Stand Structure and Composition Affect the Drought Sensitivity of Oregon White Oak (Quercus garryana Douglas ex Hook.) and Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco)


Abstract
:1. Introduction
- Douglas-fir and Oregon white oak trees growing at the same site will exhibit different relationships to climate.
- Trees growing at high densities will be more sensitive to climatic variability than will trees growing at low densities.
- Trees with broad crowns and large lower limbs (characteristic of an open canopy at the time of establishment and early growth) will exhibit different relationships to climate than will trees with narrow crowns and few lower branches (characteristic of closed canopy environments).
- Trees competing with conspecifics for resources will exhibit different relationships to climate than trees competing with heterospecifics.
2. Materials and Methods
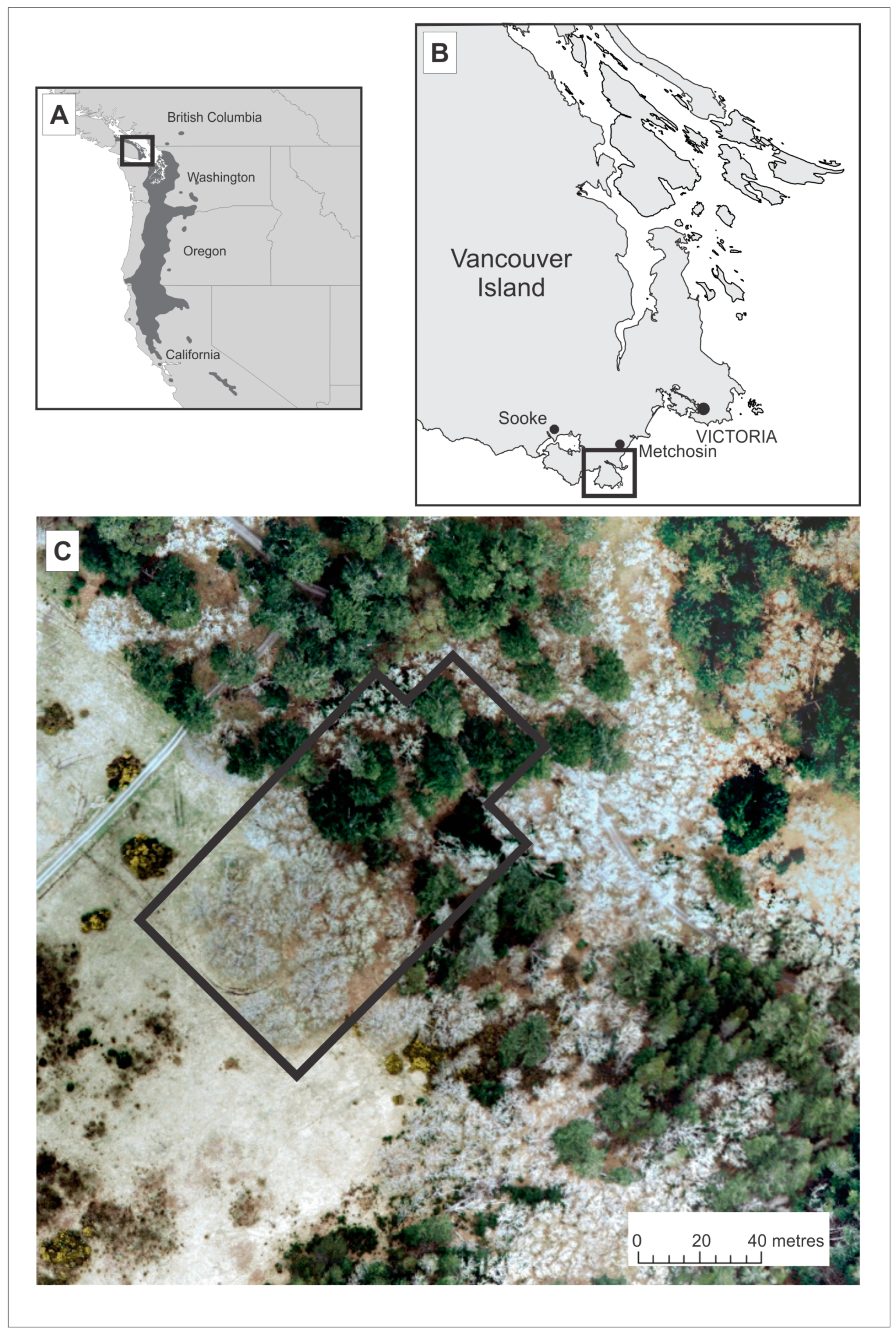
2.1. Study Site
2.2. Chronology Development
- Species Chronologies: all Oregon white oak; all Douglas-fir
- Stand Density Chronologies: Stem density was defined based on the total basal area index (BAI) within each quadrat. The upper and lower quartiles of BAI were used to distinguish quadrats of high- and low-densities respectively. All trees of a given species (i.e., Oregon white oak or Douglas-fir) within that quadrat were included in the subset used to generate the chronology.
- Morphology Chronologies: The morphology of each overstory tree was classified into two categories: open-grown (trees exhibiting large lower branches, a wide crown and a deep canopy); and closed-grown (trees exhibiting few lower branches, a narrow, tall crown and a generally erect posture). Chronologies for each species were developed for each of the two morphology types. While there is some overlap between the stand density chronologies and the morphology chronologies there were sufficient differences to merit analyzing them separately. For Oregon white oak, of the 57 cores that contributed to the closed-grown chronology 14 were also included in the high-density chronology, accounting for 64% of the high density chronology. For the open-grown chronology 7 of the 10 cores were also in the low-density chronology, accounting for 32% of that chronology.
- Intra- and Inter-Competitive Dynamics Chronologies: Separate chronologies were developed using cores from quadrats containing only a single species (i.e. only Oregon white oak or only coniferous) and quadrats containing a mix of species.
