Abstract
Spiders act as one of the major predators among arthropods in agroecosystems and are crucial to the control of pest densities. As they are almost exclusively predators, they have developed a number of hunting strategies, which have consequently impacted their complex trophic interactions. The development of molecular biology methods for environmental DNA analysis has made it easier to analyze such complex trophic networks. In order to ascertain the trophic interactions of spiders, a sampling of spider species was carried out in two olive groves in Zadar County, Croatia, under integrated and ecological pest management. To construct the trophic networks of spiders, we performed DNA metabarcoding. The combined general and spider-excluding primers were able to recognize prey from 12 distinct orders in the guts of the 57 spiders. According to our results, spiders have a complex trophic network that exhibits seasonal and site-specific variations. The results obtained from both sites having different pest management also confirmed that spiders consumed phytophagous insects in the highest ratio, including some important pests, in comparison to other prey and that management and guild type had an impact on the predation of phytophagous insects.
1. Introduction
Spiders (Arachnida, Araneae) are predatory arthropods that have a significant impact on terrestrial ecosystems. They play a key role as predators, feeding on a wide range of arthropods [1,2], mainly insects [3]. Due to their abundance, spiders are important contributors to the functioning of agroecosystems [4,5]. As generalist predators, spiders exhibit diverse hunting strategies and occupy various trophic niches [6], which allows them to effectively utilize multiple food sources and adapt to different environmental conditions. Their adaptability and capacity to occupy different trophic levels make them important contributors to ecological stability and potential regulators of pest populations [7].
Many researchers have recognized the potential of spiders for biocontrol of pest outbreaks in agroecosystems [4,8,9,10]. Spider families such as Thomisidae, Araneidae, Lycosidae, Oxyopidae, Eresidae, Clubionidae, and Hersiliidae have shown high potential as biocontrol agents due to their effective searching ability and predation on a wide variety of insect species [11].
However, intraguild predation among spiders and other predatory insects, such as carabid beetles, heteropteran bugs, and hymenopterans [2,12,13,14], may limit their effectiveness in controlling pest species in agricultural lands [14]. For instance, intraguild predation interaction between Lycosidae and Gnaphosidae spiders and carabid beetles extends beyond shared prey availability and hunting habitat [13]. In addition to pest species and other predators, spiders also prey on other species considered as alternative prey [15,16,17,18]. The density of alternative prey species can influence spider population dynamics, potentially affecting the level of predation on pests [18]. The effect of alternative prey can be twofold, either reducing predation pressure on the pest or increasing it [19]. However, recent research suggests that high rates of intraguild predation and a tendency to switch from pests to alternate prey significantly limit the biocontrol benefits of spiders [3].
Despite their abundance and potential importance in biocontrol, the trophic ecology of spiders in Mediterranean agricultural lands has been scarcely investigated and remains poorly recognized. Moreover, the Mediterranean region is highly susceptible to global climate and land changes [20]. The Mediterranean part of Croatia is characterized by carbonate parent rocks and complex water systems encompassing surface and underground ecosystems and thus even more susceptible to chemical pollution, primarily caused by pesticide use in agricultural land, which poses a significant threat to these ecosystems because contaminants may transfer via food webs [21]. The alternatives are the chemically synthesized pesticides that are used to preserve biodiversity and sustain the ecosystems. These are the main goals of The Farm to Fork Strategy within the European Green Deal [22], which aims to establish a fair, health-conscious, and environmentally friendly food systems. Within this context, the involvement and role of predatory arthropods could be a valuable addition to achieving these goals. Integrated pest management (IPM) and ecological pest management (EPM) rely on preventive strategies, including monitoring of pest populations in a field. IPM applies synthetic chemical pesticides, mainly insecticides and fertilizers, only after exceeding the threshold for economically viable damage. The goal is to balance human welfare, environmental health, and sustainable agricultural production. On the other hand, EPM generally excludes the use of synthetic chemical pesticides and fertilizers [23]. Instead, it prioritizes the augmentation of biodiversity to reinforce the natural processes and ecosystem services, aiming to enhance agricultural sustainability even more effectively than IPM.
Trophic interactions of generalist predators can be influenced by various factors, such as changes in prey density, seasonal variations of prey availability, pesticide applications, and harvest practices [1,7,24,25,26,27]. Predators in agricultural ecosystems are also influenced by management type [28]. Moreover, spider populations in agroecosystems are greatly influenced by the type of management. Caprio et al. (2015) found that organic plantations tend to have a higher population of active hunters and ambush hunters while conventional plantations support specialist spiders and web-building spiders [29]. Pesticides have a detrimental effect on spider biodiversity and population size due to migration, reduced reproductive potential, and behavioral changes, such as reduced predation [30,31]. In addition to their direct impact on spiders, pesticides from the group of insecticides also reduce prey diversity and biomass, thus negatively affecting spider populations [24,30]. Therefore, a comprehensive examination of the spider community and their trophic interactions in agroecosystems is crucial for developing effective and sustainable pest management strategies in Mediterranean agricultural areas.
This study focuses on olive groves, which are traditionally connected to the Mediterranean landscape and culture. The intensification of agriculture and the transition from traditionally managed to mono-agriculture have led to increased pressure for higher production and more aggressive management practices in Croatia [32] and across the Mediterranean part of Europe [33].
In recent years, the application of new-generation sequencing and dietary metabarcoding has accelerated research on trophic interactions and provided valuable data [34,35,36,37]. We utilize this methodology to examine the gut content of spiders in olive groves. The use of the metabarcoding approach allows to overcome challenges associated with spider fluid feeding and enables the detection of prey present in the spider’s gut. This type of approach demands careful selection of primers and downstream bioinformatics pipeline because all of this can influence final prey composition results [38,39].
To test our hypothesis that spiders, as abundant predatory arthropods in Mediterranean agriculture, can provide biocontrol services in olive groves, we analyzed their diets and prey choices in two olive groves, one with integrated and one with ecological pest management during spring and autumn. We predicted that spider families belonging to four hunting guilds—ground hunters, ambush hunters, other hunters, and web weavers—can cover the range of various prey, from the ground to the tree crown in the olive groves. More specifically, our goals were to (i) assign the prey composition within the spiders’ gut; (ii) compare the prey composition between spider groups with different hunting strategies and ecological niches; (iii) evaluate the influence of management type and season on prey composition; and (iv) assess the potential of spiders as biocontrol agents.
