
Dietary Curcumin Intake and Its Effects on the Transcriptome and Metabolome of Drosophila melanogaster

 , , , ,
, , , , 
 , and
, and
Abstract
1. Introduction
2. Results
2.1. Physiological Changes of Drosophila Melanogaster Fed with Curcumin
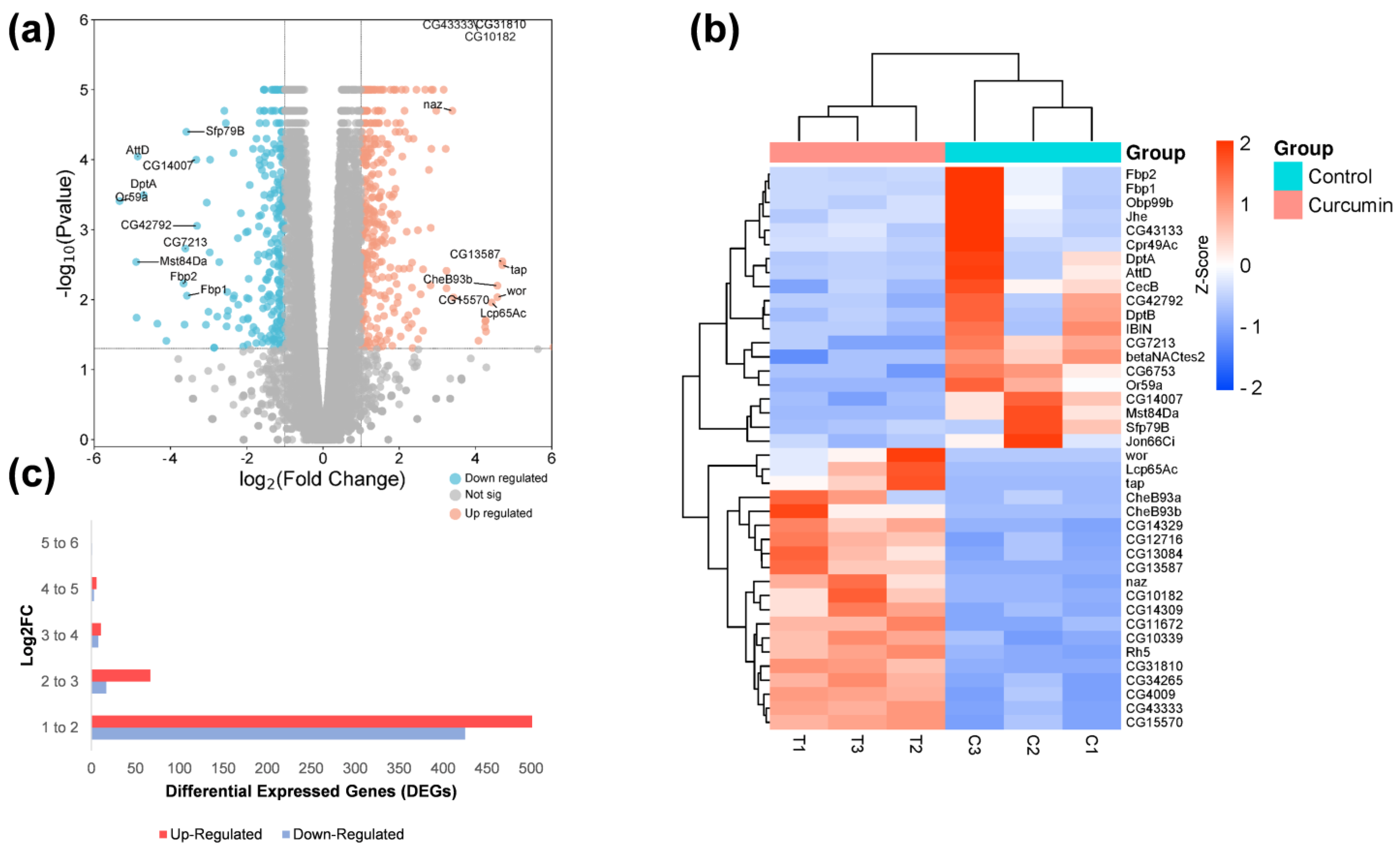
2.2. RNA-Seq Data of Flies in a Curcumin Diet
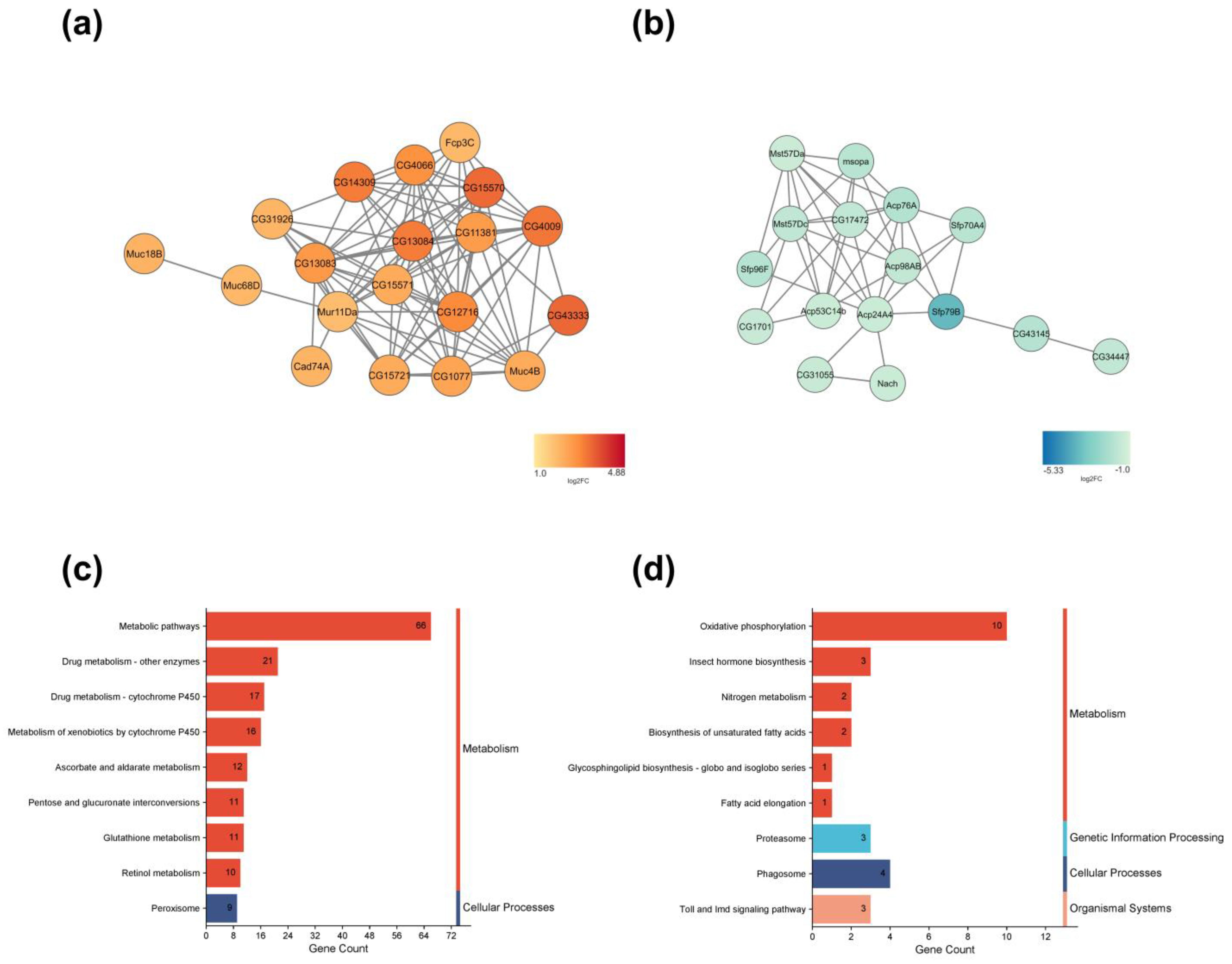
2.3. Gene Ontology, Networks, and Pathways Found in DEGs
2.4. DEGs in Response to CURCUMIN
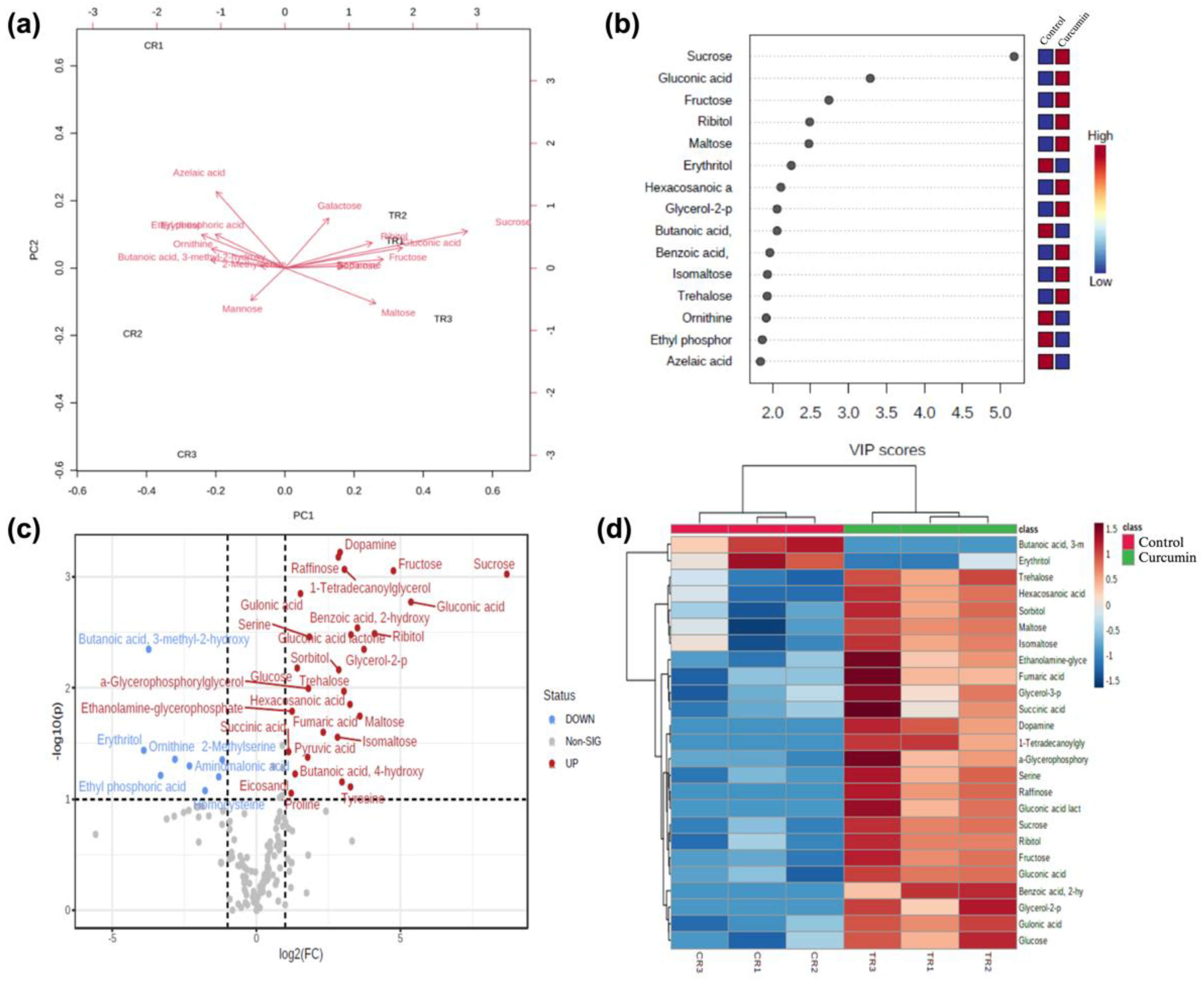
2.5. Metabolic Changes in Drosophila under Curcumin Treatment
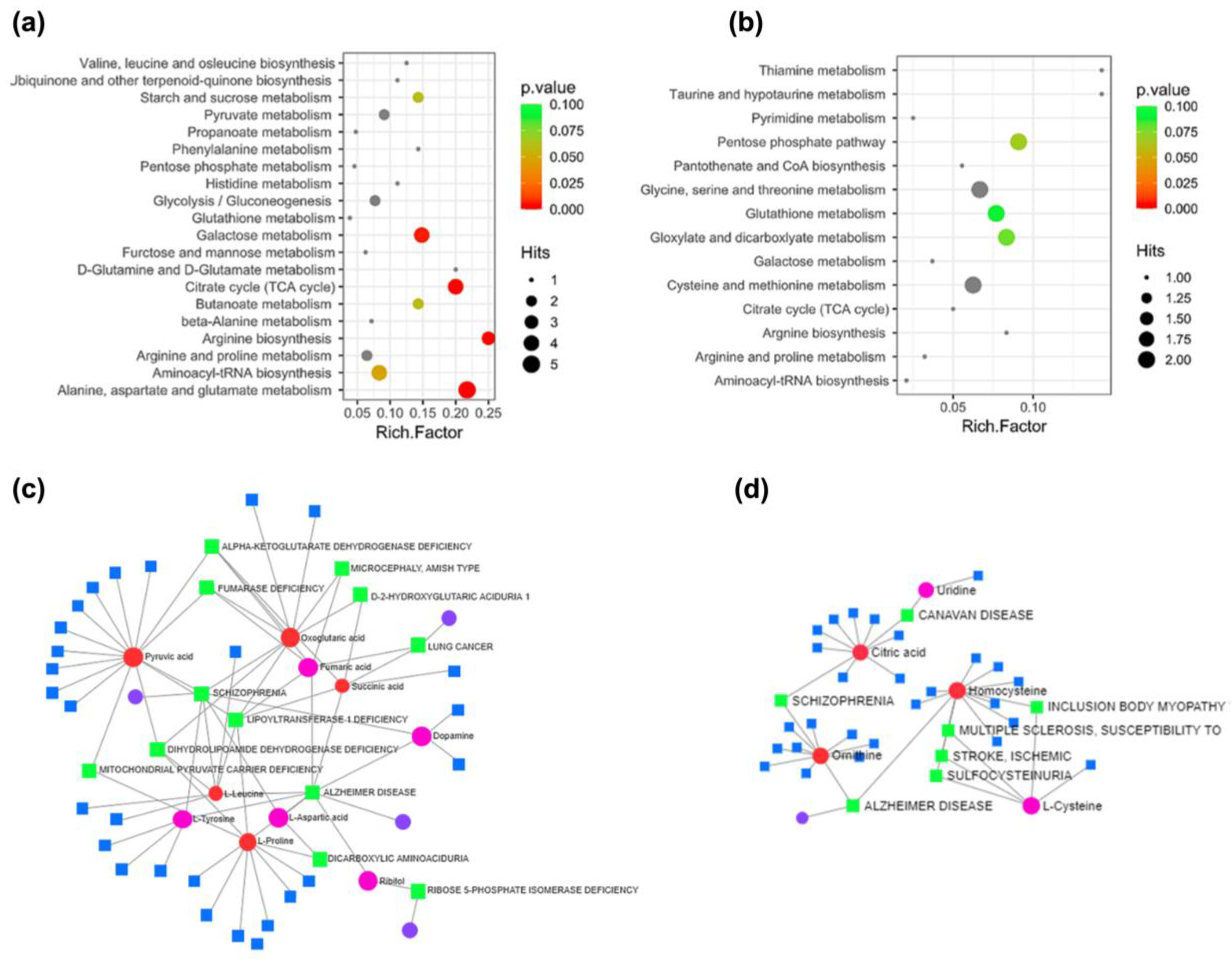
2.6. Enrichment Analysis of DAMs
2.7. Weighted Gene Co-Expression Network Analysis (WGCNA)
2.8. Transcriptome and Metabolome Integration
3. Discussion
4. Materials and Methods
4.1. Drosophila Husbandry and Experimental Diets
4.2. Food Intake and Physiological Assays
4.3. Total RNA Extraction and Transcriptomic Analysis
4.4. Metabolomics
4.5. Weighted Gene Co-Expression Network Analysis (WCGNA) and Integration of Transcriptome and Metabolome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kocaadam, B.; Şanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef]
- Salehi, B.; Stojanović-Radić, Z.; Matejić, J.; Sharifi-Rad, M.; Kumar, N.V.A.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of curcumin: A review of clinical trials. Eur. J. Med. Chem. 2019, 163, 527–545. [Google Scholar] [CrossRef]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial mechanism of curcumin: A review. Chem. Biodivers. 2020, 17, e2000171. [Google Scholar] [CrossRef]
- Tyagi, P.K.; Sharma, S.; Tyagi, S.; Mishra, A.; Gola, D. Toxicity assessment of silica nanoparticles, and their conjugates with curcumin on Drosophila melanogaster. Environ. Nanotechnol. Monit. Manag. 2022, 17, 100616. [Google Scholar]
- Sharifi-Rad, J.; Rayess, Y.E.; Rizk, A.A.; Sadaka, C.; Zgheib, R.; Zam, W.; Sestito, S.; Rapposelli, S.; Neffe-Skocińska, K.; Zielińska, D. Turmeric and its major compound curcumin on health: Bioactive effects and safety profiles for food, pharmaceutical, biotechnological and medicinal applications. Front. Pharmacol. 2020, 11, 1021. [Google Scholar] [CrossRef]
- Zia, A.; Farkhondeh, T.; Pourbagher-Shahri, A.M.; Samarghandian, S. The role of curcumin in aging and senescence: Molecular mechanisms. Biomed. Pharmacother. 2021, 134, 111119. [Google Scholar] [CrossRef]
- Kavukcu, S.B.; Çakır, S.; Karaer, A.; Türkmen, H.; Rethinam, S. Curcumin nanoparticles supported gelatin-collagen scaffold: Preparation, characterization, and in vitro study. Toxicol. Rep. 2021, 8, 1475–1479. [Google Scholar] [CrossRef]
- Ahmed, M.; Srivastava, U.; Gupta, C. Turmeric: Biological operations and medicinal applications. In Ethnopharmacological Investigation of Indian Spices; IGI Global: Hershey, PA, USA, 2020; pp. 163–172. [Google Scholar]
- Abadi, A.J.; Mirzaei, S.; Mahabady, M.K.; Hashemi, F.; Zabolian, A.; Hashemi, F.; Raee, P.; Aghamiri, S.; Ashrafizadeh, M.; Aref, A.R. Curcumin and its derivatives in cancer therapy: Potentiating antitumor activity of cisplatin and reducing side effects. Phytother. Res. 2022, 36, 189–213. [Google Scholar] [CrossRef]
- Aditi, K.; Singh, A.; Shakarad, M.N.; Agrawal, N. Management of altered metabolic activity in Drosophila model of Huntington’s disease by curcumin. Exp. Biol. Med. 2022, 247, 152–164. [Google Scholar] [CrossRef]
- Bagherniya, M.; Darand, M.; Askari, G.; Guest, P.C.; Sathyapalan, T.; Sahebkar, A. The clinical use of curcumin for the treatment of rheumatoid arthritis: A systematic review of clinical trials. In Studies on Biomarkers and New Targets in Aging Research in Iran: Focus on Turmeric and Curcumin; Springer: Cham, Switzerland, 2021; pp. 251–263. [Google Scholar]
