


Habitat and Features of Development of Plankton Communities in Salt Lakes (South-Eastern Transbaikalia, Russia)


Abstract
:1. Introduction
- -
- To determine the main physical and chemical parameters of lakes’ various chemical types;
- -
- To assess the plankton communities in studied lakes;
- -
- To identify dominant species in the different types of lakes;
- -
- To analyze the change in abundance and taxonomic diversity.
2. Materials and Methods
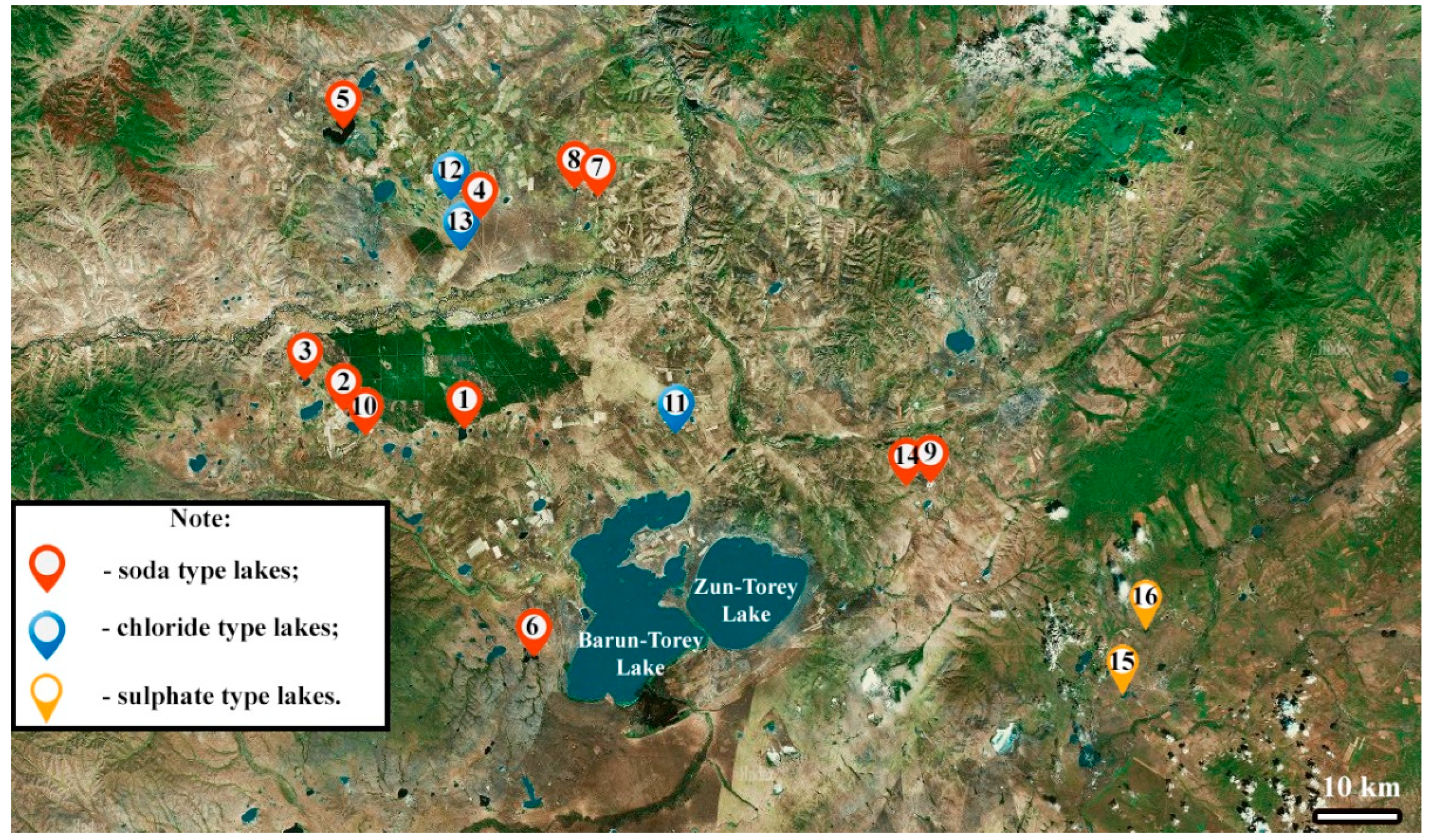
2.1. Study Area
2.2. Sampling Strategies
2.3. Phytoplankton
2.4. Zooplankton
2.5. Statistical Analysis
3. Results
3.1. Environmental Parameters
3.2. Principal Correspondence Analysis
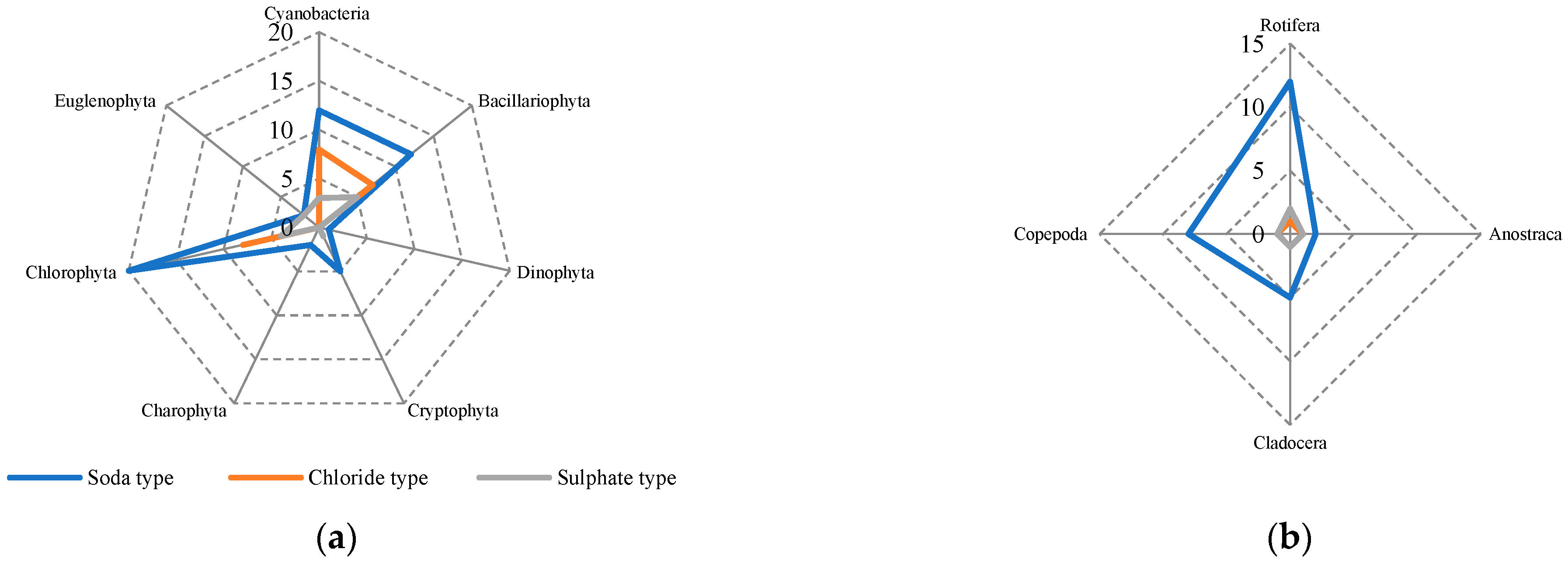
3.3. Diversity of Phyto- and Zooplankton
3.4. Dominant Complex of Phyto- and Zooplankton
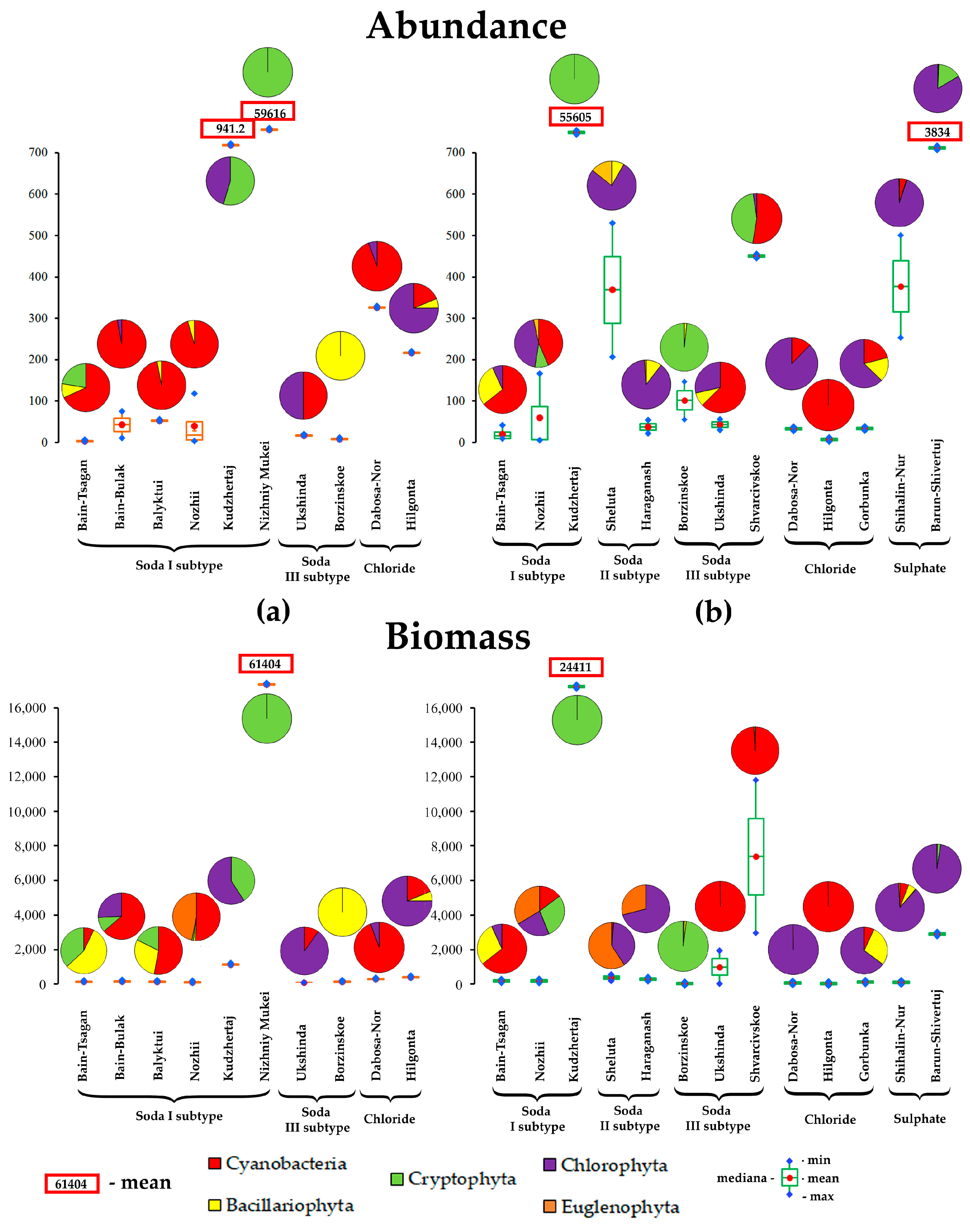
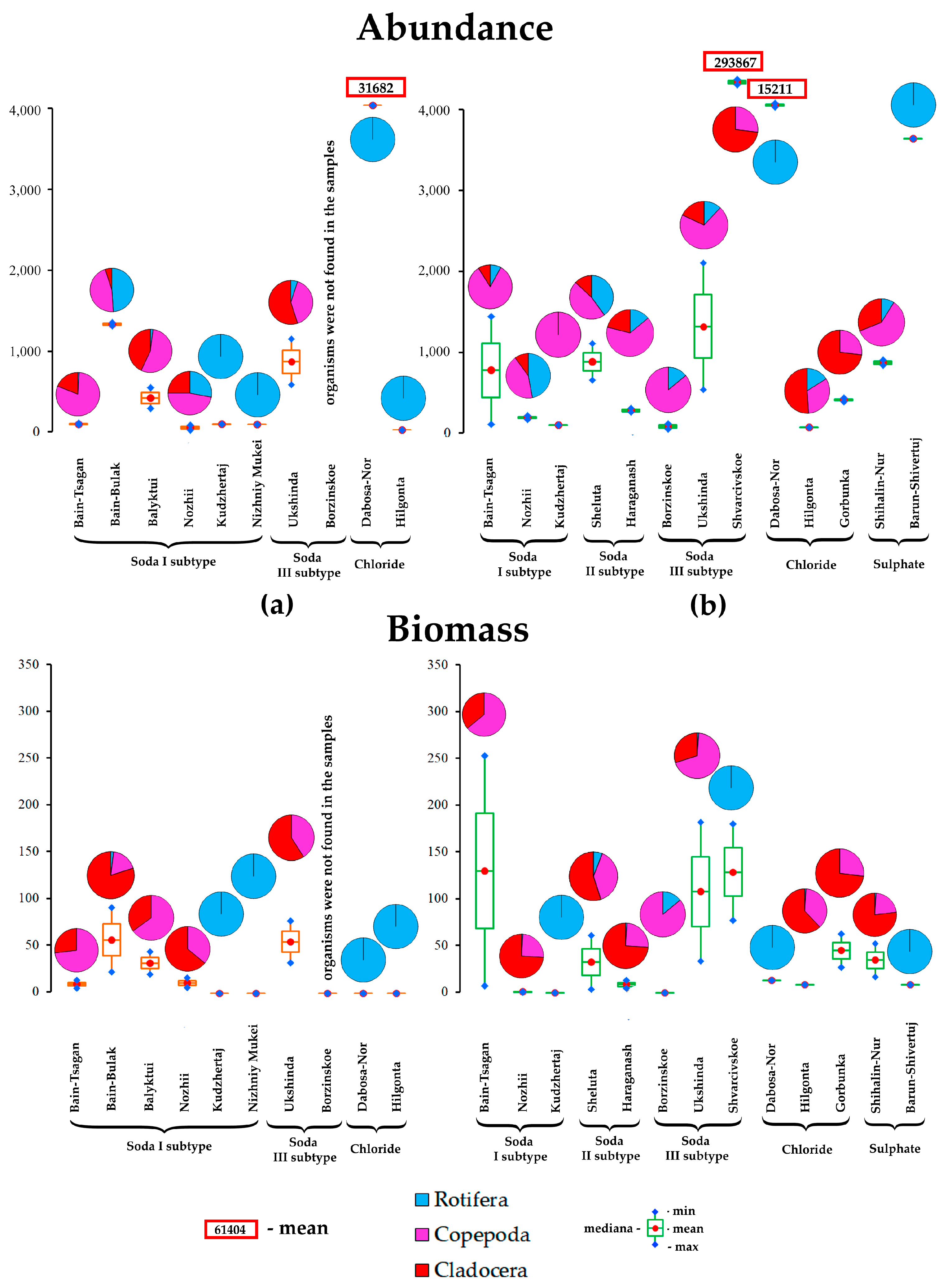
3.5. Abundance and Biomass of Phyto- and Zooplankton
3.6. Relationships between Environmental Factors and Quantitative Indicators of the Plankton
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, W.D. The largest, highest and lowest lakes of the world: Saline lakes. Verh. Int. Ver. Für Limnol. 1996, 26, 61–79. [Google Scholar] [CrossRef]
- Williams, W.D. Environmental threats to salts lakes and the likely status of inland saline ecosystems in 2025. Environ. Conserv. 2002, 29, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Zavarzin, G.A. Epicontinental alkaline water bodies as relict biotopes for the development of terrestrial biota. Mikrobiologiâ 1993, 62, 789–800. [Google Scholar]
- Brucet, S.; Boix, D.; Gascón, S.; Sala, J.; Quintana, X.D.; Badosa, A.; Søndergaard, M.; Lauridsen, T.L.; Jeppesen, E. Species Richness Crustac. Zooplankton Trophic Struct. Brac. Lagoons Contrasting Clim. Zones: North Temp. Den. Mediterr. Catalonia (Spain). Ecography 2009, 32, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Felföldi, T. Microbial communities of soda lakes and pans in the Carpathian Basin: A review. Biol. Futura 2020, 71, 393–404. [Google Scholar] [CrossRef]
- Shadrin, N.V.; Anufrieva, E.V. Dependence of halotolerance of Arctodiaptomus salinus (Calanoida, Copepoda) on exoosmolytes: New data and hypothesis. J. Mediterr. Ecol. 2013, 12, 21–26. [Google Scholar]
- Raini, J.A. The eastern Africa flamingo Lakes: Building partnerships for sustainable resource management. In Proceedings of the 11th World Lakes Conference, Nairobi, Kenya, 31 October–4 November 2005; Volume 2, pp. 276–281. [Google Scholar]
- Marden, B.; Brown, P.; Bosteels, T. Great Salt Lake Artemia: Ecosystem Functions and Services with a Global Reach. In Great Salt Lake Biology; Baxter, B., Butler, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 175–237. [Google Scholar]
