
Mechanism of Alleviating Acute Lung Injury in Mice from Serum Metabolomics Analysis of Cordyceps fumosorosea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Testing Samples
2.2. Preparation of C. fumosorosea Mycelium Solution
2.3. Grouping and Administration
2.4. Acute Lung Injury Induced by Lipopolysaccharides
2.5. Ratio of Wet and Dry Weight
2.6. Hematoxylin and Eosin Stains
2.7. Microscopic Observation
2.8. Extraction of Metabolite from Serum
2.9. UPLC-MS Analysis
2.10. Extraction and Identification Metabolite Ion Peak
3. Statistical Analysis
4. Results
4.1. Lung Wet/Dry Weight
4.2. Hematoxylin and Eosin
4.3. Transmission Electron Microscopy (TEM) Observation
4.4. Extraction and Identification Metabolite Ion Peak (UPLC-MS/MS)
4.5. Coefficient of Variation Analysis (CV)
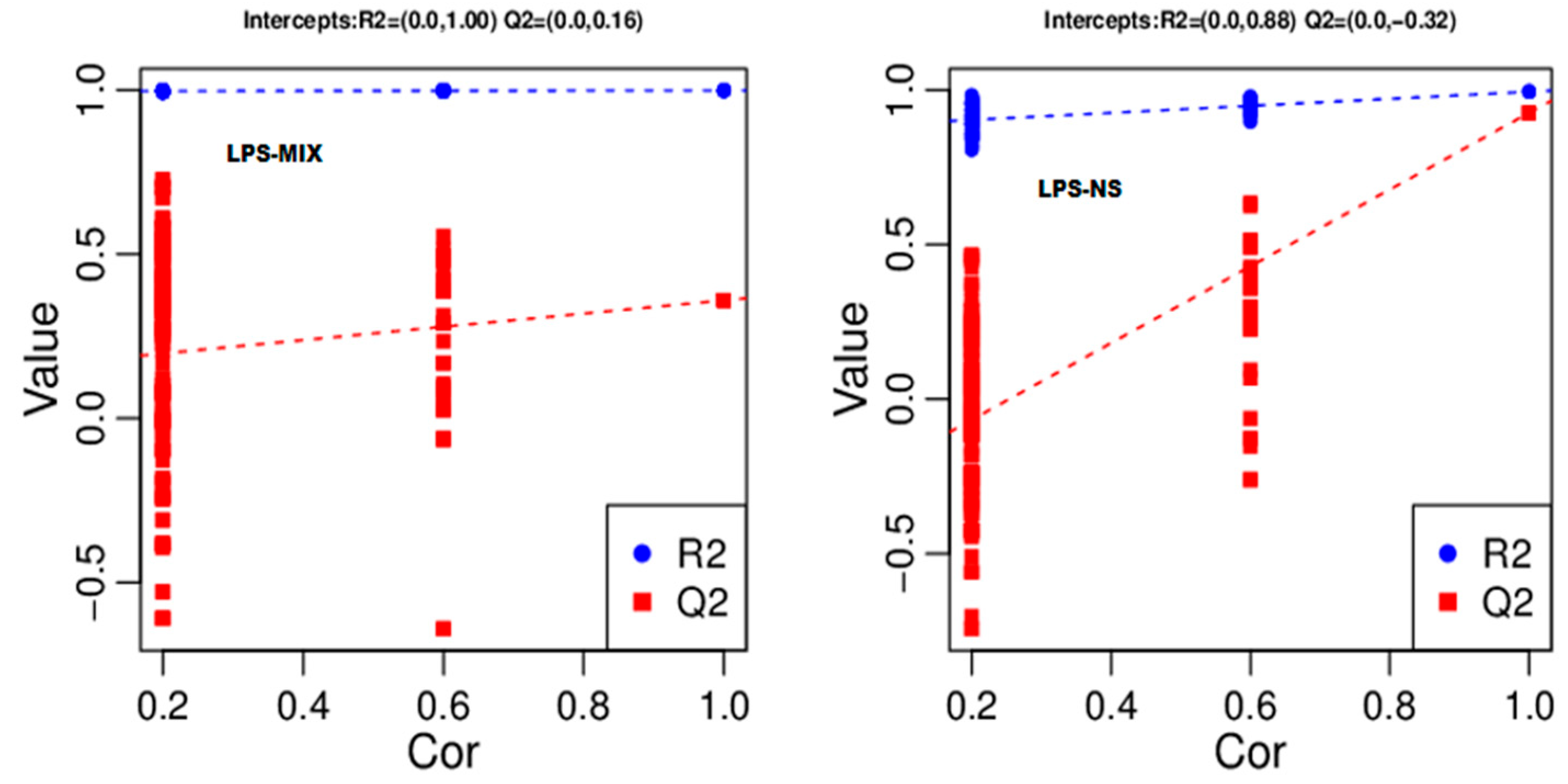
4.6. Partial Least Squares Discriminant Analysis (PLS-DA) Analysis
4.7. Metabolic Pathway Enrichment Analysis
4.8. Principal Component Analysis
4.9. Orthogonal Partial Least Squares Discriminant Analysis (OPLS-DA) Analysis
4.10. Identification of Metabolites Classes and Sub-Classes
4.11. Metabolic Pathway Analysis
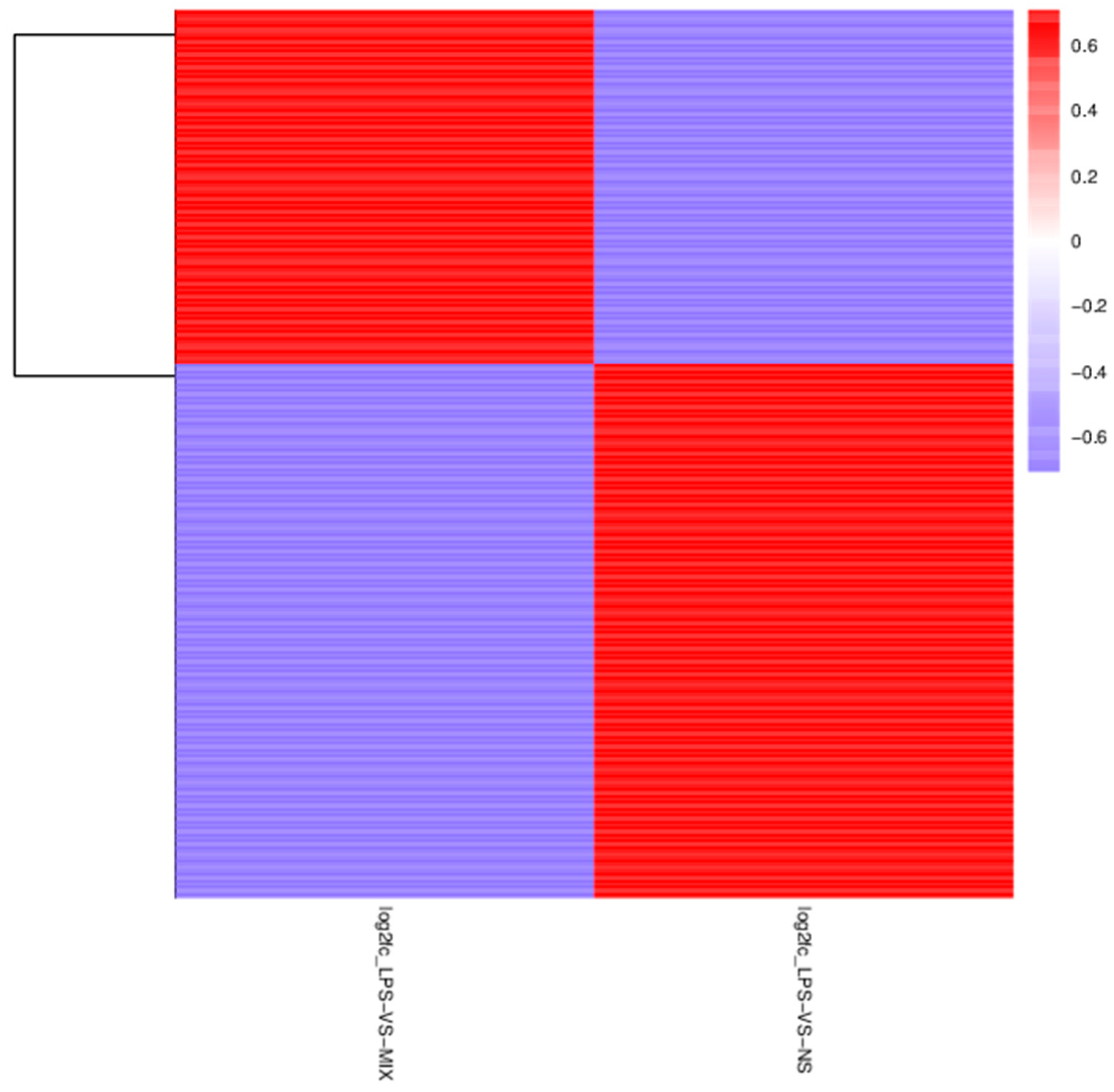
4.12. Identification of Differentially Accumulated Metabolites (DAMs)
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ping, L.; Haiping, H.; Ge, L. Bioanalysis of traditional Chinese medicines. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1026, 1. [Google Scholar]
- Graham, D.T. Regulation of proprietary traditional Chinese medicines in Australia. Chin. J. Nat. Med. 2017, 15, 12–14. [Google Scholar] [CrossRef]
- Chan, K. Chinese medicinal materials and their interface with Western medical concepts. J. Ethnopharmacol. 2005, 96, 1–18. [Google Scholar] [CrossRef]
- Coudroy, R.; Lu, C.; Chen, L.; Demoule, A.; Brochard, L. Mechanism of airway closure in acute respiratory distress syndrome: A possible role of surfactant depletion. Intens. Care Med. 2019, 45, 290–291. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, T.; Miyoshi, S.; Yamazaki, S.; Nishina, T.; Mikami, T.; Oikawa, A.; Homma, S.; Nakano, H. A murine model of acute lung injury identifies growth factors to promote tissue repair and their biomarkers. Genes Cells 2019, 24, 112–125. [Google Scholar] [CrossRef] [PubMed]
- LV, Q. Glucocorticoid combined with hyaluronic acid enhances glucocorticoid receptor activity through inhibiting p-38MAPK signal pathway activation in treating acute lung injury in rats. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3920–3929. [Google Scholar]