- Kwiecien, S.; Magierowski, M.; Majka, J.; Ptak-Belowska, A.; Wojcik, D.; Sliwowski, Z.; Magierowska, K.; Brzozowski, T. Curcumin: A potent protectant against esophageal and gastric disorders. Int. J. Mol. Sci. 2019, 20, 1477. [Google Scholar] [CrossRef]
- Pivari, F.; Mingione, A.; Brasacchio, C.; Soldati, L. Curcumin and type 2 diabetes mellitus: Prevention and treatment. Nutrients 2019, 11, 1837. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Minghetti, L.; D’Archivio, M. Interaction between gut microbiota and curcumin: A new key of understanding for the health effects of curcumin. Nutrients 2020, 12, 2499. [Google Scholar] [CrossRef]
- Mansouri, K.; Rasoulpoor, S.; Daneshkhah, A.; Abolfathi, S.; Salari, N.; Mohammadi, M.; Rasoulpoor, S.; Shabani, S. Clinical effects of curcumin in enhancing cancer therapy: A systematic review. BMC Cancer 2020, 20, 791. [Google Scholar] [CrossRef]
- Chen, M.; Du, Z.-Y.; Zheng, X.; Li, D.-L.; Zhou, R.-P.; Zhang, K. Use of curcumin in diagnosis, prevention, and treatment of Alzheimer’s disease. Neural Regen. Res. 2018, 13, 742. [Google Scholar]
- Tang, S.B.; Lee, J.C.; Jung, J.K.; Choi, M.-Y. Effect of erythritol formulation on the mortality, fecundity and physiological excretion in Drosophila suzukii. J. Insect Physiol. 2017, 101, 178–184. [Google Scholar] [CrossRef]
- Ali, K.A.; Mona, M.; Bakr, A.F.; Mahmoud, M.Y.; Abdelgawad, E.M.; Matoock, M.Y. The dual gastro-and neuroprotective effects of curcumin loaded chitosan nanoparticles against cold restraint stress in rats. Biomed. Pharmacother. 2022, 148, 112778. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Fasae, K.D.; Iwezor, C.E.; Aschner, M.; Farombi, E.O. Curcumin attenuates copper-induced oxidative stress and neurotoxicity in Drosophila melanogaster. Toxicol. Rep. 2020, 7, 261–268. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A review of its effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Bahrami, A.; Montecucco, F.; Carbone, F.; Sahebkar, A. Effects of curcumin on aging: Molecular mechanisms and experimental evidence. BioMed Res. Int. 2021, 2021, 8972074. [Google Scholar] [CrossRef]
- Mołoń, M.; Dampc, J.; Kula-Maximenko, M.; Zebrowski, J.; Mołoń, A.; Dobler, R.; Durak, R.; Skoczowski, A. Effects of temperature on lifespan of Drosophila melanogaster from different genetic backgrounds: Links between metabolic rate and longevity. Insects 2020, 11, 470. [Google Scholar] [CrossRef]
- Sandner, G.; König, A.; Wallner, M.; Weghuber, J. Alternative model organisms for toxicological fingerprinting of relevant parameters in food and nutrition. Crit. Rev. Food Sci. Nutr. 2022, 62, 5965–5982. [Google Scholar] [CrossRef]
- Bongiorni, S.; Arisi, I.; Ceccantoni, B.; Rossi, C.; Cresta, C.; Castellani, S.; Forgione, I.; Rinalducci, S.; Muleo, R.; Prantera, G. Apple polyphenol diet extends lifespan, slows down mitotic rate and reduces morphometric parameters in Drosophila melanogaster: A comparison between three different apple cultivars. Antioxidants 2022, 11, 2086. [Google Scholar] [CrossRef]
- Ogunsuyi, O.B.; Aro, O.P.; Oboh, G.; Olagoke, O.C. Curcumin improves the ability of donepezil to ameliorate memory impairment in Drosophila melanogaster: Involvement of cholinergic and cnc/Nrf2-redox systems. Drug Chem. Toxicol. 2023, 46, 1035–1043. [Google Scholar] [CrossRef]
- Akinyemi, A.J.; Oboh, G.; Ogunsuyi, O.; Abolaji, A.O.; Udofia, A. Curcumin-supplemented diets improve antioxidant enzymes and alter acetylcholinesterase genes expression level in Drosophila melanogaster model. Metab. Brain Dis. 2018, 33, 369–375. [Google Scholar] [CrossRef]
- Pisaruk, A.; Koshel, N.; Mekhova, L.; Zabuga, O.; Ivanov, S. Influense of curcumin on lifespan if it is applied at the larval stage of Drosophila melanogaster. Ageing Longev. 2020, 1, 89–96. [Google Scholar] [CrossRef]
- Dhankhar, J.; Shrivastava, A.; Agrawal, N. Amendment of Altered Immune Response by Curcumin in Drosophila Model of Huntington’s Disease. J. Huntington’s Dis. 2023, 12, 335–354. [Google Scholar] [CrossRef]