2. Materials and Methods
2.1. Field Collection and Identification of Collected Samples
Spider specimens were collected during spring and autumn in two consecutive years (2018 and 2019) from sampling sites located in Zadar County, in the Mediterranean part of Croatia. Collections were conducted in two olive orchards (located in the Poličnik area, 44°10′50.2″ N 15°21′15.9″ E, 217,000 m2 in size, and in Škabrnja, 44°04′23.7″ N 15°28′22.1″ E, 51,000 m2 in size; additional data can be found in Šerić Jelaska et al. 2022a [40]). Each olive grove represented a different management type, ecological pest management (EPM) or integrated pest management (IPM). Sample collection was conducted from April to July and from September to November in 2018 and 2019. Live spiders for molecular gut content analysis were collected using pitfall traps (plastic cups with 8 cm ø in the opening, volume 300 mL) with added small piece of sponge soaked with vinegar as an attractant and placed in the center of the plantations to avoid the edge effect on available prey. Additionally, spiders were collected by hand picking from the ground and the canopy. Traps were put in the soil, beneath the surface and opening protected by a stone from the weather conditions. The traps were emptied every 24 h to reduce potential DNA cross-contamination between samples. All collected samples were immediately stored in separate vials containing 100% ethanol to avoid cross-contamination. The ethanol for storing was replaced during field collection as well as in the laboratory to better preserve eDNA from the gut.
Morphological identification of collected specimens was performed in the laboratory under the Zeiss Stemi 305 stereomicroscope. Adult samples were identified to species level and juvenile to genus or family level using identification keys [41].
2.2. Isolation and Amplification of eDNA from Spiders’ Gut, Library Preparation and Next-Generation Sequencing
A total of 122 spiders collected in the field were processed in the laboratory for dietary DNA metabarcoding analyses. For each specimen, only the DNA from its abdomen was extracted in order to have the highest concentrations of prey DNA present compared with other body parts or the whole body [42]. DNA was isolated using the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol with some small adjustments. The abdomen was ground with sterile micro-pestles in the lysis buffer, and proteinase K was added. Subsequently, the samples were incubated at 56 °C for 12 h or more to ensure complete tissue fragmentation and better DNA yield. All extracted DNA samples were stored at −80 °C until further use. For every eleven spiders, at least one negative control, treated identically to the samples but without any tissue added, was included.
Two pairs of primers were selected for dietary metabarcoding of spider guts: BF3-BR2 [43], which amplifies a wide range of invertebrates, including spiders, and TelperionF-LaurelinR, which amplifies the DNA of a range of invertebrates but not the DNA of spiders, the so-called excluding primer [1]. The primer pair BF3-BR2 amplifies the 418 bp-long COI target region. The TelperionF-LaurelinR primer pair amplifies the 301 bp COI target region. Each primer used in PCR reactions was marked with a distinctive molecular identifier (MID) tag of length 8 bp, ensuring that each sample has a different combination of the forward and reverse primer for both primer pairs.
The PCR reaction was carried out in a total volume of 25 µL, with 12.5 µL of QIAGEN Multiplex PCR Master Mix (Qiagen, Hilden, Germany), 2.5 µL of 2 µM forward and reverse primers, 6.5 µL of sterile water, and 1 µL of DNA template. The reaction conditions were as follows: 95 °C for 15 min, followed by 35 cycles of denaturation at 94 °C for 30 s, primer annealing at 47 °C for the BF3-BR2 primer combination, or 54 °C for the TelperionF-LaurelinR primer combination, elongation at 72 °C for 90 s, and one cycle of final elongation at 72 °C for 10 min.
PCR reactions were carried out in 96-well plates, including negative and positive controls (mixture of genomic DNA of species Abax parallelus and Pacifastacus leniusculus), following the protocol from previous research [1,7]. Following the PCR reaction, the concentration of the amplified product was assessed using gel electrophoresis on a 2% agarose gel. Samples were pooled together based on band density visualized on the agarose gel in a way that ensures the equimolar concentrations of DNA. The average sample volume was used to group the negative controls. The resulting pooled sample sets were then purified using SPRIselect beads (Beckman Coulter, Brea, CA, USA) following the manufacturer’s instructions for left-side size selection. Following purification, the concentration of each pool was determined using a Qubit Fluorometer (ThermoFisher Scientific, Waltham, MA, USA) and the Qubit dsDNA High-Sensitivity Assay Kit. To ensure that the samples were roughly equimolar in terms of DNA concentration, the measured concentrations were used to merge samples in a super-pool. Illumina library was prepared at Novogene Europe (Cambridge, UK) by including Illumina Nextera adapters. Next-generation sequencing was performed on an Illumina Novaseq high-throughput instrument. Sequencing with an amplicon length of 250 bp was performed in both directions with a sequencing depth of 35,000 reads per sample.
2.3. Bioinformatic Data Processing
The bioinformatic pipeline was taken and adjusted from Drake et al. (2021) [37]. Sequencing on Illumina generated 9,234,272 reads for sequencing with the BF3-BR2 primer pair and 3,519,228 reads for sequencing with the TelperionF-LaurelinR primer pair. In FastP [44], all reads underwent quality checks and trimming. The read pairs were demultiplexed using Mothur v1.39.5 [45], eliminating the MID tags and primer sequences. Reads were demultiplexed to generate one file per sample ID using the Mothur-made files. The sample ID was added to the read headers for each file, and reads were then compiled into a single file. Sequences were denoised and grouped into operational taxonomic units (OTUs) using Unoise3 in Usearch11 [46]. Using BLASTn v2.7.1 [47] and a 97% identity threshold [48], the resulting sequences were given a taxonomic identity using data from GenBank. Any read counts lower than the largest number of reads in the relevant OTU’s negative controls were eliminated during data preparation for statistical analysis. The taxa recorded in the positive controls—distinct from those chosen for the positive controls—were computed as a percentage of the total reads in that positive control. Such a percentage was 1.6% for the BF3BR2 primer pair and 0.6% for TelperionF-LaurelinR and was used as a threshold to eliminate tag jumping and high amplification of the most prevalent taxa by removing sequences below such threshold for each sample.
2.4. Data Analyses
The diet composition of investigated spiders was compared based on their hunting guild, season (autumn or spring), and type of pest management practice. Spider hunting guilds were classified according to Cardoso et al. (2011) [49]. The analysis used binary data, i.e., the presence or absence of prey orders in the gut [50], and the prey prevalence in the gut of spiders was a response variable composed of the number of spiders in each orchard that consumed that specific prey category and those that did not consume this prey category. Because we analyzed the prey prevalence of spider guts based on the guild, the data were not independent. Therefore, we compared the data using generalized estimating equations (GEE) via “geeglm” in the “geepack” package in R v.4.2.1. [51] with binomial distribution and exchangeable correlation structure to account for the dependencies [52]. Additionally, we visualized differences using Sankey plots via “SankeyNetwork” in the “networkD3” [53] package in R v.4.2.1 [51].