- Han, S.C.; Cai, W.X.; Yang, X.K.; Jia, Y.; Zheng, Z.; Wang, H.; Li, J.L.; Ygao, J.; Fan, L.; Hu, D. ROS-mediated NLRP3 inflammasome activity is essential for burn-induced acute lung injury. Mediat. Inflamm. 2015, 2015, 720457. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.L.; Zhang, X.S.; Xue, X.; Wang, R.M. Patchouli alcohol protects against lipopolysaccharide induced acute lung injury in mice. J. Surg. Res. 2015, 194, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Marutani, E.; Shin, H.; Chen, W.; Hanaoka, K.; Xian, M.; Ichinose, F. Sodium thiosulfate attenuates acute lung injury in mice. Anesthesiology 2014, 121, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, F.; Liu, H.; Cheng, H.; Wang, W.; Wen, Q.; Wang, Y. Urinary trypsin inhibitor attenuates lipopolysaccharide-induced acute lung injury by blocking the activation of p38 mitogen-activated protein kinase. Inflamm. Res. 2011, 60, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Garbi, N.; Lambrecht, B.N. Location, function, and ontogeny of pulmonary macrophages during the steady state. Pflug. Arch. 2017, 469, 561–572. [Google Scholar] [CrossRef]
- Florentin, J.; Dutta, P. Origin and production of inflammatory perivascular macrophages in pulmonary hypertension. Cytokine 2017, 100, 11–15. [Google Scholar] [CrossRef]
- Wang, F.; Wang, R.; Liu, H. The acute pulmonary toxicity in mice induced by Staphylococcus aureus, particulate matter, and their combination. Exp. Anim. 2018, 68, 159–168. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, K.; Liao, L.; Zhang, T.; Yang, M.; Sun, C. Lipoxin A4 receptor agonist BML-111 induces autophagy in alveolar macrophages and protects from acute lung injury by activating MAPK signaling. Respir. Res. 2018, 19, 243. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.L.; Ortolan, L.S.; Sercundes, M.K.; Debone, D.; Murillo, O.; Lima, F.A.; Marinho, C.R.F.; Epiphanio, S. Association of heme oxygenase 1 with lung protection in malaria-associated ALI/ARDS. Mediat. Inflamm. 2016, 2016, 4158698. [Google Scholar] [CrossRef]
- Li, H.; Zhou, X.; Tan, H.; Hu, Y.; Zhang, L.; Liu, S.; Dai, M.; Li, Y.; Li, Q.; Mao, Z.; et al. Neutrophil extracellular traps contribute to the pathogenesis of acid-aspiration-induced ALI/ARDS. Oncotarget 2018, 9, 1772–1784. [Google Scholar] [CrossRef]
- Shi, W. The medical research advanced on Periplaneta americana. Chin. J. Ethnomed. Ethnopharm. 2012, 21, 50–51. [Google Scholar]
- Zhao, Y.; Yang, A.; Tu, P.; Hu, Z. Anti-tumor effects of the American cockroach, Periplaneta americana. Chin. Med. 2017, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Peng, F.; Song, L.; Wang, X.; Hu, M.; Zhao, Y.; Liu, G. Advances in chemical constituents and pharmacological effects of Periplaneta americana. Chin. J. Chin. Mater. Med. 2007, 32, 2326–2330. [Google Scholar]