- Pandareesh, M.; Shrivash, M.; Naveen Kumar, H.; Misra, K.; Srinivas Bharath, M. Curcumin monoglucoside shows improved bioavailability and mitigates rotenone induced neurotoxicity in cell and Drosophila models of Parkinson’s disease. Neurochem. Res. 2016, 41, 3113–3128. [Google Scholar] [CrossRef]
- Laurie, S.; Ainslie, L.; Mitchell, S.; Morimoto, J. Turmeric shortens lifespan in houseflies. Front. Insect Sci. 2024, 4, 1376011. [Google Scholar] [CrossRef]
- von Hellfeld, R.; Christie, C.; Derous, D.; Morimoto, J. Super food or super toxic? Turmeric and spirulina as culprits for the toxic effects of food dyes in Drosophila. J. Insect Physiol. 2024, 153, 104600. [Google Scholar] [CrossRef]
- Concetta Scuto, M.; Mancuso, C.; Tomasello, B.; Laura Ontario, M.; Cavallaro, A.; Frasca, F.; Maiolino, L.; Trovato Salinaro, A.; Calabrese, E.J.; Calabrese, V. Curcumin, hormesis and the nervous system. Nutrients 2019, 11, 2417. [Google Scholar] [CrossRef]
- Scuto, M.; Rampulla, F.; Reali, G.M.; Spanò, S.M.; Trovato Salinaro, A.; Calabrese, V. Hormetic Nutrition and Redox Regulation in Gut–Brain Axis Disorders. Antioxidants 2024, 13, 484. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, N.S.A.; Oskouie, M.N.; Butler, A.E.; Petit, P.X.; Barreto, G.E.; Sahebkar, A. Hormetic effects of curcumin: What is the evidence? J. Cell. Physiol. 2019, 234, 10060–10071. [Google Scholar] [CrossRef]
- Mew, M.; Caldwell, K.A.; Caldwell, G.A. From bugs to bedside: Functional annotation of human genetic variation for neurological disorders using invertebrate models. Hum. Mol. Genet. 2022, 31, R37–R46. [Google Scholar] [CrossRef] [PubMed]
- Soh, J.-W.; Marowsky, N.; Nichols, T.J.; Rahman, A.M.; Miah, T.; Sarao, P.; Khasawneh, R.; Unnikrishnan, A.; Heydari, A.R.; Silver, R.B. Curcumin is an early-acting stage-specific inducer of extended functional longevity in Drosophila. Exp. Gerontol. 2013, 48, 229–239. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Yang, T.; Korma, S.A.; Sitohy, M.; Abd El-Mageed, T.A.; Selim, S.; Al Jaouni, S.K.; Salem, H.M.; Mahmmod, Y.; Soliman, S.M. Impacts of turmeric and its principal bioactive curcumin on human health: Pharmaceutical, medicinal, and food applications: A comprehensive review. Front. Nutr. 2023, 9, 1040259. [Google Scholar] [CrossRef]
- Kamburov, A.; Cavill, R.; Ebbels, T.M.; Herwig, R.; Keun, H.C. Integrated pathway-level analysis of transcriptomics and metabolomics data with IMPaLA. Bioinformatics 2011, 27, 2917–2918. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Liu, Y.; Wang, L.; Wang, H.; Hu, Z.; Dong, H.; Yu, Z.; Yuan, Y. A review of curcumin in food preservation: Delivery system and photosensitization. Food Chem. 2023, 424, 136464. [Google Scholar] [CrossRef]
- Obeid, M.A.; Alsaadi, M.; Aljabali, A.A. Recent updates in curcumin delivery. J. Liposome Res. 2023, 33, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Morón, E.; Calderón-Montaño, J.M.; Salvador, J.; Robles, A.; López-Lázaro, M. The dark side of curcumin. Int. J. Cancer 2010, 126, 1771–1775. [Google Scholar] [CrossRef]
- Filardi, T.; Varì, R.; Ferretti, E.; Zicari, A.; Morano, S.; Santangelo, C. Curcumin: Could this compound be useful in pregnancy and pregnancy-related complications? Nutrients 2020, 12, 3179. [Google Scholar] [CrossRef]
