

Optimized Extraction, Identification and Anti-Biofilm Action of Wu Wei Zi (Fructus Schisandrae Chinensis) Extracts against Vibrio parahaemolyticus

Abstract
:1. Introduction
2. Results
2.1. Effect of Single Factor on Antibacterial Activity of WWZE against V. parahaemolyticus
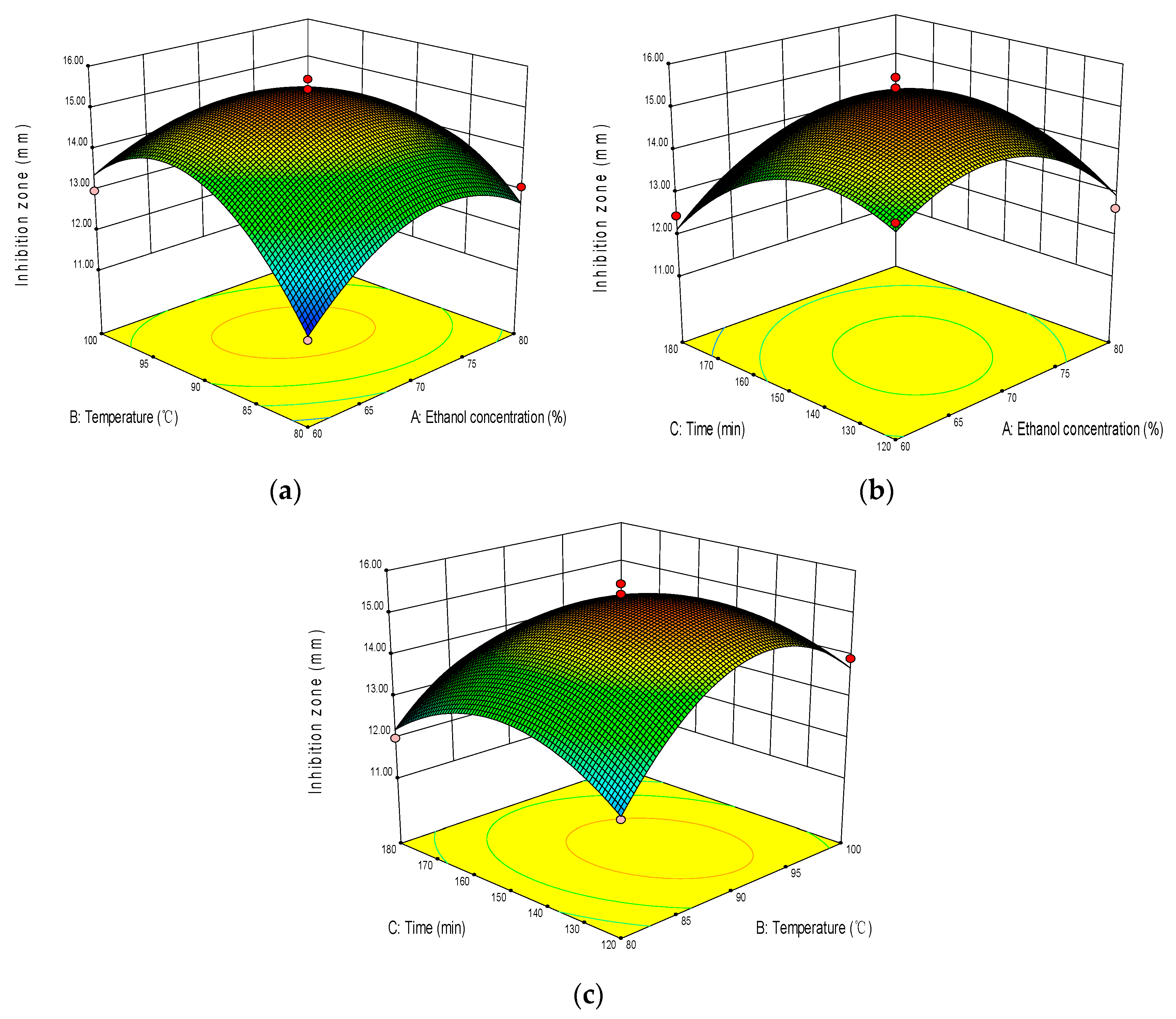
2.2. Optimization of Extraction Process
2.3. Identification of Anti-V. Parahaemolyticus Active Ingredients in WWZE
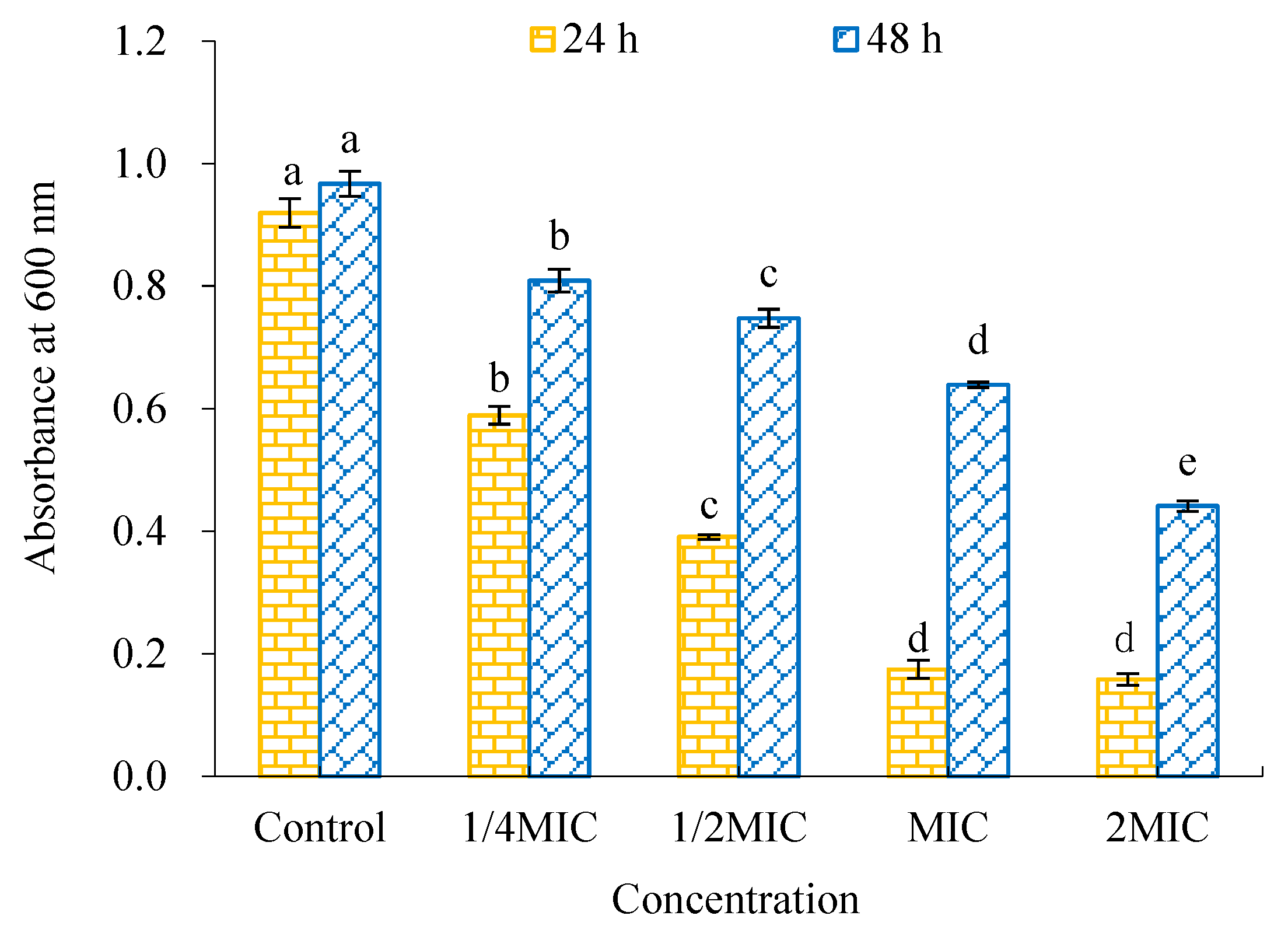
2.4. Antibiofilm Activity of WWZE against V. parahaemolyticus
2.5. Antibiofilm Mechanism of Action of WWZE against V. parahaemolyticus
2.5.1. Effect of WWZE on the Membrane Integrity of V. parahaemolyticus
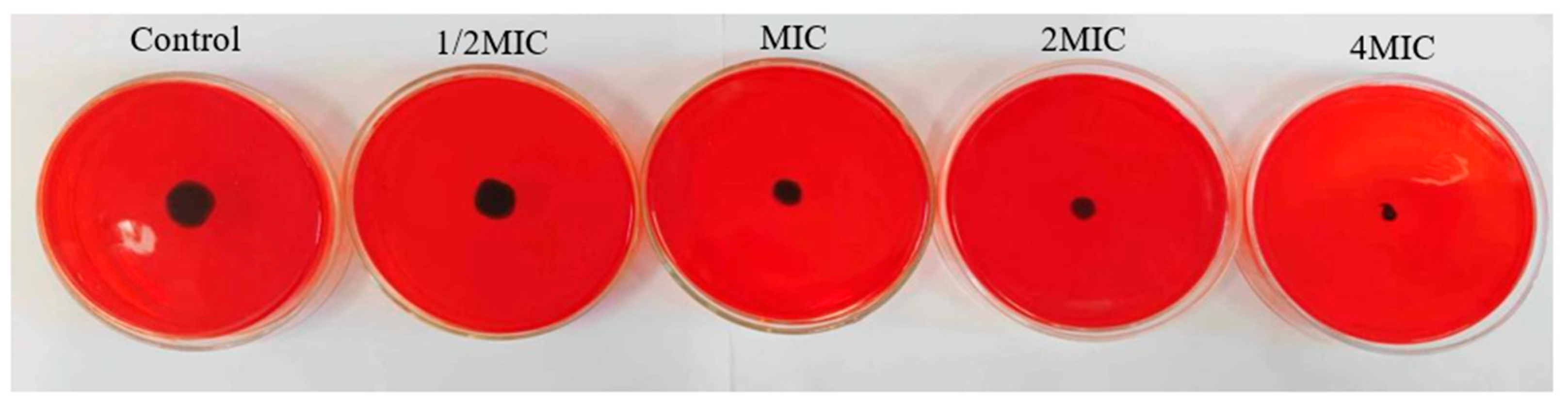
2.5.2. Effect of WWZE on Polysaccharide Intercellular Adhesin (PIA) Synthesis during Biofilm Formation of V. parahaemolyticus
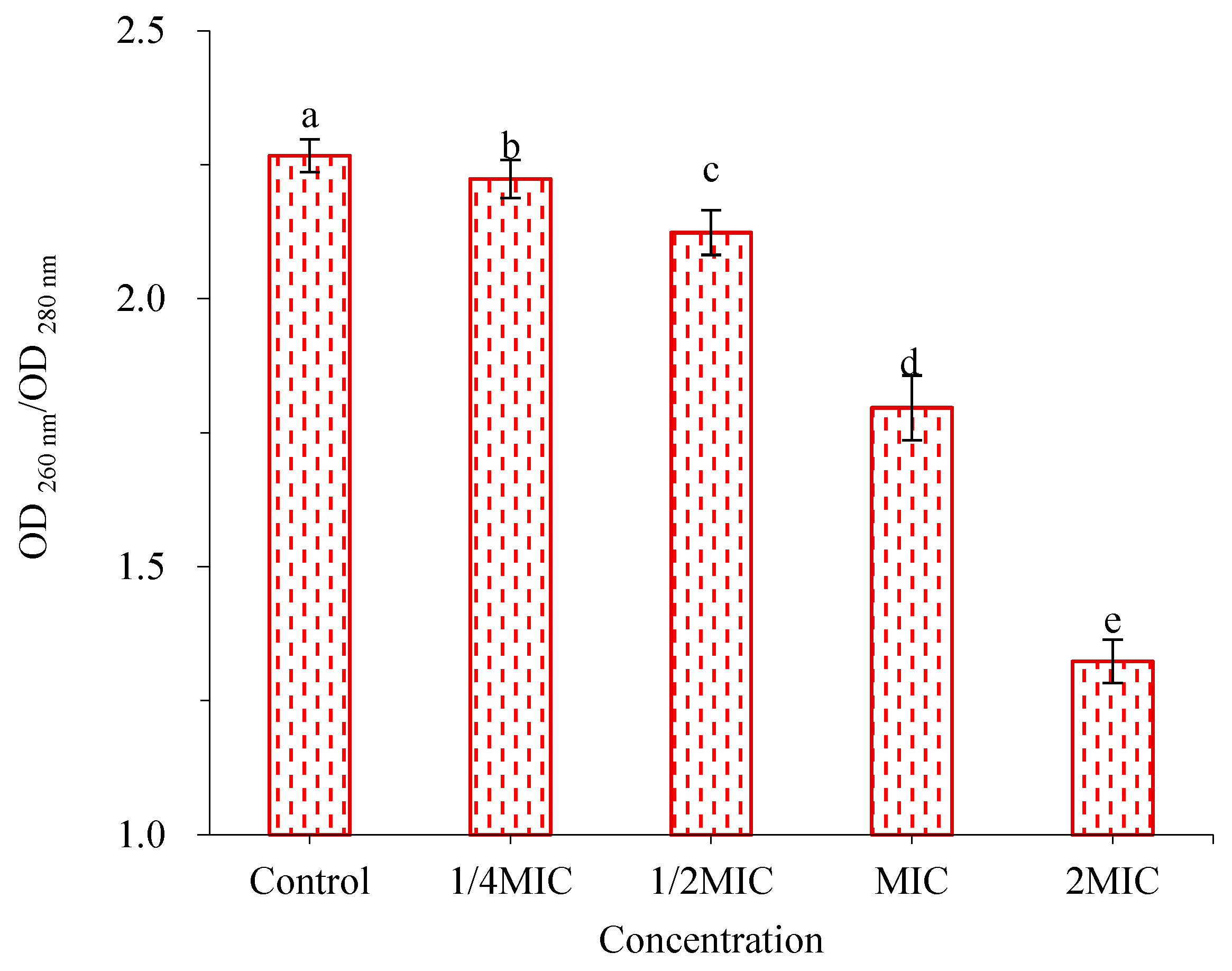
2.5.3. Effects of WWZE on Extracellular DNA (eDNA) Secretion during Biofilm Formation of V. parahaemolyticus
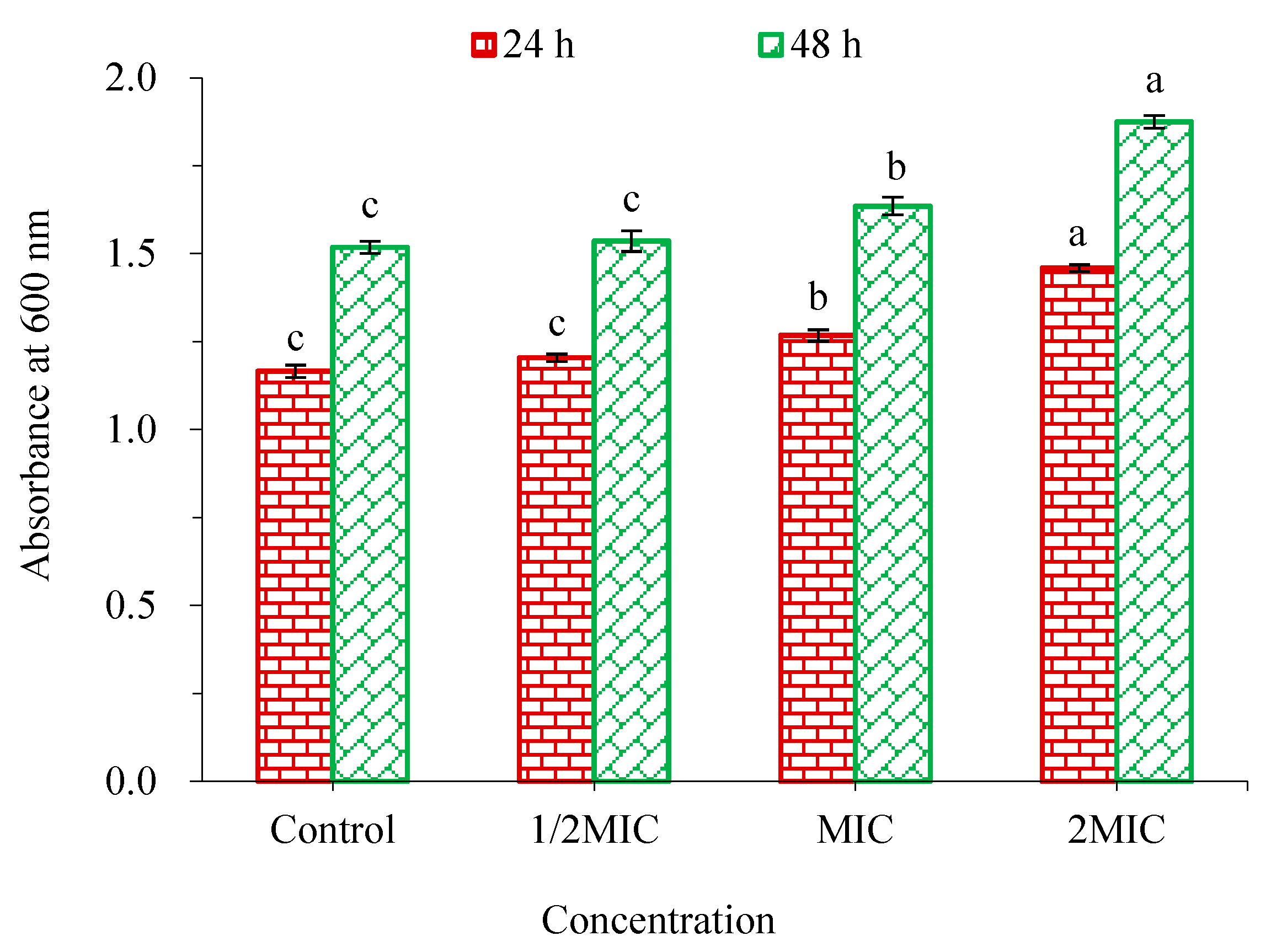
2.5.4. Effect of WWZE on Biofilm Metabolic Activity of V. parahaemolyticus
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Single-Factor Experiment
4.3. Box–Behnken Design
4.4. Agar Diffusion Method
4.5. High Performance Liquid Chromatography
4.6. Broth Microdilution Method
4.7. Crystal Violet Method
4.8. Coomassie Brilliant Blue Method
4.9. Congo Red Plate Method
4.10. Spectrophotometry
4.11. CCK-8 Method