2.3. Assessing Growth–Climate Associations
2.4. Analyses and Interpretation
3. Results
3.1. Growth–Climate Associations
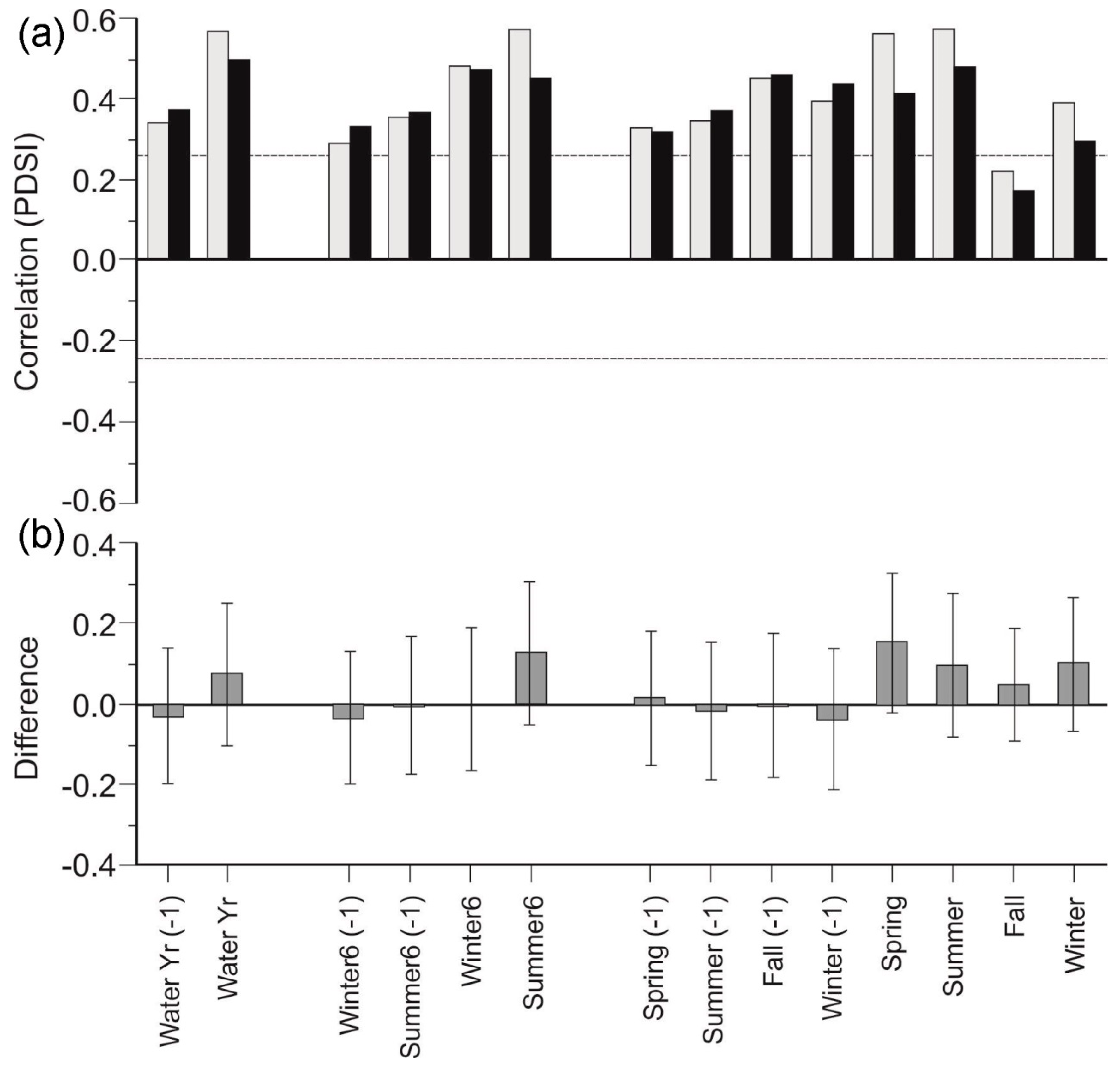
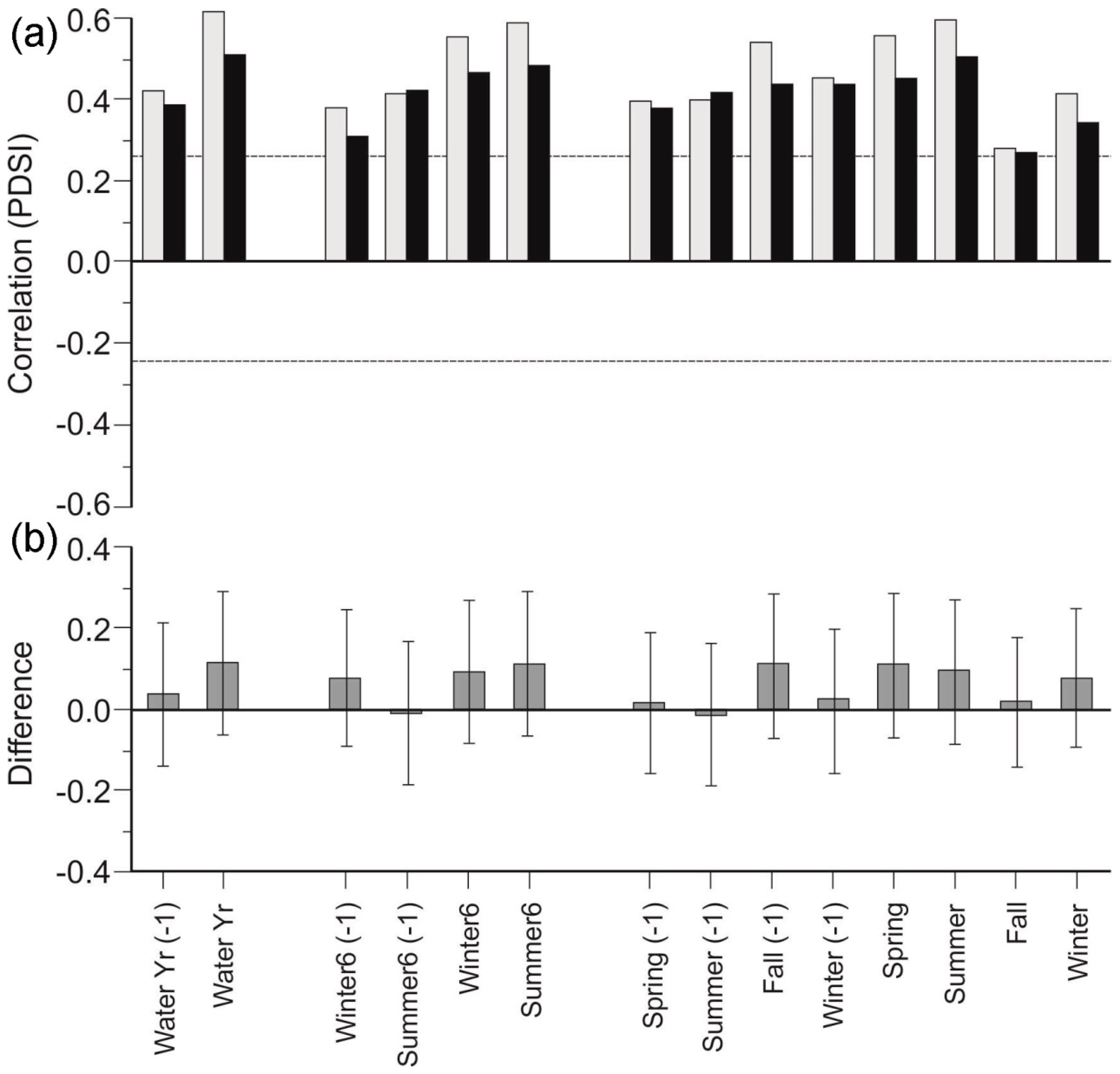
3.2. Species Chronologies
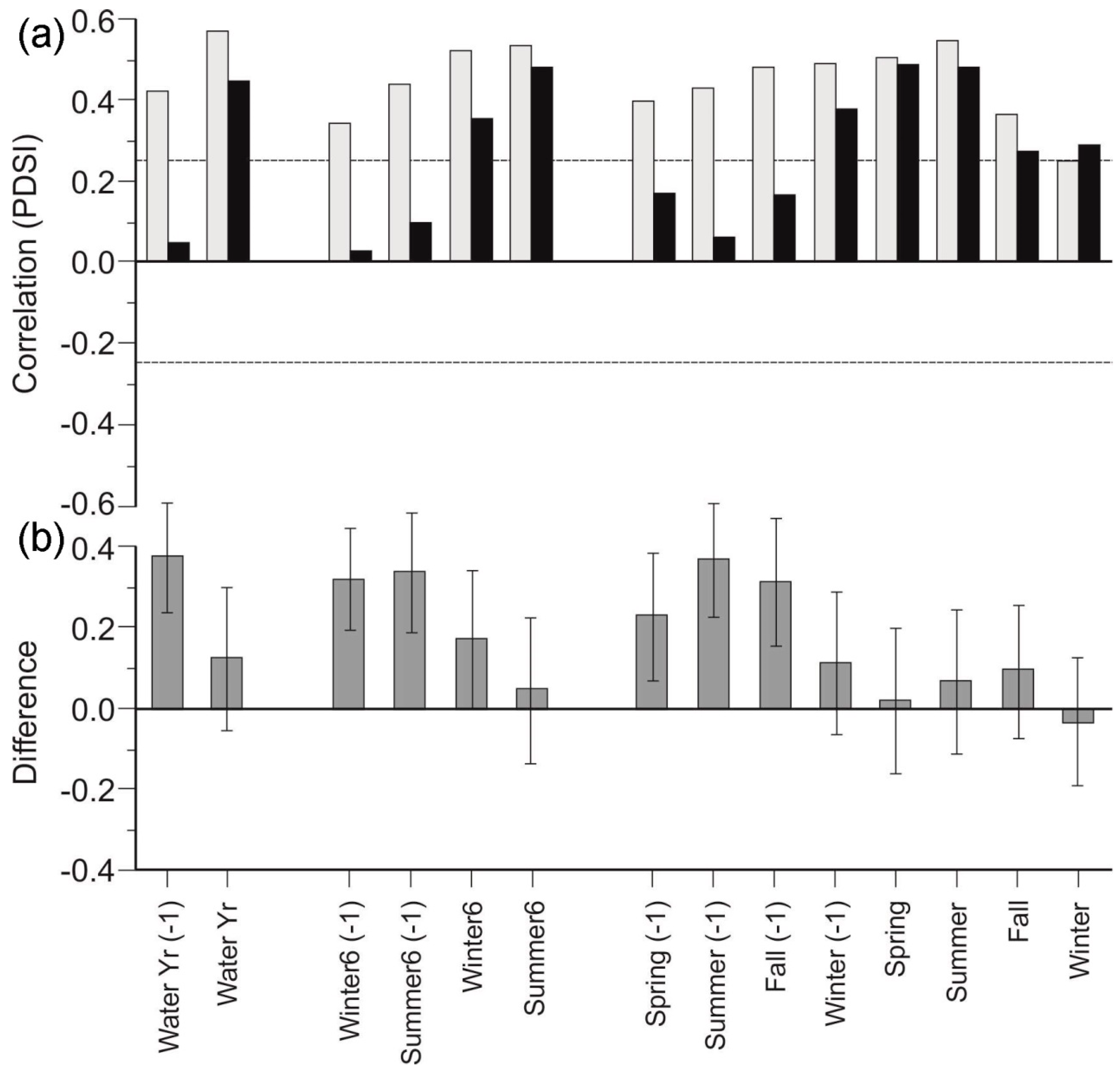
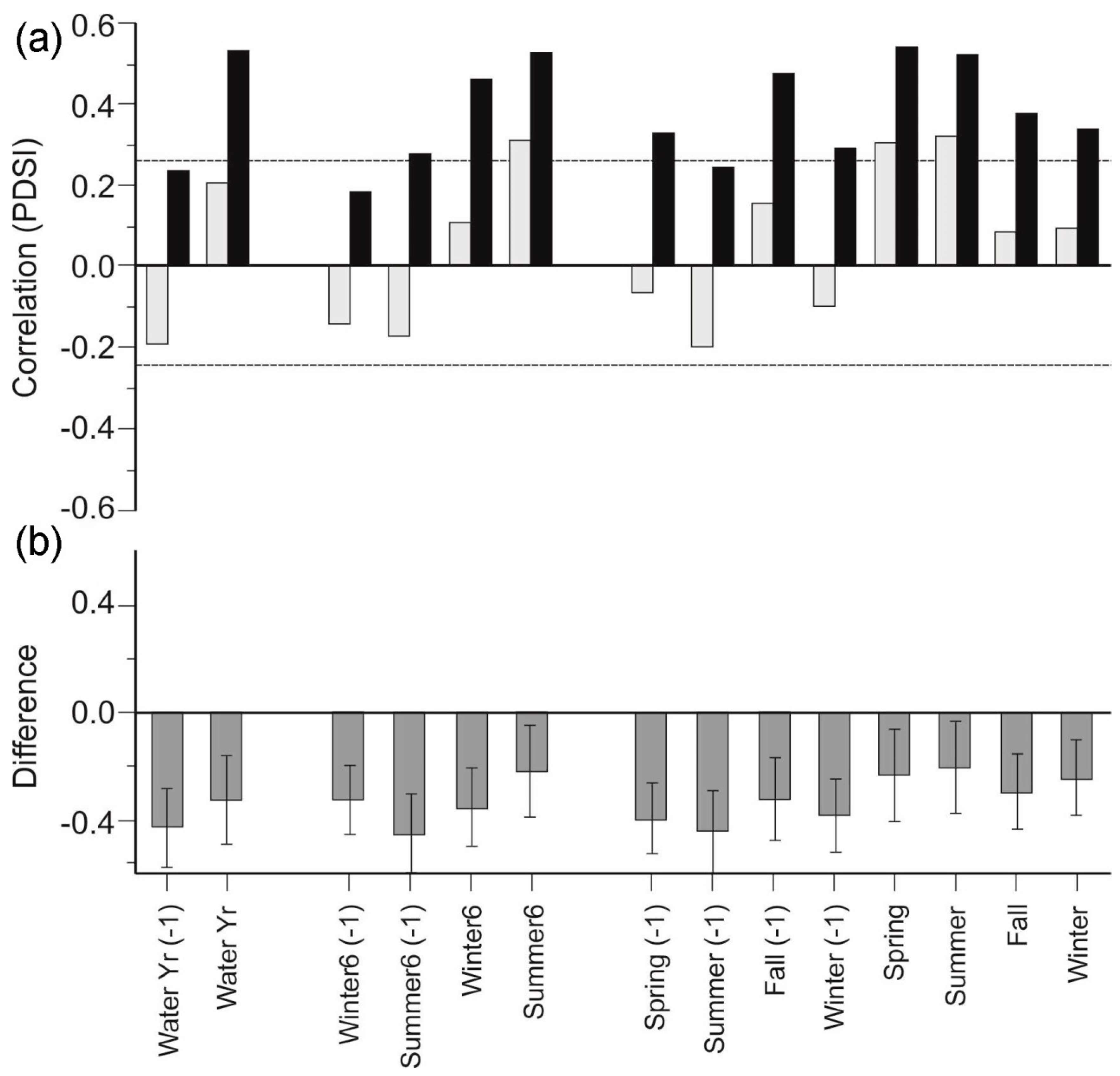
3.3. Stand Density Chronologies
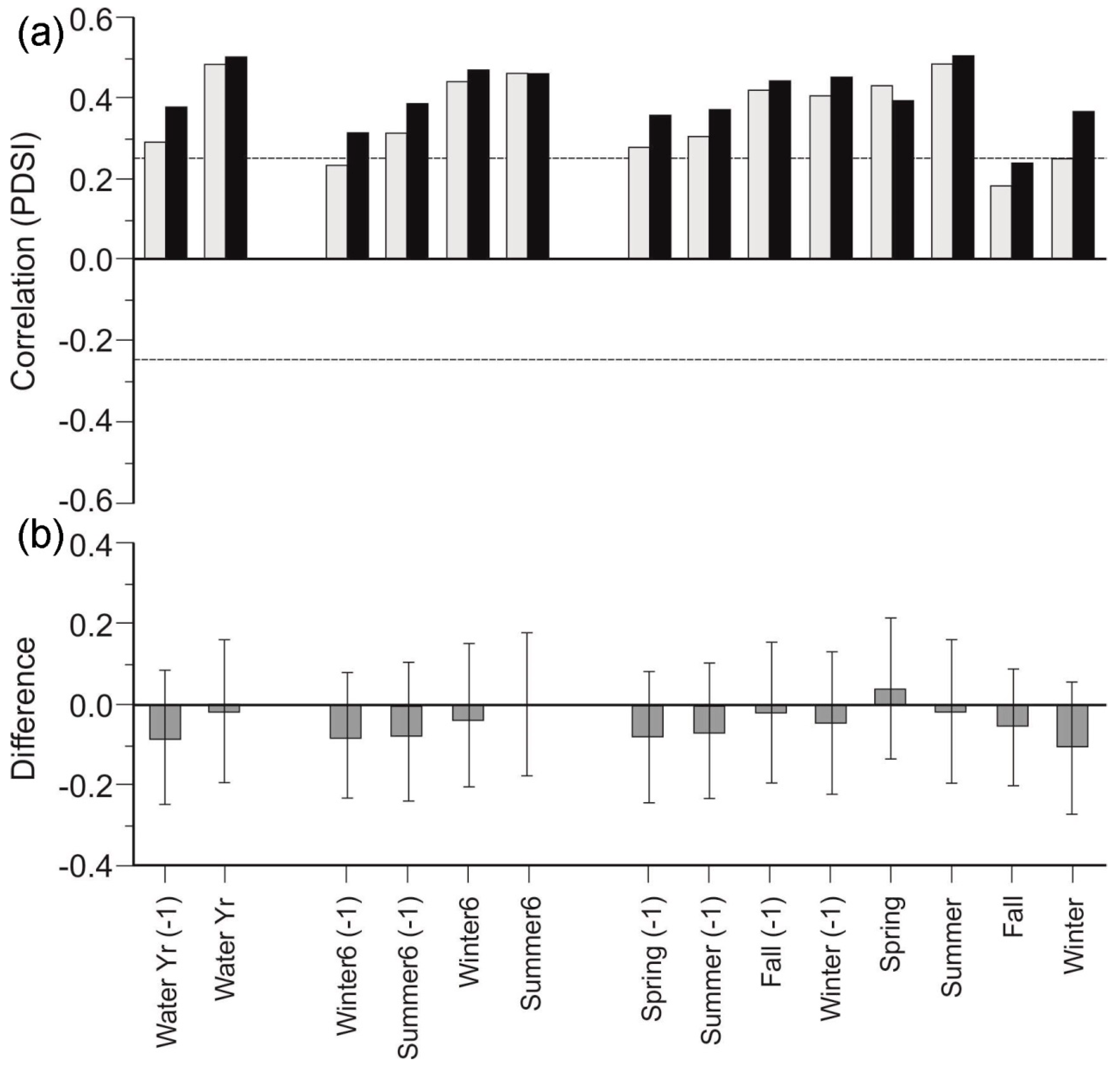
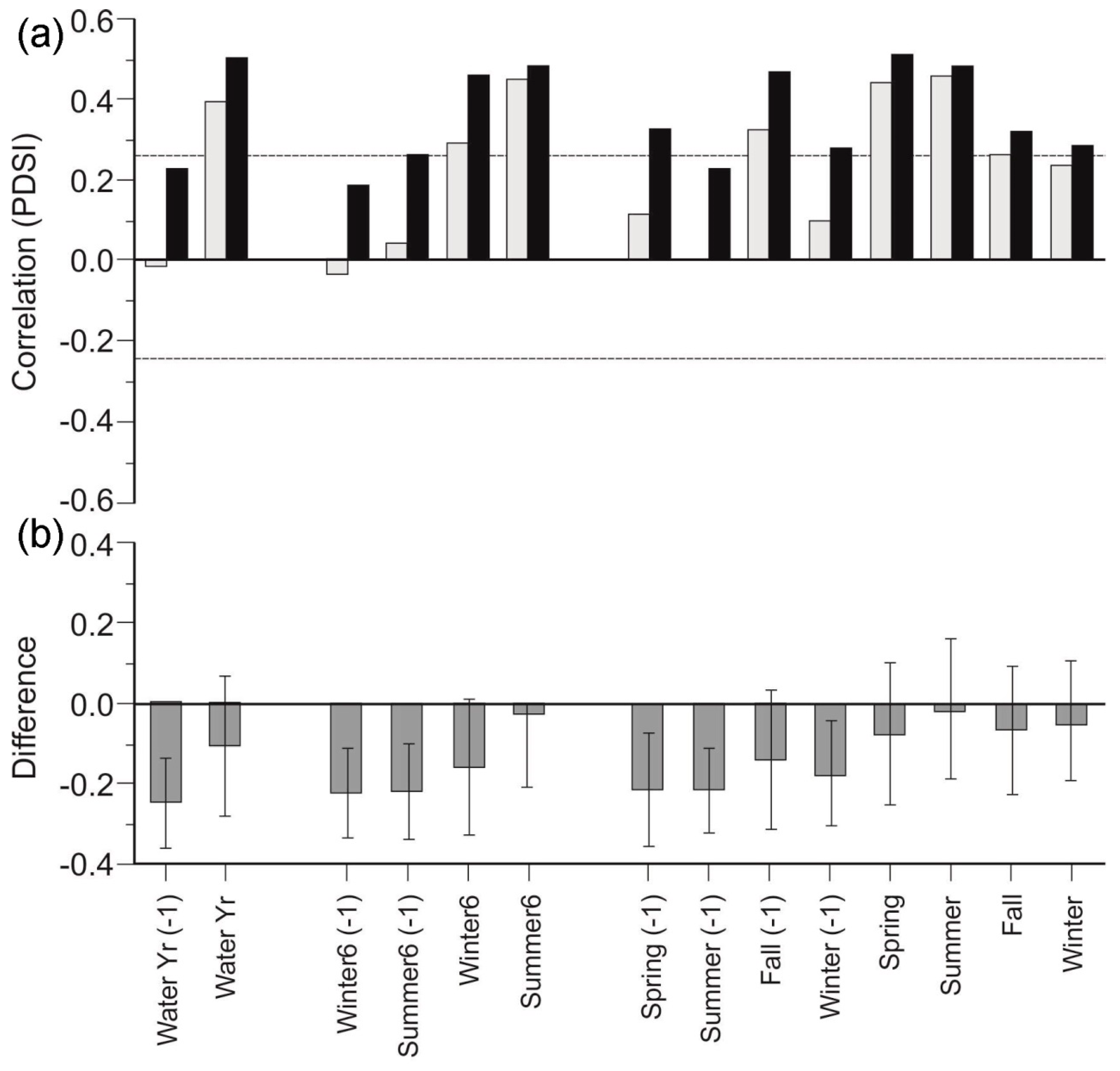
3.4. Canopy Morphology Chronologies
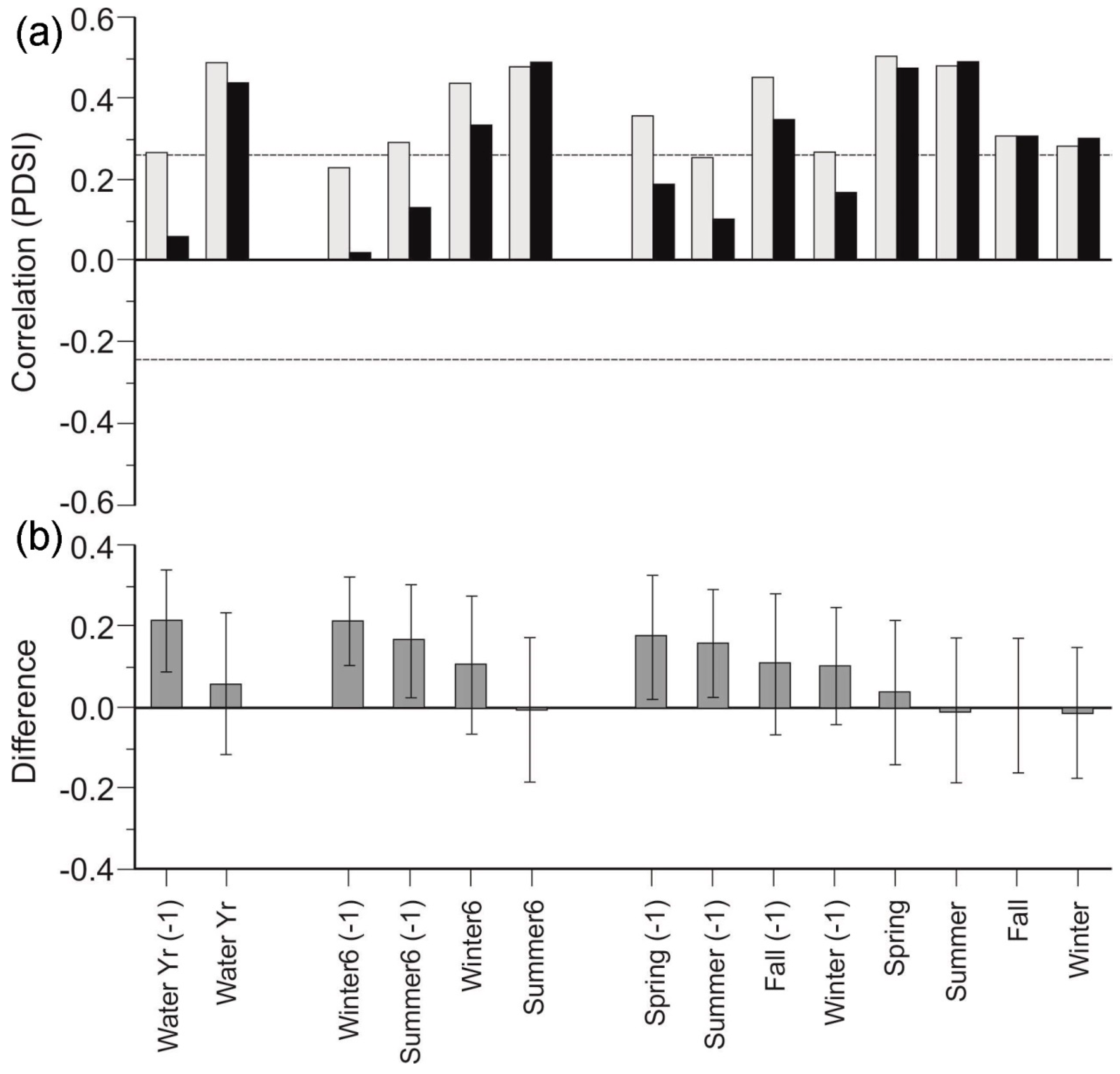
3.5. Stand Composition Chronologies
4. Discussion
4.1. Species-specific Responses to Drought
4.2. Stand Density and Responses to Drought
4.3. Morphological Responses to Drought