- Naz, R.K. Can curcumin provide an ideal contraceptive? Mol. Reprod. Dev. 2011, 78, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.G.; Klämbt, C. The Drosophila blood-brain barrier emerges as a model for understanding human brain diseases. Neurobiol. Dis. 2023, 180, 106071. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.-R.; Xiao, F.; Yuan, P.; Chen, Y.; Gao, Q.-K.; Parnell, L.D.; Meydani, M.; Ordovas, J.M.; Li, D.; Lai, C.-Q. Curcumin-supplemented diets increase superoxide dismutase activity and mean lifespan in Drosophila. Age 2013, 35, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Lee, B.-S.; Semnani, S.; Avanesian, A.; Um, C.-Y.; Jeon, H.-J.; Seong, K.-M.; Yu, K.; Min, K.-J.; Jafari, M. Curcumin extends life span, improves health span, and modulates the expression of age-associated aging genes in Drosophila melanogaster. Rejuvenation Res. 2010, 13, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Suckow, B.K.; Suckow, M.A. Lifespan extension by the antioxidant curcumin in Drosophila melanogaster. Int. J. Biomed. Sci. IJBS 2006, 2, 402. [Google Scholar] [CrossRef]
- Hoffman, T.R.; Emsley, S.A.; Douglas, J.C.; Reed, K.R.; Esquivel, A.R.; Koyack, M.J.; Paddock, B.E.; Videau, P. Assessing Curcumin Uptake and Clearance and Their Influence on Superoxide Dismutase Activity in Drosophila melanogaster. BioTech 2023, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.J.; Musachio, E.A.S.; Meichtry, L.B.; Janner, D.E.; Balok, F.R.M.; Barrientos, M.S.; Gomes, N.S.; Pinto, K.H.P.; Dahleh, M.M.M.; Rodrigues, A. Evaluation of oxidative stress indicators as toxicity parameters after chronic exposure of Drosophila melanogaster to free curcumin and curcumin-loaded nanocapsules. Food Chem. Toxicol. 2023, 181, 114109. [Google Scholar] [CrossRef] [PubMed]
- Oyetayo, B.O.; Abolaji, A.O.; Fasae, K.D.; Aderibigbe, A. Ameliorative role of diets fortified with Curcumin in a Drosophila melanogaster model of aluminum chloride-induced neurotoxicity. J. Funct. Foods 2020, 71, 104035. [Google Scholar] [CrossRef]
- Esquivel, A.R.; Douglas, J.C.; Loughran, R.M.; Rezendes, T.E.; Reed, K.R.; Cains, T.H.; Emsley, S.A.; Paddock, W.A.; Videau, P.; Koyack, M.J. Assessing the influence of curcumin in sex-specific oxidative stress, survival and behavior in Drosophila melanogaster. J. Exp. Biol. 2020, 223, jeb223867. [Google Scholar] [CrossRef]
- Khyati; Malik, I.; Agrawal, N.; Kumar, V. Melatonin and curcumin reestablish disturbed circadian gene expressions and restore locomotion ability and eclosion behavior in Drosophila model of Huntington’s disease. Chronobiol. Int. 2021, 38, 61–78. [Google Scholar] [CrossRef]
- Caesar, I.; Jonson, M.; Nilsson, K.P.R.; Thor, S.; Hammarström, P. Curcumin promotes A-beta fibrillation and reduces neurotoxicity in transgenic Drosophila. PLoS ONE 2012, 7, e31424. [Google Scholar] [CrossRef] [PubMed]
- Bayliak, M.M.; Demianchuk, O.I.; Gospodaryov, D.V.; Balatskyi, V.A.; Lushchak, V.I. Specific and combined effects of dietary ethanol and arginine on Drosophila melanogaster. Drug Chem. Toxicol. 2023, 46, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Canfield, C.-A.; Bradshaw, P.C. Amino acids in the regulation of aging and aging-related diseases. Transl. Med. Aging 2019, 3, 70–89. [Google Scholar] [CrossRef]
- Gómez-A, A.; Shnitko, T.A.; Caref, K.L.; Nicola, S.M.; Robinson, D.L. Stimuli predicting high-calorie reward increase dopamine release and drive approach to food in the absence of homeostatic need. Nutr. Neurosci. 2022, 25, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Dall’Acqua, S.; Stocchero, M.; Boschiero, I.; Schiavon, M.; Golob, S.; Uddin, J.; Voinovich, D.; Mammi, S.; Schievano, E. New findings on the in vivo antioxidant activity of Curcuma longa extract by an integrated 1H NMR and HPLC–MS metabolomic approach. Fitoterapia 2016, 109, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Acharya, A.; Ray, R.; Agrawal, R.; Raghuwanshi, R.; Jain, P. Cellular and molecular mechanisms of curcumin in prevention and treatment of disease. Crit. Rev. Food Sci. Nutr. 2020, 60, 887–939. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Yang, H.-T.; Lane, H.-Y. D-glutamate, D-serine, and D-alanine differ in their roles in cognitive decline in patients with Alzheimer’s disease or mild cognitive impairment. Pharmacol. Biochem. Behav. 2019, 185, 172760. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.-A.A.; Ren, W.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Nuru, M.; Muradashvili, N.; Kalani, A.; Lominadze, D.; Tyagi, N. High methionine, low folate and low vitamin B6/B12 (HM-LF-LV) diet causes neurodegeneration and subsequent short-term memory loss. Metab. Brain Dis. 2018, 33, 1923–1934. [Google Scholar] [CrossRef]
