
The Antifungal Potential of Niclosamide and Structurally Related Salicylanilides


Abstract
:1. Introduction
2. Methodology
3. Niclosamide: Chemistry, Pharmacology, and Antifungal Activity
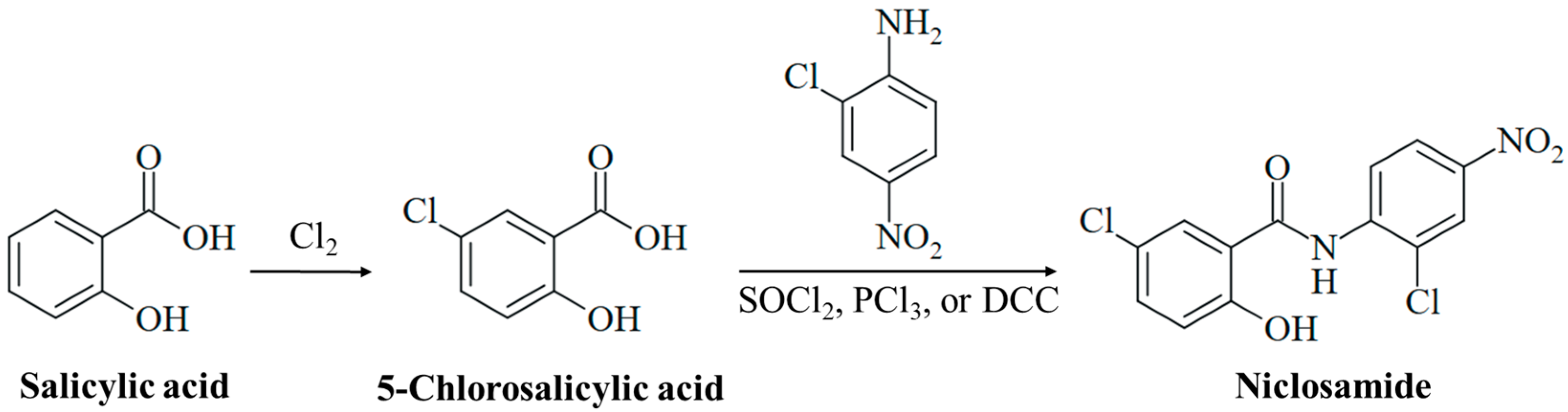
3.1. Chemistry
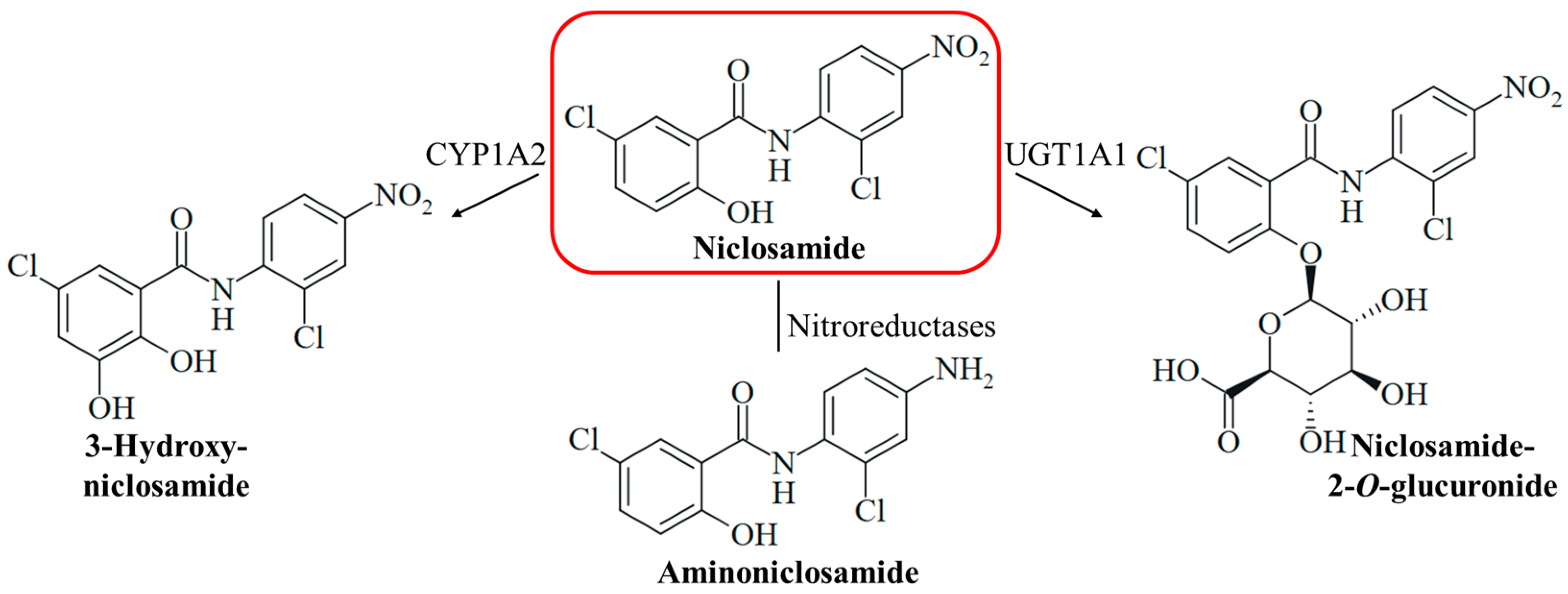
3.2. Pharmacology
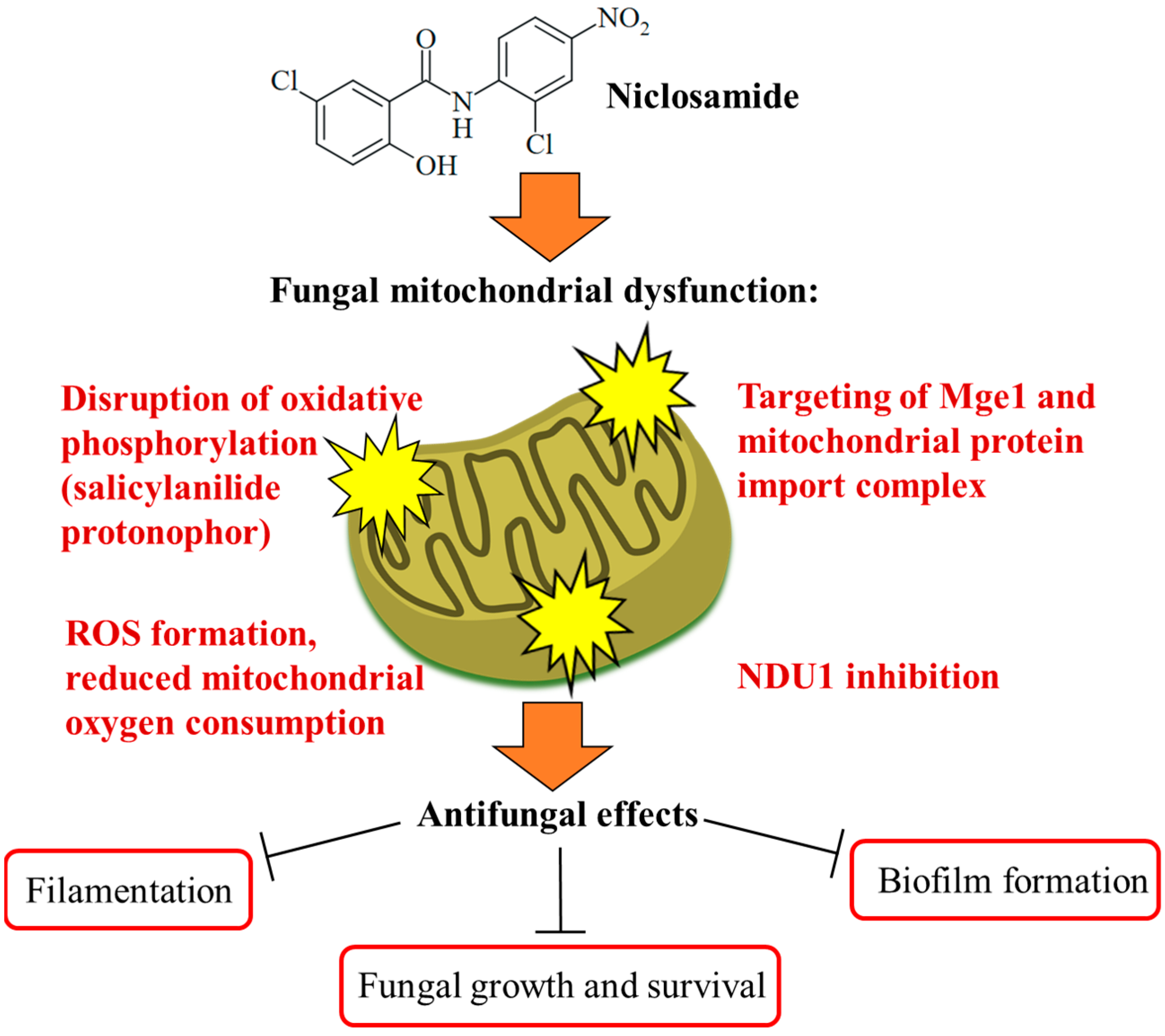
3.3. Antifungal Activity
3.3.1. Activity against Pathogenic Ascomycetes
3.3.2. Activity against Pathogenic Basidiomycetes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Fungus | Activity/Mechanism | Reference |
|---|---|---|---|
| Ascomycetes | Candida albicans (including azole-resistant strains) | Inhibition of filamentation, biofilm formation, and invasion of host cells (no growth inhibition); retrograde response; possible targets Mge1 and mitochondrial protein import complex | [51] |
| Candida albicans (including azole-resistant strains), Candida auris | Blocking of biofilm formation via NDU1 inhibition, increased ROS formation, suppressed mitochondrial oxygen consumption, and protection of mice from infection | [52] | |
| Trichophyton tonsurans | Strong growth inhibition (MIC < 1 µM) | [57] | |
| Sporothrix brasiliensis | Broad-spectrum growth inhibition (MIC = 0.625–2.5 µM, wildtype and non-wildtype strains); fungicidal | [60,61] | |
| Paracoccidioides brasiliensis | Fungicidal (1 µM) | [60] | |
| Histoplasma capsulatum | Fungicidal (1 µM) | [60] | |
| Madurella mycetomatis | Growth inhibition of SO1 and CBS131320 isolates (MIC = 0.79–1.6 µg/mL) | [66] | |
| Basidiomycetes | Cryptococcus neoformans | Growth inhibition of H99 wildtype strain (IC50 = 0.17 µM) and nine clinical isolates (IC50 = 0.17–0.52 µM) under nutrient-starvation conditions | [71] |
| Cryptococcus neoformans | Growth inhibitory and fungicidal effects on the JEC21 isolate (MIC < 0.78 µg/mL, MFC = 1.56 µg/mL); inhibition of spore germination | [72] |