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Robert-Pillot, A.; Copin, S.; Himber, C.; Gay, M.; Quilici, M.L. Occurrence of the three major Vibrio species pathogenic for human in seafood products consumed in France using real-time PCR. Int. J. Food Microbiol. 2014, 189, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Austin, B. Vibrios as causal agents of zoonoses. Vet. Microbiol. 2010, 140, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Zhang, Z.; Guo, L. Antibacterial molecules from marine microorganisms against aquatic pathogens: A concise review. Mar. Drugs 2022, 20, 230. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Ni, Q.; Chen, Z.; Li, J.; Zhao, F. Effects of short period feeding polysaccharides from marine macroalga, Ulva prolifera on growth and resistance of Litopenaeus vannamei against Vibrio parahaemolyticus infection. J. Appl. Phycol. 2019, 31, 2085–2092. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, Z.; Xu, X.; Cai, J.; Wang, W.; Guo, L. Antagonistic activity and mode of action of trypacidin from marine-derived Aspergillus fumigatus against Vibrio parahaemolyticus. 3 Biotech 2022, 12, 131. [Google Scholar] [CrossRef]
- Letchumanan, V.; Chan, K.G.; Lee, L.H. Vibrio parahaemolyticus: A review on the pathogenesis, prevalence, and advance molecular identification techniques. Front. Microbiol. 2014, 5, 705. [Google Scholar] [CrossRef] [Green Version]
- Ashrafudoulla, M.; Mizan, M.F.R.; Ha, A.J.; Park, S.H.; Ha, S.D. Antibacterial and antibiofilm mechanism of eugenol against antibiotic resistance Vibrio parahaemolyticus. Food Microbiol. 2020, 91, 103500. [Google Scholar] [CrossRef]
- Guo, L.; Wang, X.; Feng, J.; Xu, X.; Li, X.; Wang, W. Extraction, identification and mechanism of action of antibacterial substances from Galla chinensis against Vibrio harveyi. Biotechnol. Biotechnol. Equip. 2020, 34, 1215–1223. [Google Scholar] [CrossRef]