- Khayatan, D.; Razavi, S.M.; Arab, Z.N.; Hosseini, Y.; Niknejad, A.; Momtaz, S.; Abdolghaffari, A.H.; Sathyapalan, T.; Jamialahmadi, T.; Kesharwani, P. Superoxide dismutase: A key target for the neuroprotective effects of curcumin. Mol. Cell. Biochem. 2024, 479, 693–705. [Google Scholar] [CrossRef]
- Priyadarsini, K.I.; Maity, D.K.; Naik, G.; Kumar, M.S.; Unnikrishnan, M.; Satav, J.; Mohan, H. Role of phenolic OH and methylene hydrogen on the free radical reactions and antioxidant activity of curcumin. Free Radic. Biol. Med. 2003, 35, 475–484. [Google Scholar] [CrossRef]
- Patriarca, E.J.; Cermola, F.; D’Aniello, C.; Fico, A.; Guardiola, O.; De Cesare, D.; Minchiotti, G. The multifaceted roles of proline in cell behavior. Front. Cell Dev. Biol. 2021, 9, 728576. [Google Scholar] [CrossRef]
- Li, H.; Yang, P.; Knight, W.; Guo, Y.; Perlmutter, J.S.; Benzinger, T.L.; Morris, J.C.; Xu, J. The interactions of dopamine and oxidative damage in the striatum of patients with neurodegenerative diseases. J. Neurochem. 2020, 152, 235–251. [Google Scholar] [CrossRef]
- Pitoniak, A.; Bohmann, D. Mechanisms and functions of Nrf2 signaling in Drosophila. Free Radic. Biol. Med. 2015, 88, 302–313. [Google Scholar] [CrossRef]
- Urošević, M.; Nikolić, L.; Gajić, I.; Nikolić, V.; Dinić, A.; Miljković, V. Curcumin: Biological activities and modern pharmaceutical forms. Antibiotics 2022, 11, 135. [Google Scholar] [CrossRef]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Gancarz, M.; Kondracka, A.; Rusinek, R.; Oniszczuk, A. Curcumin and weight loss: Does it work? Int. J. Mol. Sci. 2022, 23, 639. [Google Scholar] [CrossRef]
- Belfort-DeAguiar, R.; Seo, D. Food cues and obesity: Overpowering hormones and energy balance regulation. Curr. Obes. Rep. 2018, 7, 122–129. [Google Scholar] [CrossRef]
- Montague, S.A.; Mathew, D.; Carlson, J.R. Similar odorants elicit different behavioral and physiological responses, some supersustained. J. Neurosci. 2011, 31, 7891–7899. [Google Scholar] [CrossRef]
- Kreher, S.A.; Kwon, J.Y.; Carlson, J.R. The molecular basis of odor coding in the Drosophila larva. Neuron 2005, 46, 445–456. [Google Scholar] [CrossRef]
- Yuan, L.; Hu, S.; Okray, Z.; Ren, X.; De Geest, N.; Claeys, A.; Yan, J.; Bellefroid, E.; Hassan, B.A.; Quan, X.-J. The Drosophila neurogenin tap functionally interacts with the Wnt-PCP pathway to regulate neuronal extension and guidance. Development 2016, 143, 2760–2766. [Google Scholar] [CrossRef]
- Ledent, V.; Gaillard, F.; Gautier, P.; Ghysen, A.; Dambly-Chaudiere, C. Expression and function of tap in the gustatory and olfactory organs of Drosophila. Int. J. Dev. Biol. 2002, 42, 163–170. [Google Scholar]
- Saadati, S.; Hatami, B.; Yari, Z.; Shahrbaf, M.A.; Eghtesad, S.; Mansour, A.; Poustchi, H.; Hedayati, M.; Aghajanpoor-Pasha, M.; Sadeghi, A. The effects of curcumin supplementation on liver enzymes, lipid profile, glucose homeostasis, and hepatic steatosis and fibrosis in patients with non-alcoholic fatty liver disease. Eur. J. Clin. Nutr. 2019, 73, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-Y.; Lucas, H.; Sagili, R.; Cha, D.H.; Lee, J.C. Effect of erythritol on Drosophila suzukii (Diptera: Drosophilidae) in the presence of naturally-occurring sugar sources, and on the survival of Apis mellifera (Hymenoptera: Apidae). J. Econ. Entomol. 2019, 112, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Chaturvedi, M.; Mishra, S.; Kumar, P.; Somvanshi, P.; Chaturvedi, R. Reductive metabolites of curcumin and their therapeutic effects. Heliyon 2020, 6, e05469. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Swelum, A.A.; Arif, M.; Abo Ghanima, M.M.; Shukry, M.; Noreldin, A.; Taha, A.E.; El-Tarabily, K.A. Curcumin, the active substance of turmeric: Its effects on health and ways to improve its bioavailability. J. Sci. Food Agric. 2021, 101, 5747–5762. [Google Scholar] [CrossRef]