- Wurtsbaugh, W.A.; Gliwicz, Z.M. Limnological control of brine shrimp population dynamics and cyst production in the Great Salt Lake, Utah. In Saline Lakes. Developments in Hydrobiology; Melack, J.M., Jellison, R., Herbst, D.B., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 162, pp. 119–132. [Google Scholar]
- Valyashko, M.G. Regularities in the Formation of Salt Deposits; Moscow State University: Moscow, Russia, 1962; p. 398. [Google Scholar]
- Zheng, M.; Liu, X. Hydrochemistry of Salt Lakes of the Qinghai-Tibet Plateau, China. Aquat. Geochem. 2009, 15, 293–320. [Google Scholar] [CrossRef]
- Borzenko, S.V.; Shvartsev, S.L. Chemical composition of salt lakes in East Transbaikalia (Russia). Appl. Geochem. 2019, 103, 72–84. [Google Scholar] [CrossRef]
- Borzenko, S.V. The main formation processes for different types of salt lakes: Evidence from isotopic composition with case studies of lakes in Transbaikalia, Russia. Sci. Total Environ. 2021, 782, 146782. [Google Scholar] [CrossRef]
- Chang, W.Y.B. Large lakes of China. J. Great Lakes Res. 1987, 13, 235–249. [Google Scholar] [CrossRef]
- Last, W.M. Continental brines and evaporites of the northern Great Plains of Canada. Sediment. Geol. 1989, 64, 207–221. [Google Scholar] [CrossRef]
- Last, W.M. Chemical composition of saline and subsaline lakes of the northern Great Plains, western Canada. Int. J. Salt Lake Res. 1992, 1, 47–76. [Google Scholar] [CrossRef]
- Zsuga, K.; Inelova, Z.; Boros, E. Zooplankton Community Structure in Shallow Saline Steppe Inland Waters. Water 2021, 13, 1164. [Google Scholar] [CrossRef]
- Last, W.M.; Ginn, F.M. Saline systems of the Great Plains of western Canada: An overview of the limnogeology and paleolimnology. Aquat. Biosyst. 2005, 1, 1–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frish, V.A. Torey “experiment”. Nature 1972, 2, 60–66. [Google Scholar]
- Zamana, L.V.; Borzenko, S.V. Hydrochemical regime of saline lakes in the southeastern Transbaikalia. Geogr. Nat. Res. 2010, 31, 370–376. [Google Scholar] [CrossRef]
- Kuklin, A.P.; Tsybekmitova, G.T.; Gorlacheva, E.P. State of lake ecosystems in Onon-Torei 601 plain in 1983–2011 (Eastern Transbaikalia). Arid. Ecosyst. 2013, 3, 122–130. [Google Scholar] [CrossRef]
- Zamana, L.V.; Vakhnina, I.L. Hydrochemistry of saline lakes of the southeastern Transbaikalia under climate aridization in the beginning of 20th century. Int. J. Appl. Fundam. Res. 2014, 11, 608–612. [Google Scholar]
- Tsibekmitova, G.T.; Matveeva, M.O. Content of nutrients in the lakes of the Onon-Torey depression during the period of climatic fluctuations. Water Sect. Russ. 2019, 3, 94–108. [Google Scholar]
- Oremland, R.S. Arsenic, prokaryotes, and closed basin soda lakes of the Western USA. AGU Fall Meet. Abstr. 2006, 2006, H51H-02. [Google Scholar]
- Tashlykova, N.A.; Afonina, E.Y. The current species composition and ecological685 geographical characteristics of plankton communities in the littoral zone of some lakes of the 686 Uldza-Torey basin (Trans-Baikal Territory). Acta Biol. Sib. 2019, 5, 102–110. [Google Scholar]
