
Novel Findings on CCR1 Receptor in CNS Disorders: A Pathogenic Marker Useful in Controlling Neuroimmune and Neuroinflammatory Mechanisms in Parkinson’s Disease
 , , and
, , and 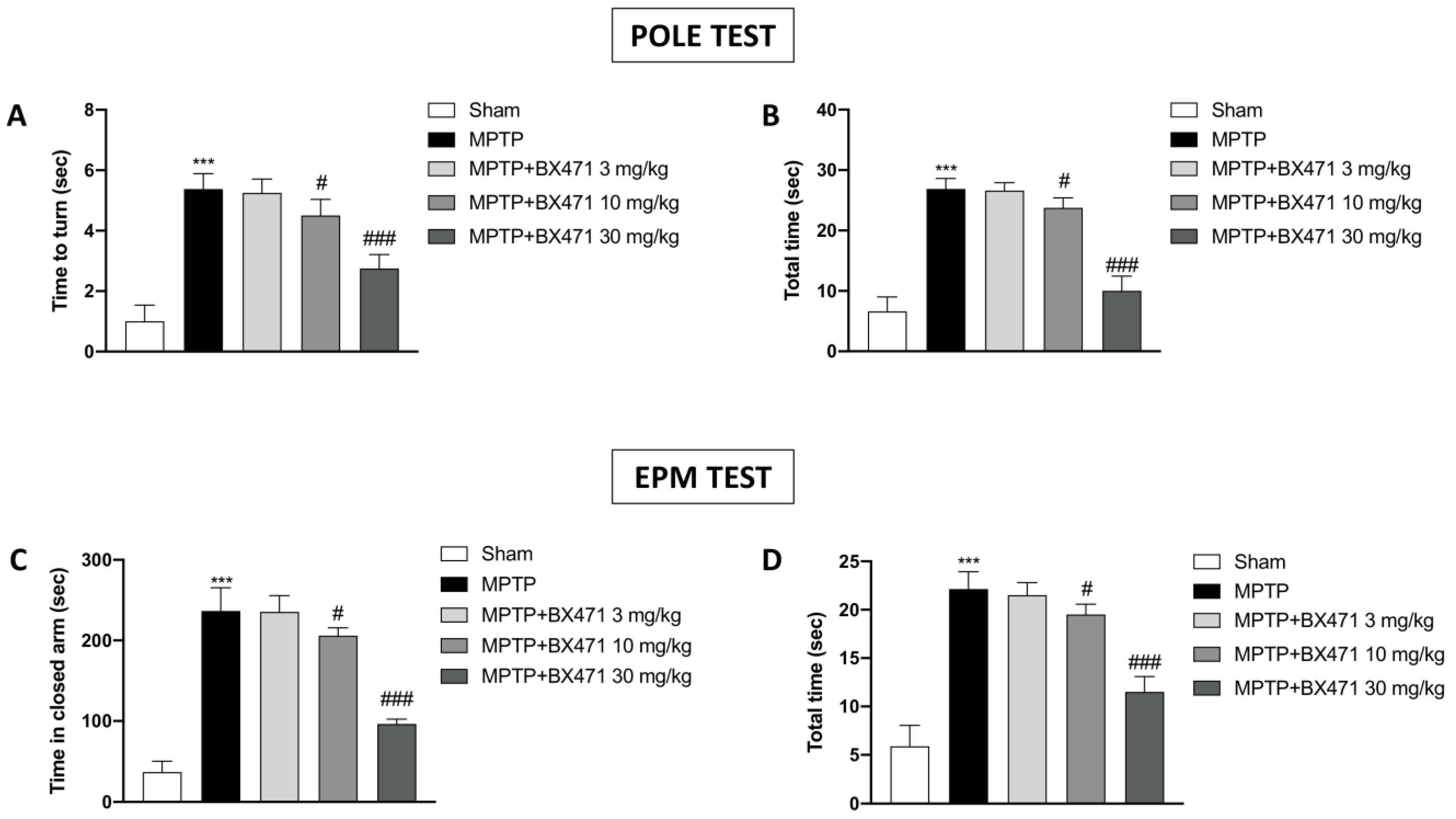{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Blocking of CCR1 Exerted by BX471 Reduced the Behavioral Deficits Caused by MPTP-Induced Nigrostriatal Degeneration
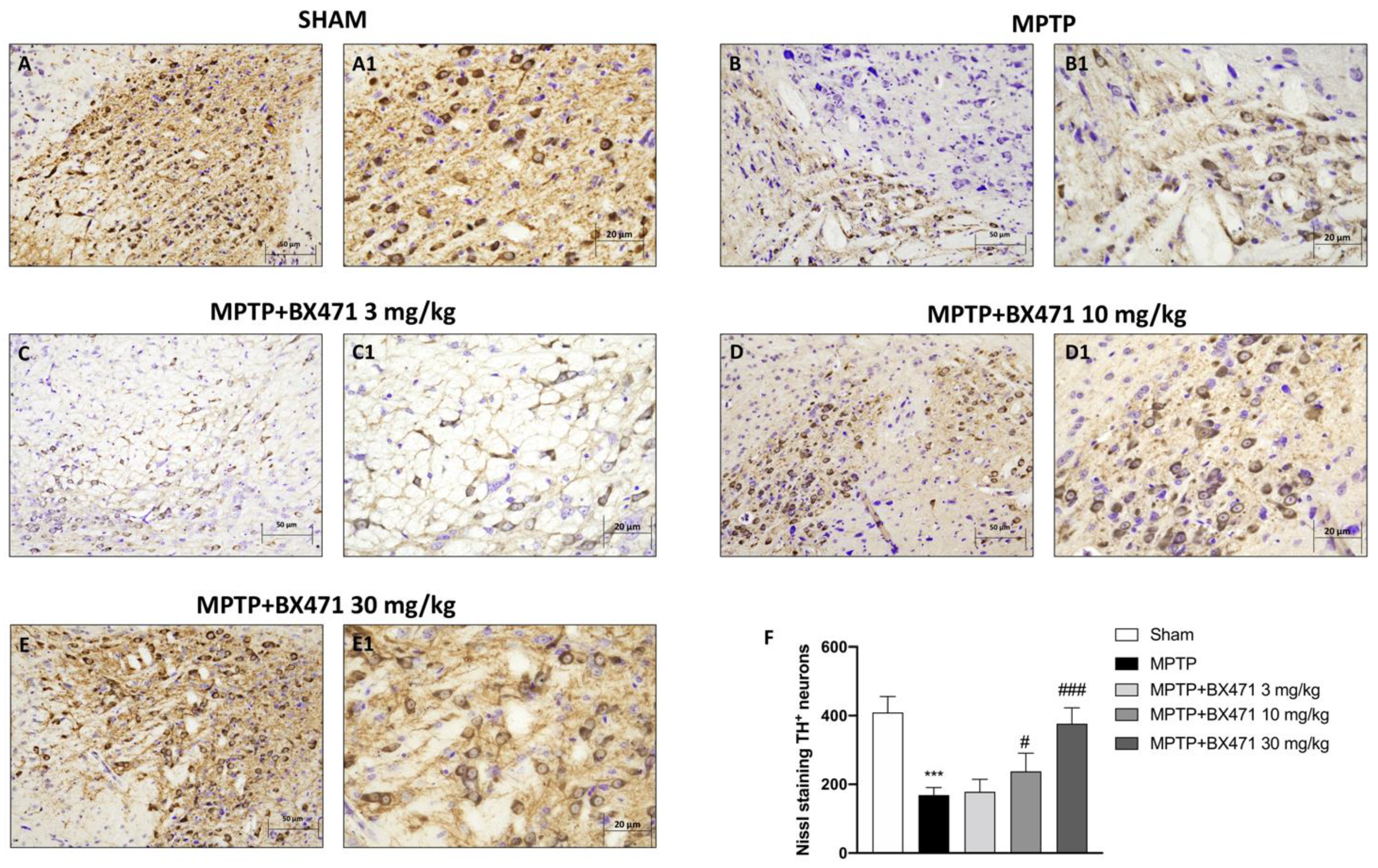
2.2. Antagonism of CCR1 by BX471 Inhibited the Loss of TH+ Neurons following MPTP-Induced Nigrostriatal Degeneration
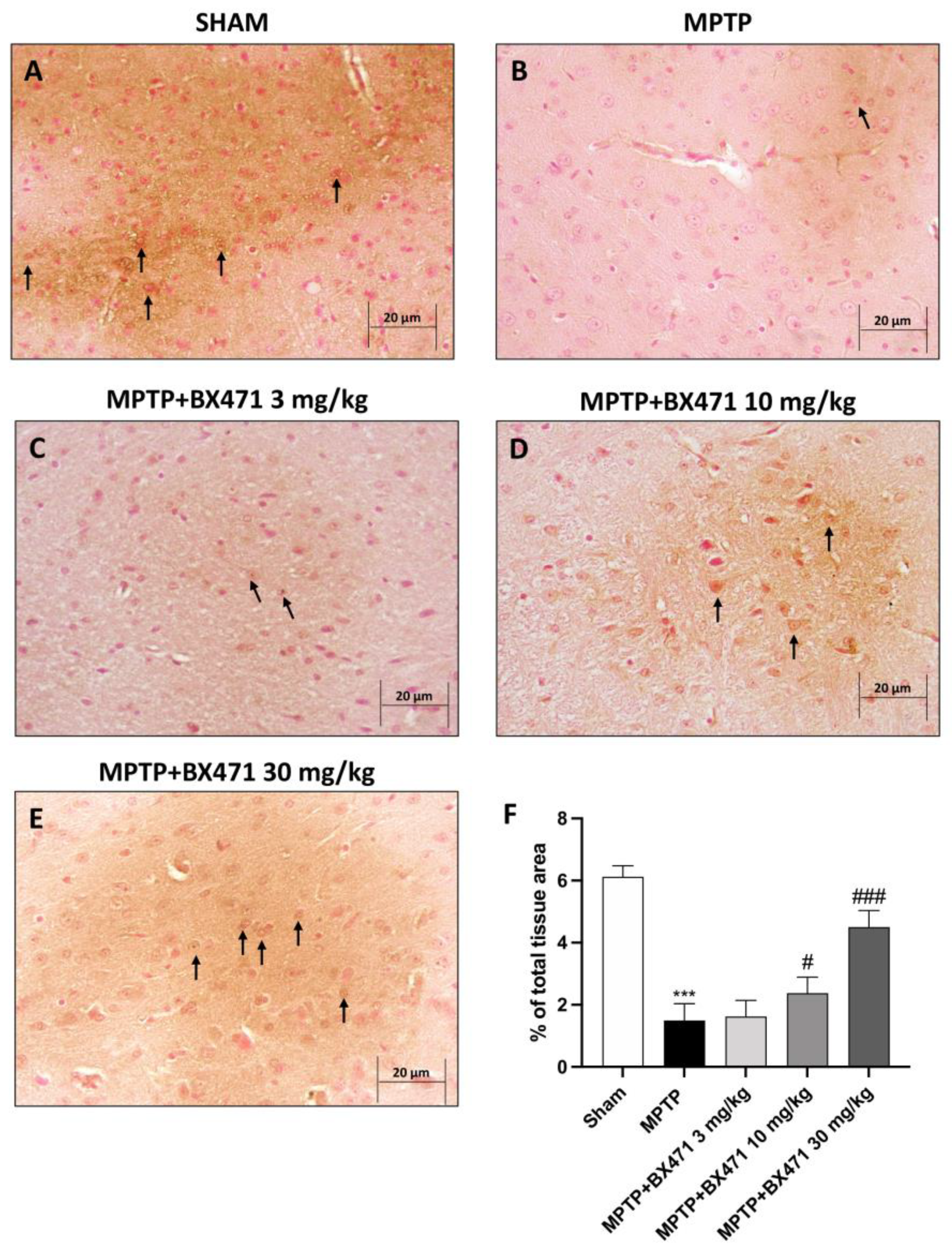
2.3. Downregulation of CCR1 by BX471 Protected against MPTP Neurotoxicity-Induced DAT Depletion
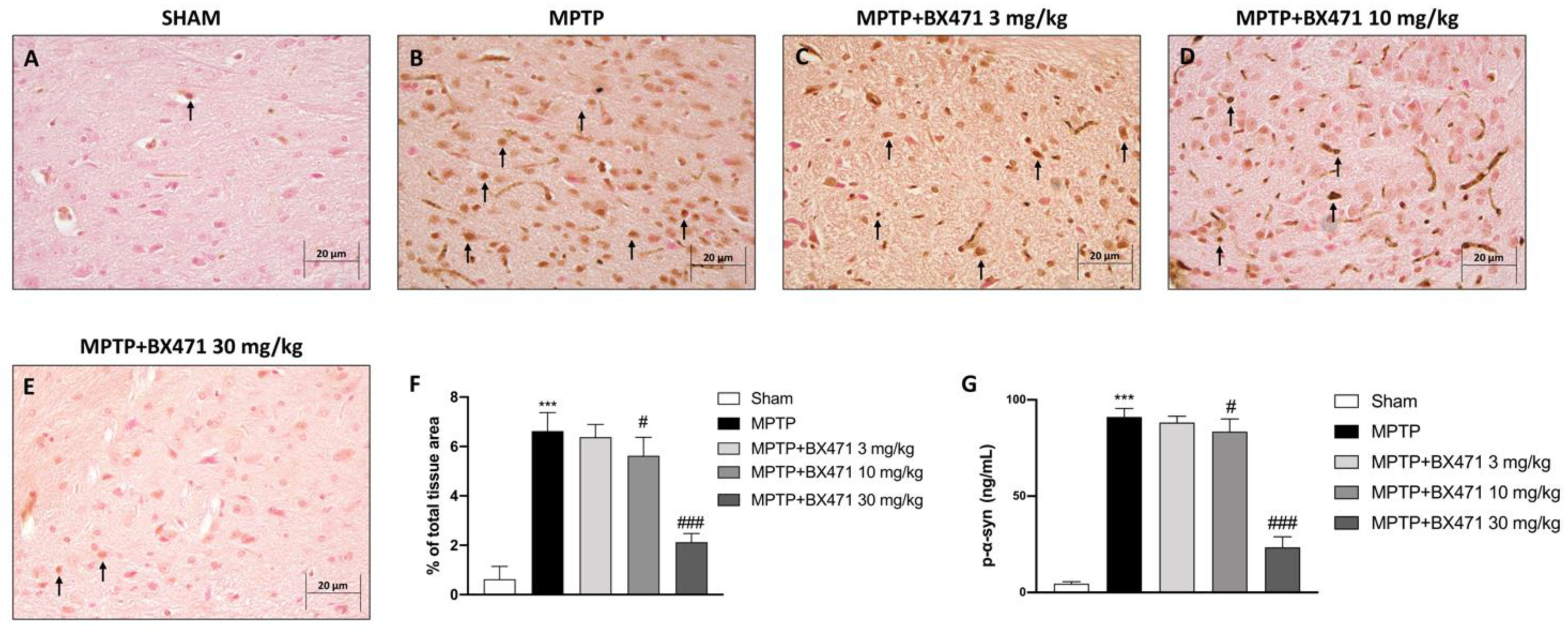
2.4. BX471 Counteracted α-Synuclein Accumulation Provoked by MPTP-Induced Nigrostriatal Degeneration
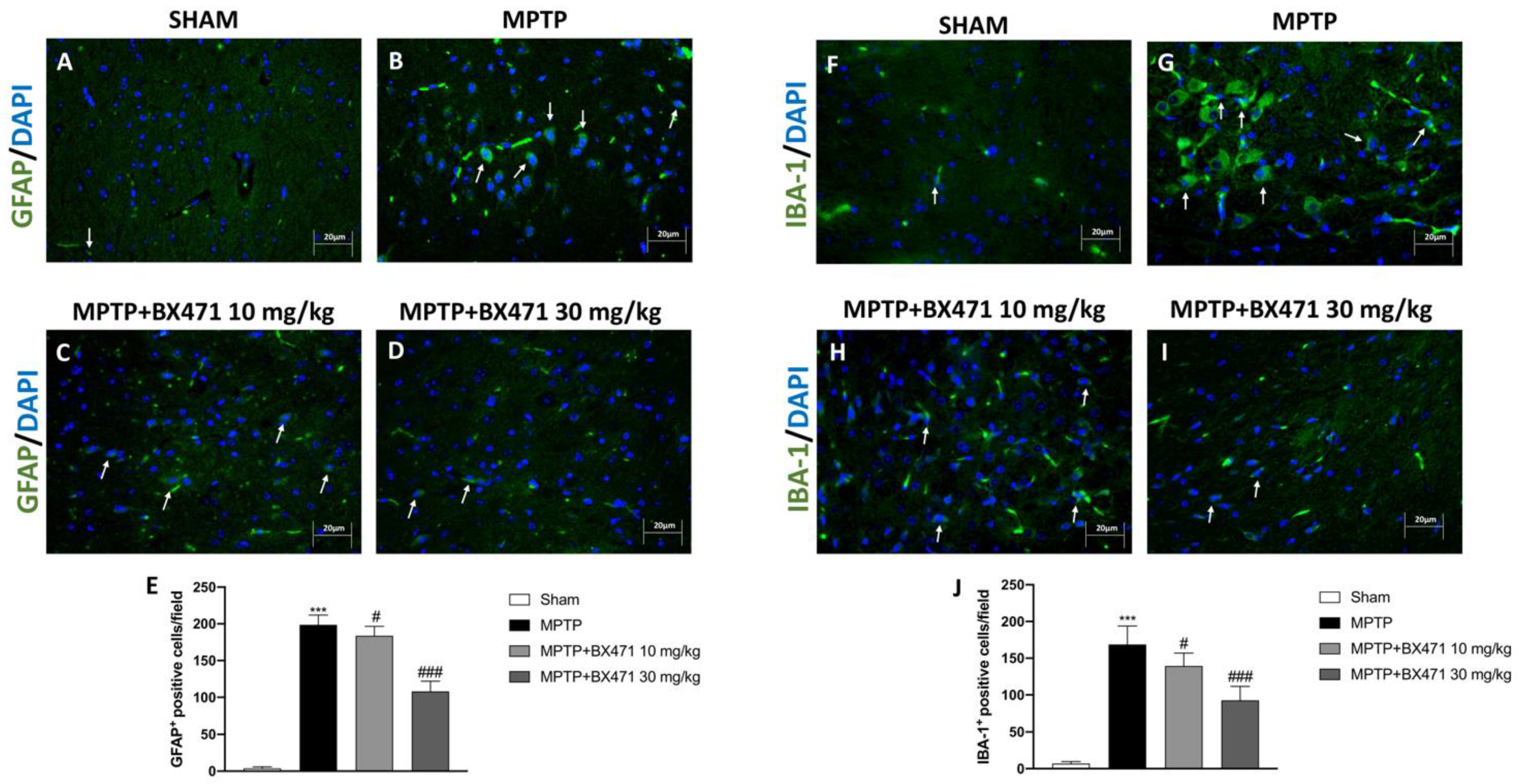
2.5. BX471 Decreased Reactive Astroglia and Microglia following MPTP-Induced Nigrostriatal Degeneration
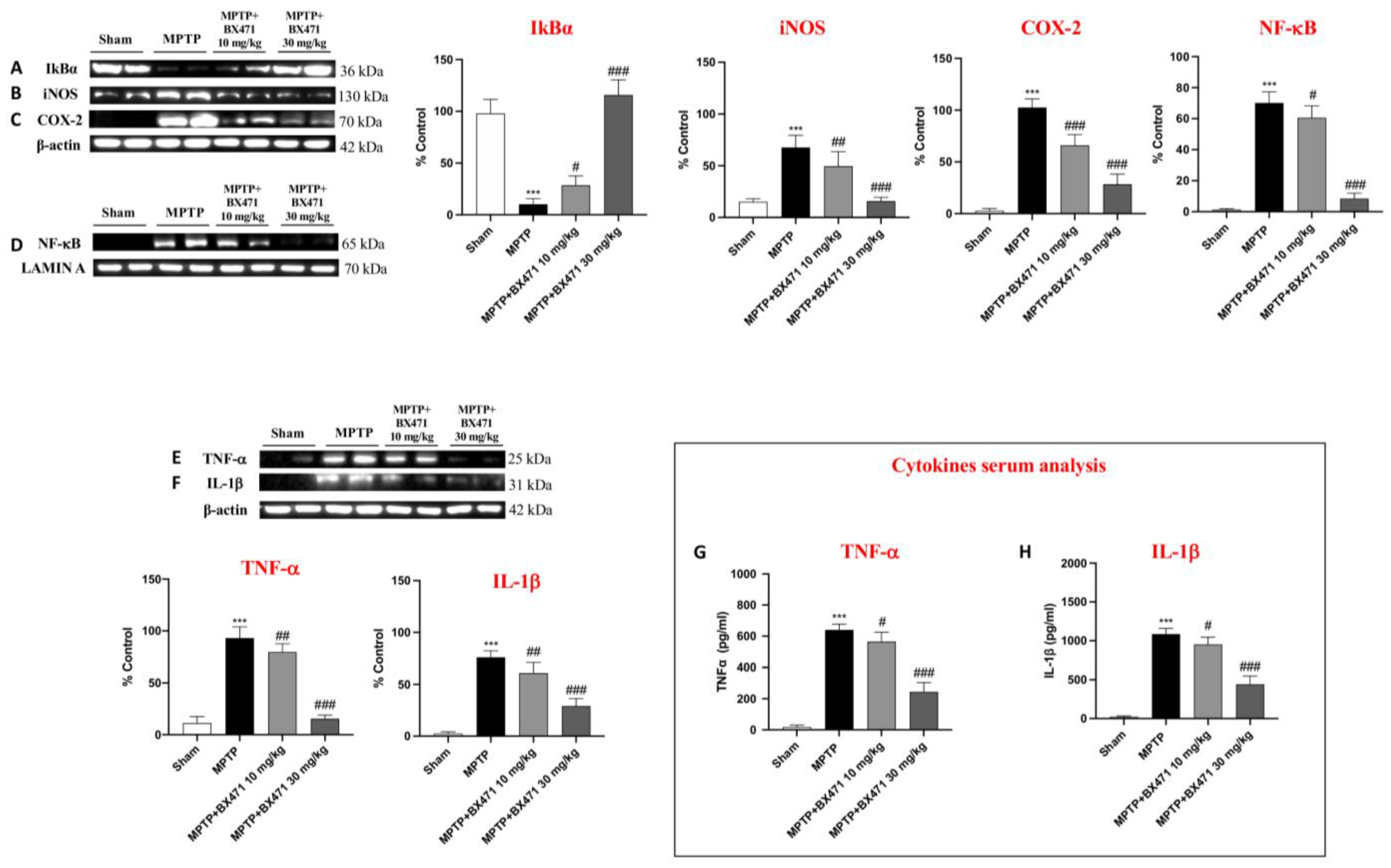
2.6. BX471 Restrained NF-kB Translocation and Reduced Pro-Inflammatory Mediators following MPTP-Induced Nigrostriatal Degeneration
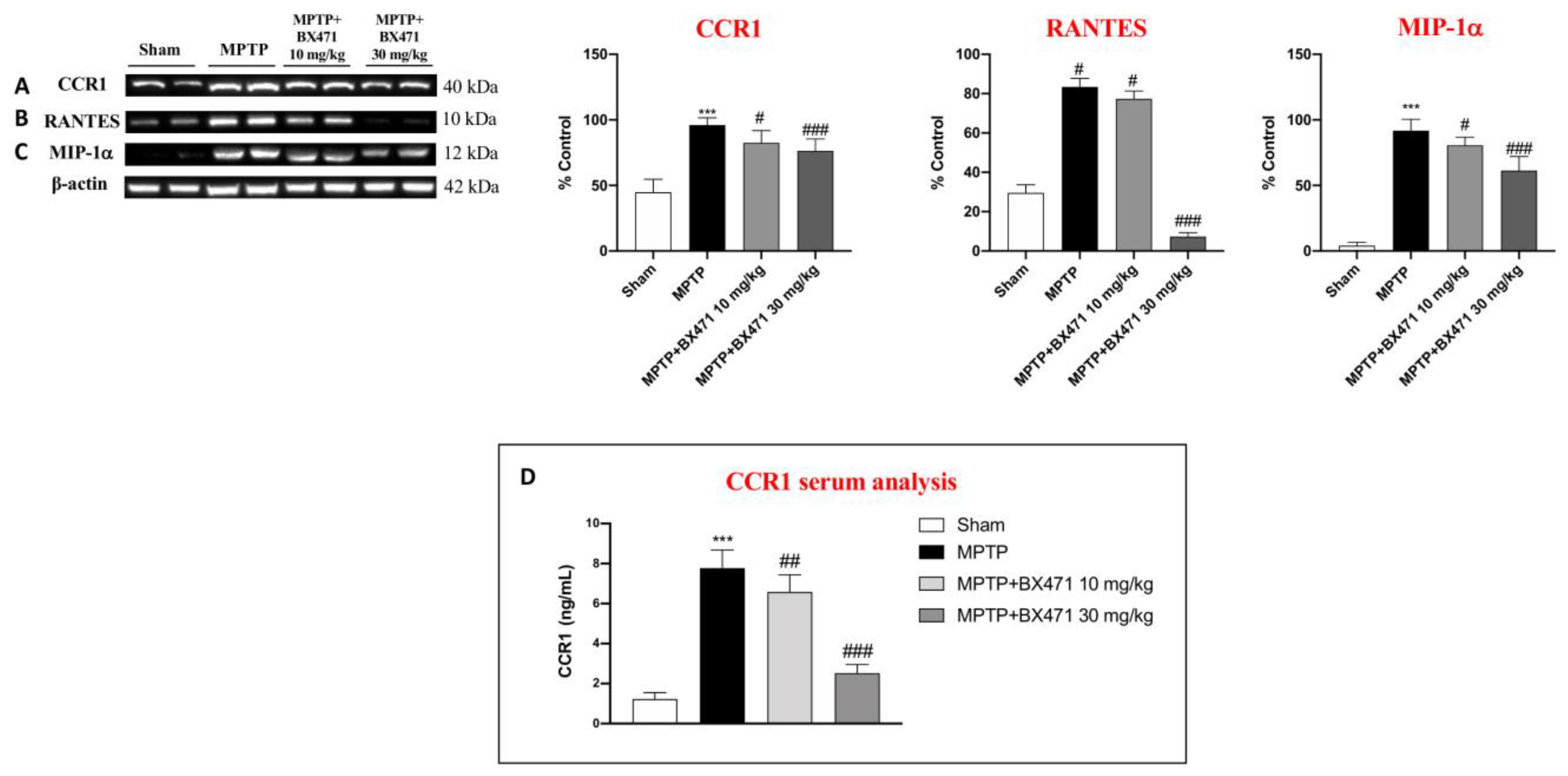
2.7. BX471 Antagonized the CCR1 Receptor and Moderated Immune-Inflammatory Mediators after MPTP-Induced Nigrostriatal Degeneration
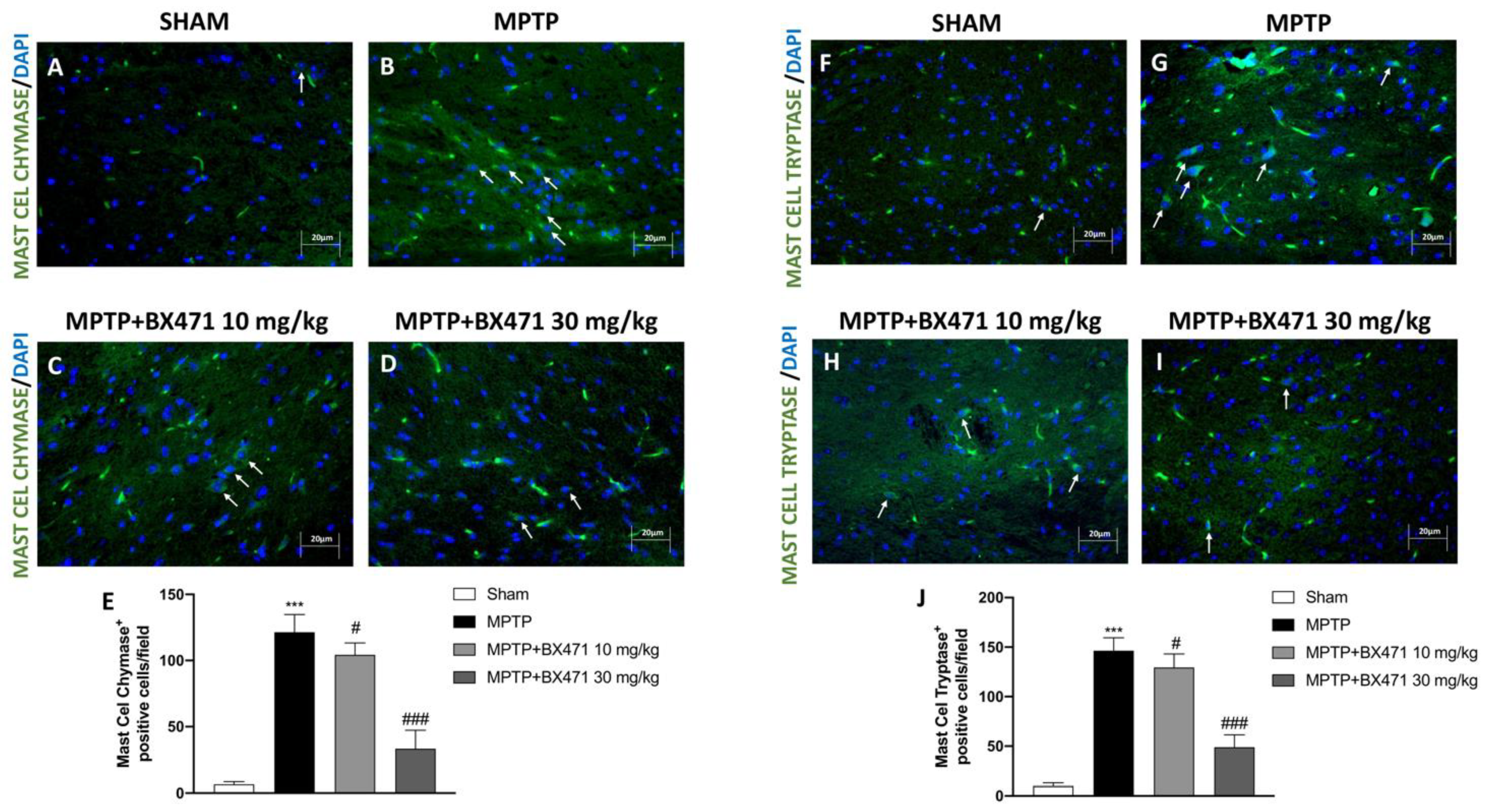
2.8. Block of CCR1 Receptor by BX471 Reduced Mast Cell Chymase and Tryptase Expression after MPTP-Induced Nigrostriatal Degeneration
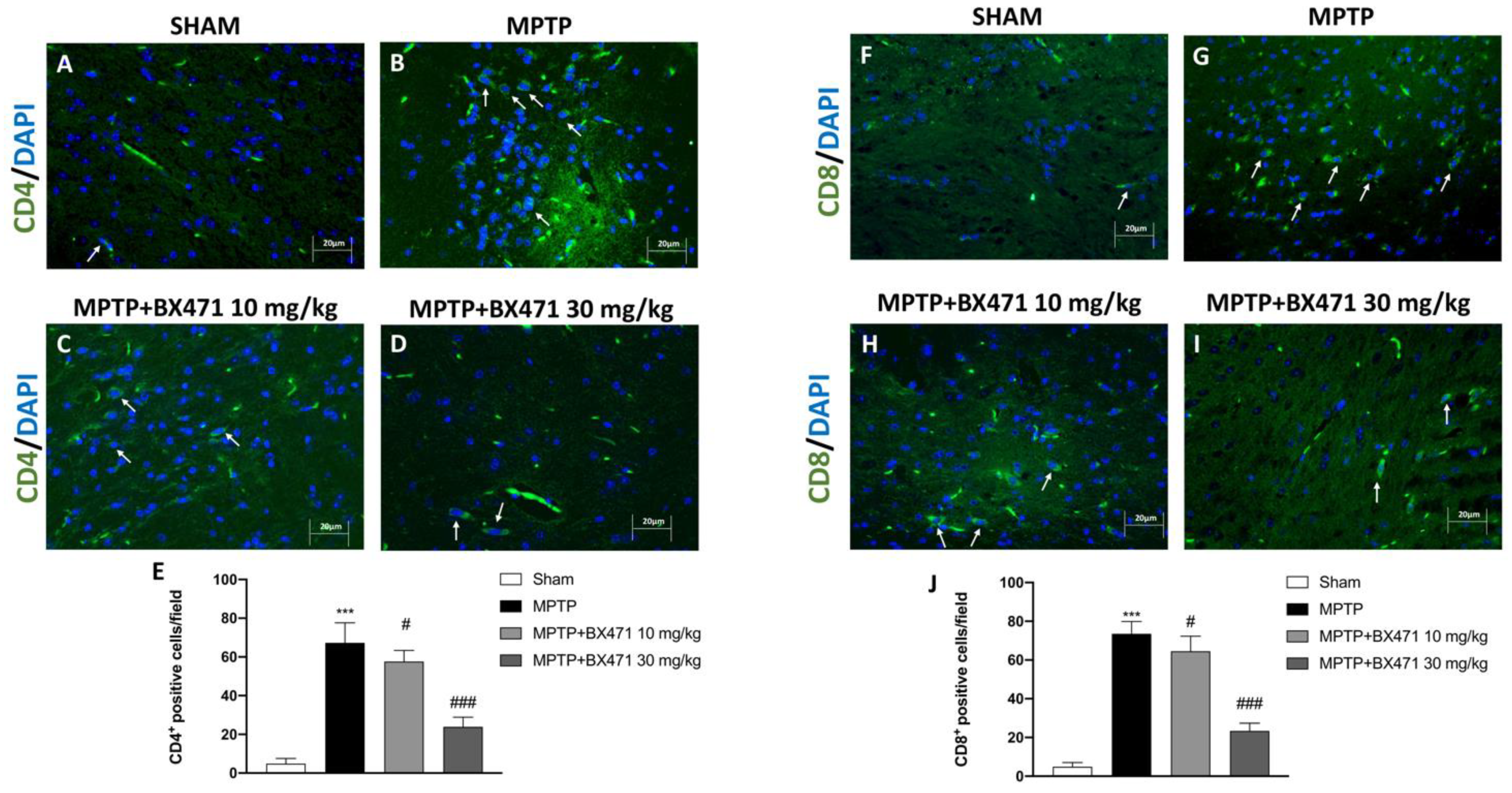
2.9. Block of CCR1 by BX471 Decreased T Lymphocyte Infiltration into the Midbrain Induced by MPTP Intoxication
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. MPTP-Induced Nigrostriatal Degeneration
4.4. Experimental Groups
4.5. Behavioral Testing
4.5.1. Pole Test
4.5.2. Elevated plus Maze (EPM)
4.6. Stereological Analysis
4.7. Immunohistochemical Localization of Dopamine Transporter (DAT) and α-Synuclein