- Davis, J.M.; Murphy, E.A.; Carmichael, M.D.; Zielinski, M.R.; Groschwitz, C.M.; Brown, A.S.; Gangemi, J.D.; Ghaffar, A.; Mayer, E.P. Curcumin effects on inflammation and performance recovery following eccentric exercise-induced muscle damage. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 292, R2168–R2173. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.M.; Almassey, M.; Burch, A.M.; Ngo, G.; Martin, J.M.; Myers, D.; Compton, D.; Archie, S.; Cross, M.; Naeger, L. Drosophila p38 MAPK interacts with BAG-3/starvin to regulate age-dependent protein homeostasis. Aging Cell 2021, 20, e13481. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Yi, P.; Li, X.; Olson, E.N. Hand, an evolutionarily conserved bHLH transcription factor required for Drosophila cardiogenesis and hematopoiesis. Development 2006, 133, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.M.; Primrose, D.A.; Hodgetts, R.B. A member of the p38 mitogen-activated protein kinase family is responsible for transcriptional induction of Dopa decarboxylase in the epidermis of Drosophila melanogaster during the innate immune response. Mol. Cell. Biol. 2008, 28, 4883–4895. [Google Scholar] [CrossRef]
- Howard, S.A.; Benhabbour, S.R. Non-Hormonal Contraception. J. Clin. Med. 2023, 12, 4791. [Google Scholar] [CrossRef]
- Koca, S.B.; Ongun, O.; Ozmen, O.; Yigit, N.O. Subfertility effects of turmeric (Curcuma longa) on reproductive performance of Pseudotropheus acei. Anim. Reprod. Sci. 2019, 202, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Giannubilo, S.R.; Marzioni, D. The multifaced actions of curcumin in pregnancy outcome. Antioxidants 2021, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.-L.; Chen, X.; Zong, Q.; Zhao, T.; Wang, J.-L.; Zheng, Y.; Zhang, M.; Wang, Z.; Brownlie, J.C.; Yang, F. Quantitative proteomic analyses of molecular mechanisms associated with cytoplasmic incompatibility in Drosophila melanogaster induced by Wolbachia. J. Proteome Res. 2015, 14, 3835–3847. [Google Scholar] [CrossRef] [PubMed]
- Findlay, G.D.; Yi, X.; MacCoss, M.J.; Swanson, W.J. Proteomics reveals novel Drosophila seminal fluid proteins transferred at mating. PLoS Biol. 2008, 6, e178. [Google Scholar] [CrossRef] [PubMed]
- Bayliak, M.M.; Lylyk, M.P.; Maniukh, O.V.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. Dietary L-arginine accelerates pupation and promotes high protein levels but induces oxidative stress and reduces fecundity and life span in Drosophila melanogaster. J. Comp. Physiol. B 2018, 188, 37–55. [Google Scholar] [CrossRef]
- Dai, C.; Lin, J.; Li, H.; Shen, Z.; Wang, Y.; Velkov, T.; Shen, J. The natural product curcumin as an antibacterial agent: Current achievements and problems. Antioxidants 2022, 11, 459. [Google Scholar] [CrossRef] [PubMed]
- Neyestani, Z.; Ebrahimi, S.A.; Ghazaghi, A.; Jalili, A.; Sahebkar, A.; Rahimi, H.R. Review of anti-bacterial activities of curcumin against Pseudomonas aeruginosa. Crit. Rev.™ Eukaryot. Gene Expr. 2019, 29, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.; Kondo, S.; Lemaitre, B. Drosophila immunity: The Drosocin gene encodes two host defence peptides with pathogen-specific roles. Proc. R. Soc. B 2022, 289, 20220773. [Google Scholar] [CrossRef] [PubMed]
- Carboni, A.L.; Hanson, M.A.; Lindsay, S.A.; Wasserman, S.A.; Lemaitre, B. Cecropins contribute to Drosophila host defense against a subset of fungal and Gram-negative bacterial infection. Genetics 2022, 220, iyab188. [Google Scholar] [CrossRef]