- Hu, J.; Wang, C.; Zu, H.; Fang, Z.; Zhou, H.; Zhou, J.; Wang, S.; Lyu, M. Nano-SiO2 inhibits the marine aquatic pathogen Vibrio parahaemolyticus. Aquacult. Rep. 2022, 23, 101015. [Google Scholar] [CrossRef]
- Abebe, G.M. The role of bacterial biofilm in antibiotic resistance and food contamination. Int. J. Microbiol. 2020, 2020, 1705814. [Google Scholar] [CrossRef]
- Flores-Vargas, G.; Bergsveinson, J.; Lawrence, J.R.; Korber, D.R. Environmental biofilms as reservoirs for antimicrobial resistance. Front. Microbiol. 2021, 12, 766242. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Song, X.; Zhang, Z.; Fu, J.; Wang, X.; Malakar, P.K.; Liu, H.; Pan, Y.; Zhao, Y. Removal of foodborne pathogen biofilms by acidic electrolyzed water. Front. Microbiol. 2017, 8, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural products as inspiration for the development of bacterial antibiofilm agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef]
- Pecoraro, C.; Carbone, D.; Deng, D.; Cascioferro, S.M.; Diana, P.; Giovannetti, E. Biofilm formation as valuable target to fight against severe chronic infections. Curr. Med. Chem. 2022, 29, 4307–4310. [Google Scholar] [CrossRef] [PubMed]
- Parrino, B.; Carbone, D.; Cascioferro, S.; Pecoraro, C.; Giovannetti, E.; Deng, D.; Di Sarno, V.; Musella, S.; Auriemma, G.; Cusimano, M.G.; et al. 1,2,4-Oxadiazole topsentin analogs as staphylococcal biofilm inhibitors targeting the bacterial transpeptidase sortase A. Eur. J. Med. Chem. 2021, 209, 112892. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Guo, J.; Xu, F. Optimized extraction process and identification of antibacterial substances from Rhubarb against aquatic pathogenic Vibrio harveyi. 