- Khan, T.; Hou, D.H.; Zhou, J.N.; Yang, Y.L.; Yu, H. Effect of Abiotic Factors on Fumosorinone Production from Cordyceps fumosorosea via Solid-State Fermentation. Mycobiology 2023, 51, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Jennifer, L.A.J.; Hywel, J.N.L. A phylogenetically based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef]
- Ali, S.; Wu, J.H.; Huang, Z. Production and regulation of extracellular chitinase from the entomopathogenic fungus Isaria fumosorosea. Biocontrol Sci. Technol. 2010, 20, 723–738. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Production of cuticle degrading enzymes by Isaria fumosorosea and their evaluation as a biocontrol agent against diamondback moth. J. Pest. Sci. 2010, 83, 361–370. [Google Scholar] [CrossRef]
- Banu, A.N.; Balasubramanian, C. Optimization synthesis of silver nanoparticles using Isaria fumosorosea against human vector mosquitoes. Parasitol. Res. 2014, 113, 3843–3851. [Google Scholar] [CrossRef]
- Pick, D.A.; Avery, P.B.; Hunter, W.B.; Powell, C.A.; Arthurs, S.P. Effect of Isaria fumosorosea (Hypocreales: Cordycipitaceae) and Lysiphlebus testaceipes, (Hymenoptera: Braconidae) on the brown citrus aphid: Preliminary assessment of a compatibility study. Fla. Entomol. 2012, 95, 764–766. [Google Scholar] [CrossRef]
- Huang, Z.; Ali, S.; Ren, S.X.; Wu, J.H. The effect of Isaria fumosorosea on mortality and fecundity of Bemisia tabaci and Plutella xylostella. Insect Sci. 2010, 17, 140–148. [Google Scholar] [CrossRef]
- Khan, T.; Dong-Hai, H.; Zhou, J.N.; Yang, Y.L.; Yu, H. Comprehensive analysis of metabolites in the mycelium of Cordyceps fumosorosea cultured on Periplaneta americana. Ann. Microbiol. 2024, 74, 9. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.-F.; Fan, Q.; Wang, Y.-B.; Yu, H. Cordycipitoid Fungi Powders Promote Mycelial Growth and Bioactive-Metabolite Production in Liquid Cultures of the Stout Camphor Medicinal Mushroom Taiwanofungus camphoratus (Agaricomycetes). Int. J. Med. Mushrooms 2020, 22, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Pi, N.; Guo, Y.; He, X.; Wang, J.; Luo, R.; Wang, M.; Yu, H. The mechanism of action of Ophiocordyceps sinensis mycelia for prevention of acute lung injury based on non-targeted serum metabolomics. PLoS ONE 2023, 18, e0287331. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. Acute lung injury in animals study g. An official american thoracic society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zou, W.; Wang, J.; Luo, R.; Wang, M.; Yu, H. Comparison of widely targeted metabolomics and untargeted metabolomics of wild Ophiocordyceps sinensis. Molecules 2022, 27, 3645. [Google Scholar] [CrossRef] [PubMed]