- Uvell, H.; Engstrüm, Y. Functional characterization of a novel promoter element required for an innate immune response in Drosophila. Mol. Cell. Biol. 2003, 23, 8272–8281. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, X.; Cheng, J.-H.; Zeng, G.; Sun, D.-W. Synthesis and antimicrobial activities of novel sorbic and benzoic acid amide derivatives. Food Chem. 2018, 268, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Ham, S.-Y.; Kim, H.-S.; Cha, E.; Lim, T.; Byun, Y.; Park, H.-D. Raffinose inhibits Streptococcus mutans biofilm formation by targeting glucosyltransferase. Microbiol. Spectr. 2022, 10, e02076-21. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ortiz, C.; Edwards, M.; Natarajan, P.; Pacheco-Valenciana, A.; Nimmakayala, P.; Adjeroh, D.A.; Sirbu, C.; Reddy, U.K. Peppers in Diet: Genome-Wide Transcriptome and Metabolome Changes in Drosophila melanogaster. Int. J. Mol. Sci. 2022, 23, 9924. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, S.; Jansen, A.; Jois, S.; Kastenholz, K.; Escarcena, L.V.; Strudthoff, N.; Scholz, H. The CApillary FEeder assay measures food intake in Drosophila melanogaster. JoVE (J. Vis. Exp.) 2017, e55024. [Google Scholar] [CrossRef]
- Sullivan, G.M.; Feinn, R. Using effect size—Or why the P value is not enough. J. Grad. Med. Educ. 2012, 4, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Pei, G.; Chen, L.; Zhang, W. WGCNA application to proteomic and metabolomic data analysis. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 585, pp. 135–158. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Symbol | Differential Expression | Function | Biological Process |
|---|---|---|---|---|
| FBgn0004552 | Akh | Up | Hormone activity | Glucose and lipid homeostasis |
| FBgn0038449 | CG17562 | Up | alcohol-forming very long-chain fatty acyl-CoA reductase activity | long-chain fatty-acyl-CoA metabolic process |
| FBgn0033216 | CG1946 | Up | diacylglycerol O-acyltransferase activity | Triglyceride biosynthetic process |
| FBgn0033204 | CG2065 | Up | NADP-retinol dehydrogenase activity | Retinal metabolic process |
| FBgn0038469 | CG4009 | Up | Peroxidase activity | Response to oxidative stress |
| FBgn0034909 | CG4797 | Up | Transport activity | Carbohydrate transport |
| FBgn0034142 | CG8306 | Up | alcohol-forming very long-chain fatty acyl-CoA reductase activity | long-chain fatty-acyl-CoA metabolic process |
| FBgn0034756 | Cyp6d2 | Up | Oxidoreductase activity | Stress response |
| FBgn0031461 | dawdl | Up | Cytokine activity | cellular response to nutrient levels |
| FBgn0029172 | Fad2 | Up | Oxidoreductase activity | Lipid metabolic process |
| FBgn0283427 | FASN1 | Up | fatty acid synthase activity | Lipid metabolic process |
| FBgn0039114 | Lsd-1 | Up | Lipid storage | |
| FBgn0023479 | tequila | Up | serine-type endopeptidase activity | Glucose homeostasis |
| FBgn0030748 | Traf-like | Up | Protein binding activity | positive regulation of lipophagy |
| FBgn0038069 | CG11608 | Down | Triglyceride lipase activity | Lipid metabolic process |
| FBgn0042207 | CG18530 | Down | Triglyceride lipase activity | Lipid metabolic process |
| FBgn0038070 | CG6753 | Down | Triglyceride lipase activity | Lipid metabolic process |
| Biological Pathway | Function | Genes | Gene Symbol | Metabolites | Metabolite Name | Joint (p-Value) |
|---|---|---|---|---|---|---|
| Arginine Proline metabolism | Biosynthesis of arginine, ornithine, proline, citrulline and glutamate | 5 | GCLC; GAD1; ALDH7A1; OAT; ODC1 | 9 | Glycine; a-Ketoglutaric acid; UREA; Succinic acid; Aspartic acid; Spermidine; GABA; Putrescine; Fumaric acid | 4.26 × 10−11 |
| Alanine and aspartate metabolism | Source of energy, gluconeogenesis, and NH3 transport | 1 | GAD1 | 7 | Tricarballylic acid citric; pyruvic acid; a-Ketoglutaric acid; Aspartic acid; Succinic acid; Asparagine; Fumaric acid | 3.87 × 10−8 |
