boundary between the two clades is the sea between continental Asia and the Japanese archipelago [12]. Pelophylax nigromaculatus “nigromaculatus” (Hallowell, 1861/1860) was described from Shimoda, Honshu, Japan [34] (Figure 3). The species is widely distributed across Kyushu, Shikoku, and Honshu but not on the Kanto and Sendai plains, and it has been introduced to Hokkaido [35]. The lineage P. n. “nigromaculatus” is the common Pelophylax lineage in Japan [36].
3.1.2. Ecology
Despite occurring in a variety of habitats, the population of P. n. “nigromaculatus” in Japan uses rice paddies as their main reproductive sites [37]. Members of the lineage are not restricted to wetlands and leave from rice paddies and migrate to woodlands during the non-breeding season [35]. Females spawn a single egg clutch every year, and the breeding season is from April to June [35]. The P. n. “nigromaculatus” lineage is a generalist predator, but arthropods make up a large proportion of its diet [36,38]. The geographical distribution of P. n. “nigromaculatus” and P. porosus brevipodus (Ito, 1941) partially overlap [18]. In addition, competition between syntopic Pelophylax species that occupy similar niches tends to influence the growth rate of P. n. “nigromaculatus”; for instance, higher growth rates were recorded in P. n. “nigromaculatus” populations without syntopic P. p. brevipodus than in populations with P. p. brevipodus [39]. Males of the species attract females using leks [40]. For the P. n. “nigromaculatus” lineage, the highest habitat suitability matched their current range in Japan, and the variable with the highest contribution was precipitation of the coldest quarter (Table 1). As winters are typically dry in the region, these results highlight the selection of habitats where the risk of drying out during hibernation is lower.
3.1.3. Threats and Conservation
Pelophylax nigromaculatus was assessed as Least Concern by The IUCN Red List of Threatened Species in 2023, being down-listed from Near Threatened [41]. The lineage P. n. “nigromaculatus” was not assessed on its own, but it is also likely to reach the Least Concern status in view of the widespread distribution of the species, the relatively large populations, and its resilience to human presence. However, the habitat used by the species is declining, especially when rice paddies are urbanised [42].
3.2. Pelophylax nigromaculatus “Reinhardtii”
3.2.1. Distribution
Pelophylax nigromaculatus “reinhardtii” [43,44] was described from the People’s Republic of China (hereafter China; Figure 4). According to Peters, the two syntypes of this species were bought from a dealer, who had obtained them from either Malacca or China, and later restricted to China [34,44]. The name “reinhardtii” is linked to the populations on the Asian continent [12], and the P. n. “reinhardtii” lineage has a broad distribution across the eastern Palearctic, including regions from Khabarovsk Krai in far-eastern Russia to the west of the Sichuan Basin in China. The lineage also ranges across the Korean Peninsula and related islands [45,46,47]. The lineage has been introduced to new localities, both in areas within its range where it was not originally present and offshore islands, such as Ulleung Island in the Republic of Korea [21] (hereafter R Korea). It has been introduced in Baiyin, Ningxia, in Yining, Xinjiang, and in Lhasa, Tibet, through trade and the practice of mercy releases [20]. In addition, P. n. “reinhardtii” populations heavily hybridise with species of the P. plancyi species complex, namely, with P. chosenicus on the Korean Peninsula, and with P. plancyi, P. hubeiensis, and P. fukienensis within their respective ranges in Eastern China [45,48].
3.2.2. Ecology
Male P. n. “reinhardtii” start calling from April in southern regions of the lineage’s range in China and R Korea, early to mid-May in R Korea, and June in the Russian Federation (hereafter Russia), although the species can be heard until July in R Korea [49]. The onset of spawning for this species is prompted by high temperatures and increased relative humidity levels [50]. The P. n. “reinhardtii” lineage primarily eats insects, gastropods, and arachnids [51]. The lineage is found up to relatively high elevations of around 2200 m above sea level (hereafter a.s.l.) in China (noting a potential misidentification with P. mongolius; [13]) but only up to 1000 m a.s.l. in R Korea, where the peak abundance is around 150 m a.s.l. [52]. Females of P. n. “reinhardtii” lay a single egg clutch every year comprising about 1000 eggs. Metamorphosis starts about six weeks after the eggs have been laid, and hibernation lasts from October to May [12]. Sexual maturity is reached at the age of two years for males and three years for females, and maximum longevity is six years in males and seven years in females [53]. Frogs from high altitudes tend to be smaller than frogs from low altitudes [54]. Within the watershed of the Geum River in R Korea, all individuals are from a single genetic population, and the important morphological diversity is attributed to environmental gradients rather than genetic structure [55]. For P. n. “reinhardtii”, the annual mean temperature was the variable with the highest contribution (Table 1), which is a common pattern for species with large distributions.
3.2.3. Threats and Conservation
Pelophylax n. “reinhardtii” was not assessed by the IUCN Red List of Threatened Species, but P. nigromaculatus is listed as Least Concern, being down-listed from Near Threatened in 2023 [41]. The severity of threats to the lineage varies by region: in some areas, such as the agricultural wetlands of the Democratic People’s Republic of Korea (hereafter DPR Korea), the lineage remains stable owing to suitable habitat conditions [52]. In R Korea and China, the lineage is not considered threatened, owing to its large population sizes and ability to withstand minor environmental changes [49]. Populations restricted to agricultural wetlands are impacted by agrochemical pollution, in terms of both tadpole development and loss of fitness [56] and gonadal development and feminisation of the population [57]. Other noted threats are invasive fish species that prey on eggs [58] and adult Lithobates catesbeianus, which feeds on adults and juveniles [59]. The lineage is also heavily impacted by road mortality in R Korea [60].
3.3. Pelophylax porosus
3.3.1. Distribution
Pelophylax porosus, or the Daruma Pond Frog (Cope 1868), was described from Kanagawa prefecture, in Eastern Honshu, Japan [35] (Figure 5). The species comprises two subspecies, the Tokyo Daruma Pond Frog, P. p. porosus (Cope 1868), and the Nagoya Daruma Pond frog, P. p. brevipodus (Ito 1941). The Tokyo Daruma Pond frog ranges from the Kanto to the Sendai Plains in central and southern Niigata Prefecture and in northern and central Nagano Prefecture [35]. The Nagoya Daruma Pond frog was described from Nagoya, Japan, and it is distributed in the Western Honshu, Tokai, central Kinki, and San-yo districts and in northern Shikoku [18,45].
3.3.2. Ecology
Both P. p. porosus and P. p. brevipodus inhabit natural and artificial semi-aquatic environments such as rice paddies, ponds, and marshes in lowland areas, and they breed in still-water environments from late April to July [35]. Longevity for this species is estimated to be three to four years in both sexes, and they start breeding at two years old [39]. The subspecies P. p. porosus is the most short-lived and the first to reach sexual maturity of all Pelophylax lineages in Japan [39]. Pelophylax p. porosus males select vegetation-shaded water surfaces as daytime calling sites and open water surfaces as night calling sites [61]. The species has been recorded up to 180 m a.s.l. [62]. For P. porosus, precipitation of the coldest quarter was the only significant variable contributing to the model, indicating a preference for wetter winter conditions.
3.3.3. Threats and Conservation
Pelophylax porosus was assessed by The IUCN Red List of Threatened Species as Least Concern in 2020, despite its decreasing population size [63]. The P. porosus population is threatened by human activities, especially habitat destruction and degradation (including agricultural wetlands), and pollution. In addition, the species is threatened by hybridisation with P. n. “nigromaculatus” [45,64]. These threats are especially acute for P. p. brevipodus, which is declining in westernmost areas and may be locally extinct on some islands [65,66].
3.4. Pelophylax mongolius
3.4.1. Distribution
In 1927, Schmidt divided the common Chinese and Japanese pond frog into three subspecies based on the shape of the head, the length of the limbs, the length of the snout, and the nature of the metatarsal tubercle, describing the northern form as Rana nigromaculata mongolia [67]. Pelophylax mongolius, or the Yellow River Pond Frog, was described from Meidaizhao, Inner Mongolia, China (Figure 6), and elevated to species level in 2022 [68] based on a holotype initially classified as P. nigromaculatus mongolia (Schmidt, 1925) and later wrongly synonymised with Pelophylax tenggerensis [11]. This species ranges across Inner Mongolia, Ningxia, Shaanxi, and eastern Gansu in China, west of the Taihang Mountains, which serve as a geographic barrier with P. n. “reinhardtii” [12].
3.4.2. Ecology
Pelophylax mongolius is present from around 300 m a.s.l. and potentially up to 2200 m a.s.l. [13]. This species is adapted to relatively arid environments and occurs in disconnected subpopulations within suitable habitats, which include grasslands and forests adjacent to wetlands, such as marshes, pools, rivers, and streams [69]. Females lay a single egg mass every year, with about 1000 eggs in each egg clutch (although this number may be based on misidentifications). Breeding starts in early May and lasts until mid-July, metamorphosis peaks in late June, and the species starts hibernating in late October [12]. Elevation was also the variable with the higher contribution for P. mongolius (Table 1), which is consistent with the restriction of the species distribution to comparatively higher elevations.
3.4.3. Threats and Conservation
The species was elevated to species level in 2022 and is not formally assessed by the IUCN Red List of Threatened Species, but as some sub-populations were assessed under different epithets, these are considered non-threatened [12]. However, some threats are present and need attention. For instance, P. mongolius is under threat of hybridisation with P. n. “reinhardtii”. It is collected for human consumption and mercy releases, and it will face a shift in its suitable habitat because of climate change [70].
3.5. Pelophylax plancyi
3.5.1. Distribution
Pelophylax plancyi, or the Eastern Golden Pond Frog, was described from Jiujiang in Jiangxi in China [13] (Figure 7). The species distribution is centred around the low-elevation plains west of the Yellow Sea, where the population density is the highest. However, the species is distributed as far south as Zhejiang and Jiangxi (where the type locality is located; Lataste, 1880), to Hunan and Hubei to the west, and to Hebei [31] and possibly the vicinity of Beijing to the north, although the clade needs to be identified using molecular tools [31]. This species is closely related to P. chosenicus to the west and P. hubeiensis to the east. The segregation between P. plancyi and P fukienensis is more ancient than that relating to other closely related species [45], noticeable variations in call properties and morphology confirm the segregation from P. hubeiensis [71,72], and the split from P. chosenicus supports the species level of each clade. However, the unexpected distribution of P. chosenicus may have resulted in the misidentification of individuals in other studies [31].
3.5.2. Ecology
Pelophylax plancyi is distributed from sea level, occurring on reclaimed tidal flats in Jiangsu, and from 50 to 200 m a.s.l. in the western areas of its range [13]. The species emerges from hibernation between late March and early April in southern areas, and in May at the northern edge of its range until June [13]. Females lay about 1500 eggs and generally prey on pest insects in agricultural wetlands [13]. Metamorphosis can take up to eight weeks in the northern boundary of its range [12]. The hibernation period lasts from late October or November to April [13]. The species reaches sexual maturity within two to three years and does not migrate between wetlands and wooded areas for hibernation [12]. Pelophylax plancyi co-occurs with other sympatric Pelophylax species [73], and frequent instances of hybridisation have been reported [48]. In addition, genetic analyses suggest the occurrence of large numbers of hybrids in some populations [45]. The variables with the highest contributions to the distribution of P. plancyi were isothermality (34%) and precipitation of the driest month (23%; Table 1).
3.5.3. Threats and Conservation
Pelophylax plancyi was assessed as Least Concern by the IUCN Red List of Threatened Species in 2019 [74]. The main threat to the species is the loss of natural habitat as it is transformed, and although the species can adjust to substitute habitats in the form of rice paddies, populations in rice paddies are impacted by agrochemicals [75]. Climate change is also likely to impact the species’ distribution and abundance at the southern edges of its range [70]. In addition, habitat loss and fragmentation caused by urbanisation have significant negative effects on the population dynamics of P. plancyi [76], which is declining in abundance with increasing urbanisation [77,78]. In addition, the P. plancyi populations in eastern China are threatened by hybridisation with P. n. “reinhardtii” [45,46].
