Extinction Resilience of Island Species: An Amphibian Case and a Predictive Model

1
Department Philosophy & Social Work, Universitat de les Illes Balears, 07122 Palma, Balearic Islands, Spain
2
Laboratori de Natura, Museu de Ciències Naturals de Barcelona, Passeig Picasso s/n, Barcelona 08003, Catalonia, Spain
Diversity 2014, 6(1), 43-71; https://doi.org/10.3390/d6010043
Submission received: 21 November 2013
/
Revised: 4 December 2013
/
Accepted: 4 December 2013
/
Published: 8 January 2014
(This article belongs to the Special Issue Biogeography and Biodiversity Conservation)
Abstract
:Extreme overall divergence and high extinction rates are typical of insular endemics. Thus, detecting and understanding nativeness is critical on islands. Resilience to extinction is explored through a mechanistic approach focusing on midwife toads (Anura: Alytidae: Alytinae), an ancient lineage that includes continental and insular species. All alytines need urgent conservation action, including control of emerging diseases and spatially explicit reserve design aimed at ensuring ecosystem health and connectivity. The only extant insular alytine is additionally affected by an introduced continental predator. This alien species acts as a driver of the prey’s near-extinction and has not elicited any evolutionary response. Both IUCN criteria and EDGE scores show that alytines are top conservation priorities. However, there is a need for also considering phenotypic and ecological uniqueness in the assessment of conservation status and urgency. The reason is that phenotypes render ecosystems functional and insular ones uniquely so. In contrast, phylogenetic relatedness is just a constraint upon, not a motor of, evolutionary novelty. Insular species are indeed particularly susceptible, but can be similarly endangered as continental ones. This paradox may be solved by recognizing the insularity syndrome in any isolated or nearly-insular ecosystem, as a function of evolutionary and dispersal potentials. This predictive model may be useful for island biogeography, invasion biology and conservation planning.
1. Introduction
Holocene extinctions have largely involved insular species, yet the underlying causes remain unclear. An important reason lies in the area affected, with two consequences. First, islands are by definition much smaller than continents, providing a smaller surface area, as well as a smaller realized niche space. Therefore, island endemics have, on average, very small ranges. Thus, any deleterious impact can extend over a whole species’ habitat on an island much more easily than on a continental area [1]. In addition, islands are numerous. Each island harbors an impoverished biota; this is shown by the empirical relationship between species richness and area that is fundamental to island biogeography. However, insular species worldwide account for a major part of global biodiversity. Indeed, prior to the wave of extinctions caused by humans, up to a fourth of mammals and a third of birds lived only on islands [2,3,4,5]. Aside from quantitative factors, the question remains of whether insular species are intrinsically prone to human-induced extinction.
It has long been widely acknowledged that many island dwellers evolved in unique ways in the absence of natural enemies. This lack of behavioral, physiological or morphological adaptations on predator avoidance has adaptive value in isolation; it represents a clear disadvantage when confronted with continental species, including humans [6,7,8]. Species evolving on continents have been subject to various selective pressures, because species distributions and whole biomes experience large shifts in relatively short periods. Thus, from a strictly continental or world-ocean perspective, it may be easy to relativize or even dismiss nativeness [9,10]. As a consequence, the role of any insularity syndrome has been also dismissed as a minor contribution to extinction risk and species endangerments [1,11,12]. This may sound counterintuitive to researchers acquainted with the depauperate and disarmed nature of insular ecosystems, yet it stems from a lack of convincing documentation. For example, it is now clear that biological invasions often impact native species at a high rate and with dire consequences; their impact can be catastrophic on islands [12,13,14,15,16]. Nevertheless, there are surprisingly few studies demonstrating a direct, driving effect of the invasive enemies on native species survival (but, see [17,18]). In contrast, some reports point to fast adaptation by native prey to novel predators [19,20]. Given such contradictory claims, it seems reasonable to address the underlying mechanistic explanations in a taxon-specific approach. This is a necessary priority for understanding general biogeographic processes [21,22,23] and human-induced extinctions [5], as well as for implementing sound conservation action [8]. Thus, focusing on a comparatively species-poor, well-studied taxon might shed some light on the subject. However, stressing the differences among lineages [24] should not deter one from searching general patterns across taxa or habitats [25,26]. From this perspective, aiming at a general explanation of extinction resilience or susceptibility of island species seems desirable.
Herein, I addressed the research need for both focus and general applicability, selecting a higher-level taxon that contains both insular and continental species with various range sizes. On this basis, I developed a model to assess the insularity syndrome. The taxon selected with these criteria is the Alytinae, or midwife toads. These amphibians are found only in south-western Europe and north-western Africa, plus an endemic on the island of Mallorca in between. They are largely terrestrial and best known for their extraordinary parental behavior, the male carrying the eggs at all times until they hatch and the tadpoles are released in pools (Figure 1).
Figure 1.
Continental and insular alytines. (A) The European midwife toad, Alytes (A.) obstetricans. This is a male carrying its developing offspring (Salt del Sallent, Catalan Pyrenees). (B) The ferreret, Alytes (Baleaphryne) muletensis, is the only extant insular alytine, now confined to a few inaccessible mountain canyons in the island of Mallorca. Notice its long limbs, the papilla-like endings of the digits and the smooth skin. (Alcanella, Serra de Tramuntana, Mallorca).
Figure 1.
Continental and insular alytines. (A) The European midwife toad, Alytes (A.) obstetricans. This is a male carrying its developing offspring (Salt del Sallent, Catalan Pyrenees). (B) The ferreret, Alytes (Baleaphryne) muletensis, is the only extant insular alytine, now confined to a few inaccessible mountain canyons in the island of Mallorca. Notice its long limbs, the papilla-like endings of the digits and the smooth skin. (Alcanella, Serra de Tramuntana, Mallorca).

Alytines constitute a distinct, ancient lineage (Figure 2). Extant alytines are usually classified within the genus, Alytes Wagler, 1830, comprising three (sub)genus-level monophyletic lineages [27,28]. The sister group of Alytinae is the Discoglossinae, which diverged at 113 Ma; it includes seven extant species of painted frogs, all in the Mediterranean region [29,30,31,32]. Together with the European and Asian Bombinatoridae fire-bellied toads (which diverged at 157 Ma and with 10 living species), they constitute the Costata. This is a lineage that split from almost all other anurans (except the basal Leiopelmatidae) at 171 Ma [30,31,32,33,34,35,36,37].
Alytines include five extant species, differing considerably in range (Figure 2) and habitat. All alytines are threatened by habitat loss and emerging infective diseases, as is the case for many anurans worldwide [38,39,40,41,42]. The European midwife toad, Alytes (A.) obstetricans Laurenti, 1768, is adapted to walking (diagonal quadrupedal limb movement, in addition to typical anuran saltatory locomotion) and has a wide distribution; in Iberia, it is represented by four subspecies, three of them endemic [43,44]. In the IUCN Red List, it is currently listed as least concern [45]. The remaining four alytine species are restricted to the extreme south-western Palearctic, an area experiencing an increase in aridity and large-scale habitat loss for amphibians [46]. Two species are Iberian endemics: the Iberian midwife toad, A. (Ammoryctis) cisternasii Boscá, 1879, and the Betic midwife toad, A. (A.) dickhilleni Arntzen and García-París, 1995. The former is a specialist of Mediterranean-type oak forests and scrub on sandy soils, where it burrows actively [47]; it is listed as near threatened [48]. The latter is restricted to a few mountain ranges [43,49] and is listed as vulnerable [50]. The sole African alytine is the Moroccan midwife toad, A. (A.) maurus Pasteur and Bons, 1962. This is also a mountain endemic, only recently recognized as a distinct species [51,52,53,54]; it is listed as near threatened [55,56]. Finally, the Mallorcan midwife toad or “ferreret”, A. (Baleaphryne) muletensis (Sanchiz and Adrover, 1979), is adapted for climbing. It is currently listed as vulnerable [57], being restricted to a handful of deep mountain canyons in northern Mallorca. However, until the 1st century BC, it occurred throughout the island. It was also present in neighboring Minorca, where it may have constituted a distinct species [58] that went extinct due to the lack of mountain refuges. Ferrerets evolved essentially without predators; this may account for their unusually smooth skin without odor or poison glands, and clutches consisting of up to twelve exceptionally large eggs. The introduction of snakes by Roman colonists, particularly the viperine water snake, Natrix maura (Linnaeus, 1758), is often considered as the cause of the ferreret’s near-extinction [2,59,60,61,62]. However, it has also been claimed that this native prey acquired heritable adaptations in the form of behavioral and developmental mechanisms of specific predator avoidance in an unusually short time span [63,64]; such adaptive responses from native prey species are unique outcomes of biological invasions [20]. Indeed, abruptly changing a selective regime that lasted 14 million years [2,28] represents a unique ecological experiment [26,65].
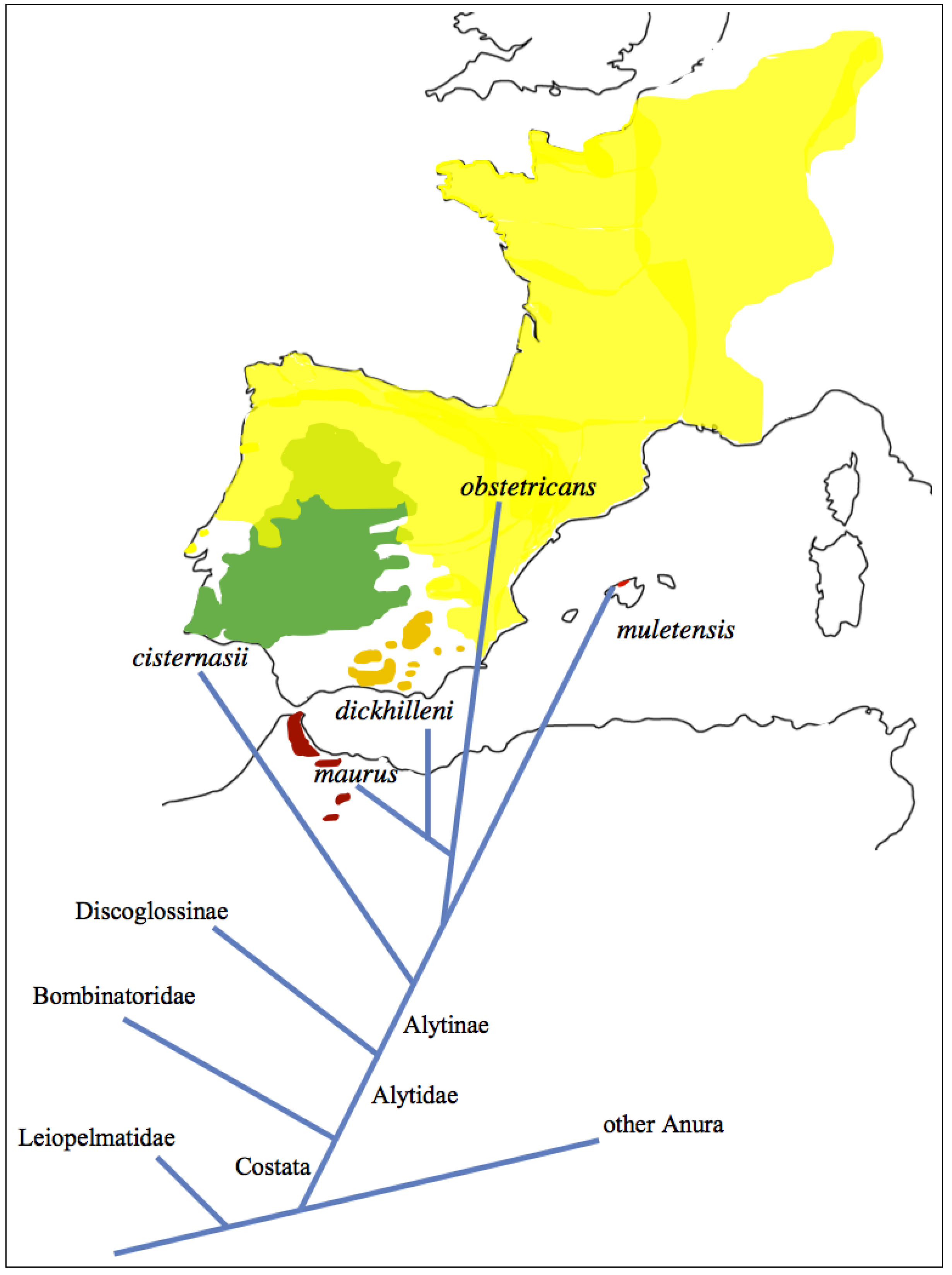
Figure 2.
Distribution and phylogenetic relationships of Midwife toads. Alytes (A.) obstetricans ranges from northern and central Iberia to north-central Germany. The closely related A. (A.) dickhilleni and A. (A.) maurus live only in small mountain areas in extreme south-eastern Iberia and northern Morocco, respectively. Alytes (Ammoryctis) cisternasii is restricted to south-western Iberia. Alytes (Baleaphryne) muletensis is the only extant insular alytine.