4.8. Immunofluorescence of Glial Fibrillary Acidic Protein (GFAP), Ionized Calcium-Binding Adapter Molecule-1 (IBA-1), Mast Cell Chymase, Mast Cell Tryptase, CD-4, and CD-8
4.9. Western Blot Analysis
4.10. ELISA Kit
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Tanner, C.M.; Goldman, S.M. Epidemiology of Parkinson’s disease. Neurol. Clin. 1996, 14, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Aarsland, D.; Creese, B.; Politis, M.; Chaudhuri, K.R.; Ffytche, D.H.; Weintraub, D.; Ballard, C. Cognitive decline in Parkinson disease. Nat. Rev. Neurol. 2017, 13, 217–231. [Google Scholar] [CrossRef]
- Gonzalez-Latapi, P.; Bayram, E.; Litvan, I.; Marras, C. Cognitive Impairment in Parkinson’s Disease: Epidemiology, Clinical Profile, Protective and Risk Factors. Behav. Sci. 2021, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Luk, K.C.; Kehm, V.; Carroll, J.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M. Pathological alpha-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science 2012, 338, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Lüth, T.; König, I.R.; Grünewald, A.; Kasten, M.; Klein, C.; Hentati, F.; Farrer, M.; Trinh, J. Age at onset of LRRK2 p. Gly2019Ser is related to environmental and lifestyle factors. Mov. Disord. 2020, 35, 1854–1858. [Google Scholar] [CrossRef]
- Ye, H.; Robak, L.A.; Yu, M.; Cykowski, M.; Shulman, J.M. Genetics and Pathogenesis of Parkinson’s Syndrome. Annu. Rev. Pathol. 2023, 18, 95–121. [Google Scholar] [CrossRef]
- Di Rocco, M.; Di Fonzo, A.; Barbato, A.; Cappellini, M.D.; Carubbi, F.; Giona, F.; Giuffrida, G.; Linari, S.; Pession, A.; Quarta, A.; et al. Parkinson’s disease in Gaucher disease patients: What’s changing in the counseling and management of patients and their relatives? Orphanet J. Rare Dis. 2020, 15, 262. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s disease and its potential as therapeutic target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, M.; Zhao, A.; Kang, W.; Chen, Z.; Luo, N.; Zhou, L.; Zhu, X.; Lu, L.; Liu, J. CXCL12 is involved in alpha-synuclein-triggered neuroinflammation of Parkinson’s disease. J. Neuroinflamm. 2019, 16, 263. [Google Scholar] [CrossRef]