- Itigilova, M.T.; Dulmaa, A.; Afonina, E.Y. Zooplankton of lakes of the Uldza and Kerulen river valleys of Northeastern Mongolia. Inland Water Biol. 2014, 7, 249–258. [Google Scholar] [CrossRef]
- Gorlacheva, E.P.; Tsybekmitova, G.T.; Afonin, A.V.; Tashlikova, N.A.; Afonina, E.Y.; Kuklin, A.P.; Saltanova, N.V. Lake-margin ecosystems of saline lakes of the Borzya group (Zabaikalsky Krai, Russia) during the initial filling phase. Chin. J. Ocean Limn. 2014, 32, 871–878. [Google Scholar] [CrossRef]
- Bazarova, B.B.; Tashlykova, N.A.; Afonina, E.Y.; Kuklin, A.P.; Matafonov, P.V.; Tsybekmitova, G.T.; Gorlacheva, E.P.; Itigilova, M.T.; Afonin, A.V.; Butenko, M.N. Long-term fluctuations of the aquatic ecosystems in the Onon-Torey plain (Russia). Acta Ecol. Sin. 2019, 39, 157–165. [Google Scholar] [CrossRef]
- Afonina, E.Y.; Tashlykova, N.A. Planktonic communities in the Torey Lakes (Zabaikalsky Krai) in a low water year. J. Sib. Fed. Univ. Biol. 2018, 11, 306–320. [Google Scholar]
- Bazhenova, O.I. Modern dynamics of lacustrine-fluvial systems of the Onon-Torey high plain (Southern Transbaikalia). Bull. TSU 2013, 371, 171–177. [Google Scholar]
- Obyazov, V.A. Change of climate and hydrological regime of the rivers and lakes in Dahurian region. In Problems of Adaptation and Climate Change in the River Basins of Dahuria: Ecological and Hydroeconomic Aspects; Kirilyuk, O., Ed.; Ekspress-izdatel’stvo: Chita, Russia, 2012; pp. 24–45. [Google Scholar]
- Abiduyeva, E.Y.; Kozyreva, L.P.; Syrenzhapova, A.S.; Namsarayev, B.B. Seasonal changes in physicochemical environmental conditions of salt lake dabasu-nur (Southeastern Trans Baikalia). Geogr. Nat. Resour. 2008, 2, 177–179. [Google Scholar]
- Tsyrenova, D.D.; Bryanskaya, A.V.; Khakhinov, V.V.; Zhavzan, C. Hydrochemical studies of brackish and saline lakes in Southern Transbaikalia. Bull. Buryat State Univ. Chem. Phys. 2009, 3, 17–19. [Google Scholar]
- Suvorova, V.A.; Abidueva, E.Y. The environmental conditions and distribution of alkalophilic bacteria-destructor in mineral lakes in Transbaikalia and Mongolian Plateau. Bull. Buryat State Univ. Chem. Phys. 2012, 3, 67–69. [Google Scholar]
- Borzenko, S.V. Basic conditions for the formation of the chemical composition of the waters of saline and brackish lakes in Eastern Transbaikalia. Geochemistry 2020, 659, 1212–1230. [Google Scholar]
- Borzenko, S.V.; Zamana, L.V.; Posokhov, V.F. Isotope composition, nature and main mechanisms of formation of salt lakes in Transbaikalia. Geol. Geophys. 2022, 63, 851–874. [Google Scholar]
- Sadchikov, A.P. Methods of Studying Freshwater Phytoplankton, 1st ed.; Publishing House “University and School”: Moscow, Russia, 2003; pp. 67–79. [Google Scholar]