Differences in the predation of spiders from different hunting guilds and management types were compared based on the economic importance of prey. The prey species recorded in the gut were classified into three groups: phytophagous prey, alternative prey (including detritophages, parasites, and pollinators), and predatory invertebrates (www.cabi.org/isc, last accessed on 29 September 2022). The number of specimens counted in the spider’s gut system and divided into the aforementioned groups was included in the analysis and was analyzed by the factorial analysis of variance (ANOVA) in TIBCO Statistica 13.5.0. program [54].
3. Results
3.1. Spiders’ Dietary Data and Consumption of Different Prey Orders
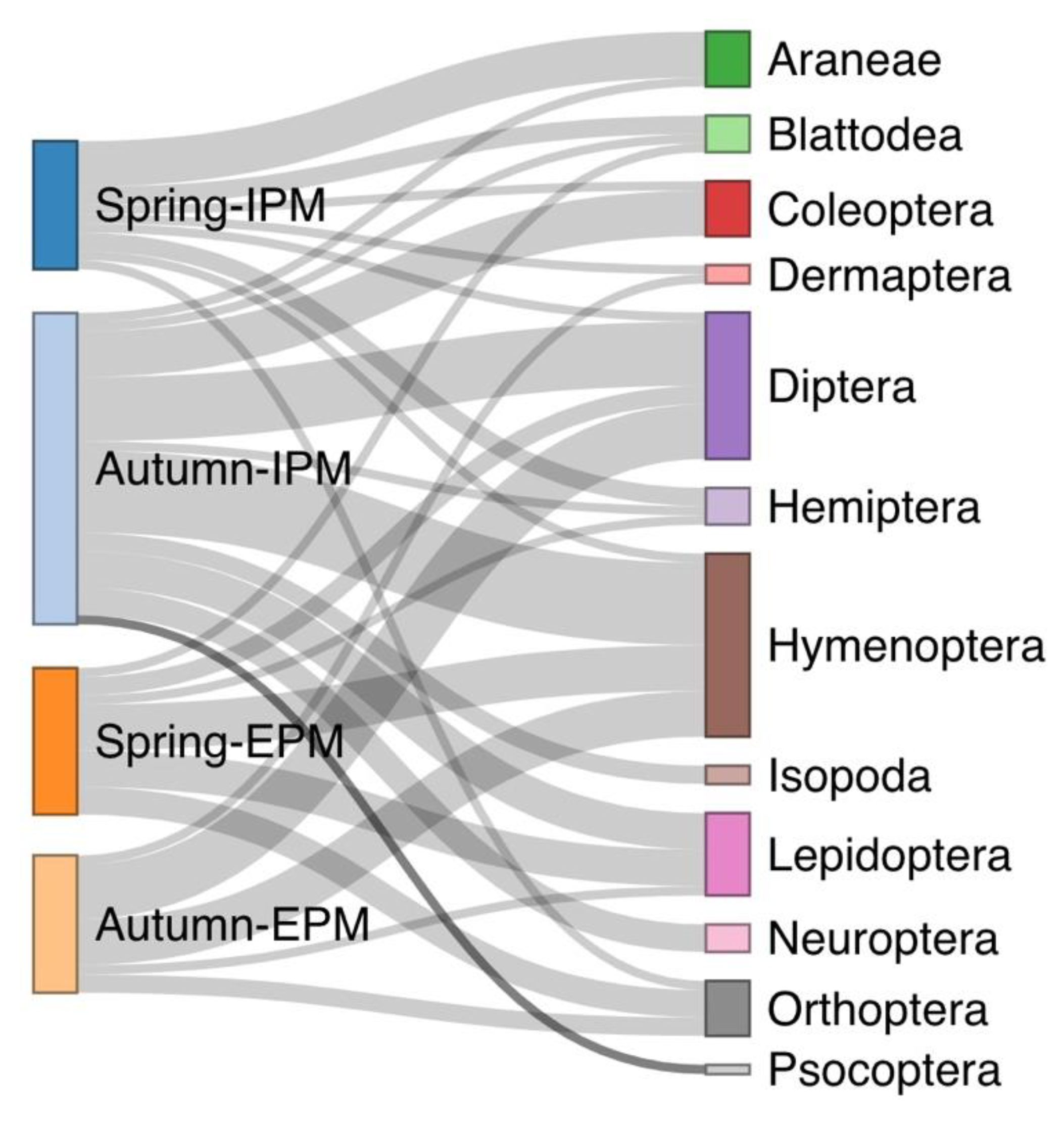
The majority of the spider specimens we collected belong to the families Gnaphosidae, Araneidae, Thomisidae, Lycosidae, and Oxyopidae, all recognized as generalist predators (Table 1). Dietary data were obtained for 57 out of the 122 screened spiders. Prey belonging to Blattodea, Dermaptera, Diptera, Hemiptera, Hymenoptera, Lepidoptera, and Orthoptera were detected in both, IPM and EPM olive groves (Table 1). Isopoda, Aranea, Coleoptera, Neuroptera, and Psocoptera were found only in the gut of spiders collected at the IPM site. Among the prey taxa detected in their gut (Table 1, Supplementary Table S1), only three species were shared between spiders from IPM and EPM sites, comprising 6,25% of the total number of species detected. Notably, the spiders’ guts have been found to contain some species that are listed in the literature as plant pests, including flies (Oscinella sp.) and moths (Cadra figulilella, Plutella xylostella, Prays oleae, Prays fraxinella). Moreover, species that are considered invasive to the study area, such as the moth species Plutella xylostella, the beetle Oxytelus sculptus, and the mosquito Aedes albopictus, were identified in the guts of tested spiders. The diet composition based on prey order in olive groves was significantly related to the type of management (GEE: χ2 = 656.8, d.f. = 11, p < 0.001) and the sampling season (GEE: χ2 = 402.74, d.f. = 11, p < 0.001) (Figure 1). In the IPM olive grove, Araneae was observed as the primary prey in spring, while Hymenoptera dominated in autumn. Conversely, the EPM olive grove showed Hymenoptera as the most common prey in spring and Diptera in autumn.

Table 1.
Spiders collected in the olive groves according to the management type, season of collection, and hunting strategies, and prey detected in their gut (species names matched by sequence OTUs—Operational Taxonomic Units results).