4.4. Competitive Interactions and Responses to Drought
4.5. Competitive Interactions and Climate Reconstructions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silvertown, J. Plant coexistence and the niche. Trends Ecol. Evol. 2004, 19, 605–611. [Google Scholar] [CrossRef]
- Fonti, P.; Cherubini, P.; Rigling, A.; Weber, P.; Biging, G. Tree rings show competition dynamics in abandoned Castanea sativa coppices after land-use changes. J. Veg. Sci. 2006, 17, 103–112. [Google Scholar] [CrossRef]
- Ettl, G.J.; Peterson, D.L. Extreme climate and variation in tree growth: individualistic response in subalpine fir (Abies lasiocarpa). Chang. Boil. 1995, 1, 231–241. [Google Scholar] [CrossRef]
- Dalen, L.; Hofgaard, A. Differential Regional Treeline Dynamics in the Scandes Mountains. Arctic Antarct. Alp. 2005, 37, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.L.; Hammer, R.D. From open to closed canopy: A century of change in a Douglas-fir forest, Orcas Island, Washington. Northwest Sci. 2001, 75, 262–269. [Google Scholar]
- Barnhart, S.J.; McBride, J.R.; Warner, P. Invasion of northern oak woodlands by Pseudotsuga menziesii (Mirb.) Franco in the Sonoma Mountains of California. Madroño 1996, 43, 28–45. [Google Scholar]