- Kiselev, I.A. Plankton of the Sea and Continental Reservoirs, 1st ed.; Nauka: Leningrad, Russia, 1969; 658p. [Google Scholar]
- Korneva, L.G. Phytoplankton of the Reservoirs of the Volga Basin; Kopylov, A.I., Ed.; Kostroma Printing House: Kostroma, Russia, 2015; 284p. [Google Scholar]
- Balushkina, E.B.; Vinberg, G.G. The relationship between body weight and length in planktonic animals. In General Principles of Study of Aquatic Ecosystems, 1st ed.; Vinberg, G., Ed.; Nauka: Leningrad, Russia, 1979; pp. 169–172. [Google Scholar]
- Ruttner-Kolisko, A. Suggestions for biomass calculation of plankton rotifers. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1977, 8, 71–76. [Google Scholar]
- Fedorov, V.D.; Gilmanov, T.G. Ecology; Publishing House of Moscow State University: Moscow, Russia, 1980; 464p. [Google Scholar]
- Dalgaard, P. Introductory Statistics with R, 1st ed.; Springer: New York, NY, USA, 2008; pp. 1–162. [Google Scholar]
- Javor, B.J. Planktonic standing crop and nutrients in a saltern ecosystem. Limnol. Oceanogr. 1986, 28, 153–159. [Google Scholar] [CrossRef]
- Edgerton, M.E.; Brimblecombe, P. Thermodynamics of halobacterial environments. Can. J. Microbiol. 1981, 27, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Baas-Becking, L.G.M. Salt effects on swarmers of Dunaliella viridis Teod. J. Gen. Physiol. 1931, 14, 765–779. [Google Scholar] [CrossRef] [Green Version]
- Bayly, I.E.A. The occurrence of calanoid copepods in athalassic saline waters in relation to salinity and anionic proportions. Verh. Int. Ver. Limnol. 1969, 17, 449–455. [Google Scholar] [CrossRef]
- Vesnina, L.; Bezmaternykh, D.; Moruzi, I.; Pishenko, E. Seasonal and interannual dynamics of zooplankton from Lake Kulundinskoye in 2017–2020. In XV International Scientific Conference “INTERAGROMASH 2022” Global Precision Ag Innovation 2022; Springer International Publishing: Cham, Switzerland, 2023; Volume 2. [Google Scholar]
- Timms, B.V. Drivers restricting biodiversity in Australian saline lakes: A review. Mar. Freshw. Res. 2020, 72, 462–468. [Google Scholar] [CrossRef]
- Csitári, B.; Bedics, A.; Felföldi, T.; Boros, E.; Nagy, H.; Máthé, I.; Székely, A.J. Anion-type modulates the effect of salt stress on saline lake bacteria. Extremophiles 2022, 26, 2–14. [Google Scholar] [CrossRef]
- Zhao, W.; Zheng, M.P.; Xu, X.Z.; Liu, X.F.; Guo, G.L.; He, Z.H. Biological and ecological features of saline lakes in northern Tibet, China. Hydrobiologia 2005, 541, 189–203. [Google Scholar]
- Vizer, L.S. Zooplankton Communities of Saline Reservoirs of the South of Western Siberia by Example of Chany Lake System. Doctoral Biology Dissertation, Digital Science & Education LP, Novosibirsk, Russia, 2016. Available online: https://www.dissercat.com/content/soobshchestva-zooplanktona-solonovatykh-vodoemov-yuga-zapadnoi-sibiri-na-primere-chanovskoi/read (accessed on 9 March 2022).