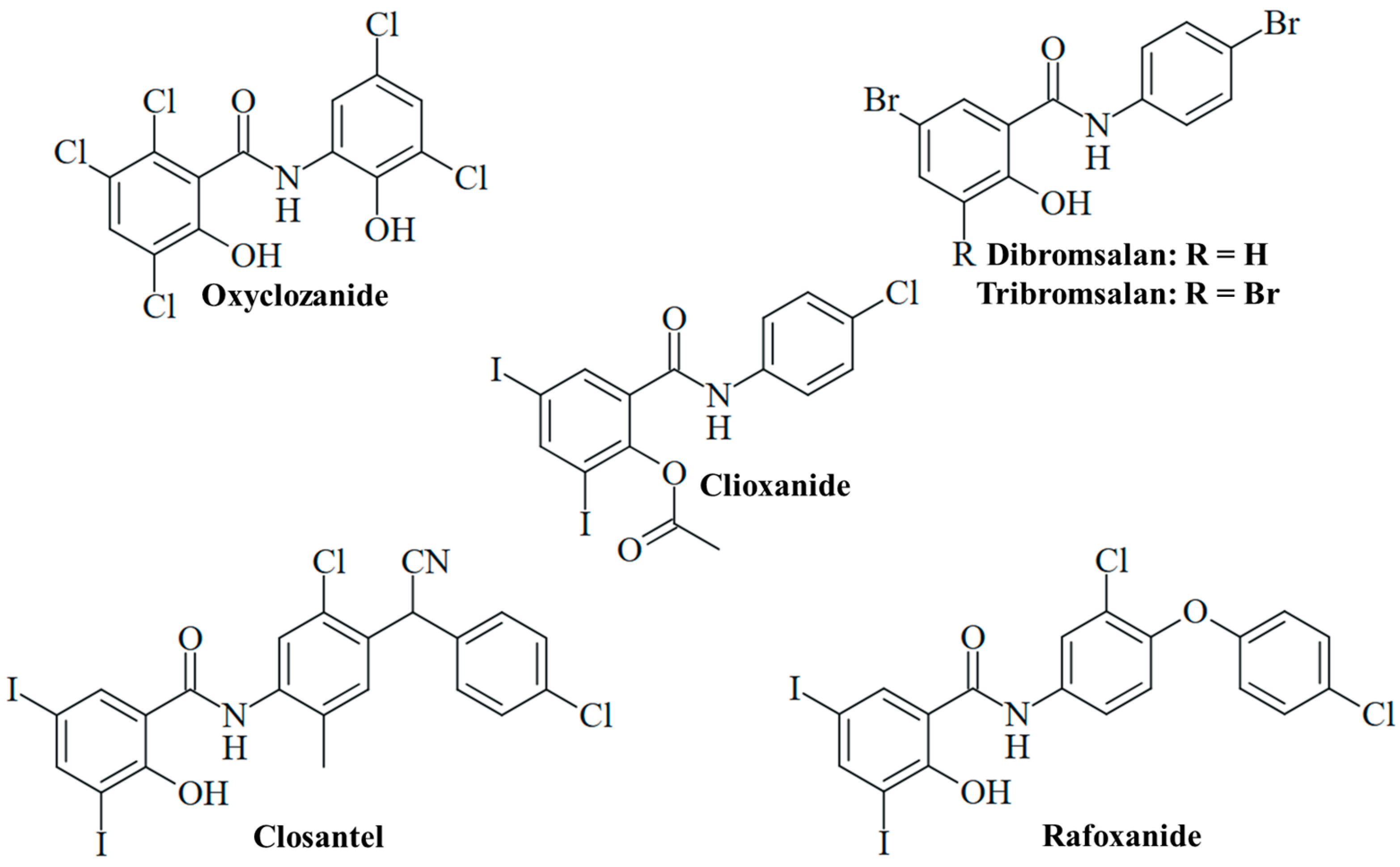
4. Antifungal Activity of Other Salicylanilides
4.1. Oxyclozanide
4.2. Further Salicylanilide Drugs with Antifungal Activity
4.2.1. 3,5-Diiodosalicylanilides
| Compound | Fungus | Activity/Mechanism | Reference |
|---|---|---|---|
| Oxyclozanide | Candida albicans | Inhibition of growth and filamentation | [51] |
| Candida albicans | Active in glucose-free medium with non-fermentable carbon sources (ethanol, glycerol); disruption of mitochondrial membrane potential | [77] | |
| Madurella mycetomatis | Low activity | [76] | |
| Closantel | Trichophyton tonsurans | Active (MIC < 1 µM) | [57] |
| Rafoxanide | Aspergillus fumigatus (resistant strains with upregulated Cyp51A) | Active (MIC = 2–8 µg/mL); suppression of Cyp51A; sensitization to fluconazole; in vivo activity (sub-MIC doses) in albino mice in combination with fluconazole (survival rate 60%) | [88] |
| Candida albicans (resistant strains with upregulated Erg11) | Active (MIC = 2–8 µg/mL); suppression of Erg11; sensitization to fluconazole; in vivo activity (sub-MIC doses) in albino mice in combination with fluconazole (survival rate 70%) | [88] | |
| Bromsalans | Aspergillus niger, Candida albicans, Pityrosporum ovale | Growth inhibition; dibromsalan more active than tribromsalan against C. albicans and P. ovale; Miranol CS formulation enhanced antifungal activity | [16] |
| Tribromsalan | C. albicans | Growth inhibition (20 µM); low inhibition of C. albicans filamentation | [51] |
4.2.2. Bromsalans
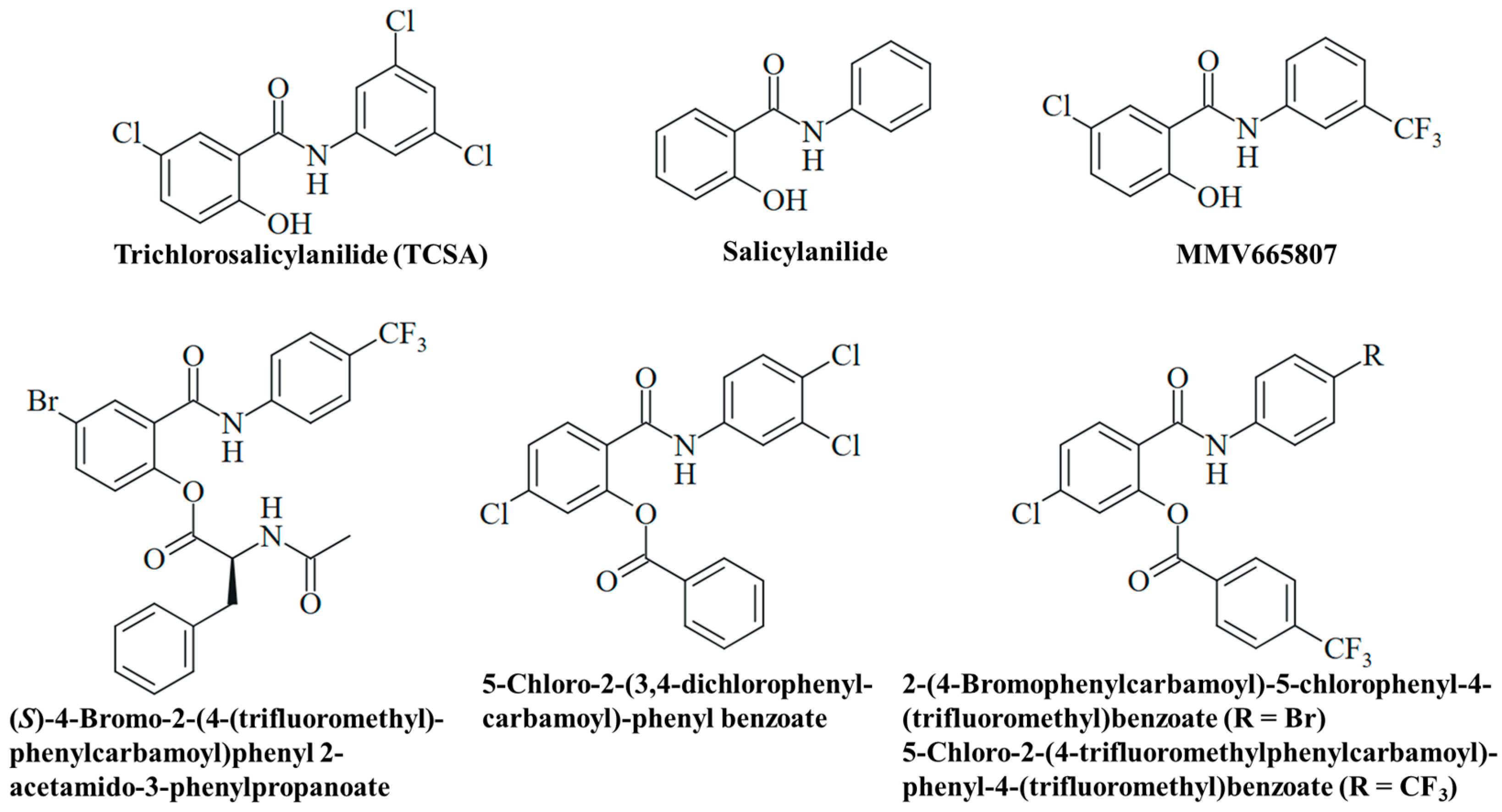
4.3. Experimental Salicylanilide Compounds
| Compound | Fungus | Activity/Mechanism | Reference |
|---|---|---|---|
| Salicylanilide | Candida albicans | Inhibition of growth and filamentation | [51] |
| TCSA | Candida albicans (including azole-resistant strains) | Inhibition of filamentation; no growth inhibition | [51] |
| MMV665907 | Madurella mycetomatis | Active (MIC = 1.6 µg/mL, SO1 and CBS131320) | [66] |
