
Phytophthora Communities Associated with Agathis australis (kauri) in Te Wao Nui o Tiriwa/Waitākere Ranges, New Zealand

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
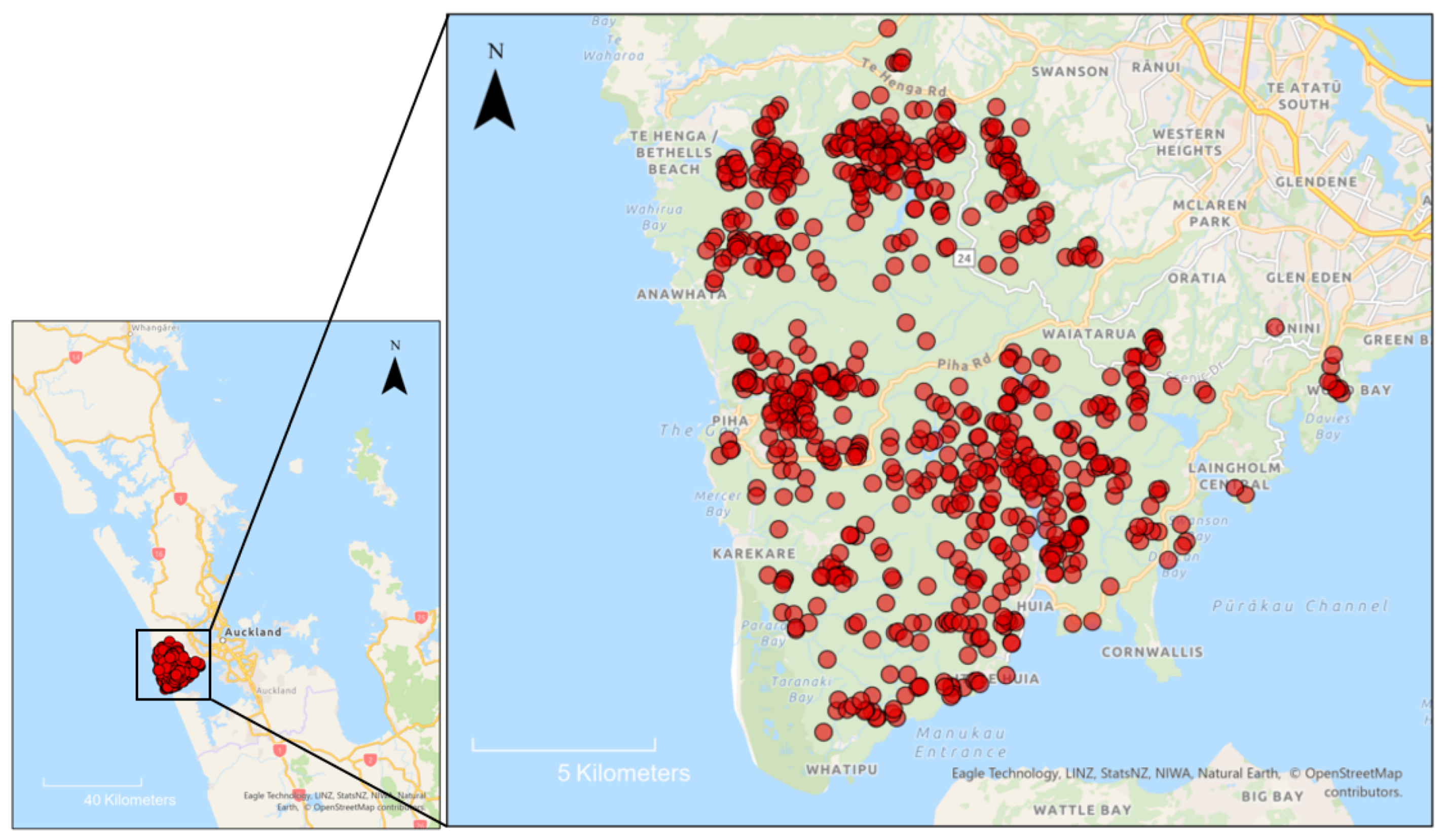
2.1. Site Characteristics and Kauri Dieback History
2.2. Survey Design and Sampling
2.3. Soil Baiting
2.4. Sequencing of Cultures
2.5. Soil DNA Extraction and PCR Amplification
2.6. Controls for Illumina Sequencing of ITS1 Gene Region
2.7. qPCR Validation of Mock Communities for the ITS1 Gene Sequencing
2.8. Bioinformatics
2.9. Data Analyses
3. Results
3.1. Characteristics of Sampled Trees
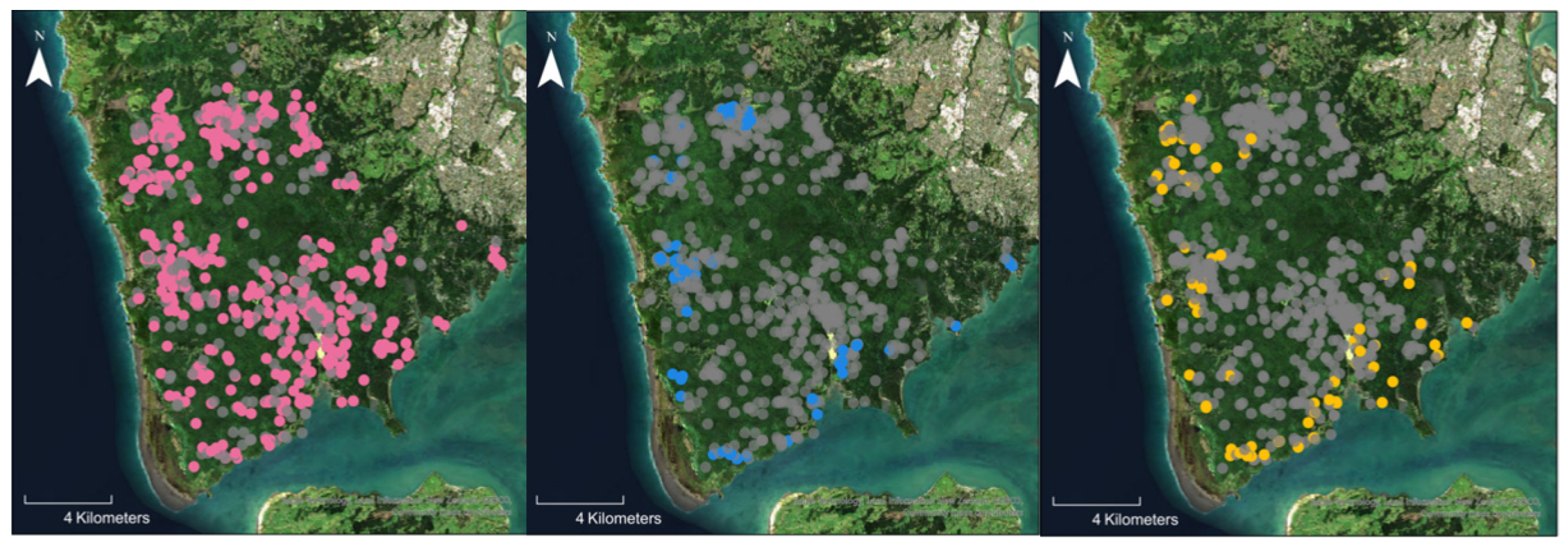
3.2. Identification of Phytophthora Species by Soil Baiting
3.3. Sequencing Output and Performance of Control Reactions
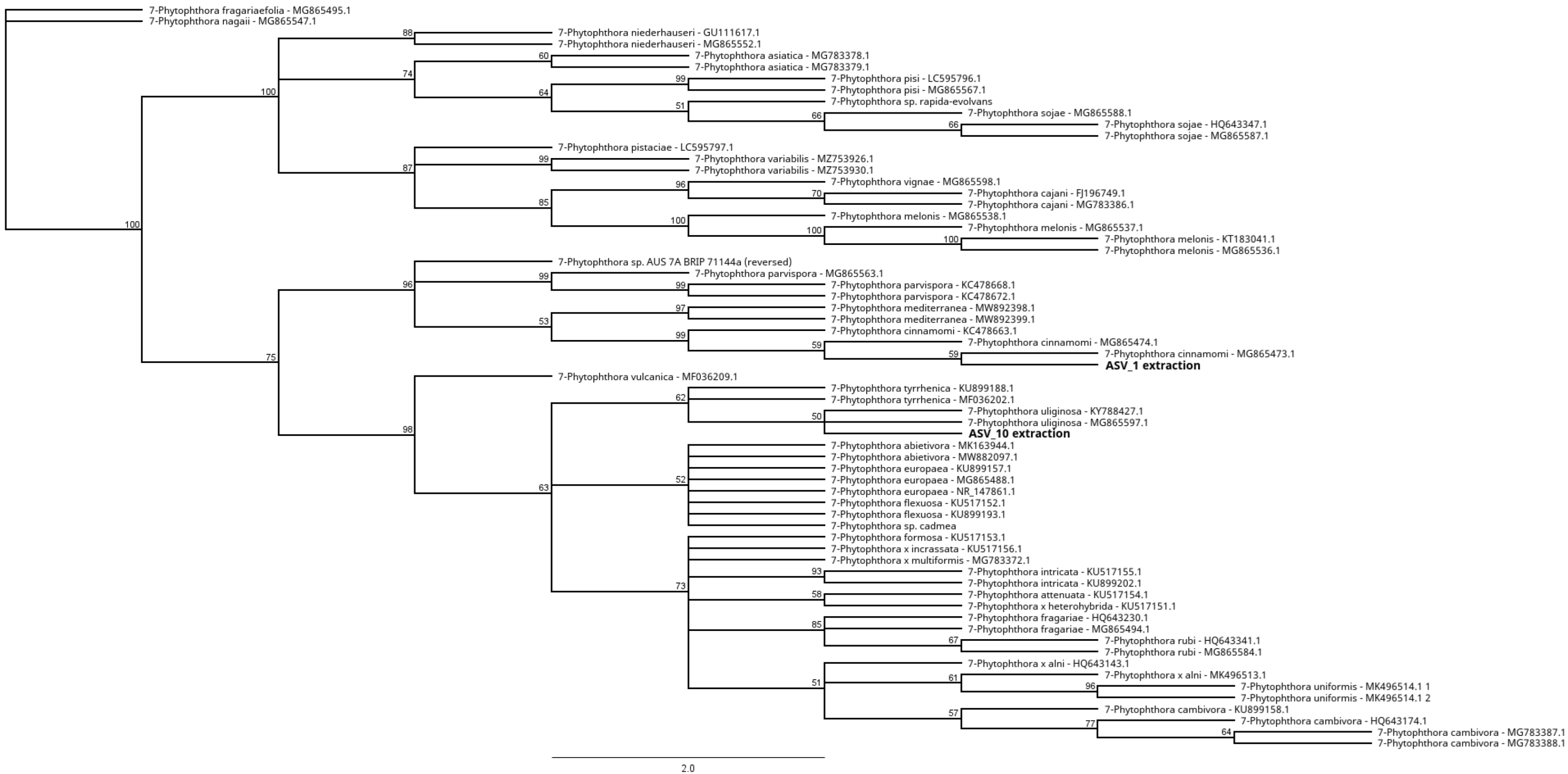
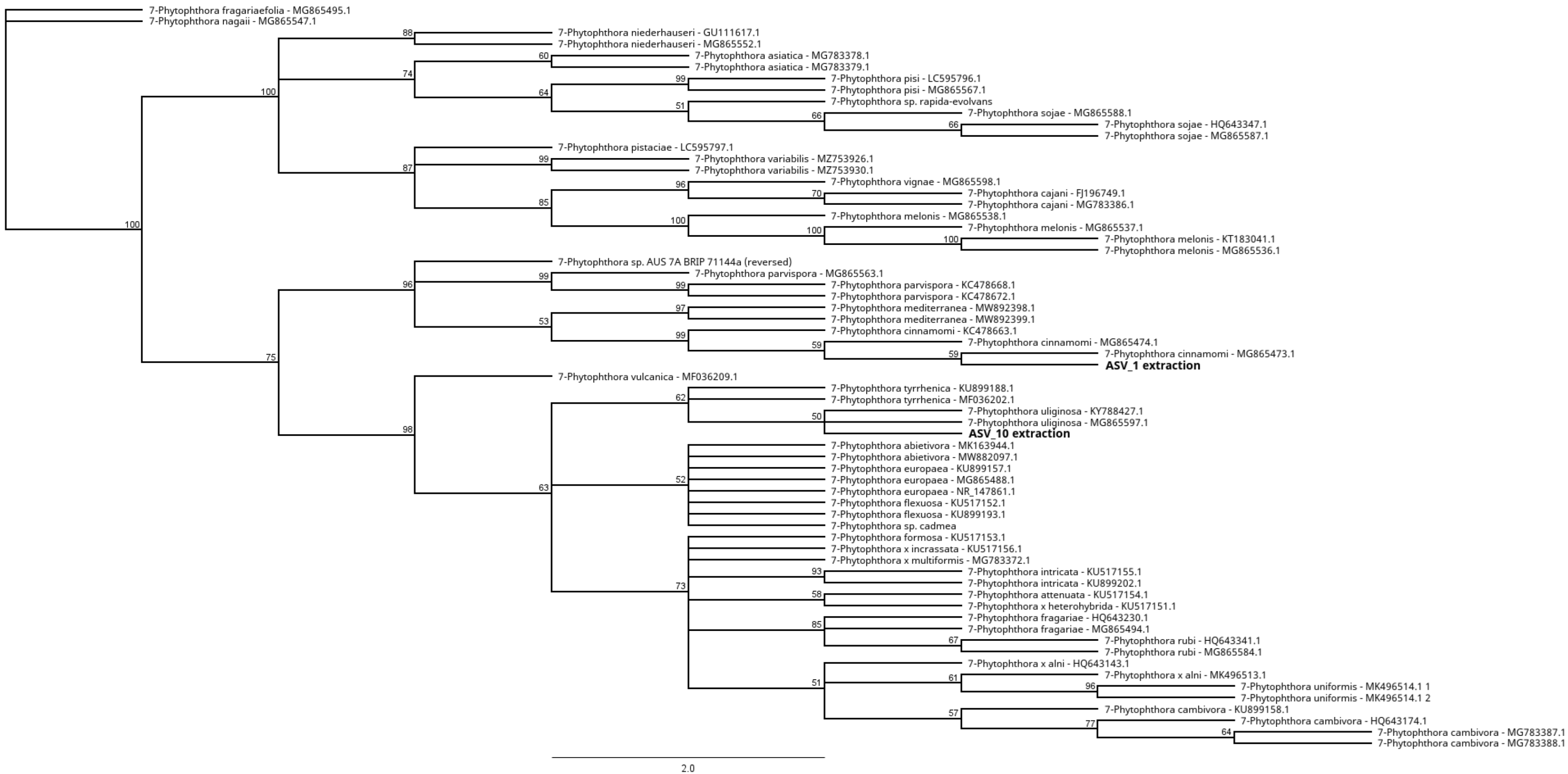
3.4. Identification of Phytophthora Phylotypes by NGS
Unknown Phytophthora Species in Clade 7
3.5. Comparison of Baiting to Metabarcoding
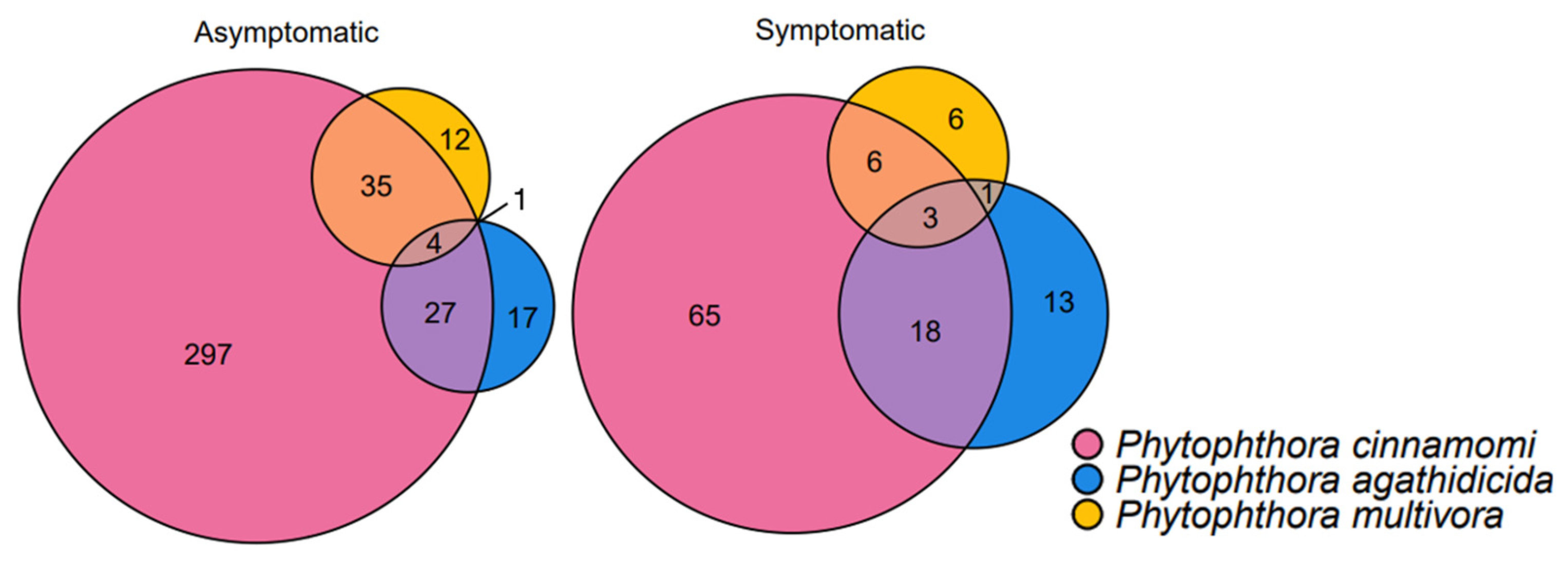
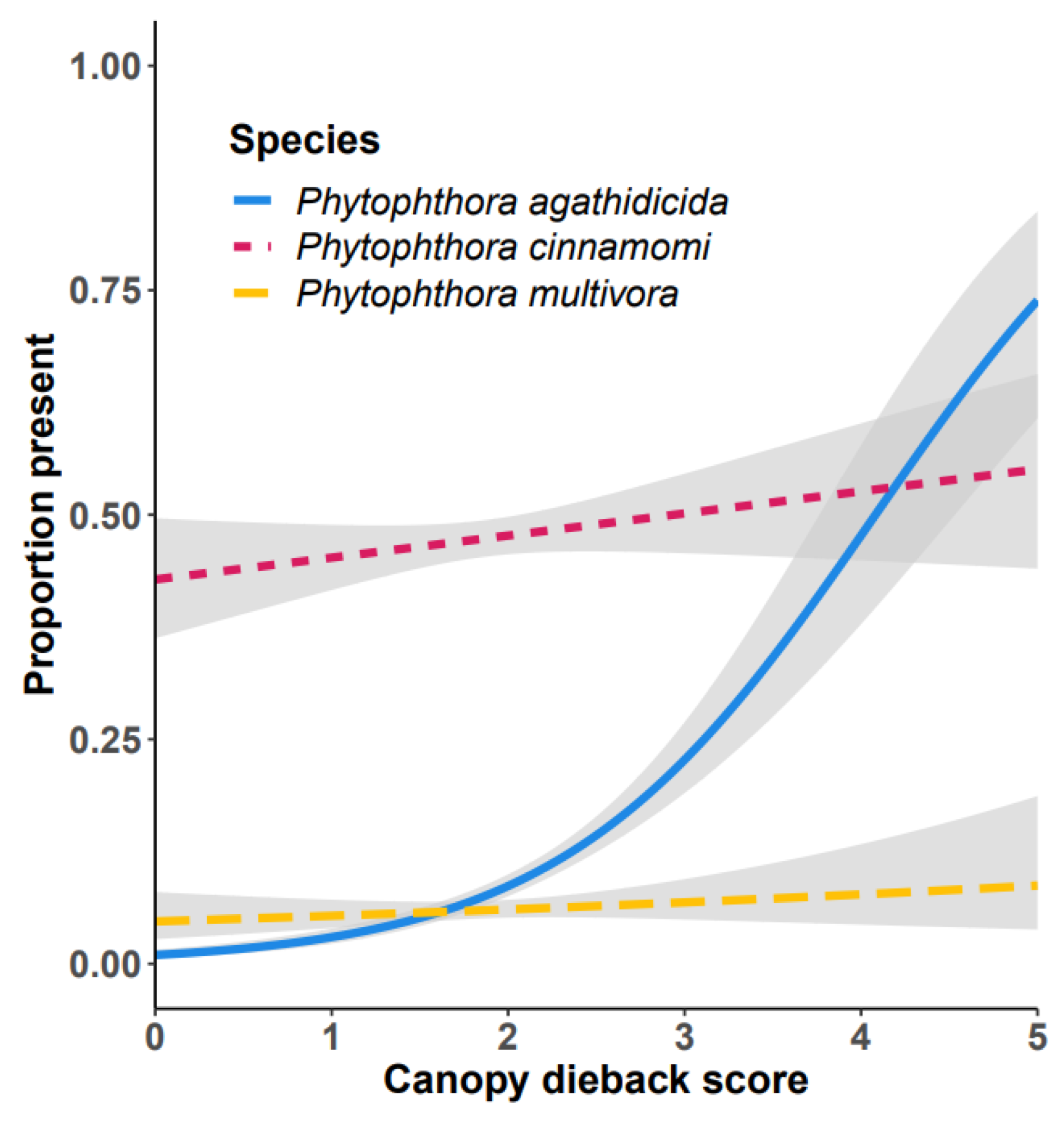
3.6. Phytophthora Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | ASV | Sequence Reads | Database | Similarity | Query Cover | Most Similar Species |
|---|---|---|---|---|---|---|
| 5359 | 24 | 24,937 | ITS1 ref | 100.00% | 100% | Phytophthora_AUS_1A_KY110340 |
| wgs (whole seq) | 93.36% | 100% | Phytophthora taxon totara | |||