- Sawada, M.; Suzumura, A.; Marunouchi, T. Cytokine network in the central nervous system and its roles in growth and differentiation of glial and neuronal cells. Int. J. Dev. Neurosci. 1995, 13, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Koprich, J.B.; Reske-Nielsen, C.; Mithal, P.; Isacson, O. Neuroinflammation mediated by IL-1beta increases susceptibility of dopamine neurons to degeneration in an animal model of Parkinson’s disease. J. Neuroinflamm. 2008, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Weiss, F.; Labrador-Garrido, A.; Dzamko, N.; Halliday, G. Immune responses in the Parkinson’s disease brain. Neurobiol. Dis. 2022, 168, 105700. [Google Scholar] [CrossRef] [PubMed]
- Callahan, V.; Hawks, S.; Crawford, M.A.; Lehman, C.W.; Morrison, H.A.; Ivester, H.M.; Akhrymuk, I.; Boghdeh, N.; Flor, R.; Finkielstein, C.V.; et al. The Pro-Inflammatory Chemokines CXCL9, CXCL10 and CXCL11 Are Upregulated Following SARS-CoV-2 Infection in an AKT-Dependent Manner. Viruses 2021, 13, 1062. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, K.; Gao, J.L.; Horuk, R.; Wada, T.; Kaneko, S.; Murphy, P.M. Chemokine receptor CCR1 regulates inflammatory cell infiltration after renal ischemia-reperfusion injury. J. Immunol. 2008, 181, 8670–8676. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef] [PubMed]
- Glabinski, A.R.; Bielecki, B.; Kolodziejski, P.; Han, Y.; Selmaj, K.; Ransohoff, R.M. TNF-alpha microinjection upregulates chemokines and chemokine receptors in the central nervous system without inducing leukocyte infiltration. J. Interferon. Cytokine Res. 2003, 23, 457–466. [Google Scholar] [CrossRef]