| (S)-4-Bromo-2-(4-(trifluoromethyl)phenylcarbamoyl)phenyl 2-acetamido-3-phenylpropanoate | Trichophyton mentagrophytes | Active (MIC = 3.9–7.8 µM) | [94] |
| 5-Chloro-2-(3,4-dichlorophenylcarbamoyl)phenyl benzoate | Trichosporon asahii | Active (MIC = 3.9 µM); medium acidification (pH 5); reduced activity | [95] |
| Candida krusei Trichophyton mentagrophytes | Moderate activity (MIC = 15.6 µM, C. krusei; MIC = 31.3 µM, T. mentagrophytes) | [95] | |
| 2-(4-Bromophenylcarbamoyl)-5-chlorophenyl-4-(trifluoromethyl)benzoate 5-Chloro-2-(4-(trifluoromethyl)phenylcarbamoyl)phenyl-4-(trifluoromethyl)benzoate | Trichophyton mentagrophytes | Active (MIC = 0.49 µM) | [96] |
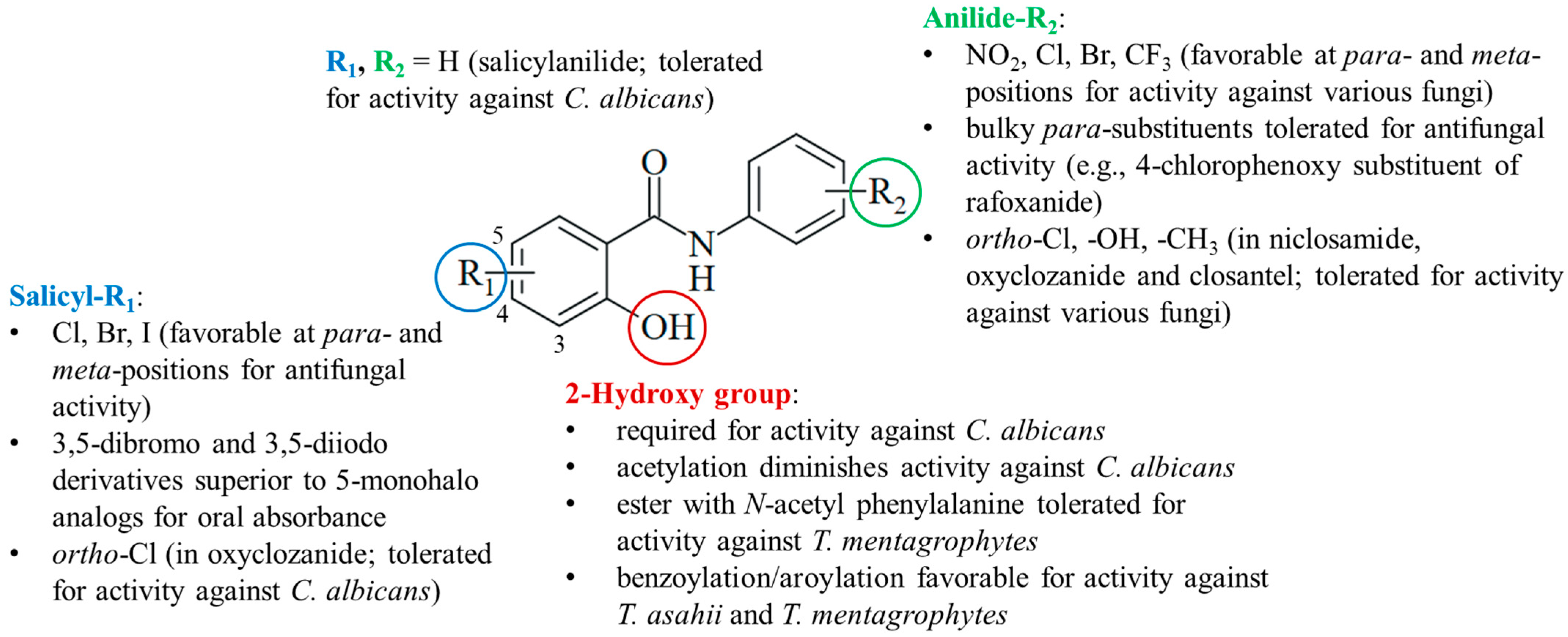
4.4. Structure–Activity Relationships
5. Discussion of Current Challenges and Opportunities
5.1. Antifungal Activity and Mechanisms
5.2. Salicylanilide Resistance and Possible Combination Partners
5.3. Adverse Effects, Drug–Drug Interactions, and Drug Availability
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal infections in humans: The silent crisis. Microb. Cell 2020, 7, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.Z.P.; Schwartz, I.S. Emerging fungal infections: New patients, new patterns, new pathogens. J. Fungi 2019, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Queiroz-Telles, F.; Fahal, A.H.; Falci, D.R.; Caceres, D.H.; Chiller, T.; Pasqualatto, A.C. Neglected endemic mycoses. Lancet Infect. Dis. 2017, 17, e367–e377. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Patterson, T.F. Multidrug-resistant Candida: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal diseases as neglected pathogens: A wake-up call to public health officials. PLoS Negl. Trop. Dis. 2020, 14, e0007964. [Google Scholar] [CrossRef] [PubMed]
- Mycetoma. Available online: https://dndi.org/diseases/mycetoma/ (accessed on 28 March 2024).