3 Biotech 2017, 7, 377. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Cai, J.; Zhang, M.; Wen, Y.; Guo, L. The exploration of anti-Vibrio parahaemolyticus substances from Phellodendri Chinensis Cortex as a preservative for shrimp storage. Front. Microbiol. 2022, 13, 1004262. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Qiu, J.; Huang, Z.; Yu, Z.; Wang, W.; Hu, H.; You, Y. A comprehensive review of ethnopharmacology, phytochemistry, pharmacology, and pharmacokinetics of Schisandra chinensis (Turcz.) Baill. and Schisandra sphenanthera Rehd. et Wils. J. Ethnopharmacol. 2022, 284, 114759. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Men, L.; Sun, Y.; Wei, M.; Fan, X. Pharmacodynamic effects and molecular mechanisms of lignans from Schisandra chinensis Turcz. (Baill.), a current review. Eur. J. Pharmacol. 2021, 892, 173796. [Google Scholar] [CrossRef]
- Nowak, A.; Zakłos-Szyda, M.; Błasiak, J.; Nowak, A.; Zhang, Z.; Zhang, B. Potential of Schisandra chinensis (Turcz.) Baill. in human health and nutrition: A review of current knowledge and therapeutic perspectives. Nutrients 2019, 11, 333. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Ekiert, R.; Ekiert, H. Current knowledge of Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine) as a medicinal plant species: A review on the bioactive components, pharmacological properties, analytical and biotechnological studies. Phytochem. Rev. 2017, 16, 195–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Cui, S.; Li, T.; Yang, S.; Wang, Q.; He, K.; Zheng, Y.; He, C. Antibacterial effects of Schisandra chinensis extract on Staphylococcus aureus and its application in food. J. Food Saf. 2018, 38, e12503. [Google Scholar] [CrossRef]