- Keshari, R.S.; Silasi-Mansat, R.; Zhu, H.; Popescu, N.I.; Peer, G.; Chaaban, H.; Lambris, J.D.; Polf, H.; Lupu, C.; Kinasewitz, G. Acute lung injury and fibrosis in a baboon model of Escherichia coli sepsis. Am. J. Respir. Cell Mol. Biol. 2014, 50, 439–450. [Google Scholar] [CrossRef]
- Cox, R.J.; Phillips, O.; Fukumoto, J.; Fukumoto, I.; Parthasarathy, P.T.; Arias, S. Enhanced Resolution of Hyperoxic Acute Lung Injury as a Result of Aspirin-Triggered Resolvin D1 Treatment. Am. J. Respir. Cell Mol. Biol. 2015, 53, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Manjunathaiah, R.N.; Ramakrishna, K.; Sirisha, V.; Divya, P.; Venkateswara Rao, A. Computer aided discovery of potential anti-inflammatory (s)-naproxen analogs as COX-2 inhibitors. Med. Chem. 2013, 9, 553–559. [Google Scholar] [CrossRef]
- Hasan, D.; Shono, A.; van Kalken, C.K.; van der Spek, P.J.; Krenning, E.P.; Kotani, T. A novel definition and treatment of hyperinflammation in COVID-19 based on purinergic signalling. Purinergic Signal 2022, 18, 13–59. [Google Scholar] [CrossRef]
- Vollbracht, C.; Kraft, K. Oxidative stress and hyper-inflammation as major drivers of severe COVID-19 and long COVID: Implications for the benefit of high-dose intravenous vitamin C. Front. Pharmacol. 2022, 13, 899198. [Google Scholar] [CrossRef]
- Anwar, S.; Almatroudi, A.; Allemailem, K.S.; Jacob Joseph, R.; Khan, A.A.; Rahmani, A.H. Protective effects of ginger extract against glycation and oxidative stress-induced health complications: An in vitro study. Processes 2020, 8, 468. [Google Scholar] [CrossRef]
- Guida, D.R.; Engel, J.; Allwood, J.W.; Weber, R.J.; Jones, M.R.; Sommer, U.; Viant, M.R.; Dunn, W.B. Non-targeted UHPLC-MS metabolomic data processing methods: A comparative investigation of normalisation, missing value imputation, transformation and scaling. Metabolomics 2016, 12, 93. [Google Scholar] [CrossRef]
- Sureshbabu, A.; Syed, M.; Das, P.; Janer, C.; Pryhuber, G.; Rahman, A.; Andersson, S.; Homer, R.J.; Bhandari, V. Inhibition of Regulatory-Associated Protein of Mechanistic Target of Rapamycin Prevents Hyperoxia-Induced Lung Injury by Enhancing Autophagy and Reducing Apoptosis in Neonatal Mice. Am. J. Respir. Cell Mol. Biol. 2016, 55, 722–735. [Google Scholar] [CrossRef]