- Hanson, P.J.; Weltzin, J.F. Drought disturbance from climate change: response of United States forests. Sci. Total Environ. 2000, 262, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Waring, R.H.; Pitman, G.B. Modifying Lodgepole Pine Stands to Change Susceptibility to Mountain Pine Beetle Attack. Ecology 1985, 66, 889–897. [Google Scholar] [CrossRef]
- Harrington, T.B.; Tappeiner, J.C.; Ii, J.C.T. Growth responses of young Douglas-fir and tanoak 11 years after various levels of hardwood removal and understory suppression in southwestern Oregon, USA. Ecol. Manag. 1997, 96, 1–11. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 701–720. [Google Scholar] [CrossRef]
- Weber, P.; Bugmann, H.; Fonti, P.; Rigling, A. Using a retrospective dynamic competition index to reconstruct forest succession. Ecol. Manag. 2008, 254, 96–106. [Google Scholar] [CrossRef]
- Scholes, R.; Archer, S.R. TREE-GRASS INTERACTIONS IN SAVANNAS1. Annu. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Fuchs, M.A. Towards a Recovery Strategy for Garry Oak and Associated Ecosystems in Canada: Ecological Assessment and Literature Review; Canadian Wildlife Service: Ladner, British; Columbia, Canada, 2001.
- Lea, T. Historical Garry Oak Ecosystems of Greater Victoria & Saanich Peninsula; Terrestrial Information Branch, B.C. Ministry of Sustainable Resource Management: Victoria, BC, Canada, 2002.