Figure 2.
Distribution and phylogenetic relationships of Midwife toads. Alytes (A.) obstetricans ranges from northern and central Iberia to north-central Germany. The closely related A. (A.) dickhilleni and A. (A.) maurus live only in small mountain areas in extreme south-eastern Iberia and northern Morocco, respectively. Alytes (Ammoryctis) cisternasii is restricted to south-western Iberia. Alytes (Baleaphryne) muletensis is the only extant insular alytine.
In this paper, I identify the threats currently and historically affecting the various alytine species, with the aim of assessing their conservation status and priority. I then place the results in a general context in search of any fundamental differences in extinction resilience between insular and continental species.
2. Experimental Section
In order to assess the conservation status of alytines, I proceeded stepwise. First, I gathered my own field observations (including anecdotal evidence) on the presence/absence, abundance and habitats actually occupied, especially relative to A. muletensis, during the last 25 years. These data span from specific searches for alytines, where individuals were searched for and counted, to unconstrained random walks through potentially favorable habitats. I also included familiarity with captive breeding.
A literature review was performed exhaustively through searches in journals, library databases (including Revistes Catalanes amb Accés Obert, RACO) and search engines (Google Scholar, Scopus), as well as in CREAIB (Centre de Recursos per a l’Educació Ambiental de les Illes Balears). In order to validate published results, I assessed the adequacy, accuracy, repeatability and logic of experimental methods, data analyses and conclusions. In particular, I wanted to know how the area of occupancy of each species has declined, whether census data are credible and if evaluations of known threats affecting alytines are evidence-based. Three issues were found to be of special interest to the latter point: (1) the evidence for the reported behavioral and developmental response by ferrerets to introduced snakes was re-evaluated; (2) available reports of chytridiomycosis among alytines were used to assess the relevance of this emerging disease; and (3) predictions of regional climate change were used to outline the expected increase in intensity of detected threats, taking into account both life history and expected habitat change in the inhabited landscape [66]. I verified my data and conclusions through the consultation of experts (S. Pinya, A. Román, G. Fernández, W. Beukema).
These results were used to quantify the threatened status of the various alytine species. This was done under two schemes. First, I applied the IUCN Red List criteria [67,68], taking into account the distinction between the area of occupancy (the sum of areas actually occupied by a species) and the extent of occurrence (the area of a convex polygon encompassing all sites where a species is present). These ranks must be supported by quantitative evidence or inferences clearly made from the available evidence, with the aim of leaving little room for subjectivity [69]. Published and reassessed rankings were used to compare variation in the Red List Index (RLI), aimed to track the trend of overall extinction risk [70,71,72].
In order to assess the conservation priority of alytines, taking into account their phylogenetic uniqueness, I also calculated evolutionary distinctiveness (ED) and EDGE scores [73]. ED is the sum of branch lengths (L, in million years between adjacent nodes) leading to a particular species divided each by the number of species (S) it includes:
and:
where GE (global endangerment) is the species’ global conservation IUCN status in a scale from one to four: least concern, vulnerable, endangered and critically endangered. This method divides the total phylogenetic diversity among its members, taking into account both the number of species in each branch and its length in time. In addition, it considers the threatened status of each species. Thus, EDGE scores are equivalent to a log-transformation of the species-specific expected loss of evolutionary history in which each increment of the Red List category represents a doubling of extinction risk [73]. The cladogram used is robust, derived from morphology, allozymes and nuclear and mitochondrial DNA sequences; it is dated with well-documented paleogeographic events ([28], Altaba submitted). The basal position of the Costata relative to most other anurans differs slightly among cladograms inferred for all amphibians [35,36,37,74]. At any rate, their sister group invariably includes virtually all living anurans. Thus, such conflict has no appreciable influence on the EDGE scores, which become asymptotic when the clade reaches ca. 100 species [73].
ED = Σ L/S
EDGE = ln(1 + ED) + GE ln(2)
There is a need for another measure of extinction risk. Red List ranks are purely phenomenological; even though successive RLI’s may document trends in conservation status, such ranks constitute mere descriptors of current reality. This is valuable, of course, and fulfils the motivation for which they were designed. In contrast, EDGE scores relate to the past, as they measure the historical legacy at risk, accounting for the phylogenetic uniqueness of endangered species. A third criterion can be devised to predict future outcomes, introducing evolutionary ecology. Insularity should then be understood in its ecological, not merely geographic, meaning. Its impact on a species’ traits can be labeled as insularity syndrome (I).
I is a function of two variables: dispersal and evolutionary potentials. The latter (E) is the magnitude of a vector in the space defined by the inverse of generation time, the inverse of effective population size and population structure. These parameters may often be very difficult to assess, but they can be replaced by proxies. In the case of alytines, the duration of the larval stage (l, in years per individual) can be substituted for generation time; fecundity (f) per male per year can replace population size, and a rank of relative range fragmentation (φ, from 1 to 4) can account for population subdivision. Dispersal potential (D) for a particular taxon in a given context is the magnitude of a vector in the space defined by the dispersal probability at the appropriate spatial and temporal scale, range size and taxonomic diversity of its native community. These can also be approximated by the ratio of the largest to the minimum (effectively a unit in most animals) distance covered by one individual in one year (d, in meters to meters), the species’ extent of occurrence (o, in Km2) and a rank of community diversity in one Km2 (δ, from 1 to 4), respectively. Using logarithmic transformations for scaling and keeping errors additive, three dimensionless variables result:
E = l−1 (1/log(f + 1)) φ
D = log(d) log(o) δ
I = f(E,D)
I developed a model of I in a four-step process. First, I drew the shape of this function intuitively as a three-dimensional graph. Then, I assessed the position in this heuristic graph of the five alytines and 13 other species, representing a wide array of continental and insular cases. Approximate values for E and D were calculated as explained above (except that fecundity of non-alytines was calculated per female). This empirical plot made sense, so I searched for an analytical expression of I.
At very low values of D, insular evolution should prevail, while at large values of D, the forces shaping strongly continental species must rule. Thus, I can be written as the addition of two terms:
I = f(E) P(D) + f’(E) D2
The first term accounts for insularity, being the product of evolutionary change and a measure of isolation. This measure is a Poisson distribution of D, meaning chance dispersal into the system. On average, colonization of islands probably occurs only once, so the mean (λ) can be set to 1. The second term is the product of the rate of evolutionary change and dispersal potential in two dimensions (i.e., geographically).
The shape of f(E) can be approximated by a Gompertz function, a sigmoid curve that accurately describes biological growth. It emerges as a result of antagonistic processes [75,76], in this case, natural selection leading to specialization and the inertia of phylogenetic constraint. It can be written as:
![Diversity 06 00043 i001]() where a is the asymptote, b is a response delay and c is acceleration. Its derivative is:
where a is the asymptote, b is a response delay and c is acceleration. Its derivative is:
![Diversity 06 00043 i002]()
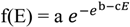
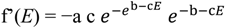
For the sake of scaling and elegance, let us set a = 1, b = 2 and c = 0.2, and let us introduce a constant for each term of I (2 and 0.05, respectively). Thus:
![Diversity 06 00043 i003]()
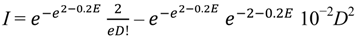
3. Results
3.1. Overall Threats Recognized
All five alytine species have experienced strong declines in the last decade (Table 1). The Red List Index of species survival for alytines dropped from 0.76 in the current Red List (published in 2011) to 0.44, as assessed herein. Even the species with the largest range (A. obstetricans) faces a variety of relevant threats [77,78,79,80,81,82]. Widespread destruction of breeding sites and landscape transformation have caused many local extinctions [49,77,78,79,80,81]. In addition, the combined action of emerging disease and climate change appears to be responsible for decimating populations, also in protected areas [42,82]. A. cisternasii is declining markedly throughout, due to habitat destruction, urban and industrial pollution and introduced predators, such as fish and crayfish [83,84]. Large expanses of suitable habitat for this species have been destroyed; over half of breeding sites were lost in less than a decade, largely due to draining of breeding pools, flooding of large areas by impoundment and stark changes in land use towards urban and industrial landscapes [47]. The highly fragmented range of A. dickhilleni suffers widespread habitat destruction, basically caused by water diversion to human uses, pollution and landscape transformation [49,85,86,87,88,89]. A. maurus lives in isolated sites that are encroached upon by human water uses and subject to increasing pollution from urban waste. It is also affected by introduced predators, such as mosquitofish. Although it is still reasonably common in some localities, it has disappeared from part of its former range [54]. Only one population was recently found with calling activity; larvae were either rare or absent at another six sites where the species had been recorded a decade earlier [90]. A. muletensis has experienced a precipitous decline [41,91,92,93,94,95,96]. Increasing water demands already claimed much of its extremely restricted and fragmented habitat in the last half century [91,96]. Together with climate change [97,98], water extraction increases the severity of periodic drought. The last wild populations are subject to effectively uncontrolled (and officially encouraged) canyoning. This is a marginal recreational activity that can exert significant pressure on small freshwater ecosystems [99]. Moreover, it directly causes intense disturbance, serious injuries to adults and larvae and spreads chytridiomycosis. Translocated populations have had a limited success; they are also affected by diffuse pollution, apparently causing abnormal development at an isolated site [100].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Threats, declines within last decade and updated conservation status with supporting IUCN criteria for the five extant alytines. Threats are range fragmentation (1), total population size (2), habitat destruction (3), water extraction (4), landscape change (5), introduced predators (6), chytridiomycosis (7), diffuse pollution (8) and climate change (9). The severity of threats and declines is given as a rank, from 0 (not appreciable) to 3 (very important). Projections for the next decade are in parentheses.
| species | threats | Δ− | status | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |||
| obstetricans | 1 | 0 | 2 | 1 | 2 | 0 | 2 | 1 | 1 | 2 | VU A2ace + 3ce |
| (2) | (1) | (1) | (2) | (2) | (0) | (3) | (1) | (2) | (2) | ||
| cisternasii | 1 | 0 | 2 | 2 | 2 | 1 | 0 | 1 | 0 | 2 | VU A4ace |
| (2) | (1) | (3) | (2) | (2) | (1) | (3) | (1) | (2) | (2) | ||
| dickhilleni | 2 | 2 | 3 | 2 | 2 | 1 | 1 | 1 | 1 | 2 | EN A4ace; B1ab(i,ii,iii,iv) + 2ab(i,ii,iii,iv) |
| (3) | (3) | (2) | (3) | (2) | (1) | (3) | (1) | (2) | (3) | ||
| maurus | 2 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 2 | EN A2abce; B1ab(i,ii,iii,iv,v) + 2ab(i,ii,iii,iv,v) |
| (3) | (3) | (2) | (3) | (2) | (1) | (3) | (2) | (2) | (3) | ||
| muletensis | 3 | 3 | 2 | 2 | 1 | 3 | 2 | 1 | 1 | 2 | CR B1ab(i,ii,iii,iv) + 2ab(i,ii,iii,iv) |
| (3) | (3) | (3) | (3) | (1) | (3) | (3) | (1) | (2) | (3) | ||
Climate projections for the next century involve a drastic increase in temperature in the Western Mediterranean region; reduced precipitation and falling ground water levels will cause freshwater availability to decrease markedly [97,101]. This, in turn, should represent the massive loss of surface water that provides breeding habitats and refuge for amphibians [46]. Nevertheless, there is a need for case-specific models [66]. In the case of A. maurus, an important range reduction can be expected from desertification by human-induced changes in the landscape hydrology and climate change. In other instances, it is difficult to make a forecast. Ferreret larvae are unique in that they do not overwinter and metamorphosis takes place in summer. Thus, occasional drying of pools already accounts for noticeable population declines in some canyons [92]. However, decreasing rainfall in the Mallorcan mountains may be coupled with an increase in the frequency of extreme events, such as severe thunderstorms [98,102]. Such sporadic events might allow deep canyon pools to persist. Even so, perennial or nearly perennial pools may harbor an increased density of predators, such as waterbugs and dragonfly larvae.
Chytridiomycosis (Batrachochytrium dendrobatidis), a globally-prevalent parasitic fungus driving amphibian decline [38,39,40,41,42,103,104,105,106,107], has imperiled the Alytinae throughout its range with very high susceptibility and high to extreme suitability [39,108]. A warming climate aggravates pathogenesis and the rate of contraction [109]. A. obstetricans has already been extirpated from montane areas, probably through the synergistic effects of increased environmental temperature and infection by the chytrid fungus [42,82]. The ruling factor, though, appears to be the pathogen’s spread, with environmental changes being a contributing factor [38]. A. cisternasii is likely to suffer heavily from chytridiomycosis [84]. The chytrid fungus has recently appeared in at least two separate populations of A. dickhilleni, causing 70% mortality [89]; it has already reached the range of A. maurus [110]. Translocations to and from zoos [111,112] brought the epidemic into the remote localities inhabited by A. muletensis [41,94], causing ongoing local extinctions. Pollution from sewage, agricultural and road runoff might increase virulence, because high concentrations of nitrate and chloride are facilitators [113]; so does low temperature [114,115], a condition that prevails in the alytine range.