- Cui, S.M.; Li, T.; Wang, Q.; He, K.K.; Zheng, Y.M.; Liang, H.Y.; Song, L.Y. Antibacterial effects of Schisandra chinensis extract on Escherichia coli and its applications in cosmetic. Curr. Microbiol. 2020, 77, 865–874. [Google Scholar] [CrossRef]
- Bai, X.; Park, I.B.; Hwang, H.J.; Mah, J.H. The ability of Schisandra chinensis fruit to inhibit the growth of foodborne pathogenic bacteria and the viability and heat resistance of Bacillus cereus spores. Int. J. Food Sci. Technol. 2015, 50, 2193–2200. [Google Scholar] [CrossRef]
- Zheng, H.; Zhao, Y.; Guo, L. A bioactive substance derived from brown seaweeds: Phlorotannins. Mar. Drugs 2022, 20, 742. [Google Scholar] [CrossRef]
- Xie, T.; Liao, Z.; Lei, H.; Fang, X.; Wang, J.; Zhong, Q. Antibacterial activity of food-grade chitosan against Vibrio parahaemolyticus biofilms. Microb. Pathog. 2017, 110, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Hakala, E.; Hanski, L.; Uvell, H.; Yrjönen, T.; Vuorela, H.; Elofsson, M.; Vuorela, P.M. Dibenzocyclooctadiene lignans from Schisandra spp. selectively inhibit the growth of the intracellular bacteria Chlamydia pneumoniae and Chlamydia trachomatis. J. Antibiot. 2015, 68, 609–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, H.; Cao, F.; Ming, D.; Zheng, Y.; Dong, X.; Zhong, X.; Mu, D.; Li, B.; Zhong, L.; Cao, J.; et al. Aloe-emodin inhibits Staphylococcus aureus biofilms and extracellular protein production at the initial adhesion stage of biofilm development. Appl. Microbiol. Biotechnol. 2017, 101, 6671–6681. [Google Scholar] [CrossRef]