- Kasotakis, G.; Purves, J.T.; Agarwal, S.; Sullenger, B.A.; Kintsurashvili, E.; Galvan, M.D.; Graham, C.; Corcoran, D.L.; Palmer, S.M.; Remick, D.G. Histone Deacetylase 7 in Murine Gram-Negative Acute Lung Injury. Shock 2020, 53, 375–377. [Google Scholar] [CrossRef]
- Canadas, O.; Keough, K.M.; Casals, C. Bacterial lipopolysaccharide promotes destabilization of lung surfactant–like films. Biophys. J. 2011, 100, 108–116. [Google Scholar] [CrossRef]
- Hiansen, J.Q.; Keating, E.; Aspros, A.; Yao, L.J.; Bosma, K.J.; Yamashita, C.M.; Lewis, J.F.; Veldhuizen, R.A. Cholesterol–mediated surfactant dysfunction is mitigated by surfactant protein A. Biochim. Biophys. Acta 2015, 1848, 813–820. [Google Scholar] [CrossRef]
- Lau, C.; Abdelmalak, B.B.; Farver, C.F.; Culver, D.A. Whole lung lavage for lipoid pneumonia. Thorax 2016, 71, 1066–1067. [Google Scholar] [CrossRef]
- Ying, J.F.; Lu, Z.B.; Fu, L.Q.; Tong, Y.; Wang, Z.; Li, W.F.; Mou, X.Z. The role of iron homeostasis and iron-mediated ROS in cancer. Am. J. Cancer Res. 2021, 11, 1895–1912. [Google Scholar] [PubMed]
- Han, X.; Wu, Y.C.; Meng, M.; Sun, Q.S.; Gao, S.M.; Sun, H. Linarin prevents LPS-induced acute lung injury by suppressing oxidative stress and inflammation via inhibition of TXNIP/NLRP3 and NF-κB pathways. Int. J. Mol. Med. 2018, 42, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, U.; Durr, R.; Koch, J. Peptides as drugs: From screening to application. Curr. Pharm. Biotechnol. 2013, 14, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Barabasi, A.L.; Gulbahce, N.; Loscalzo, J. Network medicine: A network-based approach to human disease. Nat. Rev. Genet. 2011, 12, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Ray, N.B.; Durairaj, L.; Chen, B.B.; Mcverry, B.J.; Ryan, A.J.; Donahoe, M.; Waltenbaugh, A.K.; O’Donnell, C.P.; Henderson, F.C.; Etscheidt, C.A. Dynamic regulation of cardiolipin by the lipid pump Atp8b1 determines the severity of lung injury in experimental pneumonia. Nat. Med. 2010, 16, 1120–1127. [Google Scholar] [CrossRef]
- Dequin, P.F.; Meziani, F.; Quenot, J.P.; Kamel, T.; Ricard, J.D.; Badie, J. Hydrocortisone in Severe Community-Acquired Pneumonia. N. Engl. J. Med. 2023, 388, 1931–1941. [Google Scholar] [CrossRef]
- Clark, K.D.; Snashall, P.D. Endogenous Cortisol and Lung Damage in a Predominantly Smoking Population. Am. J. Respir. Crit. Care Med. 1999, 159, 755–759. [Google Scholar] [CrossRef]
- Aljada, A.; Assian, E.; Mohanty, P.; Hamouda, W.; Garg, R.; Dandona, P. Increased IkappaB expression and diminished nuclear NF-kappaB in human mononuclear cells following hydrocortisone injection. J. Clin. Endocrinol. Metab. 1999, 84, 3386–3389. [Google Scholar]