A threat affecting the ferreret in particular is a translocated continental species. The introduction of predatory fish, snakes, frogs and crayfish into breeding pools has been reported to be responsible for local declines or extirpation among alytines [77,83,84,90,91,92]. However, indirect evidence suggests that predation by viperine water snakes was the driver of the ferreret’s near-extinction. Predator and prey do not occur together, and this prey is comparatively slow and cumbersome. Indeed, a single snake will eat all ferrerets in a pool [93,95]. Besides, the temporal pattern of its near-complete disappearance coincides with the arrival and spread of the alien predator. Eventually, no other putative cause can explain the ferreret’s historical decline and current marginal refuge [2,59,60,61,62].
3.2. EDGE Analysis and Model Predictions
The highest EDGE value belongs to A. muletensis (Table 2). However, all alytines are top priorities for preserving amphibian biodiversity. The E, D and approximate I values for alytines (Table 3) show that continental species have small negative I scores, even those with large distributions. Instead, the ferreret shows a high positive value, accounting for its strongly insular character.
Table 2.
Evolutionary distinctness (the phylogenetic measure provided by ED), conservation status (as GE, the ranked IUCN Red List categories) and composite EDGE values for the five extant species in Alytinae.
| species | ED | GE | EDGE |
|---|---|---|---|
| muletensis | 47.83 | 4 | 6.66 |
| dickhilleni | 43.06 | 3 | 5.86 |
| maurus | 43.06 | 3 | 5.86 |
| cisternasii | 50.83 | 2 | 5.33 |
| obstetricans | 45.16 | 2 | 5.22 |
Table 3.
Evolutionary potential (E), dispersal potential (D) and insularity syndrome (I) of alytines. Variables are the duration of larval stage (l), fecundity (f), range fragmentation (φ), individual dispersal (d), extent of occurrence (o) and community diversity (δ).
| Species | l | f | φ | E | d | o | δ | D | I |
|---|---|---|---|---|---|---|---|---|---|
| muletensis | 0.4 | 10 | 4 | 19.20 | 70 | 250 | 1 | 0.22 | 0.313 |
| dickhilleni | 1.3 | 60 | 3 | 2.58 | 1,000 | 10,000 | 2 | 1.20 | −0.011 |
| maurus | 1.3 | 150 | 2 | 1.42 | 1,000 | 10,000 | 2 | 1.20 | −0.005 |
| cisternasii | 0.3 | 100 | 2 | 6.66 | 100 | 140,000 | 3 | 1.54 | −0.003 |
| obstetricans | 1.7 | 200 | 1 | 1.02 | 1,500 | 800,000 | 4 | 3.75 | −0.003 |
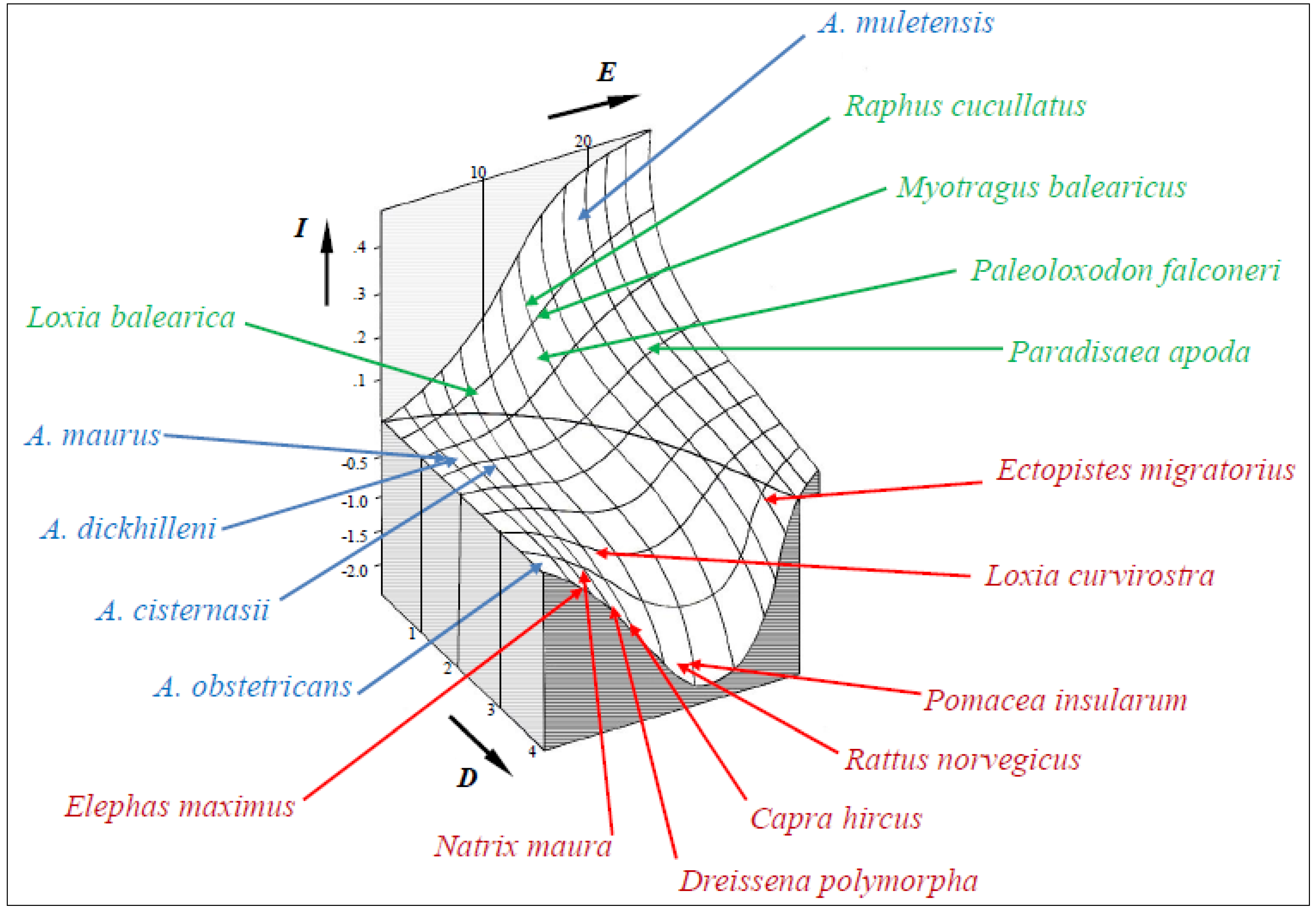
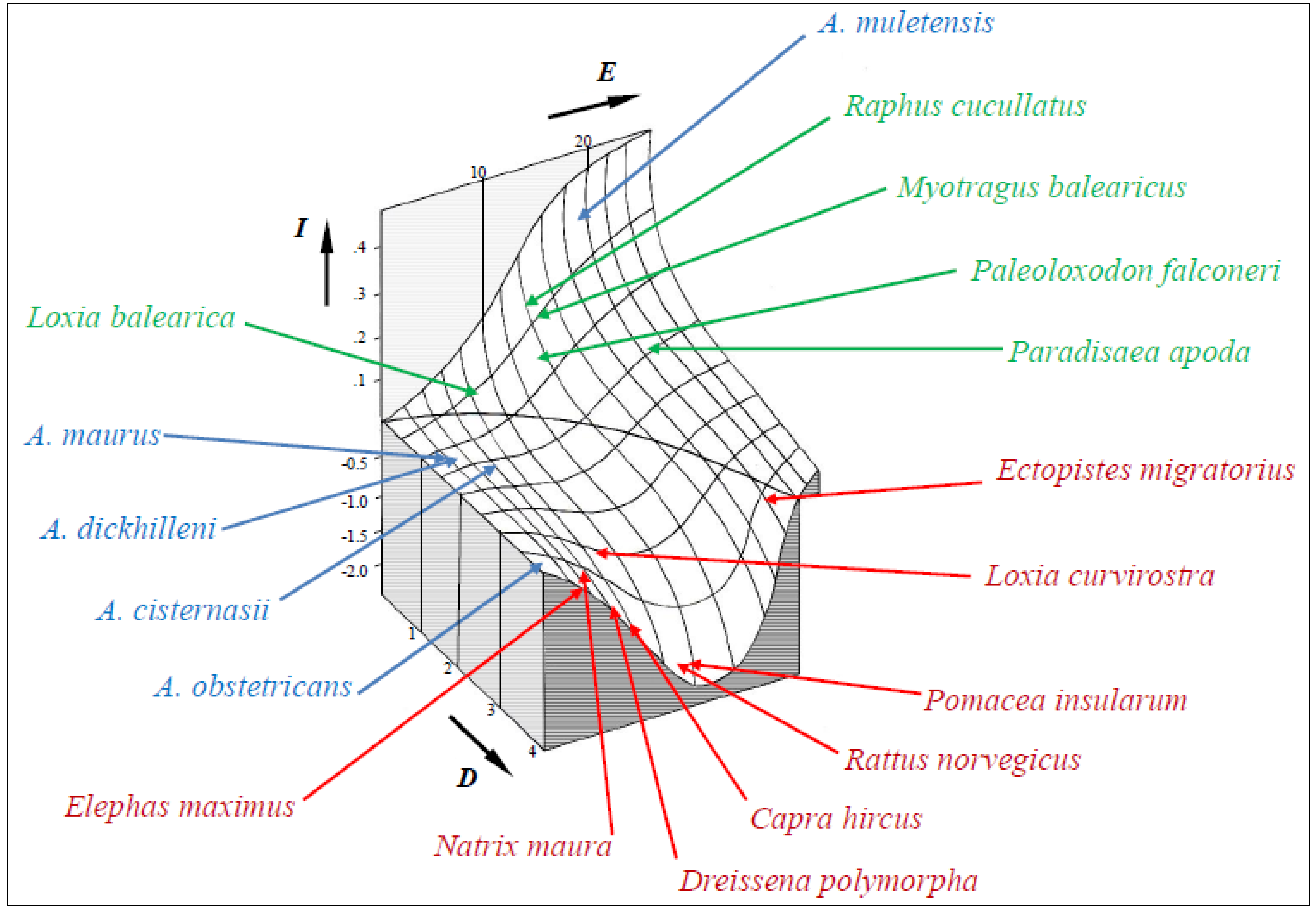
The model developed to predict insularity was validated, as only minimal corrections were needed when I scores for all species analyzed were introduced into the initial plot (Figure 3). Negative values represent continentality. At low values of D, a sigmoid depicts the increase of I with E. At high values of D, a bell-shaped valley shows strong negative values of I for intermediate values of E. The intersection of the curved surface with the E-D plane marks the border between (ecologically) insular and continental settings. Thus, it accounts for what sets island dwellers apart: living and evolving in isolation. The vector space determined by E, D and I constitutes a predictive model of the ecological consequences of insularity, beyond the tangible constraints of time and space. It may provide a conceptual framework for examining the relationship between ecological setting and extinction resilience.
The position of the species assessed on the I graph shows some interesting patterns. Insular and continental species appear on separate sectors, although there is considerable variation in scores. Three strongly insular species (Paleoloxodon falconeri, Myotragus balearicus and Raphus cucullatus) went extinct in the Holocene as a consequence of interaction with continental fauna. This is the main factor causing the near-extinction of A. muletensis. In contrast, recently diverged taxa on islands, such as Loxia balearica [116], exhibit a modest insularity syndrome. Continentality is also far from uniform. Large I negative scores belong to highly invasive species (Pomacea insularum, Rattus norvegicus, Capra hircus and Dreissena polymorpha), whose arrival into isolated ecosystems can readily have disastrous consequences for strongly insular species. However, the threat posed by invasive species may be better predicted by the differential along the I axis relative to the affected insular species. In the case of the ferreret, its large I score makes it highly vulnerable to a continental predator, such as Natrix maura.
Figure 3.
A predictive model of the insularity syndrome (I), as a function of evolutionary potential (E) and dispersal potential (D). Zero values follow a parabola separating insular and continental species. The positive range of the vertical axis is magnified five times for clarity. The position of various animal species is indicated on the surface chart; alytines in blue, insular species in green (from left to right, Mallorcan crossbill, Mauritian dodo, Balearic rat-goat, Sicilian dwarf elephant and New Guinean greater bird of paradise) and continental species in red (Asian elephant, viperine water snake, zebra mussel, feral goat, black rat, large apple snail, red crossbill and passenger pigeon).
Figure 3.