| 5480 | 26 | 19,767 | ITS1 ref | 99.50% | 100% | Phytophthora sp. in clade 12A |
| wgs (whole seq) | 99.05% | 100% | P. tubulina | |||
| 5129 | 81 | 3105 | ITS1 ref | 100.00% | 100% | P. gregata/gibbosa/gonapodyides |
| wgs (whole seq) | 98.84% | 100% | P. chlamydospora |
References
- Hansen, E.M. Phytophthora Species Emerging as Pathogens of Forest Trees. Curr. For. Rep. 2015, 1, 16–24. [Google Scholar] [CrossRef]
- Hansen, E.M.; Reeser, P.W.; Sutton, W. Phytophthora beyond agriculture. Annu. Rev. Phytopathol. 2012, 50, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Prospero, S.; Heinz, M.; Augustiny, E.; Chen, Y.Y.; Engelbrecht, J.; Fonti, M.; Hoste, A.; Ruffner, B.; Sigrist, R.; Van Den Berg, N.; et al. Distribution, causal agents, and infection dynamic of emerging ink disease of sweet chestnut in Southern Switzerland. Environ. Microbiol. 2023, 25, 2250–2265. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Jung, M.H.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef]
- Aurangzeb, W.; Guidoni, L.; Rodríguez, C.M.; Cecca, D.; Vannini, A. Exploring the diversity of Phytophthora spp. and the role of Phytophthora multivora in cork and holm oak coastal forests in Italy. Mycol. Prog. 2023, 22, 51. [Google Scholar] [CrossRef]
- Riddell, C.; Frederickson-Matika, D.; Armstrong, A.; Elliot, M.; Forster, J.; Hedley, P.; Morris, J.; Thorpe, P.; Cooke, D.; Pritchard, L.; et al. Metabarcoding reveals a high diversity of woody host-associated Phytophthora spp. in soils at public gardens and amenity woodlands in Britain. PeerJ 2019, 7, e6931. [Google Scholar] [CrossRef] [PubMed]