- Liang, M.; Mallari, C.; Rosser, M.; Ng, H.P.; May, K.; Monahan, S.; Bauman, J.G.; Islam, I.; Ghannam, A.; Buckman, B.; et al. Identification and characterization of a potent, selective, and orally active antagonist of the CC chemokine receptor-1. J. Biol. Chem. 2000, 275, 19000–19008. [Google Scholar] [CrossRef]
- Anders, H.J.; Vielhauer, V.; Frink, M.; Linde, Y.; Cohen, C.D.; Blattner, S.M.; Kretzler, M.; Strutz, F.; Mack, M.; Grone, H.J.; et al. A chemokine receptor CCR-1 antagonist reduces renal fibrosis after unilateral ureter ligation. J. Clin. Investig. 2002, 109, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.J.; Ewing, J.L.; Schuh, J.M.; Ness, T.L.; Kunkel, S.L.; Aparici, M.; Miralpeix, M.; Hogaboam, C.M. Therapeutic targeting of CCR1 attenuates established chronic fungal asthma in mice. Br. J. Pharmacol. 2005, 145, 1160–1172. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Ju, L.; Shao, Z.; Grzanna, M.; Jia, L.; Liu, M. Therapeutic Effect of C-C Chemokine Receptor Type 1 (CCR1) Antagonist BX471 on Allergic Rhinitis. J. Inflamm. Res. 2020, 13, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zuo, G.; Sherchan, P.; Huang, L.; Ocak, U.; Xu, W.; Travis, Z.D.; Wang, W.; Zhang, J.H.; Tang, J. CCR1 Activation Promotes Neuroinflammation Through CCR1/TPR1/ERK1/2 Signaling Pathway After Intracerebral Hemorrhage in Mice. Neurotherapeutics 2020, 17, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Sunnemark, D.; Eltayeb, S.; Wallstrom, E.; Appelsved, L.; Malmberg, A.; Lassmann, H.; Ericsson-Dahlstrand, A.; Piehl, F.; Olsson, T. Differential expression of the chemokine receptors CX3CR1 and CCR1 by microglia and macrophages in myelin-oligodendrocyte-glycoprotein-induced experimental autoimmune encephalomyelitis. Brain Pathol. 2003, 13, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, J.; Zhou, Z.; Ransohoff, R.M. TGFbeta1 selectively up-regulates CCR1 expression in primary murine astrocytes. Glia 2000, 30, 1–10. [Google Scholar] [CrossRef]