A predictive model of the insularity syndrome (I), as a function of evolutionary potential (E) and dispersal potential (D). Zero values follow a parabola separating insular and continental species. The positive range of the vertical axis is magnified five times for clarity. The position of various animal species is indicated on the surface chart; alytines in blue, insular species in green (from left to right, Mallorcan crossbill, Mauritian dodo, Balearic rat-goat, Sicilian dwarf elephant and New Guinean greater bird of paradise) and continental species in red (Asian elephant, viperine water snake, zebra mussel, feral goat, black rat, large apple snail, red crossbill and passenger pigeon).
4. Discussion
4.1. Conservation Status
All alytines are considerably more threatened than hitherto recognized. The two most widespread species are vulnerable, in contrast with their current listing as least concern (A. obstetricans) and near threatened (A. cisternasii). The two restricted-range continental species are endangered, but are currently listed as near threatened (A. maurus) and vulnerable (A. dickhilleni). The insular ferreret (A. muletensis) is critically endangered; it is listed as just vulnerable. Thus, the Red List Index of species survival decreased 42%. However, this large drop in just two years cannot be attributed to actual changes in status, but to the inaccurate or overly optimistic nature of previous rankings.
Likewise, the reassessed EDGE values are considerably higher than those reported in a listing of 4,340 anuran species [117]: A. muletensis ranks third of all anurans; both A. dickhilleni and A. maurus are tied in the 19th position; A. cisternasii is 96th; and A. obstetricans is in 134th place. A comparison with mammal species [73] highlights the threatened status of the Alytinae as a whole: all alytines would fall within the top 100 mammal species. Indeed, both A. maurus and A. dickhilleni would be placed among the top 25 mammals; A. muletensis has an EDGE score higher than the most endangered mammal.
It is relevant to point out that most of the alytines’ closest relatives (discoglossines and bombinatorids), together representing a major component of global amphibian biodiversity, are also endangered (e.g., [32,118,119]). Nevertheless, the alytine rankings could be even higher, given that there are several problems with the published evidence.
4.2. Limitations in the Existing Studies and Knowledgebase
Evidence for the impact of exotic predators is limited to declines in the local density of alytines, often without direct observations. In fact, there is a lack of solid experimental evidence on local population decline and its causes. As a case in point, predator-induced evolution has been suggested in A. muletensis, but on the basis of flawed experimental work. The field experiments involved comparison of tadpole behavior in pools without snakes or with four snakes bagged in a nylon mesh bag [63]. In addition, tadpole development was compared among pools with either three adult viperine snakes or stones placed inside nylon mesh bags [64]. In the laboratory, tadpole development was compared among plastic boxes having either clean or snake-conditioned water [64]. Other laboratory tests involved observation of tadpole behavior in circular pans with flowing clean or snake-conditioned water. In this case, the snakes had been captured in Mallorca and Iberia; two other amphibian-eating species of snakes were also used [63]. The reported differences among treatments were considered as evidence of a response to chemical cues from the recently invading predator. Thus, it was suggested that “it is reassuring that Mallorcan midwife toads have retained natural anti-predator behavior through several generations of captive breeding” [63]. Moreover, this led to the statement that “in certain areas plasticity in morphology may contribute to the coexistence of predator and prey” [64]. However, as shown below, the experimental design and data analysis are questionable.
In the laboratory, “snake-conditioned water” was obtained by placing either a snake in an unspecified volume during at least 2 h [63] or three netted adult snakes in just 20 L during 3 to 4 h [64]. Thus, any differences among treatments may be attributed to organic pollution by the snake’s excretions in otherwise clean spring water. Changes in behavior were reported as significant in pans with water conditioned by Natrix maura collected in Mallorca two months earlier. In contrast, no effect was observed when using mainland conspecifics (snakes captured in Iberia) or two other tadpole-eating snake species [63]. Yet, it is unclear how the recently captured snakes differed from the long-held ones in terms of predatory activity or ability to pollute water. Most importantly, that tadpoles were immobile most of the time implies that nearly a quarter of the ranked values of tadpole positioning were tied data. Such a large proportion of zero values makes it inappropriate to use the Wilcoxon–Mann–Whitney test to compare tadpole activity among treatments, as was done. Indeed, a highly skewed distribution and an increased number of ties leading to the inflation of zero values may easily lead to incorrect conclusions [120]. Overall, the laboratory experiments failed to support the proposed evolutionary response by ferrerets.
In the field experiments, artifacts were also relevant: pools with bagged snakes unavoidably experienced movements that could not exist in pools with bagged stones. The deep-canyon pools where ferrerets still survive harbor no other vertebrates, are inhabited only by a few small invertebrates and are only disturbed by rain spells. Thus, any differential behavior and development of tadpoles can be attributed to physical disturbance by the bagged snakes. This may account for differences in grazing and resting times. In addition, snakes surely enriched water with organic waste. Ferreret tadpoles exhibit considerable ecophenotypic and behavioral plasticity [121], so the recorded differences in development may be attributed to uncontrolled experimental artifacts. Such plasticity has been reported for many anurans breeding in a variety of aquatic habitats under pressure by a wide array of predators [122]. Indeed, phenotypic plasticity during larval development associated with unpredictable environments is widespread among amphibians. Likewise, the lower behavioral response of tadpoles living in pools that are reached by snakes suggests only habituation to disturbance. Although tadpoles of some species are able to learn predator-avoidance behaviors, even for recently contacted organisms [123,124,125,126,127,128], ferrerets appear to be totally naive.
All evidence points only to a pre-existing phenotypic plasticity that can be elicited by the novel predator’s behavior. In a complex spatial setting, such plasticity may allow escape from exotic predators and thus promote rapid evolutionary responses with ecosystem-wide impacts [129]. However, in the ferreret case, the necessary spatial and temporal window simply does not exist. Introduced snakes, as other non-native enemies for a variety of amphibians [130], are just one more predator that insular alytines cannot detect. There is no evidence of an evolutionary response to predatory snakes by A. muletensis; the possibility of coexistence of these two species can be discarded. Therefore, the ferreret’s present habitat is not a natural optimum in terms of survival and reproductive success (as suggested by [93]). Instead, it is just a serendipitous set of marginal refugia (as shown for European bison [131]). Eventually, the ferreret case adds evidence to the role of introduced predators as powerful drivers of extinction.
Also problematic are census data, being generally missing or unreliable. The conservation status of A. muletensis in particular appears to be underestimated in official reports (e.g., [57]). The claim of increases in population size [92] was solely based on counts of larvae. Such counts yielded fairly constant data over several years. There was no reliable measure of (admittedly large) observation error or (likely low) survival rates. This fictitious demographic recovery triggered the change in its recognized conservation status: from critically endangered to endangered and, eventually, to just vulnerable. When A. muletensis was discovered over 30 years ago, the total number of adults was less than 1,500 [95]; nowadays it is unlikely to exceed half that figure.
Data on the impact of chytridiomycosis are limited. The chytrid isolated in Mallorca already exhibits appreciable genetic and physiological differentiation, attributable to selection under local environmental conditions [107,132]. However, the consequences on the infection’s impact are unclear. In a similar way, the serious outbreaks among A. obstetricans in high-mountain areas in central Spain cannot be reasonably attributed to climate change alone (as tentatively suggested by [82]). Instead, they were probably initiated by the introduction of the chytrid, via contamination of clothes and footwear, or the release of infected amphibian pets. This human-mediated introduction probably took place in or around a small glacial lake that is easily accessible, close to the city of Madrid, and, thus, massively visited.
Finally, the impact of climate change has not been adequately assessed. For example, it is likely that breeding sites of A. dickhilleni will become scarcer. However, this is not the only factor leading to a greater endangerment (as stated in [88]).
4.3. Alytine Conservation
Without decisive conservation action, the ancient Alytinae will become greatly impoverished or even extinct in the next few decades. Two continental species have small, highly fragmented ranges that are reminiscent of those typical of island dwellers. Habitat loss and introduced enemies are critical issues for these two species. This may be the case for a majority of non-marine species living on continents, whose distributions are often ecologically and geographically limited. Even the two alytines with larger ranges have a deep phylogeographic structure [44,133], warranting an urgent (re)evaluation of the status of their constituent subunits (whether recognized as subspecies or not).
Active measures should involve the protection of extant habitats, a factor that is paramount for rapidly vanishing amphibians [134]. It must be stressed that such protection requires preservation of ecosystem health [103,135]. Indeed, all alytines are found within protected areas, but this does not always translate into effective protection. Reserve designing may benefit from gap analysis [136,137], taking into account the aquatic-terrestrial connectivity that amphibians need at a landscape level [138,139]. Identified threats should be quantitatively evaluated, in order to determine the suitability of landscape for long-term survival. This should include assessments of both the capacity of habitats to maintain viable populations and the adequacy of corridors for effective dispersal among populations [140,141,142]. The latter needs a careful identification of source populations [143]. Two other measures are critical: stringent control of introduced predators and a complete ban on harmful, superfluous activities, such as canyoning. In addition, understanding emerging diseases has become a priority.
Chytridiomycosis in alytines highlights the role of infectious disease in the decline of wild species [39,144]. It is critical to devise management strategies able to closely track this moving target [107,108]. Such monitoring needs to take into account both host and pathogen perspectives in an explicit spatial context [40,41,42]. This should include early detection, as well as strict control of translocations, if any. Although anti-fungal treatments may be successful for A. muletensis in captivity [145], prevention will only be possible through strict regulations on encroachment and management. A key factor appears to be curbing the amphibian trade [105,146], now largely unrestrained throughout the alytine range.
As a particular case of understanding the complexity of amphibian declines [42,147], it is imperative to keep a global vision of the enormous threats with which alytines are confronted. Lessons learned from this insular species could be applied to those with restricted ranges on the continent through relatively simple actions. These may include preventing encroachment, keeping exotic predators and emerging diseases away and adequate management of habitats occupied throughout their life history. This small-scale work will be useful for more ambitious plans directed at still widespread species. Indeed, habitat loss (including loss of breeding pools free of introduced enemies) constitutes a key factor for all alytines.
Conservation action cannot be implemented in the same way or pace for all alytine species. This realization stems not only from logistic issues, resource availability, legal listings, protected areas, phylogenetic considerations or threatened categories. Instead, it is largely derived from focusing on particular species in their actual habitats and distributions. Alytines live only in six of the numerous identified freshwater ecoregions [148]. However, alytines do not fit easily into this biogeographic partitioning, highlighting the need to develop conservation-oriented regionalization based on a variety of taxa [149]. In such syntheses, insular settings should be highlighted and treated as what they are: unique, isolated, evolutionarily relevant and, thus, distinct.
4.4. Island Nativeness Matters
All alytines are strongly affected by habitat loss, either in insular or continental settings. This is in contrast with the findings for taxa with much higher dispersal abilities, such as birds and mammals. In such highly mobile species, the impact of habitat loss has been overestimated on continents [150,151]. In exchange, the case of alytines supports the notion that continental species may be just as sensitive to extinction as insular species. After all, threats add up to the point where in terms of conservation status, it does not make much difference whether species live on islands or on continents. Still, this is just a partial answer.
Developing a methodological tool for including phenotypic and ecological uniqueness into conservation priorities is a top priority. This is what both the IUCN ranks and EDGE scores miss. The ferreret is not the most basal alytine, but it is definitely the most divergent in morphology, life history, behavior and habitat. Indeed, island species often have very high EDGE scores, but this is only because ancient islands often harbor taxa representing lineages that have long become extinct elsewhere. In other cases, insular uniqueness may have proceeded at a faster rate. For example, the dodo of Mauritius was a member of the pigeon family [152], and the cabbage trees of Saint Helena are nested within the composite diversification [153]. However, their extreme overall divergence makes their loss much more tragic than the extinction of an average-looking dove or goldenrod representing a lineage of similar or even older age. This is so because phylogenetic relatedness is a constraint upon, not a motor of, evolutionary novelty. Indeed, species flocks, such as helicoid snails in Madeira, cichlid fish in Lake Tanganyika or paradise birds in New Guinea, have a rather shallow history and limited phylogenetic diversity. Nevertheless, these endemic taxa include many unique life forms that represent a magnificent and valuable portion of biodiversity [154]. Eventually, it may be sensible to focus conservation on phenotypes; this is what renders ecosystems functional and biodiversity a reality.
Insular species represent many of the most divergent life forms and deserve special attention. As a case in point, the isolated evolution of the only insular alytine accounts for its historical near-extinction. This occurred through a single main factor that has a milder impact on the mainland: extreme vulnerability to continental predators. Detecting and understanding nativeness (including endemism) is thus critical for conservation biology, given that islanders account for such a large fraction of endangered species. There is an urgent need for the explicit recognition of insularity as a key factor in determining susceptibility to human-induced impacts. Doing so may help avoid the loss of the most uniquely divergent species.