- Legeay, J.; Husson, C.; Boudier, B.; Louisanna, E.; Baraloto, C.; Schimann, H.; Marcais, B.; Buee, M. Surprising low diversity of the plant pathogen Phytophthora in Amazonian forests. Environ. Microbiol. 2020, 22, 5019–5032. [Google Scholar] [CrossRef] [PubMed]
- Riolo, M.; Aloi, F.; La Spada, F.; Sciandrello, S.; Moricca, S.; Santilli, E.; Pane, A.; Cacciola, S.O. Diversity of Phytophthora Communities across Different Types of Mediterranean Vegetation in a Nature Reserve Area. Forests 2020, 11, 853. [Google Scholar] [CrossRef]
- La Spada, F.; Cock, P.J.A.; Randall, E.; Pane, A.; Cooke, D.E.L.; Cacciola, S.O. DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems. J. Fungi 2022, 8, 330. [Google Scholar] [CrossRef]
- Riit, T.; Cleary, M.; Adamson, K.; Blomquist, M.; Burokienė, D.; Marčiulynienė, D.; Oliva, J.; Poimala, A.; Redondo, M.A.; Strømeng, G.M.; et al. Oomycete Soil Diversity Associated with Betula and Alnus in Forests and Urban Settings in the Nordic–Baltic Region. J. Fungi 2023, 9, 926. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oswald, W. Involvement of soilbourne Phytophthora species in central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Jung, T. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. For. Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Podger, F.D. Phytophthora cinnamomi, a cause of lethal disease in indigenous plant communities in Western Australia. Phytopathology 1972, 62, 972–981. [Google Scholar] [CrossRef]