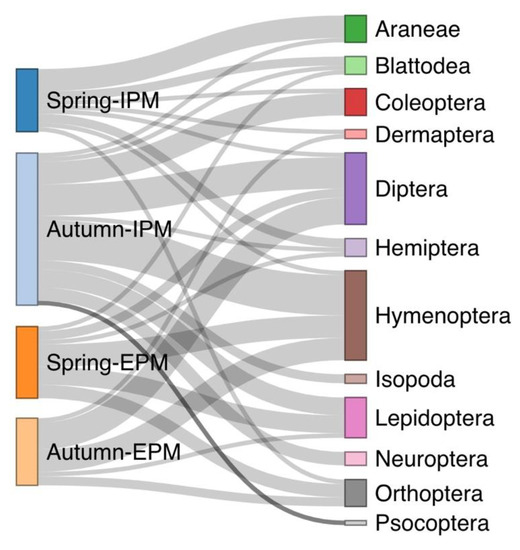
Figure 1.
Spiders’ dietary results divided by sampling site and sampling season for two olive orchards. The width of the mark for the sampling season–type of management indicates the relative abundance of spiders per group, the width of the flow paths indicates the relative importance (relative number of individuals recorded) in the diet of the spiders per group, and the width of the mark of a specific prey indicates the relative importance of that prey (relative number of individuals recorded) in the diet of spiders for all groups.
3.2. Consumption of Invertebrates as an Indicator of Spiders’ Biocontrol Potential
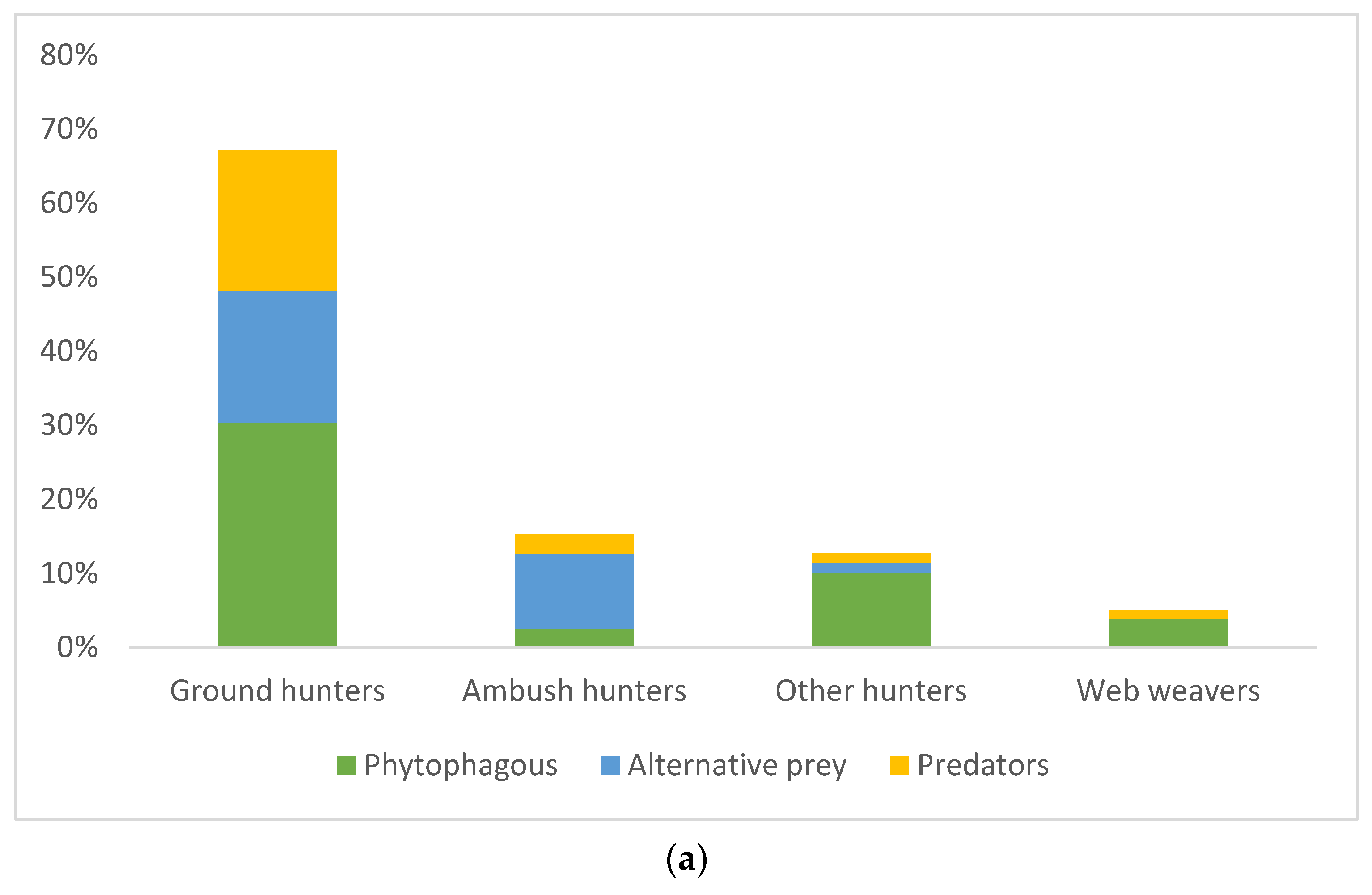
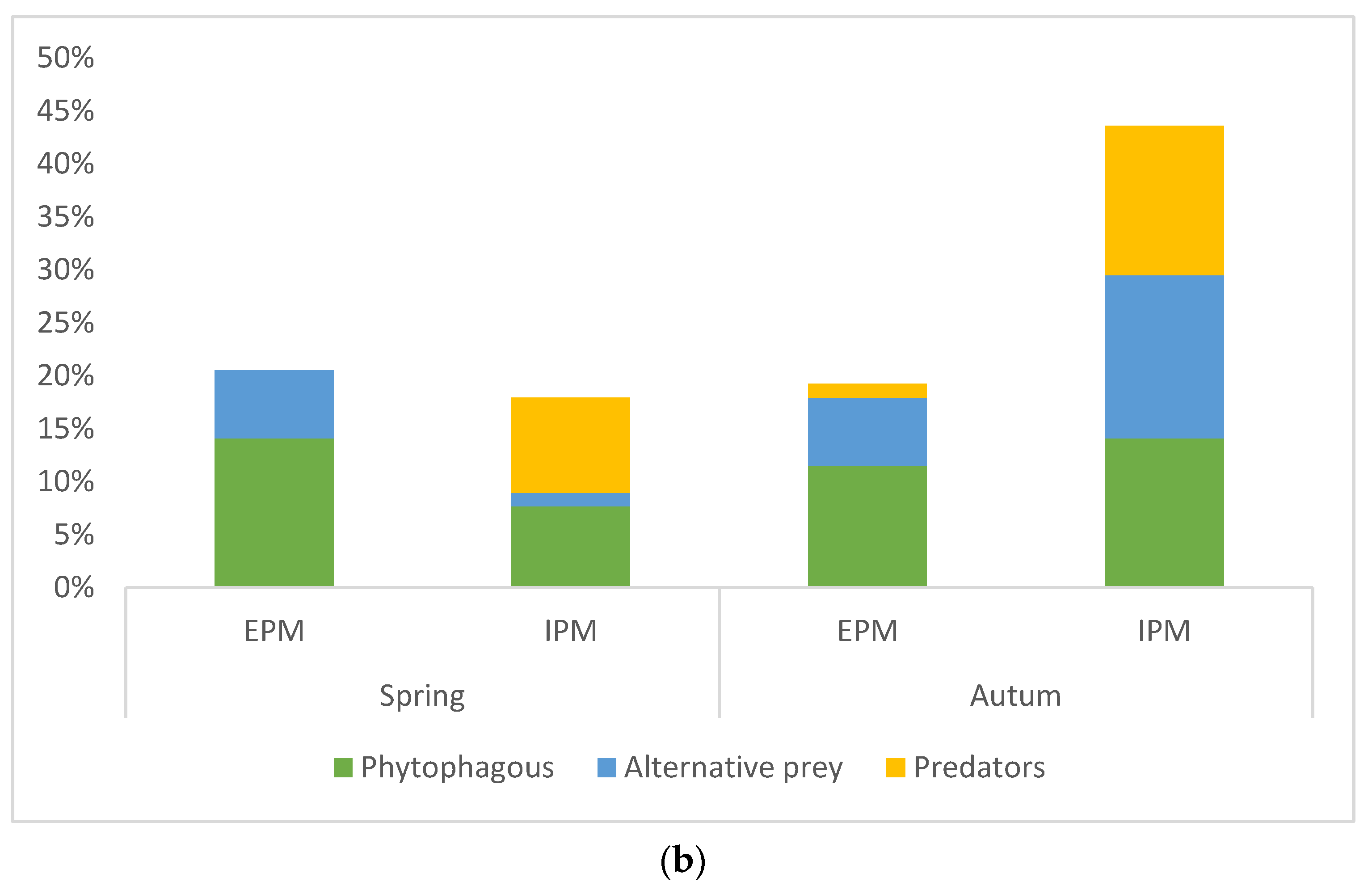
The diet composition based on economic importance of prey (phytophaga, predators, and other alternative prey) was significantly different between hunting guilds and management types (ANOVA: guild F = 3.60848, p = 0.03; management type F = 9.56146, p = 0.03). Hunting guild along with the type of pest management had a significant impact on the diet of spiders with phytophagous invertebrates and intraguild predation when compared with other alternative prey (factorial ANOVA: management*spiders guild F = 3.69335, p = 0.027) while the sampling season had no impact (Factorial ANOVA: p > 0.05, Figure 2a,b). Post hoc analyses confirmed significant differences in predation of ground hunter spiders on predatory insects at IPM site compared with ambush hunters and other hunters and between IPM and EPM sites (Tukey HSD, p < 0.05). Ground hunters had the highest intraguild predation (Figure 2a). On the other hand, the guts of other hunters showed the lowest intraguild predation. The phytophagous insects were the most abundant prey in the spiders’ guts with 46.84% at each site. At the sites with EPM, predators made up the least proportion of all prey (3.22%), whereas at the plots with IPM, alternative prey made up the smallest proportion of all prey (27.08%).