3.6. Pelophylax chosenicus
3.6.1. Distribution
Pelophylax chosenicus, the Korean Golden Pond Frog or Gold-Spotted Pond Frog, was described from Incheon in R Korea [79] (Figure 8). The species is distributed along the western coast of the Korean Peninsula and along the same coastline in Liaoning, China [12,31]. A few inland populations have also been identified in the southwest of R Korea and low-elevation plains around Shenyang in China [12,31,68]. The species may also be present on the northern bank of the Yellow Sea, potentially all the way to Beijing, but populations need to be tested using molecular tools for confirmation [31].
3.6.2. Ecology
Pelophylax chosenicus is found in small and shallow ponds at low elevations [17], generally at a maximum of 500 m a.s.l. in R Korea, with a mean elevation of about 200 m [80]. The breeding season of the species starts in May, with males calling from water bodies until late July if the water does not dry out [49]. Individuals of the species have a relatively small home range [81]. They do not migrate away from the wetlands for overwintering, and they can be active until mid-December on warm days, but they are the latest amphibians to emerge from hibernation in R Korea between late March and early April [49]. Females lay a single egg mass, and tadpole development is around six weeks long. Once metamorphosed, young individuals stay in the vicinity of the water body, and it takes about three years for individuals to reach sexual maturity [12]. Pelophylax chosenicus eats mainly small insects and insect larvae in water or at the edges of water bodies [82]. Frequent instances of hybridisation have been reported with the sympatric Pelophylax lineage [83]. The variables with the highest contribution to the distribution of P. chosenicus were isothermality (40%) and precipitation of the driest month (23%; Table 1).
3.6.3. Threats and Conservation
Pelophylax chosenicus was assessed as Vulnerable by The IUCN Red List of Threatened Species in 2020 [84]. The species was extirpated from some sites in R Korea, and it is declining in DPR Korea, including in North Pyongan and Seoncheon, because of droughts [52]. In R Korea and China, populations increasingly subsist in agricultural wetlands, where they are impacted by agrochemical pollution, resulting in the decrease or even local extirpation of populations [75]. The species cannot cope with extensive habitat changes besides agricultural wetlands [49], but it is only moderately affected by road kills [60]. Climate change models for suitable habitats for the species predict drastic decreases by 2030, 2050, and 2080 under different climate change scenarios [75]. However, conservation efforts are ongoing, with a translocation programme initiated by the National Institute of Ecology of Korea [85], and conservation plans and recommendations are available [86]. The range of this species overlaps with a few protected areas, although none are managed for wildlife, and the species is listed as Endangered category II by the Ministry of the Environment of R Korea. Finally, some individuals were found to be infected by the Chytrid fungus (Batrachochytrium dendrobatidis) [87], but the pathogen is not known to have a negative impact on the species.
3.7. Pelophylax hubeiensis
3.7.1. Distribution
Pelophylax hubeiensis, the Hubei Gold-Striped Pond Frog, was described from Lichuan in Hubei, China [88] (Figure 9). The species status of the lineage was discussed [11] and confirmed based on its morphology and phylogenetic relationship [72]. The discussion arose because of the cryptic morphological relationship with P. plancyi. For now, the species is known to be distributed in Hubei and Anhui [13], and it is present between 60 and 1070 m a.s.l. [13]. This is the Asian Pelophylax species with the smallest range.
3.7.2. Ecology
This species mainly inhabits rice paddies, lotus ponds, natural wetlands, permanent freshwater marshes, pools, and other water bodies [72]. Their diet mostly includes insects, arachnids, and small fish [13]. During the breeding season, which occurs from late April until July, female frogs spawn 1022 eggs on average and deposit them scattered in small clutches among the stems and leaves of aquatic plants [13]. Tadpoles are found in still water, often in vegetation at the bottom of the pond [88]. The variables with the highest contribution to the distribution of P. hubeiensis were annual mean temperature (28%), isothermality (25%), and precipitation of the driest month (22%; Table 1).
3.7.3. Threats and Conservation
3.8. Pelophylax fukienensis
3.8.1. Distribution
Pelophylax fukienensis, the Fukien Gold-Striped Pond Frog was described from Fuzhou in Fujian, China [90] (Figure 10). This species is only known from Zhejiang, Jiangxi, Fujian, and Taiwan, and it is present up to 1200 m a.s.l. [13,91]. Despite being distributed across a broad area, the species is not continuously distributed due to the presence of large mountains in mainland China.
3.8.2. Ecology
This species inhabits marshes, rice paddies, and ponds with vegetation [92]. Adults frequently produce advertisement calls while on emerged vegetation and hide in submerged vegetation when disturbed. Adults mainly feed on insects, earthworms, and very small crabs [13]. The species breeds in still water, and the breeding season is between April and June [13]. Females can lay 1048 eggs and may spawn twice a year [13]. Pelophylax fukienensis was initially considered a subspecies or synonym of P. plancyi [13]; however, the validity of the species was confirmed based on molecular data from mainland and island populations [53,93]. Elevation was the variable with the highest contribution for P. fukienensis, with a contribution of 60% (Table 1), matching the distribution of the species, as it is present at comparatively high elevations.
3.8.3. Threats and Conservation
3.9. Pelophylax ridibundus
3.9.1. Distribution
Pelophylax ridibundus, or the Marsh Frog, was described from Guryev in Kazakhstan (Pallas, 1771; (Figure 11). This species occurs over a vast area in Eurasia, from the United Kingdom in the west to the Kamchatka peninsula in the east, and from the outskirts of St. Petersburg (Russia) in the north to Saudi Arabia in the south [22,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111]. In mainland Europe, P. ridibundus was repeatedly introduced to Italy [112,113], France [114], Switzerland [115], Belgium [116,117], and Russia [22,118] (Figure 11).
3.9.2. Ecology
Pelophylax ridibundus is found in various biotopes with abundant herbaceous vegetation, including forests and bushland meadows. Breeding can start in mid-January and last until June in the mild climatic conditions of Türkiye [104], whereas, the breeding season starts in April in Iran [119] and Bulgaria [120] and in early May in Kazakhstan [108]. The diet of Pelophylax ridibundus includes terrestrial invertebrates belonging mostly to arthropod groups, and the most frequently consumed organisms are Curculionidae, Carabidae, Aranei, and Muscidae [121]. The egg masses are sticky and placed on the substrate [122]. The number of eggs spawned per year varies depending on the age of the female [123], and the mean fecundity was 3853 eggs (ranging from 940 to 6000) every year in a single egg clutch [104]. The species is present up to 1000 m a.s.l. [107]. The maximum lifespan of P. ridibundus varies with elevation; for instance, it is 13 years at high altitude (Sultansazlığı, Türkiye), whereas the longest lifespan recorded at low altitudes is eight years (Ulubağ, Türkiye). Similarly, individuals living at low altitudes generally reach sexual maturity at the end of their second or third year, whereas individuals living at high altitudes reach sexual maturity approximately one year later [124]. For P. ridibundus, precipitation of the driest month was the variable with the highest contribution (39%; Table 1), highlighting an ecological niche restricted by water availability.
3.9.3. Threats and Conservation
Pelophylax ridibundus was assessed as Least Concern by the IUCN Red List of Threatened Species in 2021 [125]. The species is one of the most invasive amphibians of Northern Eurasia [126], often hybridising with local clades, resulting in genetic dilution and population declines [127]. Pelophylax ridibundus can rely on agricultural landscapes as breeding habitats, and the species is highly tolerant to agricultural pollution [128,129].
3.10. Pelophylax bedriagae
3.10.1. Distribution
Pelophylax bedriagae (Camerano, 1882), the Levent Water Frog, was described from Dimashq in the current Syrian Arab Republic [34] (Figure 12). This species is known from Egypt, Israel, Türkiye, some Greek Islands, Cyprus, western Syria, Lebanon, Jordan, and western and southwestern Iran [130,131,132,133,134,135,136,137,138,139,140].
3.10.2. Ecology
Pelophylax bedriagae is a largely aquatic species that is present in permanent wetlands with rich aquatic vegetation, and it relies on permanent ponds for breeding [135,141]. The breeding period starts in April and lasts until July [138]. Pelophylax bedriagae completes metamorphosis after 111 days in Iran [138] and mainly consumes insects [142]. The species occurs at elevations up to 2500 m a.s.l. [138]. The mean temperature of the driest quarter (27%) and the mean temperature of the wettest quarter (17%) were the variables with the highest contributions among the parameters included in the P. bedriagae model (Table 1).
3.10.3. Threats and Conservation
Pelophylax bedriagae was assessed as Least Concern by the IUCN Red List of Threatened Species in 2021 [143]. However, populations are declining in the western Aegen Region of Türkiye [144]. In addition, tadpoles are parasitised by anchor worms (Lernaea cyprinacea), which has an adverse effect on the health of populations in Türkiye [144,145]. However, P. bedriagae spread outside its native range by means of introduction (e.g., for aquaculture), thereby threatening the genetic integrity of native Pelophylax clades [134], and hybridisation with P. cypriensis occurs.
3.11. Pelophylax terentievi
3.11.1. Distribution
Pelophylax terentievi (Mezhzherin 1992), the Terentiev’s Frog or Central Asian Pond Frog, was described from Obigarm Roghun district in Tajikistan [146] (Figure 13). This species is distributed south and east of the Caspian Sea in the Islamic Republic of Iran, Turkmenistan, and Uzbekistan [139,147,148]. The presence of P. terentievi is present in northern and southern Uzbekistan (Amu Darya drainage), partially overlapping with a genetically distinct Pelophylax lineage [148]. Hybrids of P. terentievi and this genetically distinct lineage are present in central Uzbekistan [148].
3.11.2. Ecology
Pelophylax terentievi typically occurs in rice paddies, rivers, and ponds and emerges from hibernation around April [147]. This species reaches sexual maturity at the age of three or four years, and each female spawns between 138 and 3317 eggs [149]. The species occurs at elevations between 400 and 822 m a.s.l. [150]. The variable with the highest contribution to the ecological model for P. terentievi was the precipitation of the warmest quarter (37%; Table 1).
3.11.3. Threats and Conservation
Pelophylax terentievi was assessed as Least Concern by the IUCN Red List of Threatened Species in 2021, although the population is inferred to be decreasing owing to the decline in habitat quality [151]. In addition, the species is threatened by overharvesting, and by hybridisation with individuals from the P. “Syr Darya” lineage in the central region of Uzbekistan.
3.12. Pelophylax “Syr Darya” Lineage
3.12.1. Distribution
This distinct Pelophylax lineage was identified based on populations in northeastern and eastern regions of Uzbekistan, and it is named after the Syr Darya drainage basin that it inhabits [148] (Figure 14). This candidate species is also found in Kyrgyzstan, Kazakhstan, western Tajikistan, and Uzbekistan [103,148,151,152,153,154]. Hybrid individuals resulting from P. terentievi and the Syr Darya lineage have been identified from central Uzbekistan [148]. In addition, there are two clades within the Syr Darya lineage, based on mitochondrial DNA, one restricted to the Syr Darya drainage basin and one to the Balkhash Basin [103].
3.12.2. Ecology
The Pelophylax “Syr Darya” lineage is present in the Syr Darya river basin in the central Asian region between 45.34° and 36.56° N and 60.05° and 77.01° E [148]. The habitat varies significantly across the range of the species, and most of the habitat in the west and centre of the range is composed of arid flatlands with desert, semi-desert, and steppe habitats and associated climates, whereas the habitat in the eastern part of the range is composed of mountains with a steep climate gradient [148]. The variable with the highest contribution to the ecological model for the P. “Syr Darya” lineage was the precipitation of the coldest quarter (34%; Table 1).