- Rea, A.J.; Jung, T.; Burgess, T.I.; Stukely, M.J.C.; Hardy, G.E.S.J. Phytophthora elongata sp. nov., a novel pathogen from the Eucalyptus marginata forest of Western Australia. Australas. Plant Path 2010, 39, 477–491. [Google Scholar] [CrossRef]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of Foundation Species: Consequences for the Structure and Dynamics of Forested Ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Weir, B.S.; Paderes, E.P.; Anand, N.; Uchida, J.Y.; Pennycook, S.R.; Bellgard, S.E.; Beever, R.E. A taxonomic revision of Phytophthora Clade 5 including two new species, Phytophthora agathidicida and P. cocois. Phytotaxa 2015, 205, 21. [Google Scholar] [CrossRef]
- Beever, R.E.; Waipara, N.; Ramsfield, T.; Dick, M.; Horner, I. Kauri (Agathis australis) under threat from Phytophthora? In Proceedings of the 4th IUFRO Working Party on Phytophthoras in Forests and Native Ecosystems, Monetrey, CA, USA, 26–31 August 2007; pp. 72–85. [Google Scholar]
- Waipara, N.W.; Hill, S.; Hill, L.M.W.; Hough, E.G.; Horner, I.J. Surveillance methods to determine tree health, distribution of kauri dieback disease and associated pathogens. N. Z. Plant Prot. 2013, 66, 235–241. [Google Scholar] [CrossRef]