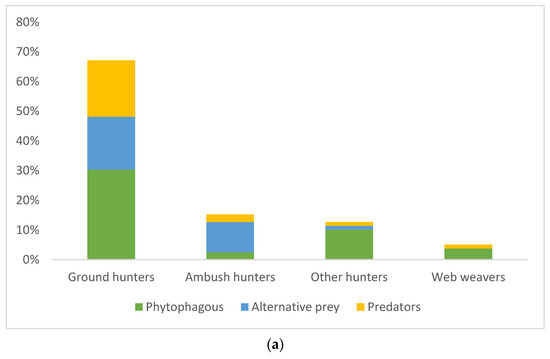
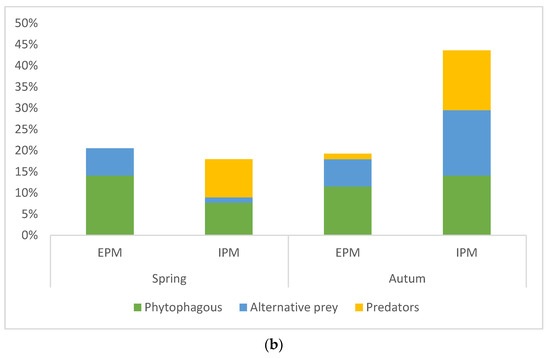
Figure 2.
(a) Percentage of occurrence of different prey groups (phytophagous, alternative, and predatory prey) in the guts of spiders from different hunting guilds, and (b) in two seasons (spring and autumn) within both pest management types (EPM—ecological pest management and IPM—integrated pest management).
4. Discussion
Understanding how management, seasonal changes, and functional traits of organisms affect trophic interactions in agroecosystems is an important factor in establishing proper ecosystem services and biological pest management measures [55,56]. This is especially the case when one is focused on generalist predators such as spiders, which can have a controversial role in biological control against pests [19,57]. Here we performed, to the best of our knowledge, the first-ever dietary metabarcoding study focusing on trophic interactions within olive groves. More precisely, this was the first study that focused on the diet of the most abundant and diverse invertebrate predators present in such agroecosystems—spiders. Furthermore, in this study, we provide the first insight into the dietary habits of different spider guilds and dietary distinctions between two management types and seasons in Mediterranean agroecosystems.
Generally, the analyzed spiders revealed a broad variety of prey, belonging to 12 distinct taxonomic orders of arthropods. The identified MOTUs were consistent with findings from our DNA barcoding-based biodiversity assessment of local fauna [58], as expected given their prevalence and abundance in agricultural ecosystems [59]. The six types of prey detected that made up over 79% of the overall diet of spiders were Hymenoptera, Diptera, Lepidoptera, Coleoptera, Araneae, and Orthoptera, which are common types of prey in similar spider guilds in pome fruit orchards [3,14] or cabbage and cauliflower farms [2,60], but in different relative frequencies.