3.12.3. Threats and Conservation
The lineage has not been assessed by the IUCN Red List of Threatened Species, as it is not formally described. However, hybridisation is already known to be a significant threat, as the lineage co-occurs with other Pelophylax lineages, and hybrid individuals were identified among numerous populations in eastern Kazakhstan as well as in Uzbekistan [103,148]. Moreover, the lineage faces threats from habitat degradation and loss. The decline in habitat suitability is attributed to anthropogenic activities that alter the natural landscape and water resources. Finally, climate change further accentuates these stressors, particularly in a region already prone to aridification and warming. Such environmental changes can lead to shifts in distribution, reduction in habitat quality, and ultimately impact the survival of the lineage.
4. Discussion
Despite taxonomic challenges surrounding some of the Pelophylax lineages in Asia [155], our review is the first to clarify the status of all currently known lineages at species level (either formally described or not) and provide the behavioural and ecological background required to further understand these species. It is important to note that we found differences in the relationships among the ecological factors that are important to the distribution of Asian Pelophylax lineages, and the primary climatic variables were different for almost all lineages. For example, isothermality, annual mean temperature, and elevation were regularly among the most significant variables in East Asia, but the most important climatic variables for central Asian Pelophylax were precipitation of the coldest quarter and precipitation of the warmest quarter. Our results can be seen in a positive light, as we define a broader range than previously known for Pelophylax lineages in Asia.
For the P. n. “nigromaculatus” lineage, the area with the highest habitat suitability matched their current range in Japan. Based on the percentage of contribution, precipitation of the coldest quarter was the variable with the highest contribution (Table 1), similar to the sympatric Japanese tree frog (Dryophytes japonicus) across the same area [42,156]. Precipitation of the coldest quarter was also a limiting factor for the distribution of this lineage. As the region is generally marked by dry winters, the high contribution of this variable highlights an adaptation to wetter winters and low variability in precipitation among seasons [157]. For the P. n. “reinhardtii” lineage, annual mean temperature was the most important variable, representing the temperature fluctuations in comparison with yearly variations [17]. Interestingly, this variable was also associated with the occurrence of the sympatric P. chosenicus [31]. For P. porosus, which is an endemic species to Japan, the precipitation of the coldest quarter was the only significant variable contributing to the model (Table 1), highlighting its importance for the occurrence of the species. For P. mongolius, elevation was the variable with the highest contribution, which is consistent with the species’ distribution, as it is present from around 300 m and likely up to 2200 m [12]. The area of highest suitability for P. plancyi was consistent with the current known range of the lineage, centred mainly on low-elevation plants west of the Yellow Sea, with the caveat of the potential misidentification of individuals in the Beijing area [31] and, in this case, the inclusion of points in the model that would not be representative of the lineage. Isothermality and precipitation of the driest month were also highly contributing variables, and this result is generally consistent with the requirements of the sister lineage P. chosenicus [31]. The most suitable habitat for P. chosenicus was on the Korean Peninsula and further north along the Eastern Coastal Yellow Sea, which is in agreement with the known distribution of the species [31]; however, the potential misidentification of individuals in the Beijing area [31] was again apparent, as the suitable habitat looped back north of the Yellow Sea and onto the western plains around Beijing and Shandong. The suitable habitat for P. hubeiensis was largely consistent with the central Yangtze Basin, an area that is notoriously hot in summer, in line with the highest annual mean temperature, which was the main variable contributing to the distribution of the species. The most suitable habitats for P. fukienensis were in Taiwan, Fujian, and Jiangxi [13]. The species is mainly distributed in hilly and mountainous areas [92], and elevation had the highest contribution in the models.
Most central Asian lineages had very broad ranges, generally also spreading across Europe, and likely resulting in numerous local adaptations. However, at the general range level, the variables of importance were typically not the same as those for Eastern Asian lineages. For instance, the distribution of P. ridibundus was principally regulated by the precipitation of the driest month. However, this consistency in ecological requirements resulted in large shifts in distribution over time; for instance, the ecological niche of P. ridibundus expanded during the late Quaternary. However, the species was restricted to glacial refugia in the northern Balkans on the northern coasts of the Black and Azov Seas and possibly in Western Europe, but post-glacial dispersal routes started from the refugia in the northeastern Balkans and the Black–Azov Seas regions [157]. The mean temperature of the driest quarter had the highest percentage of contribution for the sympatric P. bedriagae, and the range of the species followed the same variations. During the Last Glacial Maximum, the species was mainly distributed in the north of the Mediterranean region and the south of European Russia; however, in the Late Pleistocene, the suitable habitat of P. bedriagae began to decline in southern Russia [157]. The remaining lineages were generally restricted to specific river basins. P. terentievi was restricted to the Amu Darya and Kashka Darya rivers, whereas the P. “Syr Darya” lineage was present in the Syr Darya, Talas, Chu, and Ili River drainage basins. For these lineages, the precipitation of the warmest quarter was of primary importance for the distribution of P. terentievi, and this pattern regulates the distribution of other amphibians, such as Rana graeca in the Balkans [158]. In addition, precipitation of the coldest quarter was important for the P. “Syr Darya” lineage, similar to the variable principally regulating the range of P. kurtmuelleri in Europe [157].
Globally, amphibians show evidence of niche conservatism, specifically with respect to cold tolerance [159]. However, in the face of a warming climate, variables such as mean temperature, annual temperature, and mean precipitation of the warmest quarter are changing [160], and climate change is becoming one of the main drivers of threats to the extinction of amphibians [161]. Similarly, most of the focal clades are at least partially reliant on agricultural wetlands, which may not be able to sustain the species, as most species cannot adapt to the ongoing mechanisation of the landscapes [16,162,163,164] and long-term climate change [70,165]. The information provided in this review provides a clearer picture of the distribution of the Pelophylax genus in Asia and is consistent with the most recently published taxonomic revision of the genus [166], and if the boundaries of ranges and behavioural ecology are a bit more generous than originally thought, this does not reflect a genuine improvement in the status of the species, although this information will help determine the conservation needs of the species.
5. Conclusions
In conclusion, this study is the first to provide valuable insights into the phylogeographic relationships, behavioural ecology, and ecological requirements of the frog genus Pelophylax in Asia. Our findings reveal differences among the lineages in taxonomic divergence, behavioural ecology, and ecological requirements. In addition, the ecological niche models successfully identify suitable habitats for each lineage, emphasising the importance of the Syr Darya drainage area on the eastern shore of the Caspian Sea for the non-described Pelophylax “Syr Darya” lineage.
Furthermore, this study highlights the significant variations among Pelophylax lineages in terms of the threats they face. Although some lineages are expanding, others are threatened by numerous factors. Clearly, a comprehensive understanding of the ecological and phylogenetic aspects of species is crucial for assessing their conservation status, especially in the currently changing environment. The findings from this study provide a foundation for further research and conservation efforts focused on the protection and management of Pelophylax species in Asia.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d16050259/s1, Table S1 contains the GPS coordinates for all the data points used in this study. All data are open-access but combined into a single file here.
Author Contributions
Conceptualisation, A.B. and H.A.; methodology, A.B. and H.A.; software, H.A.; validation, A.B.; formal analysis, H.A. and A.B.; investigation, A.B. and H.A.; resources, A.B.; data curation, A.B. and H.A.; writing—original draft preparation, H.A. and A.B.; writing—review and editing, A.B. and H.A.; visualisation, A.B. and H.A.; supervision, A.B.; project administration, A.B.; funding acquisition, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Foreign Youth Talent Program of the Ministry of Science and Technology of the People’s Republic of China (QN2023014004L) to A.B.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
The authors are thankful to Honghua Ruan for their support and Thomas Reed, Ingrid Langer, and Martin Pickersgill for the miniatures of some of the species included in the figures.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Manne, L.; Pimm, S. Beyond eight forms of rarity: Which species are threatened and which will be next? Anim. Conserv. 2001, 4, 221–229. [Google Scholar] [CrossRef]
- Richardson, D.; Whittaker, R. Conservation biogeography—Foundations, concepts and challenges. Divers. Distrib. 2010, 16, 313–320. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.; Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Wiens, J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 2004, 58, 193–197. [Google Scholar]
- Alvarado-Serrano, D.; Knowles, L. Ecological niche models in phylogeographic studies: Applications, advances and precautions. Mol. Ecol. Resour. 2013, 14, 233–248. [Google Scholar] [CrossRef]
- Duellman, W.E. Patterns of Distribution of Amphibians: A Global Perspective; Johns Hopkins University Press: Baltimore, MD, USA, 1999; pp. 1–30. [Google Scholar]
- Fletcher, R.; Hefley, T.; Robertson, E.; Zuckerberg, B.; Mccleery, R.; Dorazio, R. A practical guide for combining data to model species distributions. Ecology 2019, 100, e02710. [Google Scholar] [CrossRef]
- Araújo, M.; Anderson, R.; Barbosa, A.M.; Beale, C.; Dormann, C.; Early, R.; Garcia, R.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef]
- Frost, D.; Grant, T.; Faivovich, J.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; Sá, R.; Channing, A.; Wilkinson, M.; Donnellan, S.; et al. The amphibian tree of life. Bull. Am. Mus. Nat. Hist. 2006, 297, 1–291. [Google Scholar] [CrossRef]