- Hammer, U.T.; Shamess, J.; Haynes, R.C. The distribution and abundance of algae in saline lakes of Saskatchewan, Canada. Hydrobiologia 1983, 105, 1–26. [Google Scholar] [CrossRef]
- Hammer, U.T.; Parker, J. Limnology of a perturbed highly saline Canadian lake. Arch. Fur Hydrobiol. 1984, 102, 31–42. [Google Scholar] [CrossRef]
- Larson, C.A.; Belovsky, G.E. Salinity and nutrients influence species richness and evenness of phytoplankton communities in microcosm experiments from Great Salt Lake, Utah, USA. J. Plankton Res. 2013, 35, 1154–1166. [Google Scholar] [CrossRef]
- Ivanova, M.B.; Kazantseva, T.I. Effect of Water pH and Total Dissolved Solids on the Species Diversity of Pelagic Zooplankton in Lakes: A Statistical Analysis. Rus. J. Ecol. 2006, 37, 264–270. [Google Scholar] [CrossRef]
- Boronat, L.; Miracle, M.R.; Armengol, X. Cladoceran assemblages in a mineralization gradient. Hydrobiologia 2001, 442, 75–88. [Google Scholar] [CrossRef]
- Frisch, D.; Moreno-Ostos, E.; Green, A. Species richness and distribution of copepods and cladocerans and their relation to hydroperiod and other environmental variables in Doñana, south-west Spain. Hydrobiologia 2006, 556, 327–340. [Google Scholar] [CrossRef]
- Williams, W.D.; Boulton, A.J.; Taffee, R.G. Salinity as a determinant of salt lake fauna: A question of scale. Hydrobiologia 1990, 197, 257–266. [Google Scholar] [CrossRef]
- Nedli, J.; De Meester, L.; Major, A.; Schwenk, K.; Szivak, I.; Forro, L. Salinity and depth as structuring factors of cryptic divergence in Moina brachiata (Crustacea: Cladocera). Fundam. Appl. Limnol. 2014, 184, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, Y.; Nakahara, H.; Katayama, Y.; Ishida, N. The phytoplankton of some saline lakes in Central Asia. Int. J. Salt Lake Res. 1997, 6, 5–16. [Google Scholar] [CrossRef]
- Popova, T.G. The main features of the algal distribution and composition in the lakes Chany and Yarkul during the high water period 1947–1948. In Algae, Mushrooms and Lichens of the South of Siberia; Naplekova, N.N., Levadnaya, G.D., Eds.; Nauka: Moscow, Russia, 1980; pp. 146–147. [Google Scholar]
- Dokulil, M. Impact of climate warming on European inland waters. Inland Waters 2013, 4, 27–40. [Google Scholar] [CrossRef]
- Stephens, D.W. Changes in lake levels, salinity and the biological community of Great Salt Lake (Utah, USA), 1847–1987. Hydrobiologia 1990, 197, 139–146. [Google Scholar] [CrossRef]
- Zotina, T.A.; Tolomeyev, A.P.; Degermendzhy, N.N. Lake Shira, a Siberian salt lake: Ecosystem structure and function. 1. Major physico-chemical and biological features. Int. J. Salt Lake Res. 1999, 8, 211–232. [Google Scholar] [CrossRef]
- Evans, J.C.; Prepas, E.E. Potential effects of climate change on ion chemistry and phytoplankton communities in prairie saline lakes. Limnol. Ocean. 1996, 41, 1063–1076. [Google Scholar] [CrossRef]
- Conte, F.P.; Geddes, M.C. Acid brine shrimp: Metabolic strategies in osmotic and ionic adaptation. Hydrobiologia 1988, 158, 191–200. [Google Scholar] [CrossRef]
- Ermolaeva, N.I.; Fetter, G.V. Influence of the ionic composition of water on the structure of the zooplankton of the lakes of the Tazheran Steppe (Western Baikalia). Arid. Ecosyst. 2021, 11, 411–420. [Google Scholar] [CrossRef]
- Derry, A.; Prepas, E.; Hebert, P. A comparison of zooplankton communities in saline lakewater with variable anion composition. Hydrobiologia 2003, 505, 199–215. [Google Scholar] [CrossRef]
- Tashlykova, N.A.; Afonina, E.Y. Diversity of plankton communities of chloride lakes of Southeastern Transbaikalia. IOP Conf. Ser. Earth Environ. Sci. 2022, 1112, 012108. [Google Scholar] [CrossRef]
- Geddes, M.C.; De Deckker, P.; Williams, W.D.; Morton, D.W.; Topping, M. On the chemistry and biota of some saline lakes in Western Australia. Hydrobiologia 1981, 82, 201–222. [Google Scholar] [CrossRef]
- Comin, F.A.; Alonso, M.; Lopez, P.; Comelles, P. Limnology of Gallocanta Lake, Aragón, NE. Spain Hydrohiol. 1983, 105, 207–221. [Google Scholar] [CrossRef]
- Stephens, D.W. A summary of biological investigations concerning the Great Salt Lake, Utah (1861–1973). Great Basin Natural. 1974, 34, 221–229. [Google Scholar]
- Seaman, M.T.; Ashton, P.J.; Williams, W.D. Inland salt waters of southern Africa. Hydrobiologia 1991, 210, 75–91. [Google Scholar] [CrossRef]
- Zhao, W.; He, Z. Biological and ecological features of inland saline waters in North Hebei, China. Int. J. Salt Lake Res. 1999, 8, 267–285. [Google Scholar]
- Gao, Q.; Xu, Z.; Zhuang, P. The relation between distribution of zooplankton and salinity in the Changjiang Estuary. Chin. J. Oceanol. Limnol. 2008, 26, 178–185. [Google Scholar] [CrossRef]
- Hammer, M.L.; Appleton, C.C. Physical and chemical characteristics and phyllopod fauna of temporary pools in north-eastern Natal, RSA. Hydrobiologia 1991, 212, 95–104. [Google Scholar] [CrossRef]
- McCulloch, G.P.; Irvine, K.; Eckardt, F.D.; Bryant, R. Hydrochemical fluctuations and crustacean community composition in an ephemeral saline lake (Sua Pan, Makgadikgadi Botswana). Hydrobiologia 2008, 596, 31–46. [Google Scholar] [CrossRef]
- Litvinenko, L.I.; Litvinenko, A.I.; Boyko, E.G.; Kutsanov, K.V. Effect of environmental factors on the structure and functioning of biocoenoses of hyperhaline water reservoirs in the South of Western Siberia. Contemp. Probl. Ecol. 2013, 6, 252–261. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli-Flores, L.; Melo, S. Towards of functional classification of the freshwater phytoplankton. J. Plankton Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Echaniz, S.A.; Vignatti, A.M.; De Paggi, S.J.; Paggi, J.C.; Pilati, A. Zooplankton seasonal abundance of south American Saline shallow lakes. Int. Rev. Hydrob. 2006, 91, 86–100. [Google Scholar] [CrossRef]
- Echaniz, S.A.; Vignatti, A.M. Seasonal variation and influence of turbidity and salinity on the zooplankton of a saline lake in central Argentina. Lat. Am. J. Aquat. Res. 2011, 39, 306–315. [Google Scholar] [CrossRef]
- Rawson, D.S.; Moore, J.E. The saline lakes of Saskatchewan. Can. J. Res. 1944, 22, 141–201. [Google Scholar] [CrossRef]
- Hammer, U.T. Zooplankton distribution and abundance in saline lakes of Alberta and Saskatchewan, Canada. Int. J. Salt Lake Res. 1993, 2, 111–132. [Google Scholar] [CrossRef]