- Winkworth, R.C.; Bellgard, S.E.; McLenachan, P.A.; Lockhart, P.J. The mitogenome of Phytophthora agathidicida: Evidence for a not so recent arrival of the “kauri killing” Phytophthora in New Zealand. PLoS ONE 2021, 16, e0250422. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Ogden, J. Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl. (Kauri) in New Zealand I. Population structures and tree growth rates in mature stands. N. Z. J. Bot. 1987, 25, 217–229. [Google Scholar] [CrossRef]
- Steward, G.A.; Beveridge, A.E. A review of New Zealand kauri (Agathis australis (D.Don) Lindl.): Its ecology, history, growth and potential for management for timber. N. Z. J. For. Sci. 2010, 40, 33–59. [Google Scholar]
- Halkett, J.C. A basis for the management of New Zealand kauri (Agathis australis (D. Don) Lindl.) forest. N. Z. J. For. 1983, 28, 15–23. [Google Scholar]
- Black, A.; Waipara, N.W.; Gerth, M. Correspondence: Save Maori people’s sacred tree species. Nature 2018, 561, 177. [Google Scholar] [CrossRef] [PubMed]
- Fadiman, M. Kauri (Agathis australis) ethnobotany: Identity, conservation and connection in New Zealand. Fla. Geogr. 2010, 41, 4–21. [Google Scholar]
- Hill, L.; Stanley, R.; Hammon, C.; Waipara, N. Kauri Dieback Report 2017: An Investigation into the Distribution of Kauri Dieback, and Implications for its Future Management, within the Waitakere Ranges Regional Park; Auckland Council: Auckland, New Zealand, 2017.