- Araújo, G.R.S.; Souza, W.; Frases, S. The hidden pathogenic potential of environmental fungi. Future Microbiol. 2017, 12, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Téllez, M.D.; Batista-Duharte, A.; Portuondo, D.; Quinello, C.; Bonne-Hernández, R.; Carlos, I.Z. Sporothrix schenkii complex biology: Environment and fungal pathogenicity. Microbiology 2014, 160, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Carmo, A.; Rocha, M.; Pereirinha, P.; Tomé, R.; Costa, E. Antifungals: From pharmacokinetics to clinical practice. Antibiotics 2023, 12, 884. [Google Scholar] [CrossRef]
- Van Daele, R.; Spriet, I.; Wauters, J.; Maertens, J.; Mercier, T.; Van Hecke, S.; Brüggemann, R. Antifungal drugs: What brings the future? Med. Mycol. 2019, 57, S328–S343. [Google Scholar] [CrossRef]
- Gintjee, T.J.; Donnelley, M.A.; Thompson, G.R., III. Aspiring antifungals: Review of current antifungal pipeline developments. J. Fungi 2020, 6, 28. [Google Scholar] [CrossRef]
- Jampilek, J. Novel avenues for identification of new antifungal drugs and current challenges. Expert Opin. Drug Discov. 2022, 17, 949–968. [Google Scholar] [CrossRef]
- 125 Years of Acetylsalicylic Acid. Available online: https://www.bayer.com/en/news-stories/125-years-of-acetylsalicylic-acid (accessed on 25 March 2024).
- Andrews, P.; Thyssen, J.; Lorke, D. The biology and toxicology of molluscicides, Bayluscide. Pharmacol. Ther. 1983, 19, 245–295. [Google Scholar] [CrossRef] [PubMed]
- Jara, M.O.; Williams III, R.O. The challenge of repurposing niclosamide: Considering pharmacokinetic parameters, routes of administration, and drug metabolism. J. Drug Deliv. Sci. Technol. 2023, 81, 104187. [Google Scholar] [CrossRef]
- Molnar, N.M. Influence of various surfactants on the antimicrobial activity of bromsalans and other ring-halogenated substances. J. Am. Oil Chem. Soc. 1968, 45, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Tan, P.; Feng, X.; Chen, M.; Kajihara, N.; Minai, M.; Hosaka, Y. Assessment of the molluscicidal activities of tribromosalan, cartap and chlorothalonil against Oncomelania hupensis. Jpn. J. Med. Sci. Biol. 1992, 45, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Swan, G.E. The pharmacology of halogenated salicylanilides and their anthelmintic use in animals. J. S. Afr. Vet. Assoc. 1999, 70, 61–70. [Google Scholar] [CrossRef]
- Pearson, I.G.; Whitlock, H.V.; DeGoosh, C.P.; Farrington, K.J.; Jones, R.C.; Haigh, J.A. Clioxanide, a new anthelmintic active against Fasciola hepatica and Haeminchus contortus in sheep. Aust. Vet. J. 1970, 46, 480–484. [Google Scholar] [CrossRef]
- Kadri, H.; Lambourne, O.A.; Mehellou, Y. Niclosamide, a drug with many (re)purposes. ChemMedChem 2018, 13, 1088–1091. [Google Scholar] [CrossRef]
- Chen, W.; Mook, R.A., Jr.; Premont, R.T.; Wang, J. Niclosamide: Beyond an antihelminthic drug. Cell Signal. 2018, 41, 89–96. [Google Scholar] [CrossRef]
- Kapale, S.S.; Chaudhari, H.K. Niclosamide & challenges in chemical modifications: A broad review on enhancement of solubility. J. Indian Chem. Soc. 2021, 98, 100262. [Google Scholar]
- Wang, Z.; Ren, J.; Du, J.; Wang, H.; Liu, J.; Wang, G. Niclosamide as a promising therapeutic player in human cancer and other diseases. Int. J. Mol. Sci. 2022, 23, 16116. [Google Scholar] [CrossRef] [PubMed]
- Kauerová, T.; Pérez-Pérez, M.-J.; Kollar, P. Salicylanilides and their anticancer properties. Int. J. Mol. Sci. 2023, 24, 1728. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, J.G.; Mahdi, A.J.; Mahdi, A.J.; Bowen, I.D. The historical analysis of aspirin discovery, its relation to the willow tree and antiproliferative and anticancer potential. Cell Prolif. 2006, 39, 147–155. [Google Scholar] [CrossRef]
- Hirwe, N.W.; Rana, K.N.; Gavankar, K.D. Derivatives of salicylic acid. Part XIII. Chlorosalicylic acids and their methyl ethers. Proc. Indian Acad. Sci. A 1938, 8, 208–213. [Google Scholar] [CrossRef]
- Chae, H.-D.; Cox, N.; Capolicchio, S.; Lee, J.W.; Horikoshi, N.; Kam, S.; Ng, A.A.; Edwards, J.; Butler, T.-L.; Chan, J.; et al. SAR optimization studies on modified salicylamides as a potential treatment for acute myeloid leukemia through inhibition of the CREB pathway. Bioorg. Med. Chem. Lett. 2019, 29, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- Juang, Y.-P.; Chou, Y.-T.; Lin, R.-X.; Ma, H.-H.; Chao, T.-L.; Jan, J.-T.; Chang, S.-Y.; Liang, P.-H. Design, synthesis and biological evaluations of niclosamide analogues against SARS-CoV-2. Eur. J. Med. Chem. 2022, 235, 114295. [Google Scholar] [CrossRef] [PubMed]
- Lal, J.; Ramalingam, K.; Meena, R.; Ansari, S.B.; Saxena, D.; Chopra, S.; Goyal, N.; Reddy, D.N. Design and synthesis of novel halogen rich salicylanilides as potential antileishmanial agents. Eur. J. Med. Chem. 2023, 246, 114996. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-R.; Lin, C.-C.; Huang, C.-Y.; Wong, Y.-H.; Hsieh, C.-H.; Wu, H.-W.; Chen, J.J.W.; Wu, Y.-S. Study of the inhibitory effects on TNF-α-induced NF-κB activation of IMD0354 analogs. Chem. Biol. Drug Des. 2017, 90, 1307–1311. [Google Scholar] [CrossRef] [PubMed]
- Niclosamide Oral. Available online: https://medicalguidelines.msf.org/en/viewport/EssDr/english/niclosamide-oral-16684320.html (accessed on 18 May 2024).