- Salm, C.R.; Saros, J.E.; Martin, C.S.; Erickson, J.M. Patterns of seasonal phytoplankton distribution in prairie saline lakes of the northern Great Plains (U.S.A.). Aquat. Biosyst. 2009, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ionescu, V.; Năstăsescu, M.; Spiridon, L.; Bulgăreanu, V.A.C. The biota of Romanian saline lakes on rock salt bodies: A review. Int. J. Salt Lake Res. 1998, 7, 45–80. [Google Scholar] [CrossRef]
- Harper, D.M.; Childress, R.B.; Harper, M.M.; Boar, R.R.; Hickley, P.; Mills, S.C.; Otieno, N.; Drane, T.; Vareschi, E.; Nasirwa, O.; et al. Aquatic biodiversity and saline lakes: Lake Bogoria National Reserve, Kenya. Hydrobiologia 2003, 500, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Alimov, A.F. (Ed.) Soda Lakes of Transbaikalia: Ecology and Productivity; Nauka: Novosibirsk, Russia, 1991; 216p. [Google Scholar]
- Namsaraev, B.B.; Gorlenko, V.M.; Namsaraev, Z.B.; Garmaev, E.Z.; Abidueva, E.Y.; Dambaev, V.B.; Barkhutova, D.D.; Khahinov, V.V.; Zamana, L.V.; Borzenko, S.V.; et al. Brackish and Saline lakes of Transbaikalia: Hydrochemistry, Biology, 1st ed.; Buryat State University Publication: Ulan-Ude, Russia, 2009; 340p. [Google Scholar]
- Krylov, A.V.; Mendsaihan, B.; Ayushsuren, C.; Tsvetkov, A.I. Zooplankton of pulsating lakes Orog and Tatsyn-Tsagaan (West Mongolia) in the period of the beginning of the stabilization of water-level regime. Inland Water Biol. 2020, 13, 242–250. [Google Scholar] [CrossRef]
- Zadereev, E.; Lipka, O.; Karimov, B.; Krylenko, M.; Elias, V.; Pinto, I.S.; Alizade, V.; Anker, Y.; Feest, A.; Kuznetsova, D.; et al. Overview of past, current, and future ecosystem and biodiversity trends of inland soda lakes of Europe and Central Asia. Inland Waters 2020, 10, 438–452. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Period | Soda | Chloride | Sulfate | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Subtype | I | II | III | ||||||||||||||
| Number of Samples | Bain-Tsagan | Bain-Bulak | Balyktui | Nozhii | Kudzhertaj | Nizhniy Mukei | Sheluta | Haraganash | Borzinskoe | Ukshinda | Shvarcivskoe | Dabosa-Nor | Hilgonta | Gorbunka | Shihalin-Nur | Barun-Shivertuj | |
| Phytoplankton | |||||||||||||||||
| In littoral | 2021 | 1 | 1 | 1 | 2 | 1 | 1 | – | – | 1 | 1 | – | 1 | 1 | – | – | – |
| 2022 | 1 | – | – | 1 | 1 | – | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| In deep-water sites | 2021 | 3 | 1 | 1 | 2 | 1 | 1 | – | – | 1 | 1 | – | 1 | 1 | – | – | – |
| 2022 | 3 | – | – | 3 | 1 | – | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| Zooplankton | |||||||||||||||||
| In littoral | 2021 | 1 | 1 | 1 | 2 | – | 1 | – | – | – | 1 | – | 1 | 1 | – | – | |
| 2022 | 1 | – | – | – | 1 | – | 1 | 1 | 1 | – | 1 | 1 | 1 | 1 | 1 | 1 | |
| In deep-water sites | 2021 | 1 | 1 | 1 | 1 | – | 1 | – | – | – | 1 | – | 1 | 1 | – | – | |
| 2022 | 1 | – | – | – | 1 | – | 1 | 1 | 1 | – | 1 | 1 | 1 | 1 | 1 | 1 | |
| Type | Subtype | Lake | Year | Alt | V | h | SDT | T | TDS | pH | Turb | O2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soda | I | Bain-Tsagan | 2021 | 632 | 35.8 | 4.75 | 1.30 | 20.00 | 8.20 | 9.30 | 1.39 | – |
| 2022 | 4.50 | 1.60 | 23.91 | 8.76 | 9.40 | 7.60 | 6.6 | |||||
| Bain-Bulak | 2021 | 655 | 33 | 0.80 | 0.80 | 26.40 | 4.68 | 9.22 | 6.67 | 17.2 | ||
| Balyktui | 2021 | 750 | 2 | 0.20 | 0.10 | 27.10 | 7.99 | 9.30 | 139.58 | 13.2 | ||
| Kudzhertaj | 2021 | 653 | 10 | 0.20 | 0.20 | 21.00 | 194.50 | 9.90 | 635.42 | – | ||
| 2022 | 0.03 | 0.03 | 33.40 | 291.61 | 9.75 | 125.57 | 2,25 | |||||
| Nozhii | 2021 | 650 | 12 | 1.98 | 0.70 | 23.10 | 4.16 | 9.46 | 6.67 | 16.4 | ||
| 2022 | 2.30 | 2.30 | 22.40 | 3.49 | 9.34 | 2.25 | 9.56 | |||||
| Nizhniy Mukei | 2021 | 700 | – | 0.10 | 0.10 | 24.40 | 128.33 | 9.90 | 927.08 | 8,4 | ||
| II | Sheluta | 2022 | 669 | 8 | 1.00 | 0.35 | 23.50 | 6.22 | 9.48 | 18.57 | 9,34 | |
| Haraganash | 2022 | 694 | 14 | 0.15 | 0.10 | 18.40 | 3.65 | 9.04 | 63.07 | 8.61 | ||
| III | Borzinskoe | 2021 | 654 | 69.2 | 0.05 | 0.05 | 34.9 | 231.30 | 9.40 | – | 3.58 | |
| 2022 | 0.05 | 0.05 | 29.10 | 138.54 | 9.46 | 16.69 | 1.92 | |||||
| Ukshinda | 2021 | 651 | 16 | 0.20 | 0.20 | 24.0 | 16.14 | 9.20 | 32.71 | 7.28 | ||
| 2022 | 0.3 | 0.3 | 22.4 | 14.95 | 9.55 | 12.2 | 7.1 | |||||
| Shvarcivskoe | 2022 | 644 | 9 | 0.5 | 0.5 | 31.10 | 22.07 | 9.25 | 72.02 | 14.3 | ||
| Chloride | Dabosa-Nor | 2021 | 662 | 1.2 | 0.2 | 0.2 | 22.4 | 39.83 | 8.90 | 1.53 | – | |
| 2022 | 0.08 | 0.08 | 24.6 | 73.06 | 8.81 | 9.70 | 10.4 | |||||
| Hilgonta | 2021 | 663 | 0.5 | 0.1 | 0.1 | 20.9 | 33.87 | 8.9 | 8.06 | – | ||
| 2022 | 0.1 | 0.1 | 25.10 | 17.13 | 8.87 | 0.92 | 10.6 | |||||
| Gorbunka | 2021 | 661 | 0.2 | 0.1 | 0.1 | 28.5 | 69.87 | 8.52 | 34.38 | – | ||
| 2022 | 0.45 | 0.45 | 22.00 | 14.45 | 8.73 | 6.25 | 9.1 | |||||
| Sulfate | Shihalin-Nur | 2022 | 652 | 11 | 0.2 | 0.2 | 24.90 | 21.26 | 8.86 | 2.05 | 5.01 | |
| Barun-Shivertuj | 2022 | 487 | 5 | 0.5 | 0.5 | 25.80 | 45.38 | 8.87 | 11.70 | 4.32 | ||
| Taxon | Lake Name | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soda I Subtype | Soda II Subtype | Soda III Subtype | Chloride | Sulfate | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
| Phytoplankton | ||||||||||||||||
| Cyanobacteria | ||||||||||||||||
| Anabaena sp. | ab | a | a | - | ab | - | - | b | - | - | - | - | - | - | - | - |
| A. sp. sp. | - | - | ab | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Drouetiella lurida (Gomont) Mai, J.R.Johansen and Pietrasiak 2018 | - | - | - | - | - | - | - | - | b | - | - | - | - | - | - | - |
| Gloeocapsa minor (Kutzing) Hollerbach 1937 | - | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Jaaginema woronichinii (Anisimova) Anagnostidis and Komárek 1988 | - | - | - | - | - | - | - | - | - | - | - | - | a | - | - | - |
| Kamptonema formosum (Bory ex Gomont) Strunecký, Komárek and J. Smarda 2014 | - | - | - | - | - | - | - | - | - | - | a | - | - | - | - | |
| Limnospira fusiformis (Voronichin) Nowicka-Krawczyk, Mühlsteinová and Hauer 2019 | - | - | - | - | - | - | - | - | - | - | - | - | - | b | - | - |
| Merismopedia elegans A.Braun ex Kützing 1849 | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Nodularia spumigena Mertens ex Bornet and Flahault 1888 | - | - | - | - | b | - | - | b | - | - | - | - | - | - | - | - |