4.5. A Predictive Model of Insularity
It may seem troublesome to find out that continental species can indeed be similarly endangered as insular ones and, at the same time, to sustain the claim for the latter’s particular susceptibility to extinction. The paradox may be solved recognizing the insularity syndrome, making species evolving on islands more susceptible to external threats. This is a condition present not only in real islands, but in any isolated ecosystem. Thus, rivers, caves, desert springs, mountaintops, islands and any other type of discrete, discontinuous habitats can be insular. In such habitats, evolution will be more uniquely divergent the longer they have remained effectively isolated for the taxa in question and the smaller they are relative to the dispersal ability of the species involved. The stronger the insular syndrome, threats directly disrupting the unique evolutionary pathway will have greater weight. Paramount among such threats, exotic invasive species emerge as a key factor in island settings.
The insularity model developed herein represents a multi-scale approach to island biogeography, including species, processes and system feedback (as proposed by [155]). Time is not considered explicitly, being represented instead by evolutionary potential (E). Besides, time is not a driving mechanism of invasion, but a scale of observation [156]. Along the second axis (dispersal potential, D), taxa with very high dispersal rates, such as most birds, will require prolonged isolation to differentiate. However, isolation is not a simple distance measure (eventually, this makes little sense for birds [157]), being, instead, a complex function of that particular taxon’s behavior. The limit between insular and continental settings is comparable to the insular distribution function [158,159,160,161]; this represents just a particular plane of the three-dimensional graph.
This model accounts for general trends of insular extinctions and endangerment [2,6,8,15]. It explains why Holocene mammal extinctions were especially intense among insular species and largely caused by introduced species. In contrast, the current general threats for mammals in continental areas are mostly habitat loss and overexploitation [5]. Likewise, the susceptibility of freshwater ecosystems to invasion-driven collapse (e.g., [162]) is a consequence of their insular character in many aspects, including invasibility [163,164,165,166]. In addition, the model includes an explicit identification of scale. This allows for considering large areas, such as whole biotic regions (as suggested by [9,167]), as well as smaller subdivisions, leading to diversification in ecological islands within islands [2,133,168,169,170].
Although the model is focused on insularity, it can be used to address an invader’s potential (i.e., combining questions, as proposed by [171,172,173]). The uniqueness of insular species results from evolution in disharmonious communities (not necessarily species-poor assemblages [7]) and is directly related to susceptibility to extinction through invasion. In contrast, the resilience of continental species is the outcome of large effective ranges encompassing long-stressed, highly diverse communities. This resilience is, in turn, directly related to the ability to invade insular settings and cause extinctions therein. The outcome of a biological invasion depends on the differential in I between invading and resident species. However, the invasion’s deployment will vary according to various factors that are essentially historical, i.e., contingent [174].
The model can also be used to address the different pathways for building an insular community. On oceanic islands [175], they start at high D and moderate to high E values, then rising fast to high I scores. On islands of continental origin, there will be variable contributions from dispersal and vicariance events [166,176]. Thus, here, they start at moderate to large D values and may have a longer trajectory before exhibiting, if ever, insular syndromes. Acquiring high I scores may proceed at disparate rates and involve a wide array of life history modifications. These may range from fast-evolving acceleration in the case of P. falconeri [177] to slow-evolving delay as in M. balearicus [178]. The latter is also the case of Paradisaea apoda [179], whose unusual morphology and behavior must be largely attributed to insularity.
Finally, conservation planning may benefit from taking into account the relative position of a given species or community. High I scores should weigh heavily in assigning conservation priorities. On the other hand, high negative values point at potentially dangerous taxa or assemblages if translocated into insular settings. This could be relevant for attempts to conserve and restore native ecosystems on both islands and continents. For example, the only extinction among the continental species analyzed can now be explained by the low absolute I score of Ectopistes migratorius. Its remarkable fast flight, large range and dispersal ability [180] were typical of a continental bird. In spite of this, its large-scale nomadism and gregarious breeding [181] conferred it with a quasi-insular character. This resulted in an unusually high sensitivity to mass predation and habitat destruction.
Eventually, this model stems from two epistemological hypotheses; first, that all biological invasions follow the same set of rules, albeit each with its own peculiarities [25,26]; and second, that biogeography can indeed be a powerful tool for biological conservation [2,65,182]. Comparisons of susceptibility to extinction and conservation efficiency become thus meaningful. These are now possible by explicitly incorporating the evolutionary, ecological and geographic variables determining insularity, spanning through various temporal and spatial scales.
5. Conclusions
Detecting and understanding nativeness is critical, because susceptibility to extinction is not equivalent in continental and insular species. Known threats to alytines are landscape transformation, water extraction, introduced predators, emerging disease, pollution and climate change. All of them contribute to habitat fragmentation and loss and are largely shared by all alytine species. Although all alytines are threatened, an introduced continental predator acts as a powerful driver of the near-extinction of the only extant insular species. This interaction has not elicited any evolutionary response in the native prey.
Continental and insular species can be similarly endangered, in spite of the latter’s particular susceptibility. This paradox may be solved by recognizing the insularity syndrome (I) in any isolated ecosystem. I can be interpreted as a function of evolutionary and dispersal potentials. The extreme overall divergence of insular endemics calls for considering phenotypic and ecological uniqueness in the assessment of conservation priorities. Phylogenetic relatedness should also be taken into account separately; although being a constraint upon evolutionary novelty, it should not mask uniquely divergent life forms. The insularity syndrome model may prove helpful for understanding the qualitative aspects of island biogeography, assessing the risk and deployment of biological invasions and guiding the conservation of biodiversity.
Acknowledgments
Lina Ponsell provided invaluable support and insights. Álvaro Román taught me much natural history of ferrerets. Carme Rosell, Roser Campeny and Eduard Filella did so with continental midwife toads. Gloria Fernández kindly shared her knowledge on captive breeding. Samuel Piña openly discussed alytine conservation. Jaume Belda at CREAIB helped in searching for the literature. Nick Isaac answered my questions on EDGE scores. Wouter Beukema discussed the threats affecting Alytes maurus. Regina Cunha critically read a previous version of the manuscript. Two anonymous reviewers provided extremely valuable criticism and suggestions.
Conflicts of Interest
The author declares no conflict of interest.
References
- Pimm, S.L. The dodo went extinct (and other ecological myths). Ann. Mo. Bot. Gard. 2002, 89, 190–198. [Google Scholar] [CrossRef]
- Altaba, C.R. Biodiversity of the Balearic Islands: A Paradigm for Conservation. In Seminar on Biodiversity and Biological Conservation; Vilà, M., Rodà, F., Ros, J., Eds.; Institut d’Estudis Catalans: Barcelona, Spain, 2004; pp. 371–389. [Google Scholar]
- Steadman, D.W. Extinction and Biogeography of Tropical Pacific Birds; University of Chicago Press: Chicago, IL, USA, 2006. [Google Scholar]
- Stork, N.E. Re-assessing current extinction rates. Biodivers. Conserv. 2010, 19, 357–371. [Google Scholar] [CrossRef]
- Turvey, S.T.; Fritz, S.A. The ghosts of mammals past: Biological and geographical patterns of global mammalian extinction across the Holocene. Philos. Trans. R. Soc. B 2011, 366, 2564–2576. [Google Scholar]
- Sax, D.F.; Gaines, S.D. Species invasions and extinction: The future of native biodiversity on islands. Proc. Natl. Acad. Sci. USA 2008, 105, 11490–11497. [Google Scholar] [CrossRef]
- Davis, M.A. Researching Invasive Species 50 Years after Elton: A Cautionary Tale. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Chichester, UK, 2011; pp. 269–276. [Google Scholar]
- Loehle, C.; Eschenbach, W. Historical bird and terrestrial mammal extinction rates and causes. Divers. Distrib. 2012, 18, 84–91. [Google Scholar] [CrossRef]
- Manne, L.L.; Brooks, T.M.; Pimm, S.L. Relative risk of extinction of passerine birds on continents and islands. Nature 1999, 399, 258–261. [Google Scholar] [CrossRef]
- Vermeij, G. Invasion as Expectation: A Historical Fact of Life. In Species Invasions: Insights into Ecology, Evolution and Biogeography; Sax, D., Stachowicz, J., Gaines, S., Eds.; Sinauer: Sunderland, MA, USA, 2005; pp. 315–339. [Google Scholar]
- Chew, M.K.; Hamilton, A.L. The Rise and Fall of Biotic Nativeness: A Historical Perspective. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Chichester, UK, 2011; pp. 121–130. [Google Scholar]
- Szabo, J.K.; Khwaja, N.; Garnett, S.T.; Butchart, S.H.M. Global patterns and drivers of avian extinctions at the species and subspecies level. PLoS One 2012, 7, e47080:1–e47080:9. [Google Scholar]
- Blackburn, T.M.; Cassey, P.; Duncan, R.P.; Evans, K.L.; Gaston, K.J. Avian extinction and mammalian introductions on oceanic islands. Science 2004, 305, 1955–1958. [Google Scholar] [CrossRef]
- Clavero, M.; Garcia-Berthou, E. Invasive species are a leading cause of animal extinctions. Trends Ecol. Evol. 2005, 20, 110. [Google Scholar] [CrossRef]
- Donlan, C.J.; Wilcox, C. Diversity, invasive species and extinctions in insular ecosystems. J. Appl. Ecol. 2008, 45, 1114–1123. [Google Scholar] [CrossRef]
- Corlett, R.I. Invasive aliens on tropical East Asian islands. Biodivers. Conserv. 2010, 19, 411–423. [Google Scholar] [CrossRef]
- Ricciardi, A. Assessing species invasions as a cause of extinction. Trends Ecol. Evol. 2004, 19, 619–619. [Google Scholar] [CrossRef]
- Roy, H.E.; Adriaens, T.; Isaac, N.J.B.; Kenis, M.; Onkelinx, T.; San Martin, G.; Brown, P.M.J.; Hautier, L.; Poland, R.; Roy, D.B.; et al. Invasive alien predator causes rapid declines of native European ladybirds. Divers. Distrib. 2012, 18, 717–725. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Townsend, C.R. Interpopulation variation in mayfly antipredator tactics: Differential effects of contrasting predatory fish. Ecology 1994, 75, 2078–2090. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Lau, J.A.; Carroll, S.P. Evolutionary responses of natives to introduced species: What do introductions tell us about natural communities? Ecol. Lett. 2006, 9, 357–374. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Lovette, I.J. The roles of island area per se and habitat diversity in the species-area relationships of four Lesser Antillean faunal groups. J. Anim. Ecol. 1999, 68, 1142–1160. [Google Scholar] [CrossRef]
- Kadmon, R.; Allouche, O. Integrating the effects of area, isolation, and habitat heterogeneity on species diversity: A unification of island biogeography and niche theory. Am. Nat. 2007, 170, 443–454. [Google Scholar] [CrossRef]
- Finlayson, C.; Monclova, A.; Carrión, J.S.; Fa, D.A.; Finlayson, G.; Rodríguez-Vidal, J.; Fierro, E.; Fernández, S.; Bernal-Gómez, M.; Giles-Pacheco, F. Ecological transitions—but for whom? A perspective from the Pleistocene. Palaeogeogr. Palaeoclimatiol. 2012, 329–330, 1–9. [Google Scholar] [CrossRef]
- Gillespie, R.G.; Baldwin, B.G. Island Biogeography of Remote Archipelagoes. Interplay between Ecological and Evolutionary Processes. In The Theory of Island Biogeography Revisited; Losos, J.B., Ricklefs, R.E., Eds.; Princeton University Press: Princeton, NJ, USA, 2010; pp. 358–387. [Google Scholar]
- Altaba, C.R. Are all mass invasions alike? Trends Ecol. Evol. 2000, 15, 248. [Google Scholar] [CrossRef]
- Richardson, D.M. Invasion Science: The Roads Travelled and the Roads Ahead. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Chichester, UK, 2011; pp. 397–407. [Google Scholar]