- Wyse, S.V.; Burns, B.R.; Wright, S.D. Distinctive vegetation communities are associated with the long-lived conifer Agathis australis (New Zealand kauri, Araucariaceae) in New Zealand rainforests. Austral Ecol. 2014, 39, 388–400. [Google Scholar] [CrossRef]
- Padamsee, M.; Johansen, R.B.; Stuckey, S.A.; Williams, S.E.; Hooker, J.E.; Burns, B.R.; Bellgard, S.E. The arbuscular mycorrhizal fungi colonising roots and root nodules of New Zealand kauri Agathis australis. Fungal Biol. 2016, 120, 807–817. [Google Scholar] [CrossRef]
- Ogden, J. The long-term conservation of forest diversity in New Zealand. Pac. Conserv. Biol. 1995, 2, 77–90. [Google Scholar] [CrossRef]
- Beever, R.E.; Bellgard, S.E.; Dick, M.A.; Horner, I.J.; Ramsfield, T.D. Detection of Phytophthora taxon Agathis; Ministry for Agriculture & Forestry, Biosecurity New Zealand: Wellington, New Zealand, 2010.
- Randall, S.D. Fishing for Phytophthora: A Yearlong Investigation into the Diversity of Phytophthora Species in the Waitakere Ranges, Auckland, NZ. MSc. Ph. D. Thesis, University of Auckland, Auckland, New Zealand, 2011. [Google Scholar]
- Podger, F.D.; Newhook, F.J. Phytophthora cinnamomi in indigenous plant communities in New Zealand. N. Z. J. Bot. 1971, 9, 625–638. [Google Scholar] [CrossRef]
- Horner, I.J. The Role of Phytophthora cinnamomi and other Fungal Pathogens in the Establishment of Kauri and kahikatea; University of Auckland: Auckland, New Zealand, 1984. [Google Scholar]
- NZOR. New Zealand Organisms Register. Available online: https://www.nzor.org.nz/ (accessed on 13 February 2024).
- Horner, I.J.; Hough, E.G. Pathogenicity of four Phytophthora species on kauri: In vitro and glasshouse trials. N. Z. Plant Prot. 2014, 67, 54–59. [Google Scholar] [CrossRef]
- Català, S.; Pérez-Sierra, A.; Abad-Campos, P. The Use of Genus-Specific Amplicon Pyrosequencing to Assess Phytophthora Species Diversity Using eDNA from Soil and Water in Northern Spain. PLoS ONE 2015, 10, e0119311. [Google Scholar] [CrossRef]
- Burgess, T.I.; White, D.; McDougall, K.M.; Garnas, J.; Dunstan, W.A.; Català, S.; Carnegie, A.J.; Worboys, S.; Cahill, D.; Vettraino, A.M.; et al. Distribution and diversity of Phytophthora across Australia. Pac. Conserv. Biol. 2017, 23, 150–162. [Google Scholar] [CrossRef]
- Català, S.; Berbegal, M.; Pérez-Sierra, A.; Abad-Campos, P. Metabarcoding and development of new real-time specific assays reveal Phytophthora species diversity in holm oak forests in eastern Spain. Plant Pathol. 2017, 66, 115–123. [Google Scholar] [CrossRef]
- Khaliq, I.; Hardy, G.; White, D.; Burgess, T.I. eDNA from roots: A robust tool for determining Phytophthora communities in natural ecosystems. FEMS Microbiol. Ecol. 2018, 94, 11. [Google Scholar] [CrossRef] [PubMed]
- Bose, T.; Wingfield, M.J.; Roux, J.; Vivas, M.; Burgess, T.I. Community composition and distribution of Phytophthora species across adjacent native and non-native forests of South Africa. Fungal Ecol. 2018, 36, 17–25. [Google Scholar] [CrossRef]
- Vannini, A.; Bruni, N.; Tomassini, A.; Franceschini, S.; Vettraino, A.M. Pyrosequencing of environmental soil samples reveals biodiversity of the Phytophthora resident community in chestnut forests. FEMS Microbiol. Ecol. 2013, 85, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Byers, A.K.; Condron, L.; Donavan, T.; O’Callaghan, M.; Patuawa, T.; Waipara, N.; Black, A. Soil microbial diversity in adjacent forest systems—Contrasting native, old growth kauri (Agathis australis) forest with exotic pine (Pinus radiata) plantation forest. FEMS Microbiol. Ecol. 2020, 96, 12. [Google Scholar] [CrossRef] [PubMed]
- Jongkind, A.G.; Buurman, P. The effect of kauri (Agathis australis) on grain size distribution and clay mineralogy of andesitic soils in the Waitakere Ranges, New Zealand. Geoderma 2006, 134, 171–186. [Google Scholar] [CrossRef]
- Froud, K.; Chew, Y.C.; Kean, J.; Jae, M.; Killick, S.; Ashby, E.; Taua-Gordon, R.; Jamieson, A.; Tolich, L. 2021 Waitakere Ranges Kauri Population Health Monitoring Survey; Auckland Council Technical Report; Auckland Council: Auckland, New Zealand, 2022.