| Glutamate Glutamine metabolism | Substrate for nucleotide synthesis, NADPH, antioxidants, and NH3 transport | 3 | GCLC; GAD1; ALDH7A1 | 6 | pyruvic acid; Glycine; a-Ketoglutaric acid; Succinic acid; Aspartic acid; GABA | 3.91 × 10−8 |
| G Protein Coupled Receptors downstream signaling | Activate cAMP and inositol phospholipids signaling | 1 | MMP1 | 17 | Nicotinic acid; glycerol-3-p; glycerol; Histamine; Palmitic acid; Pentadecanoic acid; Dopamine; stearic acid; a-Ketoglutaric acid; GABA; Tetradecanoic acid; Dodecanoic ACID; 9-Hexadecenoic acid; Succinic acid; 9-Octadecenoic acid; Adenosine; sucrose | 8.02 × 10−8 |
| Protein digestion and absorption | Proteolytic activity for amino acids and small peptides production | 1 | COL4A1 | 10 | Histamine; Threonine; Glycine; Asparagine; valine; isoleucine; Aspartic acid; Tyrosine; Putrescine; Histidine | 8.04 × 10−8 |
| Glycine_serine and threonine metabolism | Essential amino acid and intermediate of glycolysis | 3 | GNMT; SRR; ALDH7A1 | 7 | homoserine; Threonine; pyruvic acid; Glyceric acid; Glycine; glyoxylic acid; Aspartic acid | 1.32 × 10−7 |
| Signaling by GPCR | Promote heterotrimeric GTP-binding protein activation, acts as a guanine nucleotide exchange factor | 1 | MMP1 | 18 | NICOTINIC ACID; glycerol-3-p; glycerol; Histamine; Palmitic acid; Pentadecanoic acid; Dopamine; stearic acid; Dodecanoic ACID; GABA; Tetradecanoic acid; a-Ketoglutaric acid; 9-Hexadecenoic acid; Succinic acid; 9-Octadecenoic acid; Adenosine; KYNURENIC ACID; sucrose | 3.74 × 10−7 |
| Glyoxylate and dicarboxylate metabolism | Metabolism of fatty acids and glutamate | 1 | CAT | 10 | Tricarballylic acid#citric acid; Glycolic acid; pyruvic acid; Glyceric acid; Glycine; a-Ketoglutaric acid; glyoxylic acid; oxalic acid; Succinic acid; Phosphoglycolic acid | 3.80 × 10−7 |
| Purine Metabolism | Maintain optimal level of nucleotides | 2 | GART; PFAS | 9 | Hypoxanthine; Xanthine; Adenine; Guanosine; Glycine; Aspartic acid; Inosine; Adenosine; Fumaric acid | 7.91 × 10−7 |
| Purine Nucleoside Phosphorylase Deficiency | Immune system | 2 | GART; PFAS | 9 | Hypoxanthine; Xanthine; Adenine; Guanosine; Glycine; Aspartic acid; Inosine; Adenosine; Fumaric acid | 7.91 × 10−7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belcher, S.; Flores-Iga, G.; Natarajan, P.; Crummett, G.; Talavera-Caro, A.; Gracia-Rodriguez, C.; Lopez-Ortiz, C.; Das, A.; Adjeroh, D.A.; Nimmakayala, P.; et al. Dietary Curcumin Intake and Its Effects on the Transcriptome and Metabolome of Drosophila melanogaster. Int. J. Mol. Sci. 2024, 25, 6559. https://doi.org/10.3390/ijms25126559
Belcher S, Flores-Iga G, Natarajan P, Crummett G, Talavera-Caro A, Gracia-Rodriguez C, Lopez-Ortiz C, Das A, Adjeroh DA, Nimmakayala P, et al. Dietary Curcumin Intake and Its Effects on the Transcriptome and Metabolome of Drosophila melanogaster. International Journal of Molecular Sciences. 2024; 25(12):6559. https://doi.org/10.3390/ijms25126559
Chicago/Turabian StyleBelcher, Samantha, Gerardo Flores-Iga, Purushothaman Natarajan, Garrett Crummett, Alicia Talavera-Caro, Celeste Gracia-Rodriguez, Carlos Lopez-Ortiz, Amartya Das, Donald A. Adjeroh, Padma Nimmakayala, and et al. 2024. "Dietary Curcumin Intake and Its Effects on the Transcriptome and Metabolome of Drosophila melanogaster" International Journal of Molecular Sciences 25, no. 12: 6559. https://doi.org/10.3390/ijms25126559
APA StyleBelcher, S., Flores-Iga, G., Natarajan, P., Crummett, G., Talavera-Caro, A., Gracia-Rodriguez, C., Lopez-Ortiz, C., Das, A., Adjeroh, D. A., Nimmakayala, P., Balagurusamy, N., & Reddy, U. K. (2024). Dietary Curcumin Intake and Its Effects on the Transcriptome and Metabolome of Drosophila melanogaster. International Journal of Molecular Sciences, 25(12), 6559. https://doi.org/10.3390/ijms25126559