- Pols, T.W. TGR5 in inflammation and cardiovascular disease. Biochem. Soc. Trans. 2014, 42, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, T.; Yu, D.; Forman, B.M.; Huang, W. FXR protects lung from lipopolysaccharide-induced acute injury. Mol. Endocrinol. 2012, 26, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Cai, H.R.; Xue, S.; You, W.J.; Liu, B.; Jiang, H.D. Bile acids induce activation of alveolar epithelial cells and lung fibroblasts through farnesoid X receptor-dependent and independent pathways. Respirology 2016, 21, 1075–1080. [Google Scholar] [CrossRef]
- Xu, L.; Li, Y.; Wan, S.; Wang, Y.; Yu, P. Protective effects of apocynin nitrone on acute lung injury induced by lipopolysaccharide in rats. Int. Immunopharmacol. 2014, 20, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Gunther, A.; Siebert, C.; Schmidt, R.; Ziegler, S.; Grimminger, F.; Yabut, M.; Temmesfeld, B.; Walmrath, D.; Morr, H.; Seeger, W. Surfactant alterations in severe pneumonia, acute respiratory distress syndrome, and cardiogenic lung edema. Am. J. Respir. Crit. Care Med. 1996, 153, 176–184. [Google Scholar] [CrossRef]
- Spengler, D.; Winoto–Morbach, S.; Kupsch, S.; Vock, C.; Blochle, K.; Frank, S.; Rintz, N.; Diekotter, M.; Janga, H.; Weckmann, M. Novel therapeutic roles for surfactant–inositols and –phosphatidylglycerols in a neonatal piglet ARDS model: A translational study. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L32–L53. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Total Metabolites | Number Upregulated (%) | Number Downregulated (%) |
|---|---|---|---|
| LPS vs. MIX | 617 | 350 (56.72) vs. 389 (63.04) | 267 (43.27) vs. 228 (36.95) |
| LPS vs. NS | 617 | 350 (56.72) vs. 263 (42.62) | 267 (43.27) vs. 354 (57.37) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, T.; Zhou, J.; Guo, Y.; Hou, D.; Pi, N.; Yang, Y.; Yu, H. Mechanism of Alleviating Acute Lung Injury in Mice from Serum Metabolomics Analysis of Cordyceps fumosorosea. Separations 2024, 11, 74. https://doi.org/10.3390/separations11030074
Khan T, Zhou J, Guo Y, Hou D, Pi N, Yang Y, Yu H. Mechanism of Alleviating Acute Lung Injury in Mice from Serum Metabolomics Analysis of Cordyceps fumosorosea. Separations. 2024; 11(3):74. https://doi.org/10.3390/separations11030074
Chicago/Turabian StyleKhan, Tahir, Jinna Zhou, Yingqi Guo, Donghai Hou, Na Pi, Yinlong Yang, and Hong Yu. 2024. "Mechanism of Alleviating Acute Lung Injury in Mice from Serum Metabolomics Analysis of Cordyceps fumosorosea" Separations 11, no. 3: 74. https://doi.org/10.3390/separations11030074