- Altaba, C.R. Phylogeny and biogeography of midwife toads (Alytes, Discoglossidae): A reappraisal. Contrib. Zool. 1997, 66, 257–262. [Google Scholar]
- Altaba, C.R. Phylogenetic relationships and biogeography of midwife toads (Anura: Alytinae): vicariance in a dynamic insular setting preceding adaptive radiation. 2014; unpublished work. [Google Scholar]
- Zangari, F.; Cimmaruta, F.; Nascetti, G. Genetic relationships of the western Mediterranean painted frogs based on allozymes and mitochondrial markers: Evolutionary and taxonomic inferences (Amphibia, Anura, Discoglossidae). Biol. J. Linn. Soc. 2006, 87, 515–536. [Google Scholar] [CrossRef]
- Blackburn, D.C.; Bickford, D.P.; Diesmos, A.C.; Iskandar, D.T.; Brown, R.M. An ancient origin for the enigmatic flat-headed frogs (Bombinatoridae: Barbourula) from the islands of Southeast Asia. PLoS One 2010, 5, e12090. [Google Scholar]
- Pabijan, M.; Crottini, A.; Reckwell, D.; Irisarri, I.; Hauswaldt, J.S.; Vences, M. A multigene species tree for Western Mediterranean painted frogs (Discoglossus). Mol. Phylogenet. Evol. 2012, 64, 690–696. [Google Scholar] [CrossRef]
- Biton, R.; Geffen, E.; Vences, M.; Cohen, O.; Bailon, S.; Rabinovich, R.; Malka, Y.; Oron, T.; Boistel, R.; Brumfeld, V.; et al. The rediscovered Hula painted frog is a living fossil. Nat. Commun. 2013, 4, e1959:1–e1959:6. [Google Scholar]
- Roelants, K.; Bossuyt, F. Archaeobatrachian paraphyly and pangaean diversification of crown-group frogs. Syst. Biol. 2005, 54, 111–126. [Google Scholar] [CrossRef]
- San Mauro, D.; Vences, M.; Alcobendas, M.; Zardoya, R.; Meyer, A. Initial diversification of living amphibians predated the breakup of Pangaea. Am. Nat. 2005, 165, 590–599. [Google Scholar] [CrossRef]
- Frost, D.R.; Grant, T.; Faivovich, J.N.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; De Sá, R.O.; Channing, A.; Wilkinson, M.; Donnellan, S.C.; et al. The amphibian tree of life. Bull. Am. Mus. Nat. Hist. 2006, 297, 1–370. [Google Scholar] [CrossRef]
- Roelants, K.; Gower, D.J.; Wilkinson, M.; Loader, S.P.; Biju, S.D.; Guillaume, K.; Moriau, L.; Bossuyt, F. Global patterns of diversification in the history of modern amphibians. Proc. Natl. Acad. Sci. USA 2007, 104, 887–892. [Google Scholar] [CrossRef]
- Irisarri, I.; Vences, M.; San Mauro, D.; Glaw, F.; Zardoya, R. Reversal to air-driven sound production revealed by a molecular phylogeny of tongueless frogs, family Pipidae. BMC Evol. Biol. 2011, 11, 114:1–114:10. [Google Scholar]
- Skerratt, L.F.; Berger, L.; Speare, R.; Cashins, S.; McDonald, K.R.; Phillott, A.D.; Hines, H.B.; Kenyon, N. Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. EcoHealth 2007, 4, 125–134. [Google Scholar] [CrossRef]
- Rödder, D.; Kielgast, J.; Bielby, J.; Schmidtlein, S.; Bosch, J.; Garner, T.W.J.; Veith, M.; Walker, S.; Fisher, M.C.; Lötters, S. Global amphibian extinction risk assessment for the panzootic chytrid fungus. Diversity 2009, 1, 52–66. [Google Scholar] [CrossRef]
- Rosenblum, E.B.; Voyles, J.; Poorten, T.J.; Stajich, J.E. The deadly chytrid fungus: A story of an emerging pathogen. PLoS Pathog. 2010, 6, e1000550. [Google Scholar] [CrossRef]
- Rosenblum, E.B.; Fisher, M.C.; James, T.Y.; Stajich, J.E.; Longcore, J.E.; Gentry, L.R.; Poorten, T.J. A molecular perspective: Biology of the emerging pathogen Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2010, 92, 131–147. [Google Scholar]
- Blaustein, A.R.; Walls, S.C.; Bancroft, B.A.; Lawler, J.L.; Searle, C.L.; Gervasi, S.C. Direct and indirect effects of climate change on amphibian populations. Diversity 2010, 2, 281–313. [Google Scholar] [CrossRef]
- Arntzen, J.W.; García-París, M. Morphological and allozyme studies of midwife toads (genus Alytes), including the description of two new taxa from Spain. Contrib. Zool. 1995, 65, 5–34. [Google Scholar]
- García-París, M.; Martínez-Solano, I. Nuevo estatus taxonómico para las poblaciones ibero-mediterráneas de Alytes obstetricans (Anura: Discoglossidae). Rev. Esp. Herpetol. 2001, 15, 99–113. [Google Scholar]
- Bosch, J.; Beebee, T.; Schmidt, B.; Tejedo, M.; Martínez-Solano, I.; Salvador, A.; García-París, M.; Recuero Gil, E.; Arntzen, J.W.; Diaz Paniagua, C.; et al. Alytes Obstetricans . IUCN Red List of Threatened Species. Version 2011. 2. Available online: http://www.iucnredlist.org (accessed on 9 March 2012).
- Araújo, M.B.; Thuiller, W.; Pearson, R.G. Climate warming and the decline of amphibians and reptiles in Europe. J. Biogeogr. 2006, 33, 1712–1728. [Google Scholar] [CrossRef]
- Malkmus, R. Amphibians and Reptiles of Portugal, Madeira and the Azores-Archipelago; Gantner: Ruggell, Liechtenstein, 2004. [Google Scholar]
- Beja, P.; Bosch, J.; Tejedo, M.; Lizana, M.; Martínez-Solano, I.; Salvador, A.; García-París, M.; Recuero Gil, E.; Arntzen, J.W.; Márquez, R.; et al. Alytes Cisternasii . IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2. Available online: http://www.iucnredlist.org (accessed on 9 March 2012).
- París, M.; Martínez-Solano, I.; Izquierdo, E.; García-París, M. Distribución y estado de conservación de los sapos parteros (Anura: Discoglossidae: Alytes) en la provincia de Albacete (Castilla-La Mancha, España). Sabuco: Rev. Estud. Albacet. 2002, 3, 5–22. [Google Scholar]
- Bosch, J.; Tejedo, M.; Lizana, M.; Martínez-Solano, I.; Salvador, A.; García-París, M.; Recuero Gil, E.; Arntzen, J.W.; Marquez, R.; Diaz Paniagua, C.; et al. Alytes dickhilleni . IUCN Red List of Threatened Species. Version 2011.2. Available online: http://www.iucnredlist.org (accessed on 9 March 2012).
- Donaire-Barroso, D.; Bogaerts, S. Datos sobre taxonomía, ecología y biología de Alytes maurus (Pasteur and Bons, 1962) (Anura: Discoglossidae). Butll. Soc. Catalana Herpetol. 2003, 16, 25–41, 139–140. [Google Scholar]
- Fromhage, L.; Vences, M.; Veith, M. Testing alternative vicariance scenarios in Western Mediterranean discoglossid frogs. Mol. Phylogenet. Evol. 2004, 31, 308–322. [Google Scholar] [CrossRef]
- Martínez-Solano, I.; Gonçalves, H.A.; Arntzen, J.W.; García-París, M. Phylogenetic relationships and biogeography of midwife toads (Discoglossidae: Alytes). J. Biogeogr. 2004, 31, 603–618. [Google Scholar] [CrossRef]
- Beukema, W.; De Pous, P.; Donaire-Barroso, D.; Bogaerts, S.; García-Porta, J.; Escoriza, D.; Arribas, O.J.; El Mouden, E.H.; Carranza, S. Review of the systematics, distribution, biogeography and natural history of Moroccan amphibians. Zootaxa 2013, 3661, 1–60. [Google Scholar]
- Donaire-Barroso, D.; Salvador, A.; Slimani, T.; El Mouden, E.H.; Martínez-Solano, I. Alytes maurus . IUCN Red List of Threatened Species. Version 2011.2. Available online: http://www.iucnredlist.org (accessed on 9 March 2012).
- Pleguezuelos, J.M.; Brito, J.C.; Fahd, S.; Feriche, M.F.; Mateo, J.A.; Moreno-Rueda, G.; Reques, R.; Santos, X. Setting conservation priorities for the Moroccan herpetofauna: The utility of regional red listing. Oryx 2010, 44, 501–508. [Google Scholar] [CrossRef]
- Mayol Serra, J.; Griffiths, R.; Bosch, J.; Beebee, T.; Schmidt, B.; Tejedo, M.; Lizana, M.; Martínez-Solano, I.; Salvador, A.; García-París, M.; et al. Alytes muletensis . IUCN Red List of Threatened Species. Version 2011.2. Available online: http://www.iucnredlist.org (accessed on 9 March 2012).
- Sanchiz, F.J.; Alcover, J.A. Un nou discoglòssid (Amphibia: Anura) de l’holocè de Menorca. Butll. Inst. Catalana Hist. Nat. 1982, 48, 99–105. [Google Scholar]
- Mayol, J.; Alcover, J.A. Survival of Baleaphryne Sanchiz and Adrover (Amphibia: Anura: Discoglossidae) on Mallorca. Amphibia-Reptilia 1981, 1, 343–345. [Google Scholar] [CrossRef]
- Alcover, J.A.; Mayol, J. Espèces relictuelles d’amphibiens et de reptiles des îles Baléares et Pityuses: Une extension des résultats. Bull. Soc. Herpéthol. Fr. 1982, 22, 69–74. [Google Scholar]
- Alcover, J.A.; Mayol, J.; Jaume, D.; Alomar, G.; Pomar, G.; Jurado, J. Biologia i ecologia de les poblacions relictes de Baleaphryne muletensis a la muntanya mallorquina. In Història biològica del ferreret; Hemmer, H., Alcover, J.A., Eds.; Moll: Palma de Mallorca, Spain, 1984; pp. 129–151. [Google Scholar]
- Altaba, C.R. La diversitat biològica: Una perspectiva des de Mallorca; Moll: Palma de Mallorca, Spain, 1999. [Google Scholar]
- Griffiths, R.A.; Schley, L.; Sharp, P.E.; Dennis, J.L.; Roman, A. Behavioural responses of Mallorcan midwife toad tadpoles to natural and unnatural snake predators. Anim. Behav. 1998, 55, 207–214. [Google Scholar] [CrossRef]
- Moore, R.D.; Griffiths, R.A.; O’Brien, C.M.; Murphy, A.; Jay, D. Induced defences in an endangered amphibian in response to an introduced snake predator. Oecologia 2004, 141, 139–147. [Google Scholar] [CrossRef]
- Richardson, D.M.; Whittaker, R.J. Conservation biogeography—foundations, concepts and challenges. Divers. Distrib. 2010, 16, 313–320. [Google Scholar] [CrossRef]
- Akçakaya, H.R.; Butchart, S.H.M.; Mace, G.M.; Stuart, S.N.; Hilton-Taylor, C. Use and misuse of the IUCN Red List Criteria in projecting climate change impacts on biodiversity. Glob. Chang. Biol. 2006, 12, 2037–2043. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1; IUCN Species Survival Commission: Gland, Switzerland, Cambridge, UK, 2001. [Google Scholar]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria. Version 9.0. Prepared by the Standards and Petitions Subcommittee.. Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 7 June 2012).
- 2004 IUCN Red List of Threatened Species: A Global Species Assessment; Baillie, J.E.M.; Hilton-Taylor, C.; Stuart, S.N. (Eds.) IUCN-SSC: Gland, Switzerland, Cambridge, UK, 2004; p. 191.
- Butchart, S.H.M.; Stattersfield, A.J.; Bennun, L.A.; Shutes, S.M.; Akçakaya, H.R.; Baillie, J.E.M.; Stuart, S.N.; Hilton-Taylor, C.; Mace, G.M. Measuring global trends in the status of biodiversity: Red List Indices for birds. PLoS Biol. 2004, 2, e383:2294–e383:2304. [Google Scholar]
- Butchart, S.H.M.; Stattersfield, A.J.; Baillie, J.; Bennun, L.A.; Stuart, S.N.; Akçakaya, H.R.; Hilton-Taylor, C.; Mace, G.M. Using Red List Indices to measure progress towards the 2010 target and beyond. Philos. Trans. R. Soc. Lond. B 2005, 360, 255–268. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Akçakaya, H.R.; Chanson, J.; Baillie, J.E.M.; Collen, B.; Quader, S.; Turner, W.R.; Amin, R.; Stuart, S.N.; Hilton-Taylor, C. Improvements to the Red List Index. PLoS One 2007, 2, e140:1–e140:8. [Google Scholar]
- Isaac, N.J.B.; Turvey, S.T.; Collen, B.; Waterman, C.; Baillie, J.E.M. Mammals on the EDGE: Conservation priorities based on threat and phylogeny. PLoS One 2007, 2, e296:1–e296:7. [Google Scholar]
- Haas, A. Phylogeny of frogs as inferred from primarily larval characters (Amphibia: Anura). Cladistics 2003, 19, 23–89. [Google Scholar]
- Waliszewski, P.; Konarski, J. A Mystery of the Gompertz Function. In Fractals in Biology and Medicine; Losa, G.A., Merlini, D., Nonnenmacher, T.F., Weibel, E.R., Eds.; Birkhäuser: Basel, Switzerland, 2005; pp. 277–286. [Google Scholar]
- Gil, M.M.; Miller, F.A.; Brandão, T.R.S.; Silva, C.L.M. On the use of the Gompertz model to predict microbial thermal inactivation under isothermal and non-isothermal conditions. Food Eng. Rev. 2011, 3, 17–25. [Google Scholar] [CrossRef]
- Bosch, J. Alytes obstetricans . In Atlas y libro rojo de los anfibios y reptiles de España; Pleguezuelos, J.M., Márquez, R., Lizana, M., Eds.; Dirección General de Conservación de la Naturaleza & Asociación Herpetológica Española: Madrid, Spain, 2002; pp. 82–84. [Google Scholar]
- Bosch, J.; Martínez-Solano, I.; García-París, M. Evidence of a chytrid fungus infection involved in the decline of the common midwife toad (Alytes obstetricans) in protected areas of central Spain. Biol. Conserv. 2001, 97, 331–337. [Google Scholar] [CrossRef]
- Schmidt, B.R.; Zumbach, S. Rote Liste der gefährdeten Amphibien der Schweiz; Bundesamt für Umwelt, Land und Landschaft and Koordinationsstelle Amphibien- und Reptilienschutz in der Schweiz: Bern, Switzerland, 2005. [Google Scholar]
- Tobler, U.; Schmidt, B.R. Within- and among-population variation in chytridiomycosis-induced mortality in the toad Alytes obstetricans. PLoS One 2010, 5, e10927:1–e10927:8. [Google Scholar]
- Ferrer, J.; Filella, E. Sobre la presència dels escurçons Vipera latastei Boscá, 1878 i Vipera aspis (Linnaeus, 1758), i del tòtil Alytes obstetricans (Laurenti, 1768) a la península del Cap de Creus. Butll. Soc. Catalana Herpetol. 2011, 19, 184–194. [Google Scholar]
- Bosch, J.; Carrascal, L.M.; Durán, L.; Walker, S.; Fisher, M.C. Climate change and outbreaks of amphibian chytridiomycosis in a montane area of Central Spain; is there a link? Proc. R. Soc. B 2007, 274, 253–260. [Google Scholar] [CrossRef]
- Márquez, R.; Crespo, E. Alytes cisternasii . In Atlas y libro rojo de los anfibios y reptiles de España; Pleguezuelos, J.M., Márquez, R., Lizana, M., Eds.; Dirección General de Conservación de la Naturaleza & Asociación Herpetológica Española: Madrid, Spain, 2002; pp. 73–75. [Google Scholar]
- Márquez, R. Sapo partero ibérico –Alytes cisternasii. Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Ed.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2011. Available online: http://www.vertebradosibericos.org/ (accessed on 12 September 2012).