- Espinosa-Aguirre, J.J.; Reyes, R.E.; de Nava, C.C. Mutagenic activity of 2-chloro-4-nitroaniline and 5-chlorosalicylic acid in Salmonella typhimurium: Two possible metabolites of niclosamide. Mutat. Res. 1991, 264, 139–145. [Google Scholar] [CrossRef]
- Beristain-Castillo, E.; Martínez-Vázquez, M.; Camacho-Carranza, R.; Espinosa-Aguirre, J.J. CYP1A1 and Cnr nitroreductase bio-activated niclosamide in vitro. Mutagenesis 2013, 28, 645–651. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Yeh, T.-K.; Lin, K.-T.; Chen, W.-C.; Yao, H.-T.; Lan, S.-J.; Wu, Y.-S.; Hsieh, H.-P.; Chen, C.-M.; Chen, C.-T. Pharmacokinetics of anti-SARS-CoV agent niclosamide and its analogs in rats. Yaowu Shipin Fenxi 2006, 14, 329–333. [Google Scholar] [CrossRef]
- Lu, D.; Ma, Z.; Zhang, T.; Zhang, X.; Wu, B. Metabolism of the anthelmintic drug niclosamide by cytochrome P450 enzymes and UDP-glucuronosyltransferases: Metabolite elucidation and main contributions from CYP1A2 and UGT1A1. Xenobiotica 2016, 46, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.I.; Jin, G.-W.; Yoo, H.H. Pharmacokinetic considerations for enhancing drug repurposing opportunities of anthelmintics: Niclosamide as a case study. Biomed. Pharmacother. 2024, 173, 116394. [Google Scholar] [CrossRef]
- Fan, X.; Li, H.; Ding, X.; Zhang, Q.-Y. Contributions of hepatic and intestinal metabolism to the disposition of niclosamide, a repurposed drug with poor bioavailability. Drug Metab. Dispos. 2019, 47, 756–763. [Google Scholar] [CrossRef]
- Mook Jr, R.A.; Wang, J.; Ren, X.-R.; Chen, M.; Spasojevic, I.; Barak, L.S.; Lyerly, H.K.; Chen, W. Structure-activity studies of Wnt/β-catenin inhibition in the niclosamide chemotype: Identification of derivatives with improved drug exposure. Bioorg. Med. Chem. 2015, 23, 5829–5838. [Google Scholar] [CrossRef]
- Ngai, T.W.; Elfar, G.A.; Yeo, P.; Phua, N.; Hor, J.H.; Chen, S.; Ho, Y.S.; Cheok, C.F. Nitro-deficient niclosamide confers reduced genotoxicity and retains mitochondrial uncoupling activity for cancer therapy. Int. J. Mol. Sci. 2021, 22, 10420. [Google Scholar] [CrossRef]
- Chang, X.; Zhen, X.; Liu, J.; Ren, X.; Hu, Z.; Zhou, Z.; Zhu, F.; Ding, K.; Nie, J. The anthelmintic phosphate niclosamide impedes renal fibrosis by inhibiting homeodomain-interacting protein kinase 2 expression. Kidney Int. 2017, 92, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, E.J.; Löbenberg, R.; de Araujo, G.L.B.; Bou-Chacra, N.A. Niclosamide repositioning for treating cancer: Challenges and nano-based drug delivery opportunities. Eur. J. Pharm. Biopharm. 2019, 141, 58–69. [Google Scholar] [CrossRef]
- Choi, G.; Piao, H.; Rejinold, N.S.; Yu, S.; Kim, K.-Y.; Jin, G.-W.; Choy, J.-H. Hydrotalcite–niclosamide nanohybrid as oral formulation towards SARS-CoV-2 viral infections. Pharmaceuticals 2021, 14, 486. [Google Scholar] [CrossRef]
- Jara, M.O.; Warnken, Z.N.; Sahakijpijarn, S.; Moon, C.; Maier, E.Y.; Christensen, D.J.; Koleng, J.J.; Peters, J.I.; Hackman Maier, S.D.; Williams, R.O., III. Niclosamide inhalation powder made by thin-film freezing: Multi-dose tolerability and exposure in rats and pharmacokinetics in hamsters. Int. J. Pharm. 2021, 603, 120701. [Google Scholar] [CrossRef]
- Bruce, J.I.; Miller, R.; Lightner, L.; Yoganathan, S. Efficacy of niclosamide as a potential topical antipenetrant (TAP) against cercariae of Schistosoma mansoni in monkeys. Mem. Inst. Oswaldo Cruz 1992, 87, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Wulff, C.; Haeberlein, S.; Haas, W. Cream formulations protecting against cercarial dermatitis by Trichobilhazia. Parasitol. Res. 2007, 101, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Delavenne, E.; Matias, C.; Lagler, H.; Simon, D.; Li, P.; Hansen, J.U.; dos Santos, T.P.; Jana, B.; Priemel, P.; et al. Topical niclosamide (ATx201) reduces Staphylococcus aureus colonization in atopic dermatitis patients in a randomized, double-blind, placebo-controlled phase 2 trial. Clin. Transl. Med. 2022, 12, e790. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.-C.; Kim, G.Y.; Kim, D.E.; Choi, J.W.; Chun, B.C. Grafting of niclosamide and salicylanilide onto hydrophilic polyurethane for the control of fungal and barnacle growth. Polym. Bull. 2019, 76, 2041–2060. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Brandt, M.E.; Lockhart, S.R. Recent taxonomic developments with Candida and other opportunistic yeasts. Curr. Fungal Infect. Rep. 2012, 6, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Spettel, K.; Kriz, R.; Wu, C.; Achter, L.; Schmid, S.; Galazka, S.; Selitsch, B.; Camp, I.; Makristathis, A.; Lagler, H.; et al. Candida auris in Austria—What is new and what is different. J. Fungi 2023, 9, 129. [Google Scholar] [CrossRef]
- Garcia, C.; Burgain, A.; Chaillot, J.; Pic, É.; Khemiri, I.; Sellam, A. A phenotypic small-molecule screen identifies halogenated salicylanilides as inhibitors of fungal morphogenesis, biofilm formation and host cell invasion. Sci. Rep. 2018, 8, 11559. [Google Scholar] [CrossRef] [PubMed]
- Sutar, Y.; Nabeela, S.; Singh, S.; Alqarihi, A.; Solis, N.; Ghebremariam, T.; Filler, S.; Ibrahim, A.S.; Date, A.; Uppuluri, P. Niclosamide-loaded nanoparticles disrupt Candida biofilms and protect mice from mucosal candidiasis. PLoS Biol. 2022, 20, e3001762. [Google Scholar] [CrossRef]
- Mamouei, Z.; Singh, S.; Lemire, B.; Gu, Y.; Alqarihi, A.; Nabeela, S.; Li, D.; Ibrahim, A.; Uppuluri, P. An evolutionary diverged mitochondrial protein controls biofilm growth and virulence in Candida albicans. PLoS Biol. 2021, 19, e3000957. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.M.; Simon, S.; Wright, K.J.; Ndjountche, L.; Gaedigk, A. Tracking Trichophyton tonsurans through a large urban childcare center: Defining infection prevalence and transmission patterns by molecular stain typing. Pediatrics 2006, 118, 2365–2373. [Google Scholar] [CrossRef]
- Sidrim, J.J.C.; Rocha, M.F.G.; Leite, J.J.G.; da Albuquerque Maranhao, F.C.; Lima, R.A.C.; Castelo-Branco, D.S.C.M.; Bandeira, T.J.P.G.; Cordeiro, R.A.; Brilhante, R.S.N. Trichophyton tonsurans strains from Brazil: Phenotypic heterogeneity, genetic homology, and detection of virulence genes. Can. J. Microbiol. 2013, 59, 754–760. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.M.; Wright, K.J.; Navarre, H. Griseofulvin has only a modest impact on eradicating carriage of Trichophyton tonsurans. J. Pediatr. Pharmacol. Ther. 2009, 14, 94–99. [Google Scholar] [PubMed]
- Preuett, B.; Leeder, J.S.; Abdel-Rahman, S. Development and application of a high-throughput screening method to evaluate antifungal activity against Trichophyton tonsurans. J. Biomol. Screen. 2015, 20, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Waller, S.B.; Lana, D.F.D.; Quatrin, P.M.; Ferreira, M.R.A.; Fuentefria, A.M.; Mezzari, A. Antifungal resistance on Sporothrix species: An overview. Braz. J. Microbiol. 2021, 52, 73–80. [Google Scholar] [CrossRef]
- Xavier, M.O.; Poester, V.R.; Trápaga, M.R.; Stevens, D.A. Sporothrix brasiliensis: Epidemiology, therapy, and recent developments. J. Fungi 2023, 9, 921. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; de Andrade, I.B.; Ramos, M.L.M.; de Araújo Rodrigues, M.V.; do Nascimento, V.A.; Bernardes-Engemann, A.R.; Frases, S. Medicines for Malaria Venture COVID Box: A source for repurposing drugs with antifungal activity against human pathogenic fungi. Mem. Inst. Oswaldo Cruz 2021, 116, e210207. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.L.M.; Almeida-Silva, F.; de Souza Rabello, V.B.; Nahal, J.; Figueiredo-Carvalho, M.H.G.; Bernardes-Engemann, A.R.; Poester, V.R.; Xavier, M.O.; Meyer, W.; Zancopé-Oliveira, R.M.; et al. In vitro activity of the anthelmintic drug niclosamide against Sporothrix spp. strains with distinct genetic and antifungal susceptibility backgrounds. Braz. J. Microbiol. 2024; in press. [Google Scholar] [CrossRef]
- Van de Sande, W.W.J. Global burden of human mycetoma: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2013, 7, e2550. [Google Scholar] [CrossRef]
- Elkheir, L.Y.M.; Haroun, R.; Mohamed, M.A.; Fahal, A.H. Madurella mycetomatis causing eumycetoma medical treatment: The challenges and prospects. PLoS Negl. Trop. Dis. 2020, 14, e0008307. [Google Scholar] [CrossRef]
- World’s First Clinical Trial for Devastating Fungal Disease Mycetoma Shows Efficacy of New, Promising Treatment. Available online: https://dndi.org/press-releases/2023/worlds-first-clinical-trial-for-mycetoma-shows-efficacy-new-promising-treatment/ (accessed on 13 April 2024).