- Fu, J.; Wen, L. Impacts of quaternary glaciation, geological history and geography on animal species history in continental East Asia: A phylogeographic review. Mol. Ecol. 2023, 32, 4497–4514. [Google Scholar] [CrossRef]
- Dufresnes, C.; Litvinchuk, S.N. Diversity, distribution and molecular species delimitation in frogs and toads from the Eastern Palaearctic. Zool. J. Linn. Soc. 2022, 195, 695–760. [Google Scholar] [CrossRef]
- Borzée, A. Continental Northeast Asian Amphibians: Origins, Behavioural Ecology, and Conservation; Academic Press: Amsterdam, The Netherlands, 2024. [Google Scholar]
- Fei, L.; Ye, C.; Jiang, J. Colored Atlas of Chinese Amphibians and Their Distributions; Sichuan Science and Technology Press: Chengdu, China, 2012. [Google Scholar]
- Plötner, J.; Baier, F.; Akin, C.; Mazepa, G.; Schreiber, R.; Beerli, P.; Litvinchuk, S.N.; Bilgin, C.C.; Borkin, L.; Uzzell, T. Genetic data reveal that water frogs of Cyprus (Genus Pelophylax) are an endemic species of Messinian origin. Zoosyst. Evol. 2012, 88, 261–283. [Google Scholar] [CrossRef]
- Garcia, V.; Ivy, C.; Fu, J. Syntopic Frogs reveal different patterns of interaction with the landscape: A comparative landscape genetic study of Pelophylax nigromaculatus and Fejervarya limnocharis from central China. Ecol. Evol. 2017, 7, 9294–9306. [Google Scholar] [CrossRef] [PubMed]
- Borzée, A.; Andersen, D.; Jang, Y. Population trend inferred from aural surveys for calling anurans in Korea. PeerJ 2018, 2018, e5568. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.-Y.; Choi, S.; Kim, H.; Suh, J.-H.; Do, M.S. Ecological niche and interspecific competition of two frog species (Pelophylax nigromaculatus and P. chosenicus) in South Korea using the geographic information system. Korean J. Ecol. Environ. 2021, 54, 363–373. [Google Scholar] [CrossRef]
- Nakanishi, K.; Honma, A.; Furukawa, M.; Takakura, K.-I.; Fujii, N.; Morii, K.; Terasawa, Y.; Nishida, T. Habitat partitioning of two closely related pond frogs, Pelophylax nigromaculatus and Pelophylax porosus brevipodus, during their breeding season. Evol. Ecol. 2020, 34, 855–866. [Google Scholar] [CrossRef]
- Yang, S.; Kim, J.; Min, M.; Suh, J. Genetic diversity and population structure of two Korean pond frog species, Rana nigromaculata and R. plancyi (Anura, Ranidae), with a survey of temporal genetic variation in R. nigromaculata. Korean J. Biol. Sci. 1999, 3, 275–283. [Google Scholar] [CrossRef]
- Wang, S.; Liu, C.; Zhu, W.; Gao, X.; Yiming, L. Tracing the origin of the Black-Spotted Frog, Pelophylax nigromaculatus, in the Xinjiang uyghur autonomous region. Asian Herpetol. Res. 2016, 7, 69–74. [Google Scholar]
- Bae, Y.H.; Park, J.; Othman, S.N.; Yikweon, J.; Borzée, A. Record of invasive Rana huanrenensis Fei, Ye, and Huang, 1990 and Pelophylax nigromaculatus (Hallowell, 1861) on Ulleung Island, Republic of Korea. BioInvasions Rec. 2022, 11, 278–286. [Google Scholar] [CrossRef]
- Lyapkov, S.M.; Ermakov, O.A.; Titov, S.V. Distribution and origin of two forms of the Marsh Frog Pelophylax ridibundus complex (Anura, Ranidae) from Kamchatka based on mitochondrial and nuclear DNA data. Biol. Bull. 2018, 45, 699–705. [Google Scholar] [CrossRef]
- Dufresnes, C.; Mazepa, G. Hybridogenesis in Water Frogs. eLS 2020, 1, 718–726. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.; Augeri, D.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Zhu, G.; Qiao, H. Effect of the maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Kass, J.M.; Pinilla-Buitrago, G.E.; Paz, A.; Johnson, B.A.; Grisales-Betancur, V.; Meenan, S.I.; Attali, D.; Broennimann, O.; Galante, P.J.; Maitner, B.S.; et al. Wallace 2: A shiny app for modeling species niches and distributions redesigned to facilitate expansion via module contributions. Ecography 2023, 2023, 1–9. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Do, M.S.; Son, S.-J.; Choi, G.; Yoo, N.; Koo, K.-S.; Nam, H.-K. Anuran community patterns in the rice fields of the mid-western region of the Republic of Korea. Glob. Ecol. Conserv. 2021, 26, e01448. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Borzée, A.; Shin, Y.; Bae, Y.; Jeong, D.; Amin, H.; Min, M.-S.; Othman, S.N. From Korean to northeast Asian endemicity: On the occurrence of Pelophylax chosenicus in China. Front. Biogeogr. 2024, 16, e62301. [Google Scholar]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Dolgener, N.; Freudenberger, L.; Schluck, M.; Schneeweiss, N.; Ibisch, P.L.; Tiedemann, R. Environmental niche factor analysis (ENFA) relates environmental parameters to abundance and genetic diversity in ane amphibian, the fire-bellied-toad (Bombina bombina). Conserv. Genet. 2014, 15, 11–21. [Google Scholar] [CrossRef]
- Dubois, A.; Ohler, A. Frogs of the Subgenus Pelophylax (Amphibia, Anura, Genus Rana): A catalogue of available and valid scientific names, with comments on name-bearing types, complete synonymies, proposed common names, and maps showing all type localities. Zool. Pol. 1994, 39, 139–204. [Google Scholar]
- Matsui, M.; Maeda, N. Frogs and Toads of Japan; Bun-Ichi Sogo Shuppan: Tokyo, Japan, 1990. [Google Scholar]
- Hirai, T.; Matsui, M. Feeding relationships between Hyla japonica and Rana nigromaculata in rice fields of Japan. J. Herpetol. 2002, 36, 662–667. [Google Scholar] [CrossRef]
- Kanai, R.; Funao, T.; Nakanishi, K.; Sawad, H. Distribution survey of frogs by its call in paddy fields in Shiga, central Japan. Jpn. J. Environ. Entomol. 2014, 25, 121–126. [Google Scholar]
- Hirai, T.; Matsui, M. Feeding habits of the pond frog, Rana nigromaculata, inhabiting rice fields in kyoto, Japan. Copeia 1999, 1999, 940–947. [Google Scholar] [CrossRef]
- Togane, D.; Fukuyama, K.; Takai, K.; Kuramoto, N. Body size and age structure in two populations of Tokyo Daruma Pond Frog, Pelophylax porosus porosus. Curr. Herpetol. 2018, 37, 58–68. [Google Scholar] [CrossRef]
- Shimoyama, R. Preliminary report on male territoriality in the pond frog, Rana nigromaculata, in the breeding season. Jpn. J. Herpetol. 1982, 9, 99–102. [Google Scholar] [CrossRef] [PubMed]
- IUCN SSC Red List of Treatened Species. Pelophylax nigromaculatus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58679A63862076; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Tsuji, M.; Ushimaru, A.; Osawa, T.; Mitsuhashi, H. Paddy-associated frog declines via urbanization: A test of the dispersal-dependent-decline hypothesis. Landsc. Urban Plan. 2011, 103, 318–325. [Google Scholar] [CrossRef]
- Peters, W.C.H. Über Fledertheire (Pteropus gouldii, Rhinolophus deckenii, Vespertilio lobipes, Vesperugo temminckii) und Amphibien (Hypsilurus godeffroyi, Lygosoma scutatum, Stenostoma narisostre, Onychocephalus unguirostris, Ahaetulla polylepis, Pseudechis scutellatus); Monatsberichte der Königlichen Preussische Akademie des Wissenschaften zu Berlin: Berlin, Germany, 1867; pp. 703–712. [Google Scholar]
- Peter, C.; Norliza, A.; Ngeian, J.; Tuen, A.A.; Minton, G. Naturalists, Explorers and Field Scientists in South-East Asia and Australasia; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Komaki, S.; Kurabayashi, A.; Islam, M.M.; Tojo, K.; Sumida, M. Distributional change and epidemic introgression in overlapping areas of Japanese Pond Frog species over 30 Years. Zoolog. Sci. 2012, 29, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, H.T.; Ji, Y.J.; Shi, C.M.; Yang, Y.H.; Zhang, D.X. Black-Spotted Pond Frog (Pelophylax nigromaculatus) on the chinese loess plateau represents a cryptic species: Evidence from polecular phylogeny and ecological niche modeling. J. Syst. Evol. 2015, 53, 339–350. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, J.; Zhang, G.; Zhou, K. Phylogeography and demographic history of Chinese Black-Spotted Frog populations (Pelophylax nigromaculata): Evidence for independent refugia expansion and secondary contact. BMC Evol. Biol. 2008, 8, 21. [Google Scholar] [CrossRef]
- Liu, K.; Wang, F.; Chen, W.; Tu, L.; Min, M.S.; Bi, K.; Fu, J. Rampant historical mitochondrial genome introgression between two species of green pond frogs, Pelophylax nigromaculatus and P. plancyi. BMC Evol. Biol. 2010, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Groffen, J.; Andersen, D.; Borzée, A. Breeding phenology and landscape use in all amphibian species from the Republic of Korea based on open-source data. Front. Environ. Sci. 2022, 10, 846481. [Google Scholar] [CrossRef]
- Yoo, E.; Yikweon, J. Abiotic effects on calling phenology of three frog species in Korea. Anim. Cells Syst. 2011, 16, 1–8. [Google Scholar] [CrossRef]
- Yoon, I.B.; Kim, J.I.; Yang, S.Y. Study on the food habits of Rana nigromaculata Hallowell and Rana plancyi chosenica Okada (Salientia; Ranidae) in Korea. Korean J. Environ. Biol. 1998, 16, 69–76. [Google Scholar]
- Borzée, A.; Litvinchuk, S.N.; Ri, K.; Andersen, D.; Nam, T.Y.; Jon, G.H.; Man, H.S.; Choe, J.S.; Kwon, S.; Othman, S.N.; et al. Update on distribution and conservation status of amphibians in the Democratic People’s Republic of Korea: Conclusions based on field surveys, environmental modelling, molecular analyses and call properties. Animals 2021, 11, 2057. [Google Scholar] [CrossRef] [PubMed]
- Min, M.; Yan, H.; Zhiping, M.; Yanhong, L.; Zhou, C. Skeletochronological study of age, longevity and growth in a population of Rana nigromaculata (Amphibia: Anura) in Sichuan, China. Asian Herpetol. Res. 2012, 3, 258–264. [Google Scholar]
- Liao, W.B.; Zhou, C.; Yang, Z.; Hu, J.; Lu, X. Age, size and growth in two populations of the Dark-Spotted Frog Rana nigromaculata at different altitudes in southwestern China. Herpetol. J. 2010, 20, 77–82. [Google Scholar]
- Park, J.-K.; Chung, K.; Kim, J.; Do, Y. Population structure and morphological pattern of the Black-Spotted Pond Frog (Pelophylax nigromaculatus) inhabiting watershed areas of the Geum River in South Korea. Sustainability 2022, 14, 16530. [Google Scholar] [CrossRef]
- Huang, M.-Y.; Zhao, Q.; Duan, R.-Y.; Liu, Y.; Wan, Y.-Y. The effect of atrazine on intestinal histology, microbial community and short chain fatty acids in Pelophylax nigromaculatus tadpoles. Environ. Pollut. 2021, 288, 117702. [Google Scholar] [CrossRef]
- Liu, P.-Y.; Meng, T.; Li, Y.; Cai, M.; Li, X.-H.; Chen, J.; Qin, Z.-F. Tetrabromoethylcyclohexane affects gonadal differentiation and development in the frog Pelophylax nigromaculatus. Aquat. Toxicol. 2017, 192, 40–47. [Google Scholar] [CrossRef]
- Fan, X.; Lin, Z.; Li, X.; Wei, L.; Ding, G.-H. Effects of predation by invasive western mosquitofish (Gambusia affinis) on survival of eggs, embryos and tadpoles of Pelophylax nigromaculatus and Duttaphrynus melanostictus in south China. Asian Herpetol. Res. 2016, 7, 46–52. [Google Scholar]
- Groffen, J.; Kong, S.; Yikweon, J.; Borzée, A. The invasive american Bullfrog (Lithobates catesbeianus) in the Republic of Korea: History and recommendations for population control. Manag. Biol. Invasions 2019, 10, 517–535. [Google Scholar] [CrossRef]
- Shin, Y.; Kim, K.; Groffen, J.; Woo, D.; Song, E.; Borzée, A. Citizen science and roadkill trends in the Korean herpetofauna: The importance of spatially biased and unstandardized Data. Front. Ecol. Evol. 2022, 10, 944318. [Google Scholar] [CrossRef]
- Takahashi, K.; Takeuchi, H. Selection of calling sites by Pelophylax porosus porosus (Anura: Ranidae). Acta Biol. 2021, 28, 41–51. [Google Scholar] [CrossRef]