- Yan, X.; Gu, S.; Shi, Y.; Cui, X.; Wen, S.; Ge, J. The effect of emodin on Staphylococcus aureus strains in planktonic form and biofilm formation In Vitro. Arch. Microbiol. 2017, 199, 1267–1275. [Google Scholar] [CrossRef]
- Lu, C.; Liu, H.; Shangguan, W.; Chen, S.; Zhong, Q. Antibiofilm activities of the cinnamon extract against Vibrio parahaemolyticus and Escherichia coli. Arch. Microbiol. 2021, 203, 125–135. [Google Scholar] [CrossRef]
- Li, W.R.; Shi, Q.S.; Liang, Q.; Xie, X.B.; Huang, X.M.; Chen, Y.B. Antibacterial activity and kinetics of Litsea cubeba oil on Escherichia coli. PLoS ONE 2014, 9, e110983. [Google Scholar] [CrossRef]
- Speziale, P.; Pietrocola, G.; Foster, T.J.; Geoghegan, J.A. Protein-based biofilm matrices in Staphylococci. Front. Cell. Infect. Microbiol. 2014, 4, 171. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Campoccia, D.; Ravaioli, S.; Montanaro, L. Polysaccharide intercellular adhesin in biofilm: Structural and regulatory aspects. Front. Cell. Infect. Microbiol. 2015, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.; Hu, X.; Zhang, X.; Wang, W.; Sun, J.; Su, Z.; Zhu, C. Strategies to prevent, curb and eliminate biofilm formation based on the characteristics of various periods in one biofilm life cycle. Front. Cell. Infect. Microbiol. 2022, 12, 1003033. [Google Scholar] [CrossRef] [PubMed]
- Naradasu, D.; Guionet, A.; Miran, W.; Okamoto, A. Microbial current production from Streptococcus mutans correlates with biofilm metabolic activity. Biosens. Bioelectron. 2020, 162, 112236. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Wang, L.; Li, X.; Xu, X.; Guo, J.; Wang, X.; Yang, W.; Xu, F.; Li, F. Enhanced production of questin by marine-derived Aspergillus flavipes HN4-13. 