| Oscillatoria limosa C.Agardh ex Gomont 1892 | - | - | - | - | - | - | - | - | - | - | - | a | - | - | - | - |
| Oscillatoria major Vaucher ex Forti 1907 | - | - | - | - | - | - | - | - | - | - | - | a | a | - | - | - |
| O. sp. | - | a | - | - | - | - | - | - | - | a | - | - | - | - | - | - |
| O. sp. sp. | - | a | - | - | - | - | - | - | - | a | - | - | - | - | - | - |
| Phormidium breve (Kützing ex Gomont) Anagnostidis and Komárek 1988 | - | - | - | - | - | - | - | - | - | - | - | a | a | - | - | - |
| P. chalybeum (Mertens ex Gomont) Anagnostidis and Komárek 1988 | - | - | - | - | - | - | - | - | - | - | - | - | a | - | - | - |
| P. sp. | b | - | - | - | b | - | - | - | - | - | b | b | b | - | b | b |
| P. sp. sp. | - | - | - | - | - | - | - | - | - | - | - | b | - | - | - | b |
| Spirulina major Kützing ex Gomont 1892 | - | a | - | - | - | - | - | - | - | - | - | - | a | - | - | - |
| S. sp. | - | - | - | - | b | - | - | - | - | - | - | - | - | - | - | - |
| Bacillariophyta | ||||||||||||||||
| Amphora ovalis (Kützing) Kützing 1844 | b | - | a | - | - | - | - | - | - | - | - | - | a | - | - | - |
| Cymbella sp. | b | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Fragilaria crotonensis Kitton 1869 | b | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lindavia comta (Kützing) T.Nakov et al. 2015 | a | - | - | - | - | - | - | - | b | - | - | - | a | - | - | - |
| Navicula sp. | ab | - | a | - | b | - | - | - | - | b | - | - | - | b | b | - |
| N. sp. sp. | - | - | - | - | - | - | - | - | - | b | - | - | - | - | b | - |
| N. sp. sp. sp. | - | - | - | - | - | - | - | - | - | b | - | - | - | b | b | - |
| Nitzschia sigmoidea (Nitzsch) W.Smith 1853 | - | - | - | - | - | - | - | - | - | b | - | - | - | - | b | - |
| N. sp. | a | - | - | - | - | - | - | - | - | b | - | - | - | b | - | - |
| N. sp. sp. | - | - | - | - | - | - | - | - | - | b | - | - | - | b | b | - |
| Pinnularia sp. | a | - | - | - | - | - | - | - | a | - | - | - | - | - | - | - |
| Planothidium lanceolatum (Brébisson ex Kützing) Lange-Bertalot. 1999 | - | - | - | - | - | - | - | - | - | - | - | - | a | - | - | - |
| Synedra sp. | - | - | - | - | - | - | - | - | a | - | - | - | - | - | - | - |
| Peridimium sp. | - | - | - | - | b | - | - | - | - | - | - | - | - | - | - | - |
| Cryptophyta | ||||||||||||||||
| Chroomonas caudata L.Geitler 1924 | - | - | - | - | - | - | - | - | b | - | - | - | - | - | - | - |
| Cryptomonas marssonii Skuja 1948 | - | - | - | - | - | - | - | - | b | - | - | - | - | - | - | - |
| C. erosa Ehrenberg 1832 | ab | - | a | a | a | a | - | - | - | b | - | - | - | - | - | - |
| C. sp. | b | - | - | - | b | - | - | b | - | - | - | - | - | - | b | b |
| Rhodomonas salina (Wisłouch) D.R.A.Hill & R.Wetherbee 1989 | - | - | - | - | - | - | - | - | b | - | - | - | - | - | - | - |
| Chlorophyta | ||||||||||||||||
| Ankyra ancora (G.M.Smith) Fott 1957 | b | a | - | - | ab | - | - | b | - | a | - | - | - | - | b | b |
| Chlamydomonas sphagnicola (F.E.Fritsch) F.E.Fritsch and H.Takeda 1930 | - | - | - | - | - | - | - | - | - | - | - | - | a | - | - | - |
| C. sp. | - | a | - | a | - | - | - | - | - | - | - | - | a | - | - | - |
| C. sp. sp. | - | - | - | - | - | - | b | - | - | - | - | - | - | - | - | - |
| Closteriopsis acicularis (Chodat) J.H.Belcher and Swale 1926 | - | - | - | - | - | - | - | b | - | - | - | - | - | - | - | - |
| Coenocystis planctonica Korshikov 1953 | - | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Monoraphidium contortum (Thuret) Komárková-Legnerová in Fott 1969 | - | - | - | - | b | - | - | - | - | - | - | - | - | - | - | b |
| M. komarkovae Nygaard 1979 | - | - | - | - | b | - | - | - | - | - | - | - | - | - | - | b |
| M. sp. | - | - | - | - | a | - | - | - | - | a | - | - | - | - | - | - |
| Lagerheimia genevensis (Chodat) Chodat 1895 | - | - | - | - | - | - | - | - | - | - | b | - | b | - | - | - |
| Lemmermannia triangularis (Chodat) C.Bock and Krienitz 2013 | - | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| L. komarekii (Hindák) C.Bock and Krienitz 2013 | - | - | - | - | ab | - | - | - | - | - | - | - | - | - | - | - |
| Oocystis borgei J.W.Snow 1903 | - | - | - | - | - | - | - | b | - | b | b | b | - | b | b | - |
| O. rhomboidea Fott 1933 | - | - | - | - | - | - | - | - | - | - | - | a | - | - | - | - |
| O. sp. | - | - | - | - | b | - | - | b | - | b | - | b | - | - | b | - |
| Pandorina morum (O.F.Müller) Bory 1826 | - | - | - | - | - | - | - | - | - | - | - | b | - | - | - | - |
| Pseudopediastrum boryanum (Turpin) E.Hegewald 2005 | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Rhabdoderma sp. | - | - | - | - | b | - | - | - | - | - | - | - | - | - | - | - |
| Raphidocelis danubiana (Hindák) Marvan, Komárek and Comas 1984 | - | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Scenedesmus quadricauda (Turpin) Brébisson 1835 | - | - | - | - | ab | - | - | - | - | - | - | - | - | - | - | - |
| Schroederia robusta Korshikov 1953 | - | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Tetradesmus obliquus (Turpin) M.J.Wynne 2016 | - | - | - | - | - | - | - | b | - | - | - | - | - | - | - | - |
| Charophyta | ||||||||||||||||
| Spirogira sp. | - | - | - | - | - | - | - | - | - | b | - | - | - | - | - | - |
| Staurastrum sp. | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Euglenophyta | ||||||||||||||||
| Euglena sp. | - | - | - | - | b | - | - | b | - | - | - | - | - | - | b | - |
| Phacus sp. | - | - | - | - | ab | - | b | b | - | - | - | - | - | - | b | - |
| Zooplankton | ||||||||||||||||
| Rotifera | ||||||||||||||||
| Eosphora najas Ehrenberg, 1830 | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Lecane luna (Müller, 1776) | - | a | - | - | ab | - | - | b | - | - | - | - | - | - | - | - |
| Mytilina ventralis (Ehrenberg, 1832) | - | - | - | b | - | - | - | - | - | - | - | - | - | - | - | |
| Colurella adriatica Ehrenberg, 1831 | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Euchlanis dilatata Ehrenberg, 1832 | - | a | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Brachionus quadridentatus ancylognatus Schmarda, 1859 | - | - | - | b | - | - | - | - | - | - | - | - | - | - | - | |
| B. q. brevispinus Ehrenberg, 1832 | - | a | a | - | - | - | - | - | - | - | - | - | - | - | - | - |
| B. q. cluniobicularis Skorikov, 1894 | - | a | a | - | - | - | - | - | - | - | - | - | - | - | - | - |