Our findings showed that spiders engaged in both intra- and extraguild predation, mainly with a preference for other spiders, predatory coleopterans, and predatory ants. Although some of the ants that were preyed on were not predators, the majority were predatory ants from the genera Tetramorium, Aphaenogaster, and Solenopsis. Intraguild predation varied among hunting guilds and was mainly present in ground-hunting spiders. This was similarly confirmed by other studies that focused on many different spider guilds [2,61].
The spiders’ guts have been found to contain some species classified as pests, such as flies Cystiphora sonchi and Oscinella sp. in EPM olive grove, and moths Plutella xylostella in IPM, and Prays oleae and Prays fraxinella in EPM olive groves. However, these pests were found rarely, each species in only one or two spider specimens. Nevertheless, there were still a significant number of herbivores present in the guts of spiders, which are not considered general pests.
Previous dietary studies focusing on arthropod predators within agroecosystems have consistently shown the profound influence of management strategies and seasonal changes on diet composition [14,26,27,53]. In our small study, we had the same effect. The diversity of prey in the EPM olive grove was lower compared with the IPM olive grove and was dominated by only seven orders of prey: Blattodea, Dermaptera, Diptera, Hemiptera, Hymenoptera, Lepidoptera, and Orthoptera. The main reason for such low diversity of prey in EPM is most likely due to the general rocky soil and sparse vegetation at the study site, causing low species diversity [56,62]. The type of management had a significant impact on the predation of phytophagous insects. A higher predation intensity was found for spiders from the EPM olive grove, where the ratio between phytophagous and intraguild prey was higher compared with spiders from the IPM olive grove. Saqib et al. (2022) and Gajski et al. (2023) showed that the ecological type of management and the lower use of pesticides have a positive impact on the biocontrol functions of predators, which was also confirmed by this research [2,63].
In spring, the overall diet of all investigated spiders in olive groves was not really dominated by any prey type and was more even and diverse, while in autumn it was mainly dominated by dipterans and hymenopterans. Interestingly, intraguild predation on other spiders during autumn was much lower than in spring, in contrast to a study on web-building spiders in barley fields where increased intraguild predation was observed during the autumn period [1]. Generally, seasonal change showed no effect on the ratio of phytophagous insects compared with other invertebrates and predators in the spiders’ diet. Although different prey types were consumed in autumn and spring, the consumption of potential plant-related pests did not vary significantly. Higher intraguild predation in the spring season could also be due to the higher abundance of some predatory arthropods (spiders, carabid beetles, etc.) in spring [40,58] or scavenging on dead animals (i.e., after agricultural measures undertaken in spring), but gut content analyses applied in this study cannot distinguish predation from scavenging events.
5. Conclusions
In conclusion, we demonstrate diverse dietary metabarcoding results of spiders inhabiting olive groves. Management type, as well as guild type, significantly influenced predation on phytophagous insects. The ground hunters had the highest proportion of intraguild predation among all guilds. On the other hand, the overall predation on phytophagous insects remained consistent across seasons, suggesting a combined constant rate of predation in both management types. As this is the first dietary study that focused on any predators in olive groves by performing dietary metabarcoding, we motivate other researchers to continue on this path of understanding the interactions that are happening in Mediterranean agroecosystems. These ongoing efforts have the potential to strengthen advocacy for appropriate and efficient biocontrol measures in the future while preserving the rich biodiversity and potentially facilitating its growth.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15090976/s1, Table S1: Spider individuals and corresponding prey detected in their gut DNA.
Author Contributions
Conceptualization: L.Š.J., T.K., B.A.D., and D.G.; methodology: L.Š.J., B.A.D., M.J., and D.G.; validation: L.Š.J.; field sampling set up: T.K.; formal analysis: B.A.D. and L.Š.J.; taxonomic identification: D.G.; resources: L.Š.J.; data curation: B.A.D. and M.J.; writing initial draft B.A.D.; rewriting Introduction, Results and Discussion: L.Š.J. and D.G.; visualization: B.A.D. and L.Š.J.; supervision: L.Š.J.; funding acquisition: L.Š.J. All authors have commented on all previous versions of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Croatian Science Foundation under the MEDITERATRI Project (HRZZ UIP-2017-05-1046), and Unity through Knowledge (2A Grant Agreement No 10/19) granted to Lucija Šerić Jelaska and co-funded by the Department of Biology, Faculty of Science at the University of Zagreb, and CryoEntoArk project funded by Croatian Academy of Sciences and Art (HAZU) and Croatian Ecological Society (HED) (10-102/324-247-2021).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data supporting reported results can be found at Mendeley Data, V1, doi: 10.17632/37n65hx7rc.1.
Acknowledgments
We are thankful to Lara Ivanković Tatalović for her help in the field and laboratory work, Aneta Kulić for technical help, and Dale Ann Acal for English language editing. DG was a Brno PhD Talent Scholarship Holder—funded by the Brno City Municipality.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cuff, J.P.; Drake, L.E.; Tercel, M.P.; Stockdale, J.E.; Orozco-terWengel, P.; Bell, J.R.; Vaughan, I.P.; Müller, C.T.; Symondson, W.O. Money spider dietary choice in pre- and post-harvest cereal crops using metabarcoding. Ecol. Entomol. 2021, 46, 249–261. [Google Scholar] [CrossRef]
- Saqib, H.S.A.; Sun, L.; Pozsgai, G.; Liang, P.; You, M.; Gurr, G.M.; You, S. DNA metabarcoding of gut contents reveals key habitat and seasonal drivers of trophic networks involving generalist predators in agricultural landscapes. Pest Manag. Sci. 2022, 78, 5390–5401. [Google Scholar] [CrossRef]