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef]
- Gedalof, Z.; Pellatt, M.G.; Smith, D.J. From prairie to forest: Three centuries of environmental change at Rocky Point, Vancouver Island, BC. Northwest Sci. 2006, 80, 34–46. [Google Scholar]
- Stein, W.I. Quercus garryana Dougl. ex Hook. Oregon White Oak, in Silvics of North. America; Burns, R.M., Honkala, B.H., Eds.; U.S. Dept. of Agriculture, Forest Service: Washington, WA, USA, 1990.
- Regan, A.C.; Agee, J.K. Oak community and seedling response to fire at Fort Lewis, Washington. Northwest Sci. 2004, 78, 1–11. [Google Scholar]
- Brudvig, L.A.; Asbjornsen, H. Oak Regeneration Before and After Initial Restoration Efforts in a Tall Grass Oak Savanna Conference Paper. Am. Midl. Nat. 2005, 153, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Pellatt, M.G.; Gedalof, Z. Environmental change in Garry oak (Quercus garryana) ecosystems: the evolution of an eco-cultural landscape. Biodivers. Conserv. 2014, 23, 2053–2067. [Google Scholar] [CrossRef] [Green Version]
- Conedera, M.; Stanga, P.; Oester, B.; Bachmann, P. Different post-culture dynamics in abandoned chestnut orchards. For. Snow Landsc. Res. 2001, 76, 487–492. [Google Scholar]
- Schriver, M.; Sherriff, R.L.; Varner, J.M.; Quinn-Davidson, L.; Valachovic, Y. Age and stand structure of oak woodlands along a gradient of conifer encroachment in northwestern California. Ecosphere 2018, 9, e02446. [Google Scholar] [CrossRef]
- Sprague, F.I.; Hansen, H.P. Forest succession in the McDonald Forest, Willamette Valley, Oregon. Northwest Sci. 1946, 20, 89–98. [Google Scholar]
- Pellatt, M.G.; McCoy, M.M.; Mathewes, R.W. Paleoecology and fire history of Garry oak ecosystems in Canada: implications for conservation and environmental management. Biodivers. Conserv. 2015, 24, 1621–1639. [Google Scholar] [CrossRef]
- Nuszdorfer, F.C.; Klinka, K.; Demarchi, D.A. Coastal Douglas-fir Zone, in Ecosystems of British Columbia; Meidinger, D., Pojar, J., Eds.; Research Branch, Ministry of Forests: Victoria, BC, Canada, 1991.
- Cook, E.R.; Holmes, R.L. Program ARSTAN, Version 1.72P; University of Arizona Press: Tucson, AZ, USA, 1986. [Google Scholar]
- Cook, E.R.; Peters, K. The smoothing spline: A new approach to standardizing forest interior tree-ring width series for dendrochronology. Tree-Ring Bull. 1981, 41, 45–53. [Google Scholar]
- Guiot, J. ARMA techniques for modelling tree-ring response to climate and for reconstructing variations of paleoclimates. Ecol. Model. 1986, 33, 149–171. [Google Scholar] [CrossRef]
- Biondi, F.; Swetnam, T.W. Box-Jenkins models of forest interior tree-ring chronologies. Tree-Ring Bull. 1987, 47, 71–96. [Google Scholar]
- Mosteller, F.; Tukey, J.W. Data Analysis and Regression; Addison Wesley: New York, NY, USA, 1977; p. 608. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Adademic Press: London, UK, 1976; p. 567. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Palmer, W.C. Meteorological Drought; Office of Climatology, U.S. Weather Bureau: Washington, DC, USA, 1965.
- Heim, R.R. A Review of Twentieth-Century Drought Indices Used in the United States. Am. Meteorol. Soc. 2002, 83, 1149–1166. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Meko, D.M.; Stahle, D.W.; Cleaveland, M.K. Drought Reconstructions for the Continental United States*. J. Clim. 1999, 12, 1145–1162. [Google Scholar] [CrossRef]
- Watson, E.; Luckman, B.H. The dendroclimatic signal in Douglas-fir and ponderosa pine tree-ring chronologies from the southern Canadian Cordillera. Can. J. 2002, 32, 1858–1874. [Google Scholar] [CrossRef]
- Gedalof, Z.; Peterson, D.L.; Mantua, N.J. COLUMBIA RIVER FLOW AND DROUGHT SINCE 1750. JAWRA J. Am. N.a. Resour. Assoc. 2004, 40, 1579–1592. [Google Scholar] [CrossRef]
- Shabbar, A.; Skinner, W. Summer Drought Patterns in Canada and the Relationship toGlobal Sea Surface Temperatures. J. Clim. 2004, 17, 2866–2880. [Google Scholar] [CrossRef]
- Zar, J. Biostatistical Analysis, 4th ed.; Prentice Hall Press: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Speer, J.H. Fundamentals of Tree Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010; p. 352. [Google Scholar]
- Bassett, J.R. Tree Growth as Affected by Soil Moisture Availability. Soil Sci. Soc. J. 1964, 28, 436–438. [Google Scholar] [CrossRef]
- Kramer, P.J. The Role of Water in Wood Formation in The Formation of Wood in Forest Trees; Zimmerman, M.H., Ed.; Academic Press: New York, NY, USA, 1964; pp. 519–532. [Google Scholar]
- Fritts, H.C.; Smith, D.G.; Stokes, M.A. The Biological Model for Paleoclimatic Interpretation of Mesa Verde Tree-Ring Series. Memoirs Soc. Am. Archaeol. 1965, 19, 101–121. [Google Scholar] [CrossRef]
- Rigling, A.; Bräker, O.; Schneiter, G.; Schweingruber, F. Intra-annual tree-ring parameters indicating differences in drought stress of Pinus sylvestris forests within the Erico-Pinion in the Valais (Switzerland). N.a. Ecol. 2002, 163, 105–121. [Google Scholar]
- Fonti, P.; García-González, I.; Ladiges, P. Earlywood vessel size of oak as a potential proxy for spring precipitation in mesic sites. J. Biogeogr. 2008, 35, 2249–2257. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings and Environment: Dendroecology; Paul Haupt AG Bern: Berne, Switzerland, 1996; p. 609.