| B. plicatilis Müller, 1786 | - | - | a | a | b | - | b | - | - | - | b | ab | b | a | - | b |
| B. p. asplanchnoides Charin, 1947 | - | - | - | - | - | - | - | b | b | - | - | - | - | - | b | - |
| B. leydigii (Cohen, 1862) | - | - | - | - | - | - | b | b | - | - | - | - | - | - | - | - |
| Hexarthra mira (Hudson, 1871) | ab | a | - | - | ab | - | - | b | b | ab | b | - | - | - | - | - |
| Anostraca | ||||||||||||||||
| Sp. | a | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Sp. Sp. | - | - | - | - | - | - | - | - | ab | - | - | b | - | a | - | b |
| Cladocera | ||||||||||||||||
| Daphnia magna Straus, 1820 | ab | a | - | - | ab | - | - | - | - | - | - | - | - | - | - | - |
| Moina brachiata (Jurine, 1820) | ab | a | a | - | ab | a | b | b | - | ab | - | - | b | b | b | - |
| Macrothrix laticornis (Jurine, 1820) | - | - | - | - | b | - | - | - | - | - | - | - | - | - | - | - |
| M. hirsuticornis Norman and Brady, 1867 | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Coronatella rectangula (Sars, 1862) | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Copepoda | ||||||||||||||||
| Metadiaptomus asiaticus (Uljanin, 1875) | ab | - | a | b | - | - | b | - | - | ab | - | - | b | - | b | - |
| Hemidiaptomus ignatovi Sars, 1903 | - | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Arctodiaptomus bacillifer (Koelbel, 1885) | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| A. niethammeri (Mann, 1940) | - | - | - | - | ab | - | - | b | - | - | - | - | - | - | - | - |
| Mixodiaptomus incrassatus (Sars, 1903) | - | - | - | - | a | - | - | - | - | - | - | - | - | - | - | - |
| Eucyclops serrulatus (Fischer, 1851) | - | a | - | - | b | - | - | - | - | - | - | - | - | - | - | - |
| Thermocyclops dybowskii (Lande, 1890) | - | a | a | - | b | - | b | b | - | - | - | - | - | - | - | - |
| Copepodita Cyclopoida | b | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Harpacticoida gen. sp. | - | a | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Type of Lake | Lake | Phytoplankton | Zooplankton | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | 2021 | 2022 | ||||||
| Number of Species | Dominant Species/Taxa | Number of Species | Dominant Species/Taxa | Number of Species | Dominant Species/Taxa | Number of Species | Dominant Species/Taxa | ||
| Soda I subtype | Bain-Tsagan | 6 | Anabaena sp. | 11 | Anabaena sp. | 6 | Metadiaptomus asiaticus | 5 | Metadiaptomus asiaticus |
| Bain-Bulak | 10 | Merismopedia elegans Anabaena sp. | 6 | – | 14 | Hexarthra mira Euchlanis dilatata Arctodiaptomus bacillifer Eucyclops serrulatuss | – | – | |
| Balyktui | 6 | Anabaena sp. | – | – | 5 | Moina brachiate Metadiaptomus asiaticus | – | – | |
| Nozhii | 11 | Ankyra ancora Anabaena sp. | 21 | Phormidium sp. | 11 | Euchlanis dilatata Daphnia magna Arctodiaptomus niethammeri Mixodiaptomus incrassatus | 6 | Hexarthra mira Thermocyclops dybowskii | |
| Soda II subtype | Kudzhertaj | 2 | Cryptomonas erosa | 1 | Cryptomonas sp. | 1 | Brachionus plicatilis | 1 | Metadiaptomus asiaticus |
| Nizhniy Mukei | 1 | Cryptomonas marsonii | – | – | 1 | Ephippia | – | – | |
| Sheluta | – | – | 18 | Closteriopsis acicularis | – | – | 7 | Thermocyclops dybowskii | |
| Haraganash | – | – | 3 | Ankyra ancora | – | – | 5 | Thermocyclops dybowskii Moina brachiata | |
| Soda III subtype | Ukshinda | 4 | Ankyra ancora Phormidium sp. | 5 | Phormidium sp. | 3 | Moina brachiata Metadiaptomus asiaticus | 3 | Metadiaptomus asiaticus |
| Shvarcivskoe | – | – | 4 | Limnospira fusiformis Cryptomonas erosa | - | – | 2 | Brachionus plicatilis | |
| Borzinskoe | 2 | Benthic diatoms | 6 | Chroomonas caudata | 1 | Anostraca | 4 | Metadiaptomus asiaticus | |
| Sulfate | Shihalin-Nur | – | – | 17 | Oocystis sp. | - | – | 3 | Moina brachiata Metadiaptomus asiaticus |
| Brun-Shivertuj | – | – | 7 | Lagerheimia genevensis Cryptomonas sp. | - | – | 2 | B. plicatilis | |
| Chloride | Gorbunka | 4 | Oscillatoria major Phormidium sp. | 6 | Monoraphidium sp. Phormidium sp. | 1 | Anostraca | 2 | Moina brachiata |
| Hilgonta | 12 | Cryptomonas sp. | 4 | Phormidium sp. | 1 | Brachionus plicatilis | 3 | Moina brachiata Metadiaptomus asiaticus | |
| Dabosa-Nor | 3 | Phormidium sp. | 6 | Phormidium sp. | 1 | Brachionus plicatilis | 2 | Brachionus plicatilis | |
| Indicators | H | TR | pH | T | TDS | Turb | O2 |
|---|---|---|---|---|---|---|---|
| Soda lakes (n = 42) | |||||||
| nph | – | 0.7402 * | – | – | −0.9939 *** | – | – |
| Nph | – | – | −0.5210 * | – | 0.8984 *** | – | – |
| Bph | 0.7544 ** | – | −0.5930 ** | 0.7912 * | 0.9797 *** | – | – |
| nz | – | – | – | – | −0.9239 * | – | – |
| Nz | – | – | −0.7736 ** | – | – | – | – |
| Bz | – | – | 0.5022 * | – | – | – | – |
| Chloride lakes (n = 30) | |||||||
| nph | – | – | – | – | – | – | – |
| Nph | 0.9996 ** | 0.9996 ** | – | – | – | 0.7984 * | – |
| Bph | 0.9970 * | 0.9617 * | – | −0.9999 *** | – | – | – |
| nz | – | – | – | – | – | – | – |
| Nz | – | – | – | −0.9999 *** | – | – | – |
| Bz | 0.9999 *** | 0.9999 *** | – | – | – | – | – |
| Sulfate lakes (n = 20) | |||||||
| nph | – | – | −0.9530 ** | – | – | −0.7997 * | −0.7974 * |
| Nph | – | – | 0.9999 *** | – | – | – | – |
| Bph | – | – | 0.9998 ** | – | – | – | – |
| nz | – | – | 0.9998 *** | – | – | – | – |
| Nz | – | – | 0.9555 ** | – | – | −0.7976 * | – |
| Bz | – | – | −0.9699 ** | – | – | – | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tashlykova, N.A.; Afonina, E.Y. Habitat and Features of Development of Plankton Communities in Salt Lakes (South-Eastern Transbaikalia, Russia). Diversity 2023, 15, 396. https://doi.org/10.3390/d15030396
Tashlykova NA, Afonina EY. Habitat and Features of Development of Plankton Communities in Salt Lakes (South-Eastern Transbaikalia, Russia). Diversity. 2023; 15(3):396. https://doi.org/10.3390/d15030396
Chicago/Turabian StyleTashlykova, Natalya A., and Ekaterina Yu. Afonina. 2023. "Habitat and Features of Development of Plankton Communities in Salt Lakes (South-Eastern Transbaikalia, Russia)" Diversity 15, no. 3: 396. https://doi.org/10.3390/d15030396
APA StyleTashlykova, N. A., & Afonina, E. Y. (2023). Habitat and Features of Development of Plankton Communities in Salt Lakes (South-Eastern Transbaikalia, Russia). Diversity, 15(3), 396. https://doi.org/10.3390/d15030396