- Sánchez Videgaín, J.; Rubio de Lucas, J.L. Atlas preliminar de los anfibios y reptiles de las sierras prebéticas albaceteñas. Al-Basit 1996, 38, 5–30. [Google Scholar]
- García-París, M.; Arntzen, J.W. Alytes dickhilleni . In Atlas y libro rojo de los anfibios y reptiles de España; Pleguezuelos, J.M., Márquez, R., Lizana, M., Eds.; Dirección General de Conservación de la Naturaleza & Asociación Herpetológica Española: Madrid, Spain, 2002; pp. 76–78. [Google Scholar]
- García-París, M. Anura. In Amphibia Lissamphibia; García-París, M., Montori, A., Herrero, P., Eds.; CSIC: Madrid, Spain, 2004; Fauna Ibérica; Volume 24, pp. 1–639. [Google Scholar]
- Seguimiento de Alytes dickhilleni: Informe final. In Monografías SARE. Asociación Herpetológica Española; Bosch, J.; González-Miras, E. (Eds.) Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2012; p. 84.
- Bosch, J.; García-Alonso, D.; Fernández-Beaskoetxea, S.; Fisher, M.C.; Garner, T.W.J. Evidence for the introduction of lethal chytridiomycosis affecting wild betic midwife toads (Alytes dickhilleni). EcoHealth 2013, 10, 82–89. [Google Scholar] [CrossRef]
- Márquez, R.; Francisco Beltrán, J.; Slimani, T.; Radi, M.; Llusia, D.; El Mouden, H. Description of the advertisement call of the Moroccan midwife toad (Alytes maurus Pasteur and Bons, 1962). Alytes 2011, 27, 142–150. [Google Scholar]
- Román, A. Alytes muletensis . In Atlas y libro rojo de los anfibios y reptiles de España; Pleguezuelos, J.M., Márquez, R., Lizana, M., Eds.; Dirección General de Conservación de la Naturaleza & Asociación Herpetológica Española: Madrid, Spain, 2002; pp. 79–81. [Google Scholar]
- Viada, C. Libro rojo de los vertebrados de las Baleares, 3rd ed.; Conselleria de Medi Ambient: Palma, Spain, 2006. [Google Scholar]
- Moore, R.D.; Griffiths, R.; Román, A. Distribution of the Mallorcan midwife toad (Alytes muletensis) in relation to landscape topography and introduced predators. Biol. Conserv. 2004, 116, 327–332. [Google Scholar] [CrossRef]
- Walker, S.F.; Bosch, J.; James, T.Y.; Litvintseva, A.P.; Valls, J.A.O.; Pinya, S.; García, G.; Rosa, G.A.; Cunningham, A.A.; Hole, S.; et al. Introduced pathogens threaten species recovery programs. Curr. Biol. 2008, 18, R853–R854. [Google Scholar] [CrossRef]
- Alcover, J.A. El sapillo balear, un fósil viviente. Quercus 1989, 39, 14–19. [Google Scholar]
- Mayol, J.; Alcover, J.A. La conservación de Alytes muletensis. In Història biològica del ferreret; Hemmer, H., Alcover, J.A., Eds.; Moll: Palma de Mallorca, Spain, 1984; pp. 245–252. [Google Scholar]
- Climate Change and Water. In Technical Paper of the Intergovernmental Panel on Climate Change; Bates, B.C.; Kundzewicz, Z.W.; Wu, S.; Palutikof, J.P. (Eds.) IPCC Secretariat: Geneva, Switzerland, 2008.
- Homar, V.; Ramis, C.; Romero, R.; Alonso, S. Recent trends in temperature and precipitation over the Balearic Islands (Spain). Clim. Chang. 2010, 98, 199–211. [Google Scholar] [CrossRef]
- Hardiman, N.; Burgin, S. Effects of trampling on in-stream macroinvertebrate communities from canyoning activity in the Greater Blue Mountains World Heritage Area. Wetl. Ecol. Manag. 2011, 19, 61–71. [Google Scholar] [CrossRef]
- Pinya, S.; Pérez-Mellado, V.; Suárez-Fernández, J.J. First records of limb malformations in wild populations of the endangered Balearic Midwife Toad’s, Alytes muletensis. Herpetol. Rev. 2012, 43, 240–243. [Google Scholar]
- Iglesias, A.; Garrote, L.; Flores, F.; Moneo, M. Challenges to manage the risk of water scarcity and climate change in the Mediterranean. Water Resour. Manag. 2007, 21, 775–788. [Google Scholar] [CrossRef]
- Sumner, G.N.; Romero, R.; Homar, V.; Ramis, C.; Alonso, S.; Zorita, E. An estimate of the effects of climate change on the rainfall of Mediterranean Spain by the late twenty first century. Clim. Dyn. 2003, 20, 789–805. [Google Scholar]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef]
- Lips, K.R.; Brem, F.; Brenes, R.; Reeve, J.D.; Alford, R.A.; Voyles, J.; Carey, C.; Livo, L.; Pessier, A.P.; Collins, J.P. Emerging infectious disease and the loss of biodiversity in a Neotropical amphibian community. Proc. Natl. Acad. Sci. USA 2006, 103, 3165–3170. [Google Scholar] [CrossRef]
- Kriger, K.M.; Hero, J.-M. Chytridiomycosis, amphibian extinctions, and lessons for the prevention of future Panzootics. EcoHealth 2009, 6, 6–10. [Google Scholar] [CrossRef]
- Crawford, A.J.; Lips, K.R.; Bermingham, E. Epidemic disease decimates amphibian abundance, species diversity, and evolutionary history in the highlands of central Panama. Proc. Natl. Acad. Sci. USA 2010, 107, 13777–13782. [Google Scholar] [CrossRef]
- Collins, J.P.; Crump, M.L. Extinction in Our Times: Global Amphibian Decline. Oxford University Press: New York, NY, USA, 2009. [Google Scholar]
- Fisher, M.C.; Garner, T.W.J.; Walker, S.F. Global emergence of Batrachochytrium dendrobatidis and amphibian chytridiomycosis in space, time, and host. Annu. Rev. Microbiol. 2009, 63, 291–310. [Google Scholar] [CrossRef]
- Pounds, A.J.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef]
- El Mouden, E.H.; Slimani, T.; Donaire, D.; Fernández-Beaskoetxea, S.; Fisher, M.C.; Bosch, J. First record of the chytrid fungus Batrachochytrium dendrobatidis in North Africa. Herpetol. Rev. 2011, 42, 71–75. [Google Scholar]
- Bloxam, Q.M.C.; Tonge, S.J. Amphibians: Suitable candidates for breeding-release programmes. Biodiver. Conserv. 1995, 4, 636–644. [Google Scholar] [CrossRef]
- Mayol Serra, J. Rèptils i amfibis de les Balears, edició revisada; Moll: Ciutat de Mallorca, Spain, 2003. [Google Scholar]
- Boisvert, S.P.; Davidson, E.W. Growth of the amphibian pathogen, Batrachochytrium dendrobatidis, in response to chemical properties of the aquatic environment. J. Wildl. Dis. 2011, 47, 694–698. [Google Scholar] [CrossRef]
- Andre, S.E.; Parker, J.; Briggs, C.J. Effect of temperature on host response to Batrachochytrium dendrobatidis infection in the mountain yellow-legged frog (Rana muscosa). J. Wildl. Dis. 2008, 44, 716–720. [Google Scholar] [CrossRef]
- Forrest, M.J.; Schlaepfer, M.A. Nothing a hot bath won’t cure: Infection rates of amphibian chytrid fungus correlate negatively with water temperature under natural field settings. PLoS One 2011, 6, e28444:1–e28444:9. [Google Scholar]
- Altaba, C.R. Un endemisme ornitològic ignorat: el trencapinyons balear (Loxia balearica). Butll. Inst. Catalana Hist. Nat. 2001, 69, 77–90. [Google Scholar]
- EDGE of Existence: Evolutionarily Distinct and Globally Endangered. Available online: http://www.edgeofexistence.org/amphibians/default.php (accessed on 18 January 2013).