3 Biotech 2020, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Guo, S.; Chen, H.; Zhang, Z.; Li, X.; Wang, W.; Guo, L. Bioassay-guided isolation and characterization of antibacterial compound from Aspergillus fumigatus HX-1 associated with Clam. 3 Biotech 2021, 11, 193. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, H.; Lu, M.; Fang, Y.; Jiao, Y.; Li, W.; Zhao, G.; Wang, S. Dextranase from Arthrobacter oxydans KQ11-1 inhibits biofilm formation by polysaccharide hydrolysis. Biofouling 2016, 32, 1223–1233. [Google Scholar] [CrossRef]
- An, J.; Ding, N.; Zhang, Z. Mechanical and antibacterial properties of polymethyl methacrylate modified with zinc dimethacrylate. J. Prosthet. Dent. 2022, 128, 100.e1–100.e8. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | X1 | X2 | X3 | Y |
|---|---|---|---|---|
| Ethanol Concentration (%) | Temperature (°C) | Time (min) | Diameter of Inhibitory Circle (mm) | |
| 1 | −1 (60) | −1 (80) | 0 (150) | 11.36 |
| 2 | +1 (80) | −1 (80) | 0 (150) | 13.09 |
| 3 | −1 (60) | +1 (100) | 0 (150) | 12.99 |
| 4 | +1 (80) | +1 (100) | 0 (150) | 12.14 |
| 5 | −1 (60) | 0 (90) | −1 (120) | 14.06 |
| 6 | +1 (80) | 0 (90) | −1 (120) | 12.64 |
| 7 | −1 (60) | 0 (90) | +1 (180) | 12.46 |
| 8 | +1 (80) | 0 (90) | +1 (180) | 12.78 |
| 9 | 0 (70) | −1 (80) | −1 (120) | 12.03 |
| 10 | 0 (70) | +1 (100) | −1 (120) | 13.93 |
| 11 | 0 (70) | −1 (80) | +1 (180) | 11.99 |
| 12 | 0 (70) | +1 (100) | +1 (180) | 11.95 |
| 13 | 0 (70) | 0 (90) | 0 (150) | 15.69 |
| 14 | 0 (70) | 0 (90) | 0 (150) | 15.21 |
| 15 | 0 (70) | 0 (90) | 0 (150) | 15.45 |