- Halks-Miller, M.; Schroeder, M.L.; Haroutunian, V.; Moenning, U.; Rossi, M.; Achim, C.; Purohit, D.; Mahmoudi, M.; Horuk, R. CCR1 is an early and specific marker of Alzheimer’s disease. Ann. Neurol. 2003, 54, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; Wallings, R.L.; Houser, M.C.; Herrick, M.K.; Keating, C.E.; Joers, V. Inflammation and immune dysfunction in Parkinson disease. Nat. Rev. Immunol. 2022, 22, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Vaidehi, N.; Schlyer, S.; Trabanino, R.J.; Floriano, W.B.; Abrol, R.; Sharma, S.; Kochanny, M.; Koovakat, S.; Dunning, L.; Liang, M.; et al. Predictions of CCR1 chemokine receptor structure and BX 471 antagonist binding followed by experimental validation. J. Biol. Chem. 2006, 281, 27613–27620. [Google Scholar] [CrossRef]
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef]
- Mills, K.A.; Schneider, R.B.; Saint-Hilaire, M.; Ross, G.W.; Hauser, R.A.; Lang, A.E.; Halverson, M.J.; Oakes, D.; Eberly, S.; Litvan, I.; et al. Cognitive impairment in Parkinson’s disease: Associations between subjective and objective cognitive decline in a large longitudinal study. Park. Relat. Disord. 2020, 80, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, T.; Nakashima, A.; Ichinose, H.; Kobayashi, K. Human tyrosine hydroxylase in Parkinson’s disease and in related disorders. J. Neural. Transm. 2019, 126, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Salvatore, M.F.; Maiolo, S.A.; Bobrovskaya, L. Tyrosine hydroxylase as a sentinel for central and peripheral tissue responses in Parkinson’s progression: Evidence from clinical studies and neurotoxin models. Prog. Neurobiol. 2018, 165–167, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bourdy, R.; Sanchez-Catalan, M.J.; Kaufling, J.; Balcita-Pedicino, J.J.; Freund-Mercier, M.J.; Veinante, P.; Sesack, S.R.; Georges, F.; Barrot, M. Control of the nigrostriatal dopamine neuron activity and motor function by the tail of the ventral tegmental area. Neuropsychopharmacology 2014, 39, 2788–2798. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, O.V.; Amara, S.G. Dynamic regulation of the dopamine transporter. Eur. J. Pharmacol. 2003, 479, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; Galli, A. The dopamine transporter: A vigilant border control for psychostimulant action. Neurotransm. Transp. 2006; 175, 215–232. [Google Scholar] [CrossRef]
- Mannino, D.; Scuderi, S.A.; Casili, G.; Bova, V.; Cucinotta, L.; Lanza, M.; Filippone, A.; Esposito, E.; Paterniti, I. Neuroprotective effects of GSK-343 in an in vivo model of MPTP-induced nigrostriatal degeneration. J. Neuroinflamm. 2023, 20, 155. [Google Scholar] [CrossRef]
- Kumar, P.; Schilderink, N.; Subramaniam, V.; Huber, M. Membrane Binding of Parkinson’s Protein alpha-Synuclein: Effect of Phosphorylation at Positions 87 and 129 by the S to D Mutation Approach. Isr. J. Chem. 2017, 57, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Ali, J.; Khan, A.; Park, J.S.; Tahir, M.; Ahmad, W.; Choe, K.; Kim, M.O. Neuroprotective Effects of N-methyl-(2S, 4R)-trans-4-hydroxy-L-proline (NMP) against Amyloid-β-Induced Alzheimer’s Disease Mouse Model. Nutrients 2023, 15, 4986. [Google Scholar] [CrossRef]
- Yang, L.; Mao, K.; Yu, H.; Chen, J. Neuroinflammatory responses and Parkinson’disease: Pathogenic mechanisms and therapeutic targets. J. Neuroimmune Pharmacol. 2020, 15, 830–837. [Google Scholar] [CrossRef]
- Dolatshahi, M.; Ranjbar Hameghavandi, M.H.; Sabahi, M.; Rostamkhani, S. Nuclear factor-kappa B (NF-κB) in pathophysiology of Parkinson disease: Diverse patterns and mechanisms contributing to neurodegeneration. Eur. J. Neurosci. 2021, 54, 4101–4123. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Chao, Y.X.; West, A.; Chan, L.L.; Poewe, W.; Jankovic, J. Parkinson disease and the immune system-associations, mechanisms and therapeutics. Nat. Rev. Neurol. 2020, 16, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Qiao, L.; Peng, X.; Cui, Z.; Yin, Y.; Liao, H.; Jiang, M.; Li, L. The chemokine receptor CCR1 is identified in mast cell-derived exosomes. Am. J. Transl. Res. 2018, 10, 352–367. [Google Scholar] [PubMed]