- Mezőfi, L.; Markó, G.; Nagy, C.; Korányi, D.; Markó, V. Beyond polyphagy and opportunism: Natural prey of hunting spiders in the canopy of apple trees. PeerJ 2020, 8, e9334. [Google Scholar] [CrossRef] [PubMed]
- Nyffeler, M.; Sunderland, K.D. Composition, abundance and pest control potential of spider communities in agroecosystems: A comparison of European and US studies. Agriculture. Ecosyst. Environ. 2003, 95, 579–612. [Google Scholar] [CrossRef]
- Živković, P.; Lemić, D.; Samu, F.; Kos, T.; Barić, B. Spider communities affected by exclusion nets. Appl. Ecol. Environ. Res. 2019, 17, 879–887. [Google Scholar] [CrossRef]
- Sanders, D.; Vogel, E.; Knop, E. Individual and species-specific traits explain niche size and functional role in spiders as generalist predators. J. Anim. Ecol. 2015, 84, 134–142. [Google Scholar] [CrossRef]
- Cuff, J.P.; Tercel, M.P.T.G.; Drake, L.E.; Vaughan, I.P.; Bell, J.R.; Orozco-terWengel, P.; Müller, C.T.; Symondson, W.O.C. Density-independent prey choice, taxonomy, life history, and web characteristics determine the diet and biocontrol potential of spiders (Linyphiidae and Lycosidae) in cereal crops. Environ. DNA 2021, 4, 549–564. [Google Scholar] [CrossRef]
- Chapman, E.G.; Schmidt, J.M.; Welch, K.D.; Harwood, J.D. Molecular evidence for dietary selectivity and pest suppression potential in an epigeal spider community in winter wheat. Biol. Control 2013, 65, 72–86. [Google Scholar] [CrossRef]
- Gomez-Polo, P.; Alomar, O.; Castañé, C.; Agustí, N. Molecular tracking of arthropod predator–prey interactions in Mediterranean lettuce crops. Food Webs 2016, 9, 18–24. [Google Scholar] [CrossRef]
- Wagan, T.A.; Li, X.; Hua, H.; Cai, W. Starvation time and predatory efficiency of spider species on Bemisia tabaci (Homoptera: Aleyrodidae). Fla. Entomol. 2019, 102, 684–690. [Google Scholar] [CrossRef]
- Samiayyan, K. Spiders—The Generalist Super Predators in Agro-Ecosystems, Integrated Pest Management; Academic Press: Cambridge, MA, USA, 2014; pp. 283–310. [Google Scholar]
- Mestre, L.; Piñol, J.; Barrientos, J.A.; Espadaler, X.; Brewitt, K.; Werner, C.; Platner, C. Trophic structure of the spider community of a Mediterranean citrus grove: A stable isotope analysis. Basic Appl. Ecol. 2013, 14, 413–422. [Google Scholar] [CrossRef]
- Sow, A.; Haran, J.; Benoit, L.; Galan, M.; Brévault, T. DNA Metabarcoding as a Tool for Disentangling Food Webs in Agroecosystems. Insects 2020, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Hambäck, P.A.; Cirtwill, A.R.; García, D.; Grudzinska-Sterno, M.; Miñarro, M.; Tasin, M.; Yang, X.; Samnegård, U. More intraguild prey than pest species in arachnid diets may compromise biological control in apple orchards. Basic Appl. Ecol. 2021, 57, 1–13. [Google Scholar] [CrossRef]
- Agusti, N.; Shayler, S.P.; Harwood, J.D.; Vaughan, I.P.; Sunderland, K.D.; Symondson, W.O.C. Collembola as alternative prey sustaining spiders in arable ecosystems: Prey detection within predators using molecular markers. Mol. Ecol. 2003, 12, 3467–3475. [Google Scholar] [CrossRef]
- Harwood, J.D.; Sunderland, K.D.; Symondson, W.O. Prey selection by linyphiid spiders: Molecular tracking of the effects of alternative prey on rates of aphid consumption in the field. Mol. Ecol. 2004, 13, 3549–3560. [Google Scholar] [CrossRef]
- Kuusk, A.K.; Ekbom, B. Lycosid spiders and alternative food: Feeding behavior and implications for biological control. Biol. Control. 2010, 55, 20–26. [Google Scholar] [CrossRef]
- Axelsen, J.A.; Ruggle, P.; Holst, N.; Toft, S. Modelling natural control of cereal aphids. III. Linyphiid spiders and coccinellids. Acta. Jutl. 1997, 72, 221–231. [Google Scholar]
- Michalko, R.; Pekár, S.; Entling, M.H. An updated perspective on spiders as generalist predators in biological control. Oecologia 2019, 189, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Newbold, T.; Oppenheimer, P.; Etard, A.; Williams, J.J. Tropical and Mediterranean biodiversity is disproportionately sensi-tive to land-use and climate change. Nat. Ecol. Evol. 2020, 4, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Šerić Jelaska, L.; Franjević, D.; Jelaska, S.D.; Symondson, W.O.C. Prey detection in carabid beetles in woodland ecosystems by PCR analyses of gut content. E. Jour. Ent. 2014, 111, 631–638. [Google Scholar]
- European Commission. A Farm to Fork Strategy for a Fair, Healthy and Environmentally-Friendly Food System; COM (2020) 381 Final; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- European Commission (EC 889/2008) Commission Regulation (EC) No 889/2008 of 5 September 2008 Laying down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labelling of Organic Products with Regard to Organic Production, Labelling and Control, Special edition in Croatian: Chapter 15 Volume 008 P. 173–256; Latest consolidated version: 01/01/2022; European Commission: Brussels, Belgium, 2008.
- Uhl, P.; Bucher, R.; Schäfer, R.B.; Entling, M.H. Sublethal effects of imidacloprid on interactions in a tritrophic system of non-target species. Chemosphere 2015, 132, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Petráková, L.; Michalko, R.; Loverre, P.; Sentenská, L.; Korenko, S.; Pekár, S. Intraguild predation among spiders and their effect on the pear psylla during winter. Agric. Ecosyst. Environ. 2016, 233, 67–74. [Google Scholar] [CrossRef]
- Šerić Jelaska, L.; Jurasović, J.; Brown, S.D.; Vaughan, P.I.; Symondson, W.O.C. Molecular field analysis of trophic relationships in soil-dwelling invertebrates to identify mercury, lead and cadmium transmission through forest ecosystems. Mol. Ecol. 2014, 23, 3755–3766. [Google Scholar] [CrossRef]
- Šerić Jelaska, L.; Symondson, O. Predation on epigeic, endogeic and anecic earthworms by carabids active in spring and autumn. Periodicum. Biologorum. 2016, 118, 281–289. [Google Scholar] [CrossRef]
- Menalled, F.D.; Smith, R.G.; Dauer, J.T.; Fox, T.B. Impact of agricultural management on carabid communities and weed seed predation. Agric. Ecosyst. Environ. 2007, 118, 49–54. [Google Scholar] [CrossRef]
- Caprio, E.; Nervo, B.; Isaia, M.; Allegro, G.; Rolando, A. Organic versus conventional systems in viticulture: Comparative effects on spiders and carabids in vineyards and adjacent forests. Agric. Syst. 2015, 136, 61–69. [Google Scholar] [CrossRef]