| 16 | 0 (70) | 0 (90) | 0 (150) | 15.09 |
| Source | Sum of Squares | df | Mean Square | F Value | Prob > F | Sig. |
|---|---|---|---|---|---|---|
| Model | 32.57 | 9 | 3.62 | 27.32 | 0.0001 | ** |
| X1 | 0.00605 | 1 | 0.00605 | 0.05 | 0.8369 | |
| X2 | 0.81 | 1 | 0.81 | 6.09 | 0.0430 | * |
| X3 | 1.51 | 1 | 1.51 | 11.43 | 0.0117 | * |
| X1X2 | 1.66 | 1 | 1.66 | 12.56 | 0.0094 | ** |
| X1X3 | 0.76 | 1 | 0.76 | 5.71 | 0.0481 | * |
| X2X3 | 0.94 | 1 | 0.94 | 7.10 | 0.0322 | * |
| X12 | 6.22 | 1 | 6.22 | 46.96 | 0.0002 | ** |
| X22 | 12.54 | 1 | 12.54 | 94.64 | <0.0001 | ** |
| X32 | 5.43 | 1 | 5.43 | 40.99 | 0.0004 | ** |
| Lack of Fit | 0.70 | 3 | 0.23 | 4.19 | 0.1001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhao, Y.; Cai, J.; Wang, T.; Song, Y.; Lu, J.; Du, H.; Wang, W.; Zhao, Y.; Guo, L. Optimized Extraction, Identification and Anti-Biofilm Action of Wu Wei Zi (Fructus Schisandrae Chinensis) Extracts against Vibrio parahaemolyticus. Molecules 2023, 28, 2268. https://doi.org/10.3390/molecules28052268
Zhang Z, Zhao Y, Cai J, Wang T, Song Y, Lu J, Du H, Wang W, Zhao Y, Guo L. Optimized Extraction, Identification and Anti-Biofilm Action of Wu Wei Zi (Fructus Schisandrae Chinensis) Extracts against Vibrio parahaemolyticus. Molecules. 2023; 28(5):2268. https://doi.org/10.3390/molecules28052268
Chicago/Turabian StyleZhang, Zongyi, Yanan Zhao, Jing Cai, Tong Wang, Yujie Song, Jingyi Lu, Hairuo Du, Wenfang Wang, Yan Zhao, and Lei Guo. 2023. "Optimized Extraction, Identification and Anti-Biofilm Action of Wu Wei Zi (Fructus Schisandrae Chinensis) Extracts against Vibrio parahaemolyticus" Molecules 28, no. 5: 2268. https://doi.org/10.3390/molecules28052268