- Matsushima, N.; Hasegawa, M.; Nishihiro, J. Effects of landscape heterogeneity at multiple spatial scales on paddy field-breeding frogs in a large alluvial Plain in Japan. Wetlands 2022, 42, 106. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Pelophylax porosus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Naito, R.; Yamasaki, M.; Lmanishi, A.; Natuhara, Y.; Morimoto, Y. Effects of water management, connectivity, and surrounding land use on habitat use by frogs in rice paddies in Japan. Zoolog. Sci. 2012, 29, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Okochi, I. Captive breeding and the re-inforcement to an endangered population of Rana porosa brevipoda Ito (Ranidae: Amphibia). Jpn. J. Conserv. Ecol. 1997, 2, 135–146. [Google Scholar]
- Naito, J.; Sakamura, A.; Nakayama, T.; Matsubara, C. The conservation on the Daruma Pond Frog (Rana porosa brevipoda) in biotope area of Haizuka Dam. Hibakagaku 2014, 250, 1–27. [Google Scholar]
- Liu, C. Amphibians of Western China; Chicago Natural History Museum: Chicago, IL, USA, 1950; Volume 2, ISBN 0430-3776. [Google Scholar]
- Zhou, S.-B.; Zhang, Z.-H.; Hui, W.; Qing, M.; Xu, S.-J.; Guan, P.; Shi, J.-S. New record species of anura from China—Pelophylax chosenicus. Chin. J. Zool. 2022, 57, 440–446. [Google Scholar]
- Zhou, S.; He, L.; Ma, S.; Xu, S.; Zhai, Q.; Guan, P.; Wang, H.; Shi, J. Taxonomic status of Rana nigromaculata mongolia and the validity of Pelophylax tenggerensis (Anura, Ranidae). Zootaxa 2022, 5165, 486–500. [Google Scholar] [CrossRef]
- Duan, R.Y.; Kong, X.Q.; Huang, M.Y.; Varela, S.; Ji, X. The potential effects of climate change on amphibian distribution, range fragmentation and turnover in China. PeerJ 2016, 2016, e2185. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Zhao, E.M.; Jiang, Y.M. A Study of vocalization on thirteen species of four genera, Anura. Herpetology 1992, 15, 26. [Google Scholar]
- Zhou, S.-B.; Zhang, Q.-Y.; Hu, Z.-Q.; Xia, Z.-Y.; Miao, Q.; Guan, P.; Shi, J.-S. The validity of Pelophylax chosenicus (Okada, 1931) and P. hubeiensis (Fei & Ye, 1982) (Amphibia, Ranidae). Herpetozoa 2023, 36, 143–152. [Google Scholar] [CrossRef]
- Borzée, A.; Ren, J.-L.; Li, J.-T.; Groffen, J.; Yang, Y.; Jang, Y. Microhabitat segregation between Black-Spotted Pond Frogs, Pelophylax nigromaculatus, and Gold-Striped Pond Frogs, P. plancyi (Anura: Ranidae). Reptil. Amphib. 2019, 26, 119–120. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Pelophylax plancyi (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Borzée, A.; Kyong, C.N.; Kil, H.K.; Jang, Y. Impact of water quality on the occurrence of two endangered Korean anurans: Dryophytes suweonensis and Pelophylax chosenicus. Herpetologica 2018, 74, 1–7. [Google Scholar] [CrossRef]
- Wei, X.; Huang, M.; Yue, Q.; Ma, S.; Li, B.; Mu, Z.; Peng, C.; Gao, W.; Liu, W.; Zheng, J.; et al. Long-term urbanization impacts the Eastern Golden Frog (Pelophylax plancyi) in Shanghai City: Demographic history, genetic structure, and implications for amphibian conservation in intensively urbanizing environments. Evol. Appl. 2021, 14, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, B.; Shu, X.; Pei, E.; Yuan, X.; Sun, Y.; Wang, T.; Wang, Z. Responses of anuran communities to rapid urban growth in Shanghai, China. Urban For. Urban Green. 2016, 20, 365–374. [Google Scholar] [CrossRef]
- Shu, X.; Zhang, W.; Li, B.; Pei, E.; Yuan, X.; Wang, T.; Wang, Z. Major factors affecting the distribution of anuran communities in the urban, suburban and rural areas of Shanghai, China. Asian Herpetol. Res. 2016, 7, 287–294. [Google Scholar]
- Okada, Y. Frogs in Korea. J. Chosen Biol. Soc. 1928, 6, 15–46. [Google Scholar]
- Andersen, D.; Chuang, M.F.; Choe, M.; Kim, A.; Kwon, S.; Jang, Y.; Borzée, A. Elevational distribution of amphibians: Resolving distributions, patterns, and species communities in the Republic of Korea. Zool. Stud. 2022, 61, e25. [Google Scholar]
- Ra, N.Y.; Sung, H.C.; Cheong, S.K.; Lee, J.H.; Eom, J.; Park, D. Habitat use and home range of the endangered Gold-Spotted Pond Frog (Rana chosenica). Zoolog. Sci. 2008, 25, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.; Lee, J.-B.; Ra, N.-Y.; Park, D.-S. Preferred feeding sites and prey of the adult Gold-Spotted Pond Frog, Rana plancyi chosenica. J. Ecol. Environ. 2007, 30, 357–361. [Google Scholar] [CrossRef]
- Koo, K.S.; Kwon, S.; Kwak, M.; Oh, J. First record of heterospecific amplexus behaviour between Pelophylax chosenicus (Anura: Ranidae) and Dryophytes japonicus (Anura: Hylidae) in Paju, Republic of Korea. Herpetol. Notes 2021, 14, 1225–1226. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax chosenicus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Park, C.-D.; Kwon, K.; Yoo, N.; Lee, J.-H.; Kang, D.; Park, J.-S.; Yoo, J.; Kim, K.-S.; Yoon, J.-D. Post-release monitoring after reintroduction of captive-reared Korean endangered Frog, Pelophylax chosenicus. Proc. Natl. Inst. Ecol. Repub. Korea 2021, 2, 114–119. [Google Scholar]
- Yoo, N.; Do, M.S.; Nam, H.-K.; Choi, G.; Son, S.-J.; Yoo, J. Habitat characteristics of anuran species inhabiting rice fields of western Mid-South Korea. Korean J. Ecol. Environ. 2019, 52, 366–377. [Google Scholar] [CrossRef]
- Fong, J.J.; Cheng, T.L.; Bataille, A.; Pessier, A.P.; Waldman, B.; Vredenburg, V.T. Early 1900s detection of Batrachochytrium dendrobatidis in Korean amphibians. PLoS ONE 2015, 10, e0115656. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Ye, C. The distributional characteristics of amphibian in Hubei province, including description of a new frog. Acta Zool 1982, 28, 293–301. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax hubeiensis (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Pope, C. Four new frogs from Fukien Province, China. Am. Museum Novit. 1929, 1–5. [Google Scholar]
- Eo, S.H.E.; Lee, B.J.; Park, C.D.; Jung, J.H.; Hong, N.; Lee, W.S. Taxonomic identity of the Glandirana emeljanovi (Anura, Ranidae) in Korea revealed by the complete mitochondrial genome sequence analysis. Mitochondrial DNA Part B Resour. 2019, 4, 961–962. [Google Scholar] [CrossRef]
- Jang-Liaw, N.-H.; Chou, W.-H. Anuran fauna of Taiwan and adjacent islands based on valid specimen records. Coll. Res. 2015, 28, 5–53. [Google Scholar]
- Sumida, M.; Ogata, M.; Nishioka, M. Molecular phylogenetic relationships of pond frogs distributed in the Palearctic region inferred from DNA sequences of mitochondrial 12S ribosomal RNA and cytochrome b genes. Mol. Phylogenet. Evol. 2000, 16, 278–285. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Pelophylax fukienensis (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Arif, S.M. Endoparasitic nematohelminthes collected from inside frog Rana ridibunda (Pallas, 1771) in two rivers near Baghdad/Iraq. J. Pharm. Sci. Res. 2019, 11, 2080–2083. [Google Scholar]
- Jablonski, D.; Van Bemmel, S.; Masroor, R. Notes on the herpetofauna of Kunduz Province, northeastern Afghanistan, with the first record of Varanus griseus caspius (Eichwald, 1831). Herpetol. Notes 2021, 14, 63–66. [Google Scholar]
- Kaptyonkina, A.G.; Dujsebayeva, T.N.; Akhmedenov, K.M.; Khromov, V.A.; Krainyuk, V.N.; Sarzhanov, F.; Starikov, S.V.; Tarasovskaya, N.E.; Timoshenko, A.Y.; Titov, S.V. The range of Marsh Frogs (Complex Pelophylax ridibundus, Amphibia, Ranidae) in Kazakhstan: Progressive dispersal or cyclic fluctuations? Proc. Zool. Inst. Russ. Acad. Sci. 2022, 326, 211–238. [Google Scholar] [CrossRef]
- Khalaf. Reptiles of Iraq: With Some Notes on the Amphibians; Ar-Rabitta Press: Charlottesville, VA, USA, 1959. [Google Scholar]
- Mohammadi, Z.; Khajeh, A.; Ghorbani, F.; Kami, H.G. A biosystematic sudy of new records of the Marsh Frog Pelophylax ridibundus (Pallas, 1771) (Amphibia: Ranidae) from southeast of Iran. J. Asia-Pac. Biodivers. 2015, 31, 178–182. [Google Scholar] [CrossRef]
- Papežík, P.; Sciberras, A.; Benovics, M.; Sciberras, J.; Deidun, A.; Mikulicek, P. Far from home: Tracing the origin of non-native water frogs (Genus Pelophylax) in Malta by molecular markers. Biol. Invasions 2024, 26, 1045–1059. [Google Scholar] [CrossRef]
- Plötner, J. Die Westpaläarktischen Wasserfrösche: Von Märtyrern der Wissenschaft zur Biologischen Sensation; Laurenti: Bielefeld, Germany, 2005. [Google Scholar]
- Sánchez-Montes, G.; Recuero, E.; Gutiérrez-Rodríguez, J.; Gomez-Mestre, I.; Martinez-Solano, I. Species assignment in the Pelophylax ridibundus × P.perezi hybridogenetic complex based on 16 newly characterized microsatellite markers. Herpetol. J. 2016, 26, 99–108. [Google Scholar]
- Ualiyeva, D.; Ermakov, O.A.; Litvinchuk, S.N.; Guo, X.; Ivanov, A.Y.; Xu, R.; Li, J.; Xu, F.; Arifulova, I.I.; Kaptyonkina, A.G.; et al. Diversity, phylogenetic relationships and distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and northwest China. Diversity 2022, 14, 869. [Google Scholar] [CrossRef]
- Bayrakci, Y.; Çiçek, K. Breeding ecology and larval development of Marsh Frogs (Pelophylax ridibundus s.l.) from East Mediterranean, Türkiye. Commagene J. Biol. 2023, 7, 65–72. [Google Scholar] [CrossRef]
- Baran, İ.; Atatür, M.K. Turkish Herpetofauna: Amphibians and Reptiles; Turkey Ministry of Environment: Ankara, Turkey, 1997. [Google Scholar]
- Borkin, L. Cryptic species (a case study of amphibians). Entomol. Rev 2004, 84, 75–98. [Google Scholar]
- Dufresnes, C.; Leuenberger, J.; Amrhein, V.; Bühler, C.; Thiébaud, J.; Bohnenstengel, T.; Dubey, S. Invasion genetics of Marsh Frogs (Pelophylax ridibundus sensu lato) in Switzerland. Biol. J. Linn. Soc. 2018, 123, 402–410. [Google Scholar] [CrossRef]
- Dujsebayeva, T.N.; Ivanov, A.Y.; Kaptyonkina, A.G.; Ualiyeva, D.A.; Krainyuk, V.N.; Cherednichenko, A.V.; Khromov, V.A. The Marsh Frogs (Pelophylax ridibundus complex) in central Kazakhstan: Expansion and retreat. Russ. J. Ecosyst. Ecol. 2021, 6, 1–18. [Google Scholar] [CrossRef]
- Holsbeek, G.; Jooris, R. Potential impact of genome exclusion by alien species in the hybridogenetic Water Frogs (Pelophylax esculentus complex). Biol. Invasions 2010, 12, 1–13. [Google Scholar] [CrossRef]
- Hotz, H.; Beerli, P.; Uzzell, T.; Guex, G.-D.; Pruvost, N.; Schreiber, R.; Plötner, J. Balancing a cline by influx of migrants: A genetic transition in Water Frogs of eastern Greece. J. Hered. 2012, 104, 57–71. [Google Scholar] [CrossRef]
- Jablonski, D.; Khalili, F.; Masroor, R. The herpetofaunal diversity of Takhar Province, Afghanistan. Herpetozoa 2023, 90, 73–90. [Google Scholar]
- Domeneghetti, D.; Bruni, G.; Fasola, M.; Bellati, A. Discovery of alien Water Frogs (gen. Pelophylax) in Umbria, with first report of P. shqipericus for Italy. Acta Herpetol. 2013, 8, 171–176. [Google Scholar]
- Laghi, P.; Miserocchi, D.; Valli, M. Determinazione genetica della presenza delle rane verdi alloctone Pelophylax ridibundus e Pelophylax kurtmuelleri (Amphibia, Anura, Ranidae) in due località della Romagna. Quad. Mus. Stor. Nat. Ferrara 2013, 1, 75–78. [Google Scholar]
- Pagano, A.; Joly, P.; Plénet, S.; Lehman, A.; Grolet, O. Breeding habitat partitioning in the Rana esculenta complex: The intermediate niche hypothesis supported. Ecoscience 2001, 8, 294–300. [Google Scholar] [CrossRef]
- Dubey, S.; Leuenberger, J.; Perrin, N. Multiple origins of invasive and ‘native’ Water Frogs (Pelophylax spp.) in Switzerland. Biol. J. Linn. Soc. 2014, 112, 442–449. [Google Scholar] [CrossRef]
- Holsbeek, G.; Mergeay, J.; Hotz, H.; Plötner, J.; Volckaert, F.; De Meester, L. A cryptic invasion within an invasion and widespread introgression in the European Water Frog complex: Consequences of uncontrolled commercial trade and weak international legislation. Mol. Ecol. 2008, 17, 5023–5035. [Google Scholar] [CrossRef]