- Schaller, T.H.; Batich, K.A.; Suryadevara, C.M.; Desai, R.; Sampson, J.H. Chemokines as adjuvants for immunotherapy: Implications for immune activation with CCL3. Expert Rev. Clin. Immunol. 2017, 13, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Potashkin, J.A.; Blume, S.R.; Runkle, N.K. Limitations of animal models of Parkinson’s disease. Park. Dis. 2010, 2011, 658083. [Google Scholar] [CrossRef] [PubMed]
- Zipp, F.; Hartung, H.P.; Hillert, J.; Schimrigk, S.; Trebst, C.; Stangel, M.; Infante-Duarte, C.; Jakobs, P.; Wolf, C.; Sandbrink, R.; et al. Blockade of chemokine signaling in patients with multiple sclerosis. Neurology 2006, 67, 1880–1883. [Google Scholar] [CrossRef] [PubMed]
- Ardizzone, A.; Bova, V.; Casili, G.; Filippone, A.; Campolo, M.; Lanza, M.; Esposito, E.; Paterniti, I. SUN11602, a bFGF mimetic, modulated neuroinflammation, apoptosis and calcium-binding proteins in an in vivo model of MPTP-induced nigrostriatal degeneration. J. Neuroinflamm. 2022, 19, 107. [Google Scholar] [CrossRef]
- Campolo, M.; Casili, G.; Lanza, M.; Filippone, A.; Cordaro, M.; Ardizzone, A.; Scuderi, S.A.; Cuzzocrea, S.; Esposito, E.; Paterniti, I. The inhibition of mammalian target of rapamycin (mTOR) in improving inflammatory response after traumatic brain injury. J. Cell. Mol. Med. 2021, 25, 7855–7866. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Lanza, M.; Paterniti, I.; Filippone, A.; Ardizzone, A.; Casili, G.; Scuderi, S.A.; Puglisi, C.; Mare, M.; Memeo, L.; et al. PEA-OXA Mitigates Oxaliplatin-Induced Painful Neuropathy through NF-kappaB/Nrf-2 Axis. Int. J. Mol. Sci. 2021, 22, 3927. [Google Scholar] [CrossRef]