- Zimmermann, L.; Zeugin, F.; Zweifel, R.; Newbery, D.M. Intra-annual radial growth and water relations of trees: implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [Green Version]
- Owens, J.N.; Singh, H. Vegetative bud development and the time and method of cone initiation in subalpine fir ( Abies lasiocarpa ). Can. J. Bot. 1982, 60, 2249–2262. [Google Scholar] [CrossRef]
- Abrams, M.D. Adaptations and responses to drought in Quercus species of North America. Tree Physiol. 1990, 7, 227–238. [Google Scholar] [CrossRef]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Bräker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from mediterranean climates. Boil. Rev. 2003, 78, 119–148. [Google Scholar] [CrossRef]
- Hermann, R.K.; Lavender, D.P. Pseudotsuga menziesii (Mirb.) Franco. Douglas-fir, in Silvics of North; U.S. Dept. of Agriculture, Forest Service: Washington, WA, USA, 1990; pp. 527–540.
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Walker, I.J.; Sydneysmith, R. British Columbia, in From Impacts to Adaptation: Canada in a Changing Climate 2007; Government of Canada: Ottawa, ON, Canada, 2008; pp. 329–386.
- Teskey, R.O.; Hinckley, T.M. Influence of temperature and water potential on root growth of white oak. Physiol. Plant. 1981, 52, 363–369. [Google Scholar] [CrossRef]
- Eis, S. Root Grafts and their Silvicultural Implications. Can. J. 1972, 2, 111–120. [Google Scholar] [CrossRef]
- Lanner, R.M. Living Stumps in the Sierra Nevada. Ecology 1961, 42, 170–173. [Google Scholar] [CrossRef]
- Korakaki, E.; Vanderklein, D.; Mencuccini, M.; Martinez-Vilalta, J.; Hamid, H.A. Evidence for age- and size-mediated controls of tree growth from grafting studies. Tree Physiol. 2007, 27, 463–473. [Google Scholar] [Green Version]
- Zahner, R.; Donnelly, J.R. Refining Correlations of Water Deficits and Radial Growth in Young Red Pine. Ecology 1967, 48, 525–530. [Google Scholar] [CrossRef]
- Devine, W.D.; Harrington, C.A. Root system morphology of Oregon white oak on a glacial outwash soil. Northwest. Science 2005, 79, 179–188. [Google Scholar]
- Briffa, K.R.; Schweingruber, F.H.; Jones, P.D.; Osborn, T.J.; Harris, I.C.; Shiyatov, S.G.; Vaganov, E.A.; Grudd, H.; Osborn, T. Trees tell of past climates: but are they speaking less clearly today? Philos. Trans. Soc. B: Boil. Sci. 1998, 353, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.T.; Cufar, K.; Levanic, T. Temporal stability and dendroclimatology in silver fir and red spruce. Phyton Ann. Rei Bot. 1999, 39, 117–122. [Google Scholar]
- Biondi, F. Are Climate-Tree Growth Relationships Changing in North-Central Idaho, U.S.A.? Arctic, Antarct. Alp. 2000, 32, 111. [Google Scholar] [CrossRef]
- Szeicz, J.M.; Macdonald, G.M. Age-dependent tree-ring growth responses of subarctic white spruce to climate. Can. J. 1994, 24, 120–132. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chronology | N cores | Max age | Mean age | Mean r-bar | EPS | Lag-1 ACF | Mean S |
|---|---|---|---|---|---|---|---|
| All Douglas-fir | 33 | 136 | 100 | 0.419 | 0.951 | 0.429 | 0.152 |
| All Oregon white oak | 90 | 277 | 116 | 0.478 | 0.987 | 0.528 | 0.244 |
| Open-grown Douglas-fir | 6 | 123 | 119 | 0.302 | 0.706 | 0.393 | 0.175 |
| Open-grown Garry oak | 10 | 277 | 140 | 0.533 | 0.922 | 0.561 | 0.244 |
| Closed-grown Douglas-fir | 13 | 136 | 86 | 0.292 | 0.803 | 0.458 | 0.153 |
| Closed-grown Oregon white oak | 57 | 148 | 114 | 0.485 | 0.982 | 0.543 | 0.180 |
| High Density Douglas-fir | 8 | 133 | 110 | 0.574 | 0.906 | 0.195 | 0.211 |
| Low Density Douglas-fir | 8 | 123 | 79 | 0.224 | 0.717 | 0.443 | 0.182 |
| High Density Oregon white oak | 23 | 145 | 113 | 0.398 | 0.943 | 0.566 | 0.165 |
| Low Density Oregon white oak | 22 | 140 | 114 | 0.534 | 0.953 | 0.425 | 0.193 |
| Heterospecific Competition Douglas-fir | 17 | 129 | 91 | 0.494 | 0.951 | 0.388 | 0.167 |
| Heterospecific Competition Oregon white oak | 22 | 148 | 112 | 0.422 | 0.955 | 0.603 | 0.190 |
| Conspecific Competition Douglas-fir | 8 | 136 | 118 | 0.292 | 0.787 | 0.484 | 0.162 |
| Conspecific Competition Oregon white oak | 43 | 277 | 120 | 0.485 | 0.972 | 0.520 | 0.244 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gedalof, Z.; Franks, J.A. Stand Structure and Composition Affect the Drought Sensitivity of Oregon White Oak (Quercus garryana Douglas ex Hook.) and Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco). Forests 2019, 10, 381. https://doi.org/10.3390/f10050381
Gedalof Z, Franks JA. Stand Structure and Composition Affect the Drought Sensitivity of Oregon White Oak (Quercus garryana Douglas ex Hook.) and Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco). Forests. 2019; 10(5):381. https://doi.org/10.3390/f10050381
Chicago/Turabian StyleGedalof, Ze’ev, and Jennifer A. Franks. 2019. "Stand Structure and Composition Affect the Drought Sensitivity of Oregon White Oak (Quercus garryana Douglas ex Hook.) and Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco)" Forests 10, no. 5: 381. https://doi.org/10.3390/f10050381