- Holsbeek, G.; Mergeay, J.; Volckaert, F.; De Meester, L. Genetic detection of multiple exotic Water frog species in Belgium illustrates the need for monitoring and immediate action. Biol. Invasions 2010, 12, 1459–1463. [Google Scholar] [CrossRef]
- Ermakov, O.; Ivanov, A.; Titov, S.; Svinin, A.; Litvinchuk, S.N. New Multiplex PCR method for identification of East European Green Frog species and their hybrids. Russ. J. Herpetol. 2019, 26, 367–370. [Google Scholar] [CrossRef]
- Sah, A.K.; Gautam, B.; Bhattarai, S. Biological aspects of the Marsh Frog Pelophylax ridibundus population from the ahadegan wetland, Khuzestan Province, Iran. J. Anim. Divers. 2020, 2, 18–26. [Google Scholar]
- Lukanov, S.P.; Tzankov, N.D.; Simeonovska-Nikolova, D.M. Effects of environmental factors on mating call characteristics of the Marsh Frog Pelophylax ridibundus (Pallas, 1771) (Amphibia: Ranidae) in Bulgaria. Acta Zool. Bulg. 2014, 66, 209–216. [Google Scholar]
- Balint, N.; Citrea, L.; Memetea, A.; Jurj, N.; Condure, N. Feeding biology of the Pelophylax ridibundus Pallas 1771, (Anura, Ranidae) in Dobromir, Romania. Biharean Biol. 2008, 2, 27–37. [Google Scholar]
- Najibzadeh, M.; Darvish, J. Comparison of habitat, mating behavior and laying of three species of anuran amphibians (Rana (Pelophylax) ridibunda, Hyla savignyi and Bufo (Pseudepidalea) variabilis) in the Lorestan Province. J. Anim. Res. 2014, 27, 291–299. [Google Scholar]
- Erismis, U. Abundance, demography and population structure of Pelophylax ridibundus (Anura: Ranidae) in 26-August National Park (Turkey). North. West. J. Zool. 2011, 7, 5–16. [Google Scholar]
- Altun, C.; Altunışık, A. Comparison of the Marsh Frog (Pelophylax ridibundus) populations living in different altitudes in terms of age, size, and some growth parameters. Biol. Bull. 2023, 2023, 1–8. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax ridibundus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Quilodran, C.; Montoya-Burgos, J.; Currat, M. Modelling interspecific hybridization with genome exclusion to identify conservation actions: The case of native and invasive Pelophylax Waterfrogs. Evol. Appl. 2015, 8, 199–210. [Google Scholar] [CrossRef]
- Quilodran, C.; Austerlitz, F.; Currat, M.; Montoya-Burgos, J. Cryptic biological invasions: A general model of hybridization. Sci. Rep. 2018, 8, 2414. [Google Scholar] [CrossRef]
- Zhelev, Z.; Tsonev, S.; Georgieva, K. Health status of Pelophylax ridibundus (Amphibia: Ranidae) in a rice paddy ecosystem in southern Bulgaria and its importance in assessing environmental state: Haematological parameters. Environ. Sci. Pollut. Res. 2019, 25, 7884–7895. [Google Scholar] [CrossRef]
- Zhelev, Z.; Tsonev, S.; Boyadz, P. Using of fluctuating asymmetry in adult Pelophylax ridibundus (Amphibia: Anura:Ranidae) meristic traits as a method for a developmental stability of populationand environmental quality of their habitat: Industrial area in southern Bulgaria. Turkish J. Zool. 2022, 46, 220–227. [Google Scholar] [CrossRef]
- Lukanov, S.; Popgeorgiev, G.; Tzankov, N. First bioacoustic and morphological data for the presence of Pelophylax bedriagae in Bulgaria. Acta Sci. Nat. 2018, 5, 54–63. [Google Scholar] [CrossRef]
- Schneider, H. Calls and reproductive behaviour of the Water Frogs of Damascus, Syria (Amphibia: Anura: Rana Bedriagae Camerano, 1882). Zool. Middle East 1997, 15, 51–66. [Google Scholar] [CrossRef]
- Pesarakloo, A.; Najibzadeh, M.; Rastegar-Pouyani, N.; Rastegar-Pouyani, E. Taxonomic survey of Water Frog populations of Pelophylax bedriagae (Anura: Ranidae) in Western Iran: A morphometric and bioacoustic approach. Biologia 2018, 73, 673–681. [Google Scholar] [CrossRef]
- Schneider, H. Calls of the Levantine Frog, Rana bedriagae, at Birket Ata, Israel (Amphibia: Anura). Zool. Middle East 1999, 19, 101–116. [Google Scholar] [CrossRef]
- Plötner, J.; Akın Pekşen, Ç.; Baier, F.; Uzzell, T.; Bilgin, C. Genetic evidence for human-mediated introduction of Anatolian Water Frogs (Pelophylax cf. bedriagae) to Cyprus (Amphibia: Ranidae). Zool. Middle East 2015, 61, 125–132. [Google Scholar] [CrossRef]
- Goldberg, S. Notes on reproduction of Levant Green Frogs, Pelophylax bedriagae (Anura: Ranidae), from Israel. Bull. Chic. Herpetol. Soc. 2023, 58, 26–27. [Google Scholar]
- Dufresnes, C.; Denoël, M.; Di Santo, L.; Dubey, S. Multiple uprising invasions of Pelophylax Water Frogs, potentially inducing a new hybridogenetic complex. Sci. Rep. 2017, 7, 6506. [Google Scholar] [CrossRef]
- Ibrahim, A. First Record of Pelophylax bedriagae (Amphibia: Ranidae) in the Suez Canal area, Egypt. Herpetol. Notes 2011, 4, 331–332. [Google Scholar]
- Pesarakloo, A.; Najibzadeh, M. Life history of the Levant Water Frog, Pelophylax bedriagae (Amphibia: Anura: Ranidae) in Western Iran. J. Anim. Divers. 2019, 1, 11–19. [Google Scholar] [CrossRef]
- Pesarakloo, A.; Rastegar-Pouyani, E.; Rastegar-Pouyani, N.; Kami, H.; Najibzadeh, M.; Khosravani, A.; Oraie, H. The first taxonomic revaluation of the Iranian Water Frogs of the genus Pelophylax (Anura: Ranidae) using sequences of the mitochondrial genome. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2017, 28, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Khaloei, M.; Fathinia, B.; Shafaeipour, A. Skeletochronology of the Levant Green Frog, Pelophylax bedriagae (Camerano, 1882), in southern Iran. Curr. Herpetol. 2023, 42, 55–63. [Google Scholar] [CrossRef]
- Disi, A.; Amr, Z. Morphometrics, distribution and ecology of the amphibians in Jordan. Vertebr. Zool. 2010, 60, 147–162. [Google Scholar] [CrossRef]
- Özcan, L.; Yorulmaz, B.; Ayaz, D. Data on Food Composition of the Levant Marsh Frog (Pelophylax bedriagae) in Southwestern Anatolia. North. West. J. Zool. 2021, 17, 227–231. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax bedriagae (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Başkale, E.; Kaya, U. Decline of the Levantine Frog, Pelophylax bedriagae Camerano, 1882, in the western Aegean region of Turkey changes in population size and implications for conservation: (Amphibia: Ranidae). Zool. Middle East. 2012, 57, 69–76. [Google Scholar] [CrossRef]
- Innal, D.; Sozbilen, D.; Ozmen, O. Prevalence and pathology of Lernaea cyprinacea (Crustacea: Lernaeidae) parasitizing tadpoles of Pelophylax bedriagae (Anura: Ranidae) in Antalya, Turkey. Phyllomedusa 2019, 18, 177–183. [Google Scholar] [CrossRef]
- Mezhzherin, S.V. New species of Green Frogs Rana terentievi sp. nova (Amphibia, Ranidae) from South Tajikistan. Proc. Acad. Sci. Ukr. 1992, 5, 150–153. [Google Scholar]
- Pesarakloo, A. Central Asia or Minor Asia: Origin of the Water Frogs in the Southern Coast of the Caspian Sea. Soc. Sci. Res. Netw. 2022, preprint, 4178613. [Google Scholar]
- Mazepa, G. Evolution of Water Frogs Pelophylax in Central Asia: How Hybridization and Mitochondrial Introgression among Ecologically Divergent Species Promote Occupation of Novel Environment. Master’s Thesis, Uppsala University, Uppsala, Sweden, 2013. [Google Scholar]
- Kidov, A.A.; Ivolga, R.A.; Kondratova, T.E.; Ivanov, A.A. Age, Growth, and fertility in Terentiev’s Frog (Pelophylax terentievi, Amphibia, Ranidae). Biol. Bull. 2023, 50, 1854–1863. [Google Scholar] [CrossRef]
- Wagner, P.; Bauer, A.; Leviton, A.; Wilms, T.; Böhme, W. A Checklist of the amphibians and reptiles of Afghanistan—Exploring herpetodiversity using biodiversity archives. Proc. Calif. Acad. Sci. 2016, 63, 457–565. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax terentievi (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Akin, C.P. Molecular Evolution and Phylogeography of the Eastern Mediterranean Water Frog (Pelophylax) Complex. Ph.D. Thesis, School of Natural and Applied Sciences of Middle East Technical University, Ankara, Turkey, 2015. [Google Scholar]
- Akin, C.; Bilgin, C.C.; Beerli, P.; Westaway, R.; Ohst, T.; Litvinchuk, S.N.; Uzzell, T.; Bilgin, M.; Hotz, H.; Guex, G.D.; et al. Phylogeographic patterns of genetic diversity in eastern mediterranean water frogs were determined by geological processes and climate change in the late cenozoic. J. Biogeogr. 2010, 37, 2111–2124. [Google Scholar] [CrossRef] [PubMed]
- Ualiyeva, D.A.; Ivanov, A.Y.; Ermakov, O.A. A development of a PCR-RFLP test system for the identification of mitochondrial lines of the Pelophylax ridibundus Lake Frog in Kazakhstan. Univ. Proc. Volga Reg. Nat. Sci. 2022, 37, 76–84. [Google Scholar] [CrossRef]
- Lymberakis, P.; Poulakakis, N.; Manthalou, G.; Tsigenopoulos, C.S.; Magoulas, A.; Mylonas, M. Mitochondrial phylogeography of Rana (Pelophylax) populations in the eastern Mediterranean region. Mol. Phylogenet. Evol. 2007, 44, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.; Maslova, I.; Purevdorj, Z.; Li, J.T.; Messenger, K.R.; Ren, J.L.; Jang, Y.; Borzée, A. East palearctic treefrog past and present habitat suitability using ecological niche models. PeerJ 2022, 10, e12999. [Google Scholar] [CrossRef] [PubMed]
- Litvinchuk, S.; Skorinov, D.; Ivanov, A.; Ermakov, O. Detection of glacial refugia and post-glacial colonization routes of morphologically cryptic Marsh Frog species (Anura: Ranidae: Pelophylax) using environmental niche modeling. Diversity 2024, 16, 94. [Google Scholar] [CrossRef]
- Jablonski, D.; Gkontas, I.; Poursanidis, D.; Lymberakis, P.; Poulakakis, N. Stability in the Balkans: Phylogeography of the endemic Greek Stream Frog, Rana graeca. Biol. J. Linn. Soc. 2021, 132, 829–846. [Google Scholar] [CrossRef]
- Olalla-Tárraga, M.; McInnes, L.; Bini, L.; Diniz-Filho, J.A.; Fritz, S.; Hawkins, B.; Hortal, J.; Orme, D.; Rahbek, C.; Rodriguez, M.; et al. Climatic niche conservatism and the evolutionary dynamics in species range boundaries: Global congruence across mammals and amphibians. J. Biogeogr. 2011, 38, 2237–2247. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Future Global Climate: Scenario-Based Projections and Near-Term Information; Cambridge University Press: Cambridge, UK, 2023. [Google Scholar]
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; Borzée, A.; Hamidy, A.; Aowphol, A.; Jean, A. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature 2023, 622, 308–314. [Google Scholar] [CrossRef]
- Borzée, A.; Heo, K.; Jang, Y. Relationship between agro-environmental variables and breeding Hylids in rice paddies. Sci. Rep. 2018, 8, 8049. [Google Scholar] [CrossRef]
- Naito, R.; Sakai, M.; Morimoto, Y. Negative effects of deep roadside ditches on Pelophylax porosa brevipoda dispersal and migration in comparison with Hyla japonica in a rice paddy area in Japan. Zoolog. Sci. 2012, 29, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Naito, R.; Sakai, M.; Natuhara, Y.; Morimoto, Y.; Shibata, S. Microhabitat use by Hyla japonica and Pelophylax porosa brevipoda at levees in rice paddy areas of Japan. Zoolog. Sci. 2013, 30, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Borzée, A.; Andersen, D.; Groffen, J.; Kim, H.T.; Bae, Y.; Jang, Y. Climate change-based models predict range shifts in the distribution of the only Asian Plethodontid salamander: Karsenia koreana. Sci. Rep. 2019, 9, 11838. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Monod-Broca, B.; Bellati, A.; Canestrelli, D.; Ambu, J.; Wielstra, B.; Dubey, S.; Crochet, P.A.; Denoël, M.; Jablonski, D. Piecing the barcoding puzzle of Palearctic Water Frogs (Pelophylax) sheds light on amphibian biogeography and global invasions. Glob. Chang. Biol. 2024, 30, e17180. [Google Scholar] [CrossRef]
Figure 1.