- Ardizzone, A.; Filippone, A.; Mannino, D.; Scuderi, S.A.; Casili, G.; Lanza, M.; Cucinotta, L.; Campolo, M.; Esposito, E. Ulva pertusa, a Marine Green Alga, Attenuates DNBS-Induced Colitis Damage via NF-kappaB/Nrf2/SIRT1 Signaling Pathways. J. Clin. Med. 2022, 11, 4301. [Google Scholar] [CrossRef]
- Ardizzone, A.; Fusco, R.; Casili, G.; Lanza, M.; Impellizzeri, D.; Esposito, E.; Cuzzocrea, S. Effect of Ultra-Micronized-Palmitoylethanolamide and Acetyl-l-Carnitine on Experimental Model of Inflammatory Pain. Int. J. Mol. Sci. 2021, 22, 1967. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Ardizzone, A.; Campolo, M.; Conoci, S.; Esposito, E.; Paterniti, I. Beneficial Effect of Tempol, a Membrane-Permeable Radical Scavenger, on Inflammation and Osteoarthritis in In Vitro Models. Biomolecules 2021, 11, 352. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Repici, A.; Capra, A.P.; Hasan, A.; Bulzomì, M.; Campolo, M.; Paterniti, I.; Esposito, E.; Ardizzone, A. Novel Findings on CCR1 Receptor in CNS Disorders: A Pathogenic Marker Useful in Controlling Neuroimmune and Neuroinflammatory Mechanisms in Parkinson’s Disease. Int. J. Mol. Sci. 2024, 25, 4337. https://doi.org/10.3390/ijms25084337
Repici A, Capra AP, Hasan A, Bulzomì M, Campolo M, Paterniti I, Esposito E, Ardizzone A. Novel Findings on CCR1 Receptor in CNS Disorders: A Pathogenic Marker Useful in Controlling Neuroimmune and Neuroinflammatory Mechanisms in Parkinson’s Disease. International Journal of Molecular Sciences. 2024; 25(8):4337. https://doi.org/10.3390/ijms25084337
Chicago/Turabian StyleRepici, Alberto, Anna Paola Capra, Ahmed Hasan, Maria Bulzomì, Michela Campolo, Irene Paterniti, Emanuela Esposito, and Alessio Ardizzone. 2024. "Novel Findings on CCR1 Receptor in CNS Disorders: A Pathogenic Marker Useful in Controlling Neuroimmune and Neuroinflammatory Mechanisms in Parkinson’s Disease" International Journal of Molecular Sciences 25, no. 8: 4337. https://doi.org/10.3390/ijms25084337