- Dick, M.; Bellgard, S.E. Soil survey method for Phytophthora taxon Agathis. In Report Prepared for the Ministry of Agriculture and Forestry on behalf of Kauri Dieback Joint Agency, Version 2.1; Beauchamp, A.J., Ed.; Department of Conservation: Wellington, New Zealand, 2012; p. 13. [Google Scholar]
- O’Brien, P.A.; Williams, N.; Hardy, G.E.S. Detecting Phytophthora. Crit. Rev. Microbiol. 2009, 35, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996. [Google Scholar]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. 1990, 31, 315–322. [Google Scholar]
- Robideau, G.P.; De Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef]
- Legeay, J.; Husson, C.; Cordier, T.; Vacher, C.; Marcais, B.; Buée, M. Comparison and validation of Oomycetes metabarcoding primers for Phytophthora high throughput sequencing. J. Plant Pathol. 2019, 101, 743–748. [Google Scholar] [CrossRef]
- Scibetta, S.; Schena, L.; Chimento, A.; Cacciola, S.O.; Cooke, D.E.L. A molecular method to assess Phytophthora diversity in environmental samples. J. Microbiol. Meth. 2012, 88, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.N.; Coffey, M.D. Mitochondrial Haplotype Analysis for Differentiation of Isolates of Phytophthora Cinnamomi. Phytopathology 2012, 102, 229–239. [Google Scholar] [CrossRef]
- Foster, Z.S.L.; Albornoz, F.E.; Fieland, V.J.; Larsen, M.M.; Jones, F.A.; Tyler, B.M.; Nguyen, H.D.T.; Burgess, T.I.; Riddell, C.; Voglmayr, H.; et al. A New Oomycete Metabarcoding Method Using the rps10 Gene. Phytobiomes J. 2022, 6, 214–226. [Google Scholar] [CrossRef]
- Illumina. llumina 16S Metagenomic Sequencing Library Preparation (IlluminaTechnical Note 15044223). 2013. Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 24 January 2024).
- McDougal, R.; Bellgard, S.E.; Scott, P.; Ganley, B. Comparison of a Real Time PCR Assay and a Soil Bioassay Technique for Detection of Phytophthora taxon Agathis from Soil; Scion: Rotorua, New Zealand, 2014. [Google Scholar]
- Than, D.; Hughes, K.; Boonhan, N.; Tomlinson, J.; Woodhall, J.; Bellgard, S. A TaqMan real-time PCR assay for the detection of Phytophthora ‘taxon Agathis’ in soil, pathogen of Kauri in New Zealand. For. Pathol. 2013, 43, 324–330. [Google Scholar] [CrossRef]
- Kunadiya, M.; White, D.; Dunstan, W.A.; St, G.E.; Hardy, J.; Andjic, V.; Burgess, T.I. Pathways to false positive diagnoses using molecular genetic detection methods; Phytophthora cinnamomi a case study. Fems Microbiol. Lett. 2017, 364, fnx009. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Version 0.12.0. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 20 February 2024).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Venables, B.; Ripley, B. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Zuur, A.F.; Ieno, E.N. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 2016, 7, 636–645. [Google Scholar] [CrossRef]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. Code Snippets 2016, 69, 1–17. [Google Scholar] [CrossRef]
- Larsson, J. eulerr: Area-Proportional Euler and Venn Diagrams with Ellipses. R Package Version 7.0.0. 2022. Available online: https://rdrr.io/cran/eulerr/ (accessed on 20 February 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Gardner, J.F.; Dick, M.A.; Bader, M.K.-F. Susceptibility of New Zealand flora to Phytophthora kernoviae and its seasonal variability in the field. N. Z. J. For. Sci. 2015, 45, 23. [Google Scholar] [CrossRef]
- Scott, P.; Williams, N. Phytophthora diseases in New Zealand forests. N. Z. J. For. 2014, 59, 14–21. [Google Scholar]
- Bregant, C.; Batista, E.; Hilário, S.; Linaldeddu, B.T.; Alves, A. Phytophthora Species Involved in Alnus glutinosa Decline in Portugal. Pathogens 2023, 12, 276. [Google Scholar] [CrossRef]
- Seddaiu, S.; Brandano, A.; Ruiu, P.A.; Sechi, C.; Scanu, B. An overview of Phytophthora species inhabiting declining Quercus suber stands in Sardinia (Italy). Forests 2020, 11, 971. [Google Scholar] [CrossRef]
- Corcobado, T.; Cech, T.L.; Daxer, A.; Ďatková, H.; Janoušek, J.; Patra, S.; Jahn, D.; Hüttler, C.; Milenković, I.; Tomšovský, M.; et al. Phytophthora, Nothophytophthora and Halophytophthora diversity in rivers, streams and riparian alder ecosystems of Central Europe. Mycol. Prog. 2023, 22, 50. [Google Scholar] [CrossRef] [PubMed]
- Greslebin, A.G.; Hansen, E.M.; Winton, L.M.; Rajchenberg, M. Phytophthora species from declining Austrocedrus chilensis forests in Patagonia, Argentina. Mycologia 2005, 97, 218–228. [Google Scholar] [CrossRef]
- Lyubenova, A.; Kostov, K.; Tsvetkov, I.; Slavov, S. Possible contribution of Phytophthora spp. in decline of Norway spruce (P. abies (L.) H. Karst.) stands in Vitosha Mountain. Silva Balc. 2014, 15, 100–109. [Google Scholar]
- Riddell, C.E.; Dun, H.F.; Elliot, M.; Armstrong, A.C.; Clark, M.; Forster, J.; Hedley, P.E.; Green, S. Detection and spread of Phytophthora austrocedri within infected Juniperus communis woodland and diversity of co-associated Phytophthoras as revealed by metabarcoding. For. Pathol. 2020, 50, e12602. [Google Scholar] [CrossRef]