- Ma, J.; Todd, M.; van de Sande, W.W.J.; Biersack, B. Antifungal activity of natural naphthoquinones and anthraquinones against Madurella mycetomatis. Chem. Biodivers. 2023, 20, e202300151. [Google Scholar] [CrossRef]
- Mahmoud, A.B.; Abd Algaffar, S.; van de Sande, W.; Khalid, S.; Kaiser, M.; Mäser, P. Niclosamide is active in vitro against mycetoma pathogens. Molecules 2021, 26, 4005. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Blackwell, M.; James, T.Y.; Spatafora, J.W.; Taylor, J.W.; Vilgalys, R. Phylogenetic taxon definitions for Fungi, Dikarya, Ascomycota and Basidiomycota. IMA Fungi 2018, 9, 291–298. [Google Scholar] [CrossRef]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. Aids 2009, 23, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Bahn, Y.-S.; Sun, S.; Heitman, J.; Lin, X. Microbe profile: Cryptococcus neoformans species complex. Microbiology 2020, 166, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Bicanic, T.; Meintjes, G.; Wood, R.; Hayes, M.; Rebe, K.; Bekker, L.G.; Harrison, T. Fungaly burden, early fungicidal activity, and outcome in cryptococcal meningitis in antiretroviral-naïve and antiretroviral-experienced patients treated with amphotericin B or fluconazole. Clin. Infect. Dis. 2007, 45, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Dehdashti, S.J.; Abbott, J.; Nguyen, D.-T.; McKew, J.C.; Williamson, P.R.; Zheng, W. A high throughput screening assay for assessing viability of Cryptococcus neoformans under nutrient starvation condition. Anal. Bioanal. Chem. 2013, 405, 6823–6829. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, S.C.; Huang, M.; Hull, C.M. Spore germination as a target for antifungal therapeutics. Antimicrob. Agents Chemother. 2019, 63, e00994-19. [Google Scholar] [CrossRef] [PubMed]
- Beck, D.P.; Greenawalt, J.W. Biogenesis of mitochondrial membranes in Neurospora crassa during cellular differentiation: Changes in oxidative phosphorylation and synthesis of mitochondrial phospholipids. J. Gen. Microbiol. 1976, 92, 111–119. [Google Scholar] [CrossRef]
- Terenzi, H.F.; Storck, R. Stimulation of fermentation and yeast-like morphogenesis in Mucor rouxii by phenethyl alcohol. J. Bacteriol. 1969, 97, 1249–1261. [Google Scholar] [CrossRef]
- Broome, A.W.; Jones, G.M. A new drug for the treatment of fascioliasis in sheep and cattle. Nature 1966, 210, 744–745. [Google Scholar] [CrossRef]
- OpenSourceMycetoma. Available online: https://github.com/OpenSourceMycetoma (accessed on 28 March 2024).
- Pic, E.; Burgain, A.; Sellam, A. Repurposing the anthelmintic salicylanilide oxyclozanide against susceptible and clinical resistant Candida albicans strains. Med. Mycol. 2019, 57, 387–390. [Google Scholar] [CrossRef]
- Peyclit, L.; Yousfi, H.; Rolain, J.-M.; Bittar, F. Drug repurposing in medical mycology: Identification of compounds as potential antifungals to overcome the emergence of multidrug-resistant fungi. Pharmaceuticals 2021, 14, 488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bai, Y.; Li, B.; Zhou, X.; Si, H.; Zhang, J. Determination and pharmacokinetics study of oxyclozanide suspension in cattle by LC-MS/MS. BMC Vet. Res. 2019, 15, 210. [Google Scholar] [CrossRef]
- Wang, W.; Dong, Z.; Zhang, J.; Zhou, X.; Wei, X.; Cheng, F.; Li, B.; Zhang, J. Acute and subacute toxicity assessment of oxyclozanide in Wistar rats. Front. Vet. Sci. 2019, 6, 294. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.N.; Maia Filho, F.S.; Ferreira, G.F.; Mendes, J.F.; Goncalves, C.L.; Villela, M.M.; Pereira, D.I.B.; Nascente, P.S. In vitro susceptibility of nematophageous fungi to antiparasitic drugs: Interactions and implications for biological control. Braz. J. Biol. 2017, 77, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Michiels, M.; Meuldermans, W.; Heykants, J. The metabolism and fate of closantel (Flukiver) in sheep and cattle. Drug Metab. Rev. 1987, 18, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Swan, G.E.; Koeleman, H.A.; Steyn, H.S.; Mülders, M.S. Intravascular plasma disposition and salivary secretion of closantel and rafoxanide in sheep. J. S. Afr. Vet. Assoc. 1999, 70, 75–79. [Google Scholar] [CrossRef]
- Leary, E.O.; Gasior, S.; McElnea, E. Closantel toxicity. BMJ Case Rep. 2023, 16, e249626. [Google Scholar] [CrossRef]
- Bazvand, F.; Riazi-Esfahani, H.; Salari, F. Presumed veterinary niclosamide-induced retinal toxicity in a human: A case report. J. Med. Case Rep. 2023, 17, 110. [Google Scholar] [CrossRef] [PubMed]
- Swan, G.E.; Schröder, J. A safety trial with rafoxanide in sheep. J. S. Afr. Vet. Assoc. 1981, 52, 123–125. [Google Scholar]
- Schröder, J. The safety of injectable rafoxanide in cattle. J. S. Afr. Vet. Assoc. 1982, 53, 29–31. [Google Scholar]
- Bendary, M.M.; Abd El-Hamid, M.I.; Abousaty, A.I.; Elmanakhly, A.R.; Alshareef, W.A.; Mosbah, R.A.; Alhomrani, M.; Ghoneim, M.M.; Elkelish, A.; Hashim, N.; et al. Therapeutic switching of rafoxanide: A new approach to fighting drug-resistant bacteria and fungi. Microbiol. Spectr. 2023, 11, e0267922. [Google Scholar] [CrossRef] [PubMed]
- Norbury, H.M.; Waring, R.H. The disposition and metabolism of 3,4′,5-tribromosalicylanilide and 4′,5-dibromosalicylanilide in the rat. Xenobiotica 1981, 11, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Daidone, G.; Raffa, D.; Plescia, S.; Matera, M.; Caruso, A.; Leone, V.; Amico-Roxas, M. Synthesis and evaluation of the analgesic and antiinflammatory activities of N-substituted salicylamides. Farmaco 1989, 44, 465–473. [Google Scholar] [PubMed]
- Marhold, J. Prehled Prumyslove Toxicologie: Organicke Latky; Avicenum: Prague, Czech Republic, 1986; p. 663. [Google Scholar]
- Mitchell, S.C.; Norbury, H.M.; Waring, R.H.; Gadsden, P.M.; Wood, P.B. A comparison of the metabolism and elimination of benzanilide and salicylanilide in the rat. Xenobiotica 1982, 12, 93–99. [Google Scholar] [CrossRef]
- Krátký, M.; Vinšova, J. Salicylanilide ester prodrugs as potential antimicrobial agents–a review. Curr. Pharm. Des. 2011, 17, 3494–3505. [Google Scholar] [CrossRef] [PubMed]