Distribution of 12 Pelophylax lineages in central Asia. The species ranges are based on the IUCN Red List of Threatened Species [12], where available; otherwise, a minimum convex polygon is used. Black dots indicate populations where the species identity has not been clarified.
Figure 1.
Distribution of 12 Pelophylax lineages in central Asia. The species ranges are based on the IUCN Red List of Threatened Species [12], where available; otherwise, a minimum convex polygon is used. Black dots indicate populations where the species identity has not been clarified.

Figure 2.
Literature selection flow diagram used in the review. The literature was searched using Google Scholar (http://www.scholar.google.com; accessed on 15 December 2023). We used GBIF (http://www.gbif.org; accessed on 15 December 2023) and iNaturalist (http://www.inaturalist.org; accessed on 15 December 2023) independently for occurrence points, as some of the data that are not reaching the “Research Grade” on iNaturalist are not uploaded onto GBIF, but we were able to curate some additional records and clarify the species identification.
Figure 2.
Literature selection flow diagram used in the review. The literature was searched using Google Scholar (http://www.scholar.google.com; accessed on 15 December 2023). We used GBIF (http://www.gbif.org; accessed on 15 December 2023) and iNaturalist (http://www.inaturalist.org; accessed on 15 December 2023) independently for occurrence points, as some of the data that are not reaching the “Research Grade” on iNaturalist are not uploaded onto GBIF, but we were able to curate some additional records and clarify the species identification.

Figure 3.
Distribution of the Pelophylax nigromaculatus “nigromaculatus” lineage in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].
Figure 3.
Distribution of the Pelophylax nigromaculatus “nigromaculatus” lineage in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].

Figure 4.
Distribution of the Pelophylax nigromaculatus “reinhardtii” lineage in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on [12].
Figure 4.
Distribution of the Pelophylax nigromaculatus “reinhardtii” lineage in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on [12].

Figure 5.
Distribution of Pelophylax porosus in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
Figure 5.
Distribution of Pelophylax porosus in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).

Figure 6.
Distribution of Pelophylax mongolius in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (yellow and above); (B) Range of the species (red outline) based on (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].
Figure 6.
Distribution of Pelophylax mongolius in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (yellow and above); (B) Range of the species (red outline) based on (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].

Figure 7.
Distribution of Pelophylax plancyi in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].
Figure 7.
Distribution of Pelophylax plancyi in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].

Figure 8.
Distribution of Pelophylax chosenicus in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].
Figure 8.
Distribution of Pelophylax chosenicus in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on (https://www.iucnredlist.org/; accessed on 15 December 2023) and [12].

Figure 9.
Distribution of Pelophylax hubeiensis in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
Figure 9.
Distribution of Pelophylax hubeiensis in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).

Figure 10.
Distribution of Pelophylax fukienensis in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
Figure 10.
Distribution of Pelophylax fukienensis in Eastern Asia. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).