- Newhook, F.J. The association of Phytophthora spp. with mortality of Pinus radiata and other conifers. N. Z. J. Agric. Res. 1959, 2, 808–843. [Google Scholar] [CrossRef]
- Brien, R.M.; Dingley, J.M. Fourth supplement to “A revised list of plant diseases recorded in New Zealand”, 1957–1958. N. Z. J. Agric. Res. 1959, 2, 406–413. [Google Scholar] [CrossRef]
- Burgess, T.I.; Scott, J.K.; McDougall, K.L.; Stukely, M.J.C.; Crane, C.; Dunstan, W.A.; Brigg, F.; Andjic, V.; White, D.; Rudman, T.; et al. Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Glob. Chang. Biol. 2017, 23, 1661–1674. [Google Scholar] [CrossRef]
- Scott, P.M.; Burgess, T.I.; Barber, P.A.; Shearer, B.L.; Stukely, M.J.C.; Hardy, G.E.S.; Jung, T. Phytophthora multivora sp nov., a new species recovered from declining Eucalyptus, Banksia, Agonis and other plant species in Western Australia. Persoonia 2009, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tsykun, T.; Prospero, S.; Schoebel, C.N.; Rea, A.; Burgess, T.I. Global invasion history of the emerging plant pathogen Phytophthora Multivora. BMC Genom. 2022, 23, 153. [Google Scholar] [CrossRef] [PubMed]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi. Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I.; White, D.; Sapsford, S.J. Comparison of Primers for the Detection of Phytophthora (and Other Oomycetes) from Environmental Samples. J. Fungi 2022, 8, 980. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.R.; McComb, J.; Burgess, T.I.; Hardy, G.E.S.J. Timing and abundance of sporangia production and zoospore release influences the recovery of different Phytophthora species by baiting. Fungal Biol. 2021, 125, 477–484. [Google Scholar] [CrossRef]
- Ruiz Gómez, F.J.; Pérez-de-Luque, A.; Sánchez-Cuesta, R.; Quero, J.L.; Navarro Cerrillo, R.M. Differences in the Response to Acute Drought and Phytophthora cinnamomi Rands Infection in Quercus ilex L. Seedlings. Forests 2018, 9, 634. [Google Scholar] [CrossRef]



| Primer or Probe Name | Target Species | Sequence (5′ to 3′) | Notes * | References |
|---|---|---|---|---|
| ITS_PTA_F2 | P. agathidicida | AACCAATAGTTGGGGGCGA | [54,55] | |
| ITS_PTA_R3 | CTCGCCATGATAGAGCTCGTC | [54] | ||
| ITS_PTA_probe2 | AGCCAAAGCCAGCAGCCG | 5′ FAM, 3′ BHQ1 | [54] | |
| PCIN F6 | P. cinnamomi | CGTGGCGGGCCCTATC | [56] | |
| PCIN R2 | AAAAGAGAGGCTACTAGCTCAGTTCCC | [56] | ||
| PCIN probe | TGGCGAGCGTTTGGGTCCCTCT | 5′ HEX, 3′ BHQ2 | [56] |
| Phytophthora Species | Clade | Soil Baiting | Metabarcoding (Soil eDNA) | Total Detections | Total Detections (%) |
|---|---|---|---|---|---|
| P. cinnamomi | 7 | 404 | 231 | 455 | 59.3 |
| P. agathidicida | 5 | 79 | 44 | 84 | 11.0 |
| P. multivora | 2 | 63 | 6 | 68 | 8.9 |
| P. sp. (P. europaea-like) | 7 | 0 | 20 | 20 | 2.6 |
| P. cactorum/P. aleatoria | 1 | 0 | 4 | 4 | 0.5 |
| P. pseudocryptogea/P. cryptogea | 8 | 1 | 2 | 2 | 0.3 |
| P. kernoviae | 10b | 0 | 2 | 2 | 0.3 |
| Reads (%) * | |||||
|---|---|---|---|---|---|
| Sample ID | DNA (ng/µL) | P. cinnamomi (H1454) | P. agathidicida (H1453) | P. pseudocryptogea (H1258) | P. multivora (H1467) |
| 6000 | 0.1 | 53.3 | 16.0 | 27.9 | 0.0 |
| 6001 | 0.01 | 51.0 | 19.4 | 27.4 | 0.0 |
| 6002 | 0.001 | 55.9 | 15.3 | 26.8 | 0.7 |
| 6011 | 0.0001 | 60.7 | 2.8 | 33.4 | 0.0 |
| 6003 | 0.01 | - | 0.3 | - | - |
| 0.1 | 58.3 | - | 38.2 | 0.0 | |
| 6004 | 0.001 | - | 31.4 | - | - |
| 0.1 | 16.5 | - | 48.7 | 1.1 | |
| 6005 | 0.01 | 3.3 | - | - | - |
| 0.1 | - | 37.5 | 57.6 | 1.0 | |
| 6006 | 0.001 | 69.6 | - | - | - |
| 0.1 | - | 19.2 | 7.0 | 0.0 | |
| 6007 | 0.01 | - | - | 0.7 | - |
| 0.1 | 71.0 | 22.8 | - | 0.3 | |
| 6008 | 0.001 | - | - | 30.2 | - |
| 0.1 | 52.0 | 17.5 | - | 0.0 | |
| 6009 | 0.01 | - | - | - | 0.0 |
| 0.1 | 53.2 | 17.1 | 29.7 | - | |
| 6010 | 0.001 | - | - | - | 0.0 |
| 0.1 | 62.2 | 9.1 | 26.6 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunter, S.; Horner, I.; Hosking, J.; Carroll, E.; Newland, J.; Arnet, M.; Waipara, N.; Burns, B.; Scott, P.; Williams, N. Phytophthora Communities Associated with Agathis australis (kauri) in Te Wao Nui o Tiriwa/Waitākere Ranges, New Zealand. Forests 2024, 15, 735. https://doi.org/10.3390/f15050735
Hunter S, Horner I, Hosking J, Carroll E, Newland J, Arnet M, Waipara N, Burns B, Scott P, Williams N. Phytophthora Communities Associated with Agathis australis (kauri) in Te Wao Nui o Tiriwa/Waitākere Ranges, New Zealand. Forests. 2024; 15(5):735. https://doi.org/10.3390/f15050735
Chicago/Turabian StyleHunter, Shannon, Ian Horner, Jack Hosking, Ellena Carroll, Jayne Newland, Matthew Arnet, Nick Waipara, Bruce Burns, Peter Scott, and Nari Williams. 2024. "Phytophthora Communities Associated with Agathis australis (kauri) in Te Wao Nui o Tiriwa/Waitākere Ranges, New Zealand" Forests 15, no. 5: 735. https://doi.org/10.3390/f15050735