- Krátky, M.; Vinšova, J.; Buchta, V.; Horvati, K.; Bösze, S.; Stolaříková, J. New amino acid esters of salicylanilides active against MDR-TB and other microbes. Eur. J. Med. Chem. 2010, 45, 5106–6113. [Google Scholar] [CrossRef] [PubMed]
- Krátký, M.; Vinšová, J.; Buchta, V. In vitro antibacterial and antifungal activity of salicylanilide benzoates. Sci. World. J. 2012, 2012, 290628. [Google Scholar] [CrossRef] [PubMed]
- Krátký, M.; Vinšová, J. Antifungal activity of salicylanilides and their esters with 4-(trifluoromethyl)benzoic acid. Molecules 2012, 17, 9426–9442. [Google Scholar] [CrossRef] [PubMed]
- Krátký, M.; Vinšová, J.; Buchta, V. In vitro antibacterial and antifungal activity of salicylanilide pyrazine-2-carboxylates. Med. Chem. 2012, 8, 732–741. [Google Scholar] [CrossRef]
- Krátký, M.; Vinšová, J. Salicylanilide N-monosubstituted carbamates: Synthesis and in vitro antimicrobial activity. Bioorg. Med. Chem. 2016, 24, 1322–1330. [Google Scholar] [CrossRef]
- Appetecchia, F.; Consalvi, S.; Scarpecci, C.; Biava, M.; Poce, G. SAR analysis of small molecules interfering with energy-metabolism in Mycobacterium tuberculosis. Pharmaceuticals 2020, 13, 227. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulos, G.; Monteiro, S.; Vosátka, R.; Krátký, M.; Navrátilová, L.; Treitnar, F.; Stolaříková, J.; Vinšová, J. Novel salicylanilides from 4,5-diahalogenated salicylic acids: Synthesis, antimicrobial activity and cytotoxicity. Bioorg. Med. Chem. 2017, 25, 1524–1532. [Google Scholar] [CrossRef]
- Ng, M.Y.; Song, Z.J.; Tan, C.H.; Bassetto, M.; Hagen, T. Structural investigations on the mitochondrial uncouplers niclosamide and FCCP. FEBS Open Bio 2024, in press. [CrossRef] [PubMed]
- Prichard, R.K. The metabolic profile of adult Fasciola hepatica obtained from rafoxanide-treated sheep. Parasitology 1978, 76, 277–288. [Google Scholar] [CrossRef]
- Hossain, S.; Veri, A.O.; Liu, Z.; Iyer, K.R.; O’Meara, T.R.; Robbins, N.; Cowen, L.E. Mitochondrial perturbation reduces susceptibility to xenobiotics through altered efflux in Candida albicans. Genetics 2021, 219, iyab095. [Google Scholar] [CrossRef]
- Cheng, S.; Clancy, C.J.; Nguyen, K.T.; Clapp, W.; Nguyen, M.H. A Candida albicans petite mutant strain with uncoupled oxidative phosphorylation overexpresses MDR1 and has diminished susceptibility to fluconazole and voriconazole. Antimicrob. Agents Chemother. 2007, 51, 1855–1858. [Google Scholar] [CrossRef]
- Liston, S.D.; Whitesell, L.; Kapoor, M.; Shaw, K.J.; Cowen, L.E. Enhanced efflux pump expression in Candida mutants results in decreased manogepix susceptibility. Antimicrob. Agents Chemother. 2020, 64, e00261-20. [Google Scholar] [CrossRef]
- Koltai, T. The complex relationship between multiple drug resistance and the tumor pH gradient: A review. Cancer Drug Resist. 2022, 5, 277–303. [Google Scholar] [CrossRef]
- Lionakis, M.S. Exploiting antifungal immunity in the clinical context. Semin. Immunol. 2023, 67, 101752. [Google Scholar] [CrossRef] [PubMed]
- Al-Gareeb, A.I.A.; Gorial, F.I.; Mahmood, A.S. Niclosamide as an adjuvant to etanercept in treatment patients with active rheumatoid arthritis: An 8-week randomized controlled pilot study. Clin. Rheumatol. 2018, 37, 2633–2641. [Google Scholar] [CrossRef]
- Krátký, M.; Dzurková, M.; Janoušek, J.; Konecná, K.; Trejtnar, F.; Stolaříková, J.; Vinšová, J. Sulfadiazine salicylaldehyde-based Schiff bases: Synthesis, antimicrobial activity and cytotoxicity. Molecules 2017, 22, 1573. [Google Scholar] [CrossRef]
- Bonilla, J.J.A.; Honorato, L.; Haranahalli, K.; Gremiao, I.D.F.; Pereira, S.A.; Guimaraes, A.; de Souza Baptista, A.R.; de Melo Tavares, P.; Rodrigues, M.L.; Miranda, K.; et al. Antifungal activity of acylhydrazone derivatives against Sporothrix spp. Antimicrob. Agents Chemother. 2021, 65, e02593-20. [Google Scholar]
- Ferreira, E.S.; Cordeiro, L.V.; Silva, D.F.; Souza, H.D.S.; de Athayde-Filho, P.F.; Barbosa-Filho, J.M.; Scotti, L.; Lima, E.O.; de Castro, R.D. Antifungal activity and mechanism of action of 2-chloro-N-phenylacetamide: A new molecule with activity against strains of Aspergillus flavus. An. Acad. Bras. Cienc. 2021, 93 (Suppl. 3), e20200997. [Google Scholar] [CrossRef]
- Diniz-Neto, H.; Silva, S.L.; Cordeiro, L.V.; Silva, D.F.; Oliveira, R.F.; Athayde-Filho, P.F.; Oliveira-Filho, A.A.; Guerra, F.Q.S.; Lima, E.O. Antifungal activity of 3-chloro-N-phenylacetamide: A new molecule with fungicidal and antibiofilm activity against fluconazole-resistant Candida spp. Braz. J. Biol. 2024, 84, e255080. [Google Scholar] [CrossRef]
- Silva, S.L.; de Oliveira Pereira, F.; Cordeiro, L.V.; Diniz-Neto, H.; dos Santos Maia, M.; da Silva Souza, H.D.; de Athayde-Filho, P.F.; Scotti, M.T.; Scotti, L.; de Oliveira Lima, E. Antifungal activity of 2-chloro-N-phenylacetamide, docking and molecular dynamics studies against clinical isolates of Candida tropicalis and Candida parapsilosis. J. Appl. Microbiol. 2022, 132, 3601–3617. [Google Scholar] [CrossRef]
- Dos Santos Ferreira, E.; Cordeiro, L.V.; de Figueredo Silva, D.; Diniz-Neto, H.; de Sousa, A.P.; da Silva Souza, H.D.; de Athayde-Filho, P.F.; Guerra, F.Q.S.; Barbosa-Filho, J.M.; de Oliveira Filho, A.A.; et al. Evaluatuon of antifungal activity, mechanisms of action and toxicological profile of the synthetic amide 2-chloro-N-phenylacetamide. Drug Chem. Toxicol. 2024, 47, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Copp, J.N.; Pletzer, D.; Brown, A.S.; van der Heijden, J.; Miton, C.M.; Edgar, R.J.; Rich, M.H.; Little, R.F.; Williams, E.M.; Hancock, R.E.W.; et al. Mechanistic understanding enables the rational design of salicylanilide combination therapies for Gram-negative infections. mBio 2020, 11, e02068-20. [Google Scholar] [CrossRef]