Figure 11.
Distribution of Pelophylax ridibundus. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
Figure 11.
Distribution of Pelophylax ridibundus. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).

Figure 12.
Distribution of Pelophylax bedriagae. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
Figure 12.
Distribution of Pelophylax bedriagae. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).

Figure 13.
Distribution of Pelophylax terentievi. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).
Figure 13.
Distribution of Pelophylax terentievi. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on the IUCN Red List of Threatened Species (https://www.iucnredlist.org/; accessed on 15 December 2023).

Figure 14.
Distribution of the Pelophylax “Syr Darya” lineage. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on a maximum convex polygon.
Figure 14.
Distribution of the Pelophylax “Syr Darya” lineage. (A) Maxent habitat suitability models using the maximum training sensitivity plus specificity threshold (MTSS) of the models as the cut-off for moderate suitability (orange and red); (B) Range of the species (red outline) based on a maximum convex polygon.


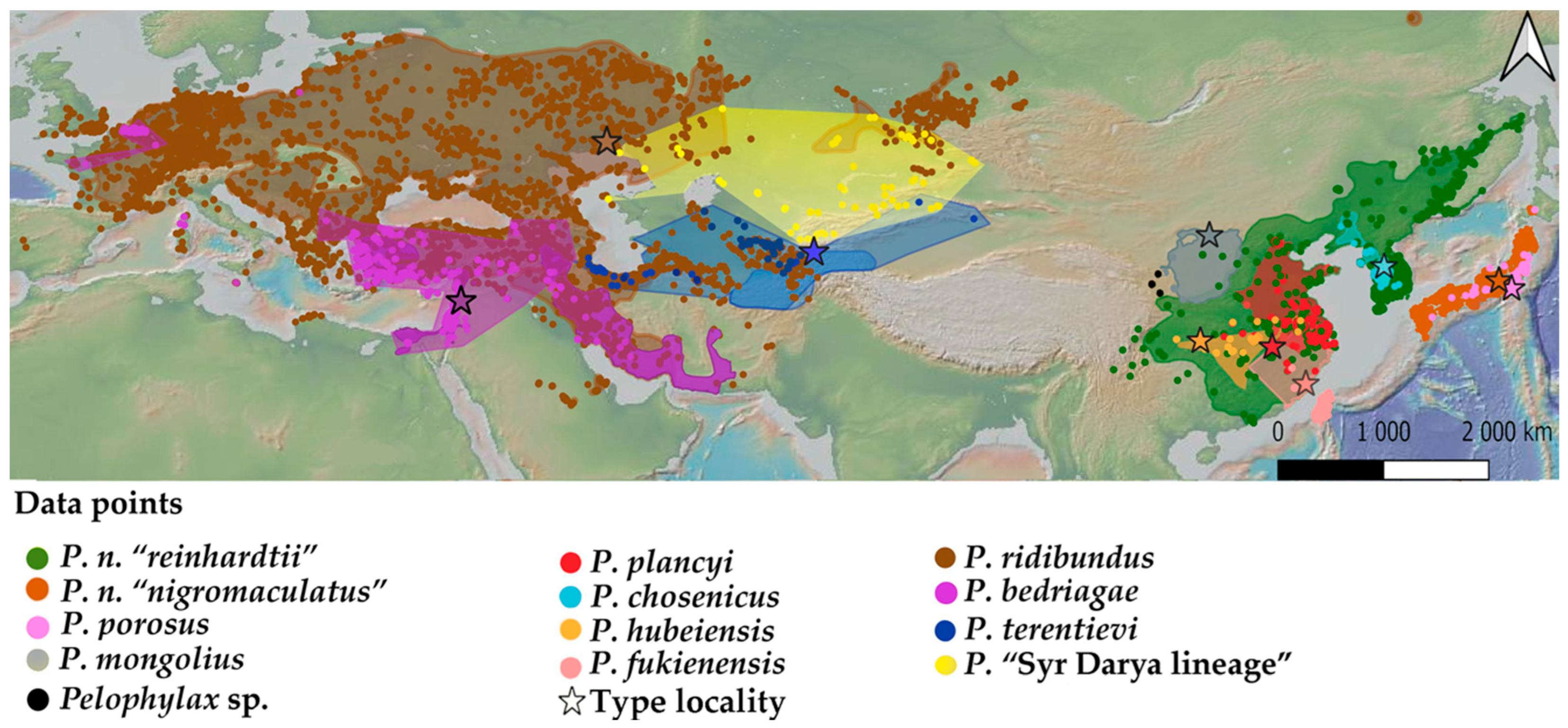{kind=link}
{kind=link}
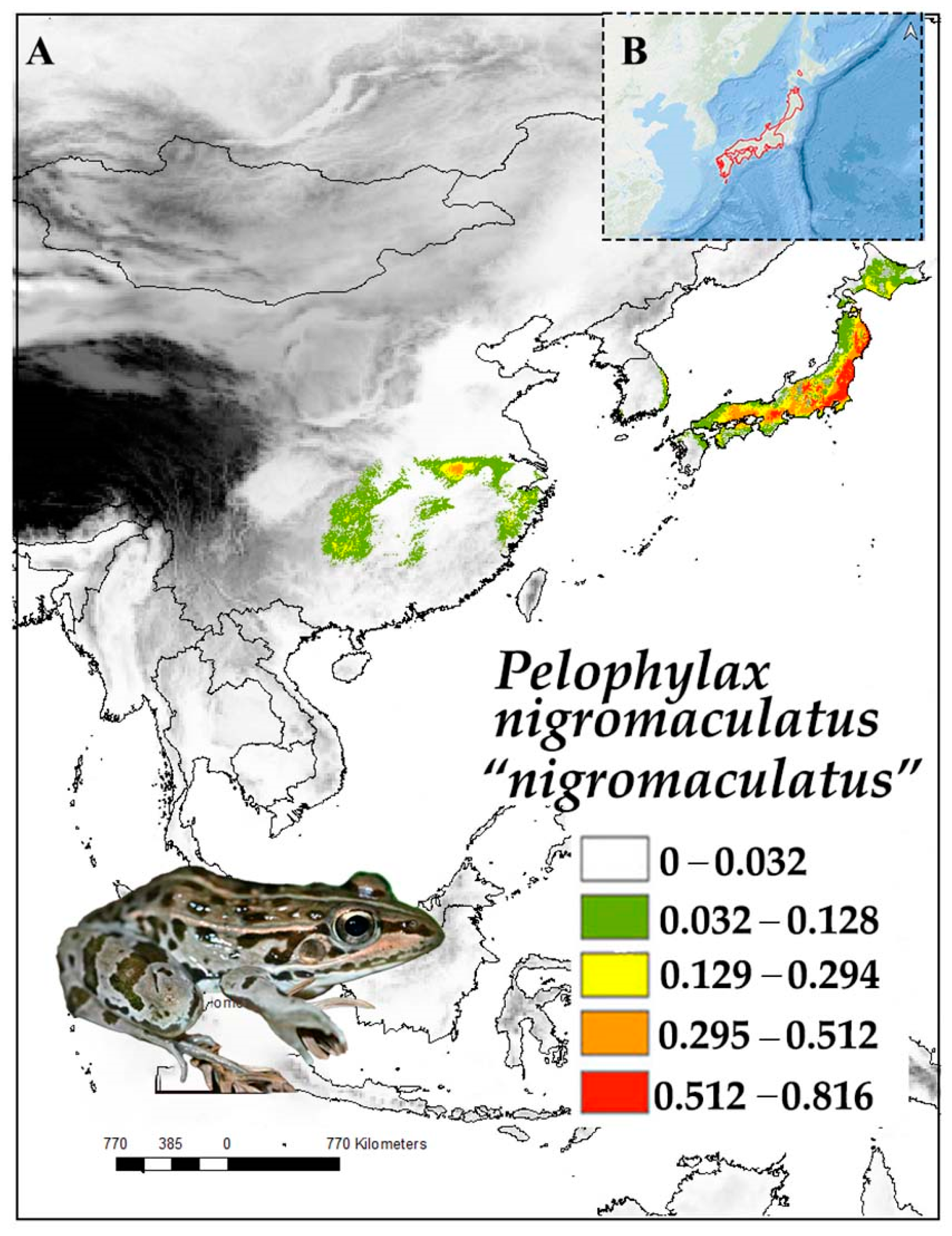{kind=link}
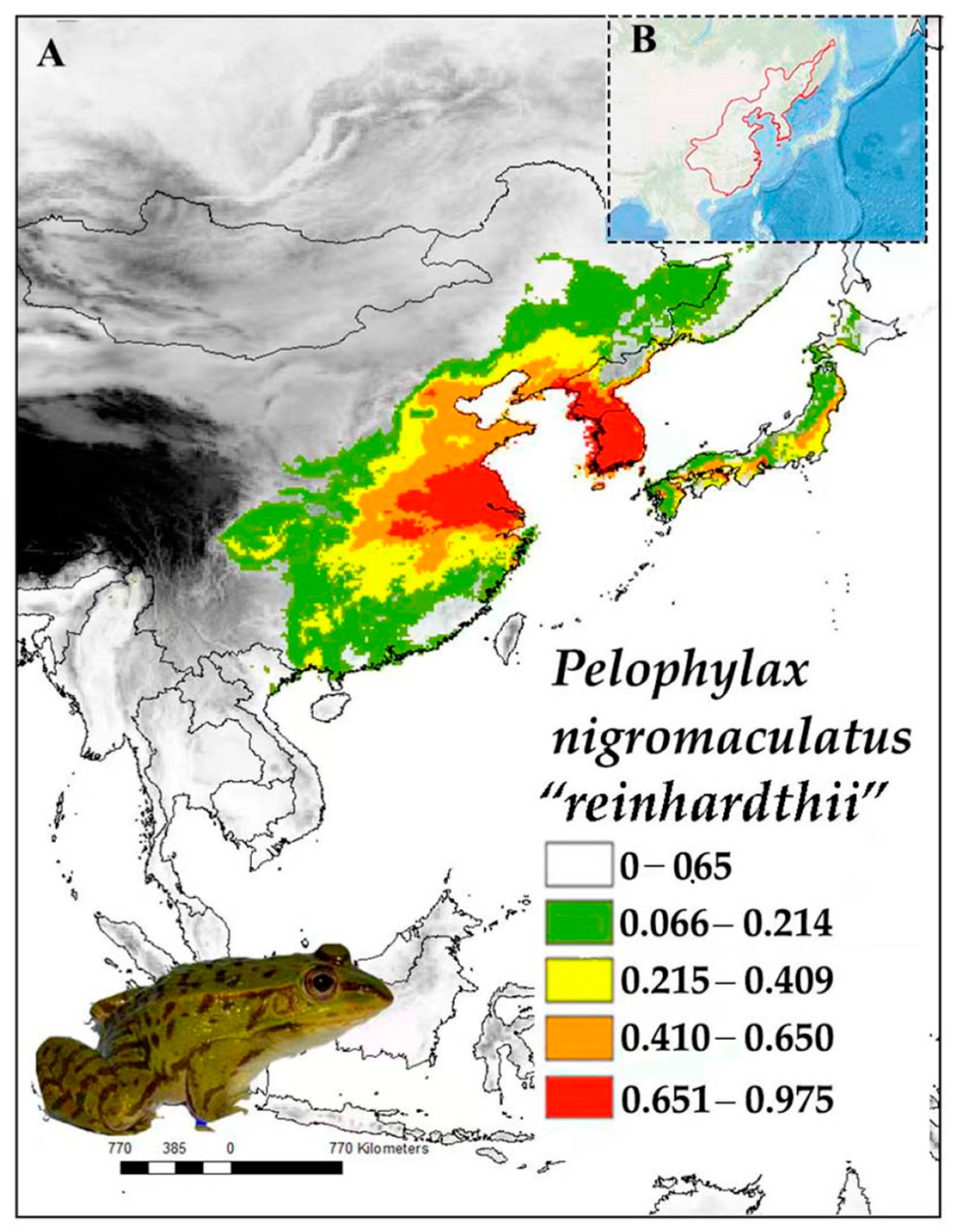{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
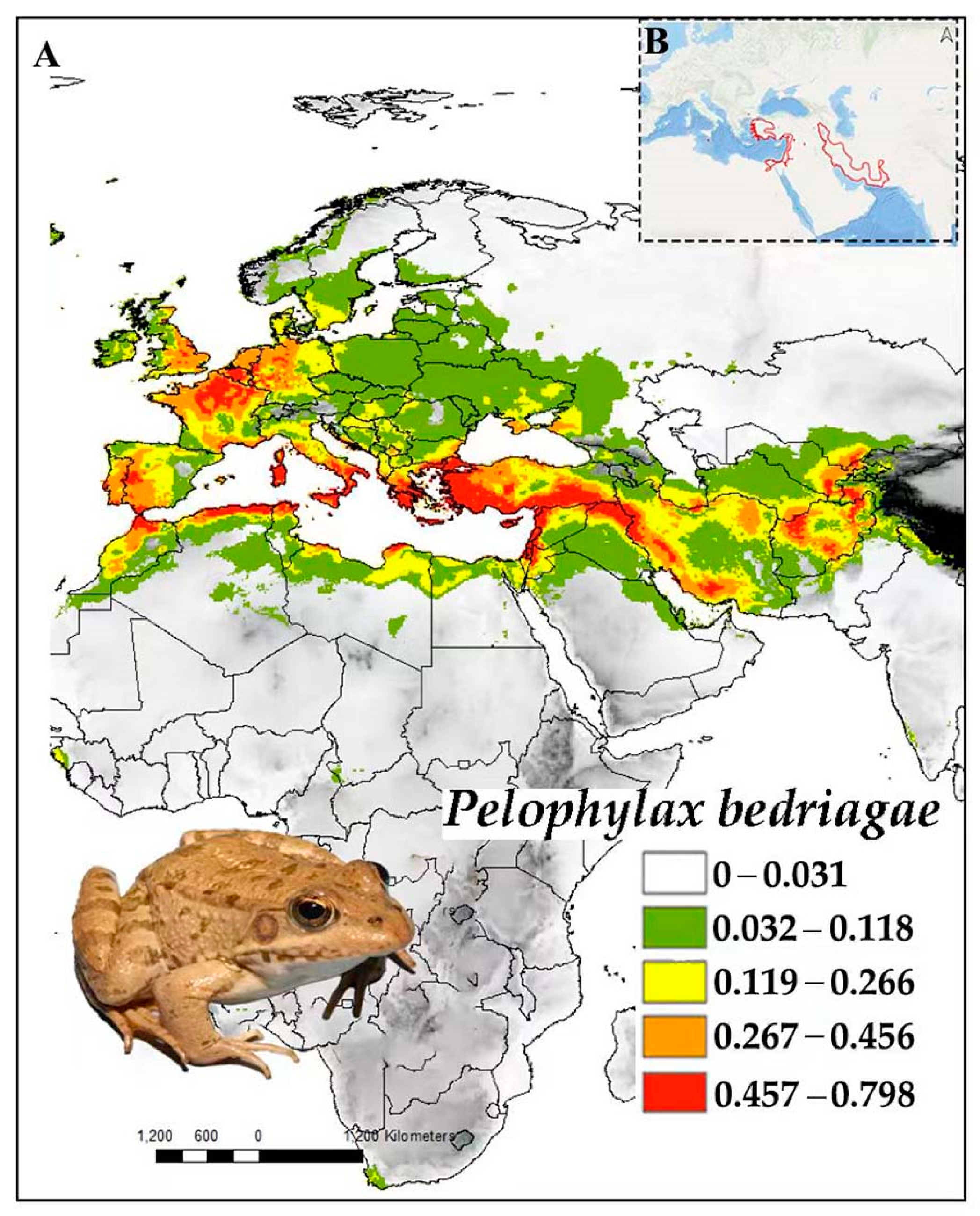{kind=link}
{kind=link}
{kind=link}
Table 1.
Percentage of contribution for each of the bioclimatic variables from the WorldClim 2.1 database used in the ecological models for each of the focal Pelophylax lineages.
Table 1.
Percentage of contribution for each of the bioclimatic variables from the WorldClim 2.1 database used in the ecological models for each of the focal Pelophylax lineages.
| Variables | P. ridibundus | P. bedriagae | P. terentievi | Pelophylax “Syr Darya lineage” | P. n. “reinhardtii” | P. n. “nigromaculatus” | P. porosus | P. mongolius | P. plancyi | P. chosenicus | P. hubeiensis | P. fukienensis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Elevation | 0.6 | 0.8 | 0.9 | 2.2 | 18 | 2.74 | 1 | 59 | 11 | 2.9 | 4.8 | 60 |
| Bio 1 | 5.2 | 3.1 | 17 | 0.1 | 21 | 10.2 | 12 | 2.9 | 2.87 | 3 | 28 | 1 |
| Bio 2 | 4.4 | 1.6 | 8.2 | 11 | 1.5 | 0.6 | 1 | 0.3 | 8.8 | 8.7 | 12 | 0 |
| Bio 3 | 1 | 10.4 | 1.4 | 5.5 | 11 | 0.1 | 3 | 6 | 34.3 | 40 | 25 | 7 |
| Bio 5 | 1 | 5.8 | 0 | 1.7 | 16 | 2.4 | 0 | 2.3 | 1 | 0 | 0 | 2 |
| Bio 8 | 17 | 17.2 | 9 | 0.5 | 1 | 1.1 | 1 | 1 | 14 | 19 | 13 | 1 |
| Bio 9 | 6.1 | 27.8 | 1.3 | 4.9 | 0.6 | 0 | 2 | 0.2 | 1.3 | 1.1 | 1.3 | 0 |
| Bio 11 | 12.8 | 0.1 | 3.6 | 4.3 | 0 | 0 | 1 | 14 | 0.4 | 0.9 | 5.1 | 20 |
| Bio 12 | 4.8 | 12.6 | 10 | 14 | 6 | 0.2 | 3 | 2.9 | 3 | 0.2 | 3.9 | 2 |
| Bio 14 | 39 | 0.3 | 1.1 | 0.9 | 0.7 | 0.1 | 2 | 6 | 18.4 | 23 | 22 | 5 |
| Bio 15 | 1.2 | 1.2 | 10 | 1 | 5 | 16.1 | 12 | 2.1 | 2.63 | 0.2 | 1.2 | 1 |
| Bio 18 | 0.6 | 1.3 | 37 | 9.2 | 0 | 0 | 0 | 0.2 | 0 | 1.3 | 1.5 | 0 |
| Bio 19 | 1 | 1.7 | 0.2 | 34 | 20 | 65.2 | 60 | 0.6 | 1.9 | 0.3 | 1.5 | 1 |
| AUC | 0.992 | 0.985 | 0.994 | 0.976 | 0.895 | 0.953 | 0.997 | 0.98 | 0.99 | 0.997 | 0.981 | 0.997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Amin, H.; Borzée, A. Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax. Diversity 2024, 16, 259. https://doi.org/10.3390/d16050259
AMA Style
Amin H, Borzée A. Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax. Diversity. 2024; 16(5):259. https://doi.org/10.3390/d16050259
Chicago/Turabian StyleAmin, Hina, and Amaël Borzée. 2024. "Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax" Diversity 16, no. 5: 259. https://doi.org/10.3390/d16050259
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.