- Pekár, S. Spiders (Araneae) in the pesticide world: An ecotoxicological review. Pest Manag. Sci. 2012, 68, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Benamú, M.; Schneider, M.; González, A.; Sánchez, N. Short and long-term effects of three neurotoxic insecticides on biological and behavioural attributes of the orb-web spider Alpaida veniliae (Araneae, Araneidae): Implications for IPM programs. Ecotoxicology 2013, 22, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Šimunović, V. Stanje maslinarstva i uljarstva u Republici Hrvatskoj. Pomologia Croatica 2005, 11, 69–78. [Google Scholar]
- Concepción, E.D.; Díaz, M.; Baquero, R.A. Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landscape Ecol. 2008, 23, 135–148. [Google Scholar] [CrossRef]
- Ando, H.; Mukai, H.; Komura, T.; Dewi, T.; Ando, M.; Isagi, Y. Methodological trends and perspectives of animal dietary studies by noninvasive fecal DNA metabarcoding. Environ. DNA 2020, 2, 391–406. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Tabarlet, P. Who is eating what: Diet assessment using next generation sequencing? Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef]
- Kennedy, S.R.; Prost, S.; Overcast, I.; Rominger, A.J.; Gillespie, R.G.; Krehenwinkel, H. High-throughput sequencing for community analysis: The promise of DNA barcoding to uncover diversity, relatedness, abundances and interactions in spider communities. Dev. Genes Evol. 2020, 230, 185–201. [Google Scholar] [CrossRef] [PubMed]
- Drake, L.E.; Cuff, J.P.; Young, R.E.; Marchbank, A.; Chadwick, E.A.; Symondson, W.O.C. Post-bioinformatic methods to identify and reduce the prevalence of artefacts in metabarcoding data. Authorea 2021. [Google Scholar] [CrossRef]
- Clare, E.L.; Chain, F.J.; Littlefair, J.E.; Cristescu, M.E. The effects of parameter choice on defining molecular operational taxonomic units and resulting ecological analyses of metabarcoding data. Genome 2016, 59, 981–990. [Google Scholar] [CrossRef]
- Dušátková, L.P.; Pekár, S.; Michálek, O.; Líznarová, E.; Symondson, W.O.C. Estimation of trophic niches in myrmecophagous spider predators. Sci. Rep. 2020, 10, 8683. [Google Scholar] [CrossRef]
- Šerić Jelaska, L.; Ivanković Tatalović, L.; Kostanjšek, F.; Kos, T. Ground beetle assemblages and distribution of functional traits in olive orchards and vineyards depend on the agricultural management practice. BioControl 2022, 67, 275–286. [Google Scholar] [CrossRef]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 3. 2022. Available online: https://www.araneae.nmbe.ch. (accessed on 20 February 2023).
- Macías-Hernández, N.; Athey, K.; Tonzo, V.; Wangensteen, O.S.; Arnedo, M.; Harwood, J.D. Molecular gut content analysis of different spider body parts. PLoS ONE 2018, 13, e0196589. [Google Scholar] [CrossRef] [PubMed]
- Elbrecht, V.; Braukmann, T.W.A.; Ivanova, N.V.; Prosser, S.W.J.; Hajibabaei, M.; Wright, M.; Zakharov, E.V.; Hebert, P.D.N.; Steinke, D. Validation of COI metabarcoding primers for terrestrial arthropods. PeerJ 2019, 7, e7745. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Alberdi, A.; Aizpurua, O.; Gilbert, M.T.P.; Bohmann, K. Scrutinizing key steps for reliable metabarcoding of environmental samples. Methods Ecol. Evol. 2017, 9, 134–147. [Google Scholar] [CrossRef]
- Cardoso, P.; Pekár, S.; Jocqué, R.; Coddington, J.A. Global patterns of guild composition and functional diversity of spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef] [PubMed]
- Deagle, B.E.; Thomas, A.C.; McInnes, J.C.; Clarke, L.J.; Vesterinen, E.J.; Clare, E.L.; Kartzinel, T.R.; Eveson, J.P. Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Mol. Ecol. 2019, 28, 391–406. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R—A Language and Environment for Statistical Computing. Available: R Foundation for Statistical Computing, Vienna. Available online: http://www.R-project.org/ (accessed on 2 September 2022).
- Pekár, S.; Brabec, M. Generalized estimating equations: A pragmatic and flexible approach to the marginal GLM modelling of correlated data in the behavioural sciences. Ethology 2018, 124, 86–93. [Google Scholar] [CrossRef]
- Allaire, J.J.; Gandrud, C.; Russell, K.; Yetman, C.J. NetworkD3: D3 JavaScript Network Graphs from R. R Package Version 0.4. Available online: https://CRAN.R-project.org/package=networkD3. (accessed on 2 September 2022).
- TIBCO Software Inc. Data Science Workbench. 2020. Available online: http://tibco.com (accessed on 1 May 2023).
- Diehl, E.; Mader, V.L.; Wolters, V.; Birkhofer, K. Management intensity and vegetation complexity affect web-building spiders and their prey. Oecologia 2013, 173, 579–589. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2003, 45, 175–201. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef]
- Anđelić Dmitrović, B.; Jelić, M.; Rota, E.; Šerić Jelaska, L. DNA Barcoding of Invertebrates Inhabiting Olive Orchards and Vineyards Accelerates Understudied Mediterranean Biodiversity Assessment. Diversity 2022, 14, 182. [Google Scholar] [CrossRef]
- Kubiak, K.L.; Pereira, J.A.; Tessaro, D.; Santos, S.A.P.; Benhadi-Marín, J. Functional diversity of epigeal spiders in the olive grove agroecosystem in northeastern Portugal: A comparison between crop and surrounding semi-natural habitats. Entomol. Exp. Appl. 2022, 170, 449–458. [Google Scholar] [CrossRef]
- Saqib, H.S.A.; Liang, P.; You, M.; Gurr, G.M. Molecular gut content analysis indicates the inter- and intraguild predation patterns of spiders in conventionally managed vegetable fields. Ecol. Evol. 2021, 11, 9543–9552. [Google Scholar] [CrossRef]
- Birkhofer, K.; Djoudi, E.A.; Schnerch, B.; Michalko, R. Climatic conditions and functional traits affect spider diets in agricultural and non-agricultural habitats worldwide. Ecography 2022, 3, e06090. [Google Scholar] [CrossRef]
- Aguilera, G.; Roslin, T.; Tamburini, G.; Birkhofer, K.; Caballero-Lopez, B.; Lindstrom, S.A.M.; Bommarco, R. Crop diversity benefits carabid and pollinator communities in landscapes with semi-natural habitats. J. Appl. Eco. 2000, 57, 2170–2179. [Google Scholar] [CrossRef]
- Gajski, D.; Mifková, T.; Košulič, O.; Michalek, O.; Štarhova-Serbina, L.; Michalko, R.; Pekar, S. Brace yourselves, winter is coming: The winter activity, natural diet, and prey preference of winter-active spiders on pear trees. J. Pest Sci. 2023. In press. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).