- Lateef, M.; Zargar, S.A.; Khan, A.R.; Nazir, M.; Shoukat, A. Successful treatment of niclosamide- and praziquantel-resistant beef tapeworm infection with nitazoxanide. Int. J. Infect. Dis. 2008, 12, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Song, H.-N.; Jeong, D.-G.; Bang, S.-Y.; Paek, S.-H.; Park, B.-C.; Park, S.-G.; Woo, E.-J. Crystal structure of the fungal nitroreducates Frm2 from Saccharomyces cerevisiae. Protein Sci. 2015, 24, 1159–1163. [Google Scholar] [CrossRef]
- Zhou, Y.; Lv, H.; Li, H.; Li, J.; Yan, Y.; Liu, F.; Hao, W.; Zhou, Z.; Wang, P.; Zhou, S. Nitroreductase increases menadione-mediated oxidative stress in Aspergillus nidulans. Appl. Environ. Microbiol. 2021, 87, e01758-21. [Google Scholar] [CrossRef]
- Wojcicki, A.; Chae, H.-D.; Han, K.; Youn, M.; Wilkes, M.C.; Lacayo, N.J.; Bassik, M.; Sakamoto, K.M. Genetic modulators of niclosamide sensitivity and resistance in acute myeloid leukemia. Blood 2020, 136 (Suppl. 1), 29. [Google Scholar] [CrossRef]
- Stover, K.R.; Hawkins, B.K.; Keck, J.M.; Barber, K.E.; Cretella, D.A. Antifungal resistance, combinations and pipeline: Oh my! Drugs Context 2023, 12, 2023-7-1. [Google Scholar]
- Wiederhold, N.P. Review of the novel investigational antifungal olorofim. J. Fungi 2020, 6, 122. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Orduña, G.R.; Ledesma-Martínez, E.; Aguiñiga-Sánchez, I.; Mora-García, M.d.L.; Weiss-Steider, B.; Santiago-Osorio, E. Inhibitors of chemoresistance pathways in combination with ara-C to overcome multidrug resistance in AML. A mini review. Int. J. Mol. Sci. 2021, 22, 4955. [Google Scholar] [CrossRef]
- Guo, Y.; Zhu, H.; Xiao, Y.; Guo, H.; Lin, M.; Yuan, Z.; Yang, X.; Huang, Y.; Zhang, Q.; Bai, Y. The anthelmintic drug niclosamide induces GSK-β-mediated β-catenin degradation to potentiate gemcitabine activity, reduce immune evasion ability and suppress pancreatic cancer progression. Cell Death Dis. 2022, 13, 112. [Google Scholar] [CrossRef]
- Vermes, A.; Guchelaar, H.-J.; Dankert, J. Flucytosine: A review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J. Antimicrob. Chemother. 2000, 46, 171–179. [Google Scholar] [CrossRef]
- Nitzsche, B.; Höpfner, M.; Biersack, B. Synthetic small molecule modulators of Hsp70 and Hsp40 chaperones as promising anticancer agents. Int. J. Mol. Sci. 2023, 24, 4083. [Google Scholar] [CrossRef]
- Rouges, C.; Asad, M.; Laurent, A.D.; Marchand, P.; Le Pape, P. Is the C-terminal domain an effective and selective target for the design of Hsp90 inhibitors against Candida yeast? Microorganisms 2023, 11, 2837. [Google Scholar] [CrossRef] [PubMed]
- Behboudi, H.; Alizadeh, Y.; Medghalchi, A.; Soltani-Moghadam, R.; Azaripour, E.; Moravvej, Z. Closantel retinal toxicity: Recovery from severe vision loss after corticosteroid therapy. Eur. J. Ophthalmol. 2023, 33, NP91–NP94. [Google Scholar] [CrossRef]
- Kang, M.-J.; Kim, M.J.; Kim, A.; Koo, T.-S.; Lee, K.-R.; Chae, Y.-J. Pharmacokinetic interactions of niclosamide in rats: Involvement of organic anion transporters 1 and 3 and organic cation transporter 2. Chem. Biol. Interact. 2024, 390, 110886. [Google Scholar] [CrossRef]
- Vermeer, L.M.M.; Isringhausen, C.D.; Ogilvie, B.W.; Buckley, D.B. Evaluation of ketoconazole and its alternative clinical CYP3A4/5 inhibitors as inhibitors of drug transporters: The in vitro effects of ketoconazole on 13 clinically-relevant drug transporters. Drug Metab. Dispos. 2016, 44, 453–459. [Google Scholar] [CrossRef]
- Xu, S.; Feliu, M.; Lord, A.K.; Lukason, D.P.; Negoro, P.E.; Khan, N.S.; Dagher, Z.; Feldman, M.B.; Reedy, J.L.; Steiger, S.N.; et al. Biguanides enhance antifungal activity against Candida glabrata. Virulence 2018, 9, 1150–1162. [Google Scholar] [CrossRef]
- Kang, H.E.; Seo, Y.; Yun, J.S.; Song, S.H.; Han, D.; Cho, E.S.; Cho, S.B.; Jeon, Y.; Lee, H.; Kim, H.S.; et al. Metformin and niclosamide synergistically suppress Wnt and YAP in APC-mutated colorectal cancer. Cancers 2021, 13, 343. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Weiss, A.; Goodman, J.; Fisk, M.; Kulkarni, S.; Lu, I.; Gray, J.; Smith, R.; Sommer, M.; Cheriyan, J. Niclosamide–a promising treatment for COVID-19. Br. J. Pharmacol. 2022, 179, 3250–3267. [Google Scholar] [CrossRef] [PubMed]
- Niyomdecha, N.; Suptawiwat, O.; Boonarkart, C.; Jitobaom, K.; Auewarakul, P. Inhibition of human immunodeficiency virus type 1 by niclosamide through mTORC1 inhibition. Heliyon 2020, 6, e04050. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Bano, G.; Malik, A.; Rasool, Y.; Manzar, S.; Singh, T.; Maity, M. Opportunistic fungal invasion in COVID-19 pandemic: A critical review in diagnosis and management. Avicenna J. Med. 2023, 13, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, Z.A.; Ghazanfar, H.; Altaf, F.; Ghazanfar, A.; Hasan, K.Z.; Kandhi, S.; Fortuzi, K.; Dileep, A.; Shrivistava, S. Cryptococcosis and cryptococcal meningitis: A narrative review and the up-to-date management approach. Cureus 2024, 16, e55498. [Google Scholar] [CrossRef]
- Needham, D. Extraction of niclosamide from commercial approved tablets into aqueous buffered solution creates potentially approvable oral and nasal sprays against COVID-19 and other respiratory infections. AAPS Open 2023, 9, 9. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biersack, B. The Antifungal Potential of Niclosamide and Structurally Related Salicylanilides. Int. J. Mol. Sci. 2024, 25, 5977. https://doi.org/10.3390/ijms25115977
Biersack B. The Antifungal Potential of Niclosamide and Structurally Related Salicylanilides. International Journal of Molecular Sciences. 2024; 25(11):5977. https://doi.org/10.3390/ijms25115977
Chicago/Turabian StyleBiersack, Bernhard. 2024. "The Antifungal Potential of Niclosamide and Structurally Related Salicylanilides" International Journal of Molecular Sciences 25, no. 11: 5977. https://doi.org/10.3390/ijms25115977
APA StyleBiersack, B. (2024). The Antifungal Potential of Niclosamide and Structurally Related Salicylanilides. International Journal of Molecular Sciences, 25(11), 5977. https://doi.org/10.3390/ijms25115977