- Rachmayuningtyas, B.A.; Bickford, D.P.; Kamsi, M.; Kutty, S.N.; Meier, R.; Arifin, U.; Rachmansah, A.; Iskandar, D.T. Conservation status of the only lungless frog Barbourula kalimantanensis Iskandar, 1978 (Amphibia: Anura: Bombinatoridae). J. Threat. Taxa 2011, 3, 1981–1989. [Google Scholar] [CrossRef]
- Galán, P. Redescubierto el sapillo pintojo de vientre negro o sapillo de Hula. Quercus 2012, 313, 46–48. [Google Scholar]
- McElduff, F.; Cortina-Borja, M.; Chan, S.-K.; Wade, A. When t-tests or Wilcoxon-Mann-Whitney tests won’t do. Adv. Physiol. Educ. 2010, 34, 128–133. [Google Scholar] [CrossRef]
- Lea, J.; Dyson, M.; Halliday, T. The effects of cohort structure and density on larval growth and development in Alytes muletensis: Implications for conservation. Herpetol. J. 2002, 12, 155–164. [Google Scholar]
- Carfagno, G.L.F.; Carithers, J.M.; Mycoff, L.J.; Lehtinen, R.M. How the cricket frog lost its spot: The inducible defense hypothesis. Herpetologica 2011, 67, 386–396. [Google Scholar] [CrossRef]
- Marquis, O.; Saglio, P.; Neveu, A. Effects of predators and conspecific chemical cues on the swimming activity of Rana temporaria and Bufo bufo tadpoles. Arch. Hydrobiol. 2004, 160, 153–170. [Google Scholar] [CrossRef]
- Mandrillon, A.L.; Saglio, P. Prior exposure to conspecific chemical cues affects predator recognition in larval Common Toad (Bufo bufo). Arch. Hydrobiol. 2005, 164, 1–12. [Google Scholar] [CrossRef]
- Ferrari, M.C.O.; Messier, F.; Chivers, D.P. First documentation of cultural transmission of predator recognition by larval amphibians. Ethology 2007, 113, 621–627. [Google Scholar] [CrossRef]
- Gonzalo, A.; López, P.; Martin, J. Iberian Green Frog tadpoles may learn to recognize novel predators from chemical alarm cues of conspecifics. Anim. Behav. 2007, 74, 447–453. [Google Scholar] [CrossRef]
- Jara, F.B.; Perotti, M.G. Risk of predation and behavioural response in three anuran species: Influence of tadpole size and predator type. Hydrobiologia 2010, 644, 313–324. [Google Scholar] [CrossRef]
- Shah, A.A.; Ryan, M.J.; Bevilacqua, E.; Schlaepfer, M.A. Prior experience alters the behavioral response of prey to a nonnative predator. J. Herpetol. 2010, 44, 185–192. [Google Scholar] [CrossRef]
- Scoville, A.G.; Pfrender, M.E. Phenotypic plasticity facilitates recurrent rapid adaptation to introduced predators. Proc. Natl. Acad. Sci. USA 2010, 107, 4260–4263. [Google Scholar] [CrossRef]
- Gomez-Mestre, I.; Díaz-Paniagua, C. Invasive predatory crayfish do not trigger inducible defences in tadpoles. Proc. R. Soc. B 2011, 278, 3364–3370. [Google Scholar] [CrossRef] [Green Version]
- Cromsigt, J.P.G.M.; Kerley, G.I.H.; Kowalczyk, R. The difficulty of using species distribution modelling for the conservation of refugee species –the example of European bison. Divers. Distrib. 2012, 18, 1253–1257. [Google Scholar] [CrossRef]
- Fisher, M.C.; Bosch, J.; Yin, Z.K.; Stead, D.; Walker, J.; Selway, L.; Brown, A.J.P.; Walker, L.A.; Gow, N.A.R.; Stajich, J.E.; et al. Proteomic and phenotypic profiling of an emerging pathogen of amphibians, Batrachochytrium dendrobatidis, shows that genotype is linked to virulence. Mol. Ecol. 2009, 18, 415–429. [Google Scholar] [CrossRef]
- Gonçalves, H.; Martínez-Solano, I.; Pereira, R.J.; Carvalho, B.; García-París, M.; Ferrand, N. High levels of population subdivision in a morphologically conserved Mediterranean toad (Alytes cisternasii) result from recent, multiple refugia: Evidence from mtDNA, microsatellites and nuclear genealogies. Mol. Ecol. 2009, 18, 5143–5160. [Google Scholar]
- Chen, X.; Jiao, J.; Tong, X. A generalized model of island biogeography. Sci. China Life Sci. 2011, 54, 1055–1061. [Google Scholar] [CrossRef]
- Gewin, V. Riders of a modern-day ark. PLoS Biol. 2008, 6, e24:18–e24:21. [Google Scholar] [CrossRef]
- Scott, J.M.; Davis, F.; Csuti, B.; Noss, R.; Butterfield, B.; Groves, C.; Anderson, H.; Caicco, S.; D'Erchia, F.; Edwards, T.C., Jr.; et al. Gap analysis: A geographic approach to protection of biological diversity. Wildl. Monogr. 1993, 123, 3–41. [Google Scholar]
- Rodrigues, A.S.L.; Akçakaya, H.R.; Andelman, S.J.; Bakkarr, M.I.; Boitani, L.; Brooks, T.M.; Chanson, J.S.; Fishpool, L.D.C.; da Fonseca, G.A.B.; Gaston, K.J.; et al. Global gap analysis: Priority regions for expanding the global protected-area network. Bioscience 2004, 54, 1092–1100. [Google Scholar] [CrossRef]
- Lowe, W.H.; Nislow, K.H.; Likens, G.E. Forest structure and stream salamander diets: Implications for terrestrial-aquatic connectivity. Verh. Int. Verein Limnol. 2005, 29, 279–286. [Google Scholar]
- Baldwin, R.F.; deMaynadier, P.G. Assessing threats to pool-breeding amphibian habitat in an urbanizing landscape. Biol. Conserv. 2009, 142, 1628–1638. [Google Scholar] [CrossRef]
- Beier, P.; Noss, R.F. Do habitat corridors provide connectivity? Conserv. Biol. 1998, 12, 1241–1252. [Google Scholar] [CrossRef]
- Surasinghe, T.D. Major issues in threat analysis and resolving such problems: An addendum to the GAP analysis. J. Threat. Taxa 2012, 4, 2545–2550. [Google Scholar] [CrossRef]
- Crooks, K.R.; Sanjayan, M. Connectivity Conservation; Cambridge University Press: Cambridge, UK, New York, NY, USA, 2006; p. 732. [Google Scholar]
- Stevens, V.M.; Baguette, M. Importance of habitat quality and landscape connectivity for the persistence of endangered natterjack toads. Conserv. Biol. 2008, 22, 1194–1204. [Google Scholar] [CrossRef]
- Smith, K.F.; Sax, D.F.; Lafferty, K.D. Evidence for the role of infectious disease in species extinction and endangerment. Conserv. Biol. 2006, 20, 1349–1357. [Google Scholar] [CrossRef]
- Garner, T.W.J.; Garcia, G.; Carroll, B.; Fisher, M.C. Using itraconazole to clear Batrachochytrium dendrobatidis infection, and subsequent depigmentation of Alytes muletensis tadpoles. Dis. Aquat. Organ. 2009, 83, 257–260. [Google Scholar] [CrossRef]
- Kriger, K.M.; Hero, J.-M. After the horse has bolted: A reply to Garner et al. EcoHealth 2009, 6, 152. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Han, B.A.; Relyea, R.A.; Johnson, P.T.J.; Buck, J.C.; Gervasi, S.S.; Kats, L.B. The complexity of amphibian population declines: Understanding the role of cofactors in driving amphibian losses. Ann. N. Y. Acad. Sci. 2011, 1223, 108–119. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Contreras Balderas, S.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. Bioscience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Bass, M.S.; Finer, M.; Jenkins, C.N.; Kreft, H.; Cisneros-Heredia, D.F.; McCracken, S.F.; Pitman, N.C.A.; English, P.H.; Swing, K.; Villa, G.; et al. Global conservation significance of Ecuador’s Yasuní National Park. PLoS One 2010, 5, e8767. [Google Scholar] [CrossRef]
- He, F.; Hubbell, S.P. Species-area relationships always overestimate extinction rates from habitat loss. Nature 2011, 473, 368–371. [Google Scholar] [CrossRef]
- Fisher, D.O.; Blomberg, S.P. Correlates of rediscovery and the detectability of extinction in mammals. Philos. Trans. R. Soc. B 2011, 278, 1090–1097. [Google Scholar]
- Pereira, S.L.; Johnson, K.P.; Clayton, D.H.; Baker, A.J. Mitochondrial and nuclear DNA sequences support a Cretaceous origin of Columbiformes and a dispersal-driven radiation in the Paleogene. Syst. Biol. 2007, 56, 656–672. [Google Scholar] [CrossRef]
- Eastwood, A.; Gibby, M.; Cronk, Q.C.B. Evolution of St Helena arborescent Astereae (Asteraceae): Relationships of the genera Commidendrum and Melanodendron. Bot. J. Linn. Soc. 2004, 144, 69–83. [Google Scholar] [CrossRef]
- Altaba, C.R. Documenting biodiversity: The need for species identifications. Trends Ecol. Evol. 1997, 12, 358–359. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Brown, J.H. The reticulating phylogeny of island biogeography theory. Q. Rev. Biol. 2009, 84, 357–390. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef]
- Kalmar, A.; Currie, D.J. A global model of island biogeography. Glob. Ecol. Biogeogr. 2006, 15, 72–81. [Google Scholar] [CrossRef]
- Lomolino, M.V. A call for a new paradigm of island biogeography. Glob. Ecol. Biogeogr. 2000, 9, 1–6. [Google Scholar] [CrossRef]
- Lomolino, M.V. A species-based theory of insular zoogeography. Glob. Ecol. Biogeogr. 2000, 9, 39–58. [Google Scholar] [CrossRef]
- Lomolino, M.V. Body size evolution in insular vertebrates: Generality of the island rule. J. Biogeogr. 2005, 32, 1683–1699. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Brown, J.H.; Sax, D.F. Island Biogeography Theory. Reticulations of “a Biogeography of the Species”. In The Theory of Island Biogeography Revisited; Losos, J.B., Ricklefs, R.E., Eds.; Princeton University Press: Princeton, NJ, USA, 2010; pp. 13–51. [Google Scholar]
- Cox, J.G.; Lima, S.L. Naiveté and an aquatic-terrestrial dichotomy in the effects of introduced predators. Trends Ecol. Evol. 2006, 21, 674–680. [Google Scholar] [CrossRef]
- Margalef, R. Limnología; Omega: Barcelona, Spain, 1983. [Google Scholar]
- Margalef, R.; Alcover, J.A.; Altaba, C.R.; Camarasa, J.M. De l’aïllament a la insularitat (in Catalan). In Biosfera; Folch, R., Camarasa, J.M., Eds.; Enciclopèdia Catalana: Barcelona, Spain, 1995; Volume 9, pp. 328–337, [English version: Gale Group, Madison, WI, USA, 2000.]. [Google Scholar]
- Altaba, C.R. Testing vicariance: Melanopsid snails and Neogene tectonics in the Western Mediterranean. J. Biogeogr. 1998, 25, 541–551. [Google Scholar]
- Ricciardi, A.; MacIsaac, H.J. Biodiversity as a Bulwark against Invasion: Conceptual Threads since Elton. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Chichester, UK, 2011; pp. 211–224. [Google Scholar]
- Fridley, J.D. Biodiversity as a Bulwark against Invasion: Conceptual Threads since Elton. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Chichester, UK, 2011; pp. 121–130. [Google Scholar]
- Buckley, R. The habitat-unit model of island biogeography. J. Biogeogr. 1982, 9, 339–344. [Google Scholar] [CrossRef]
- Losos, J.B.; Parent, C.E. The Speciation-Area Relationship. In The Theory of Island Biogeography Revisited; Losos, J.B., Ricklefs, R.E., Eds.; Princeton University Press: Princeton, NJ, USA, 2010; pp. 415–438. [Google Scholar]
- Rosindell, J.; Phillimore, A.B. A unified model of island biogeography sheds light on the zone of radiation. Ecol. Lett. 2011, 14, 552–560. [Google Scholar] [CrossRef]
- Vermeij, G. An agenda for invasion biology. Biol. Conserv. 1996, 78, 3–9. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Tilman, D. What Species Invasions Tell Us about the Relationship between Community Saturation, Species Diversity and Ecosystem Functioning. In Species Invasions: Insights into Ecology, Evolution and Biogeography; Sax, D., Stachowicz, J., Gaines, S., Eds.; Sinauer: Sunderland, MA, USA, 2005; pp. 41–64. [Google Scholar]
- Van Kleunen, M.; Dawson, W.; Schlaepfer, D.; Jeschke, J.M.; Fischer, M. Are invaders different? A conceptual framework of comparative approaches for assessing determinants of invasiveness. Ecol. Lett. 2010, 13, 947–958. [Google Scholar]
- Gould, S.J. The Structure of Evolutionary Theory; Belknap Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Whittaker, R.J.; Triantis, K.A.; Ladle, R.J. A general dynamic theory of oceanic island biogeography. J. Biogeogr. 2008, 35, 977–994. [Google Scholar] [CrossRef]
- Morrone, J.J. Evolutionary Biogeography: An Integrative Approach with Case Studies; Columbia University Press: New York, NY, USA, 2009; p. 301. [Google Scholar]
- Raia, P.; Barbera, C.; Conte, M. The fast life of a dwarfed giant. Evol. Ecol. 2003, 17, 293–312. [Google Scholar] [CrossRef]
- Marín-Moratalla, N.; Jordana, X.; García-Martínez, R.; Köhler, M. Tracing the evolution of fitness components in fossil bovids under different selective regimes. C. R. Palevol 2011, 10, 469–478. [Google Scholar] [CrossRef]
- Irestedt, M.; Jønsson, K.A.; Fjeldså, J.; Christidis, L.; Ericson, P.G.P. An unexpectedly long history of sexual selection in birds-of-paradise. BMC Evol. Biol. 2009, 9, e235. [Google Scholar] [CrossRef]
- Johnson, K.P.; Clayton, D.H.; Dumbacher, J.P.; Fleischer, R.C. The flight of the passenger pigeon: Phylogenetics and biogeographic history of an extinct species. Mol. Phylogenet. Evol. 2010, 57, 455–458. [Google Scholar] [CrossRef]
- Bucher, E.H. The Causes of Extinction of the Passenger Pigeon. In Current Onrnithology; Power, D.M., Ed.; Plenum Press: New York, NY, USA, 1992; Volume 9, pp. 1–36. [Google Scholar]
- Van Kleunen, M.; Richardson, D.M. Invasion biology and conservation biology: Time to join forces to explore the links between species traits and extinction risk and invasiveness. Prog. Phys. Geogr. 2007, 31, 447–450. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Altaba, C.R. Extinction Resilience of Island Species: An Amphibian Case and a Predictive Model. Diversity 2014, 6, 43-71. https://doi.org/10.3390/d6010043
AMA Style
Altaba CR. Extinction Resilience of Island Species: An Amphibian Case and a Predictive Model. Diversity. 2014; 6(1):43-71. https://doi.org/10.3390/d6010043
Chicago/Turabian StyleAltaba, Cristian R. 2014. "Extinction Resilience of Island Species: An Amphibian Case and a Predictive Model" Diversity 6, no. 1: 43-71. https://doi.org/10.3390/d6010043