

Assessing the Safety and Therapeutic Efficacy of Cannabidiol Lipid Nanoparticles in Alleviating Metabolic and Memory Impairments and Hippocampal Histopathological Changes in Diabetic Parkinson’s Rats

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Physical Stability of Cannabidiol Lipid Nanoparticle (CBD-LNP)
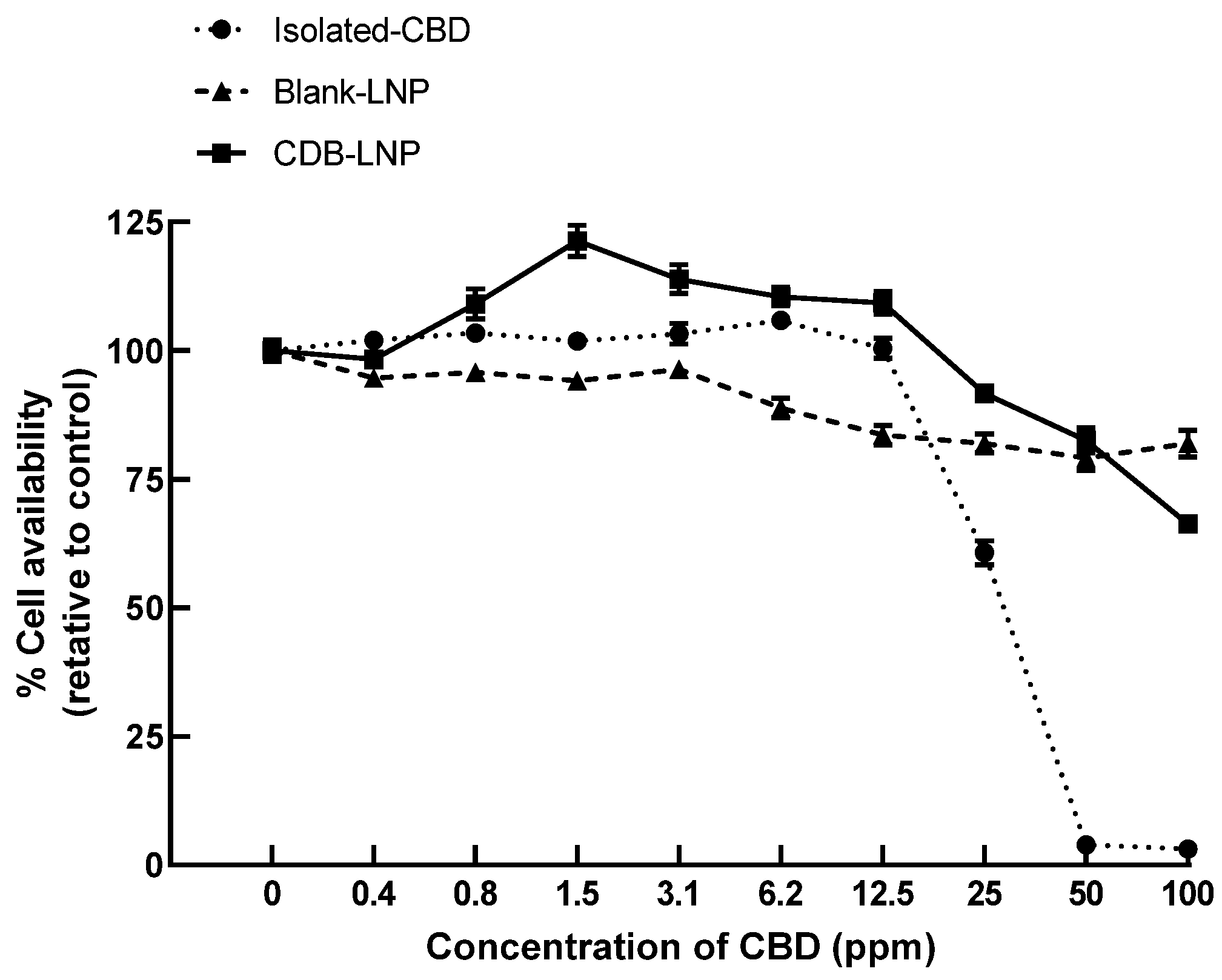
2.2. Assessment of Cytotoxicity
2.3. Animals
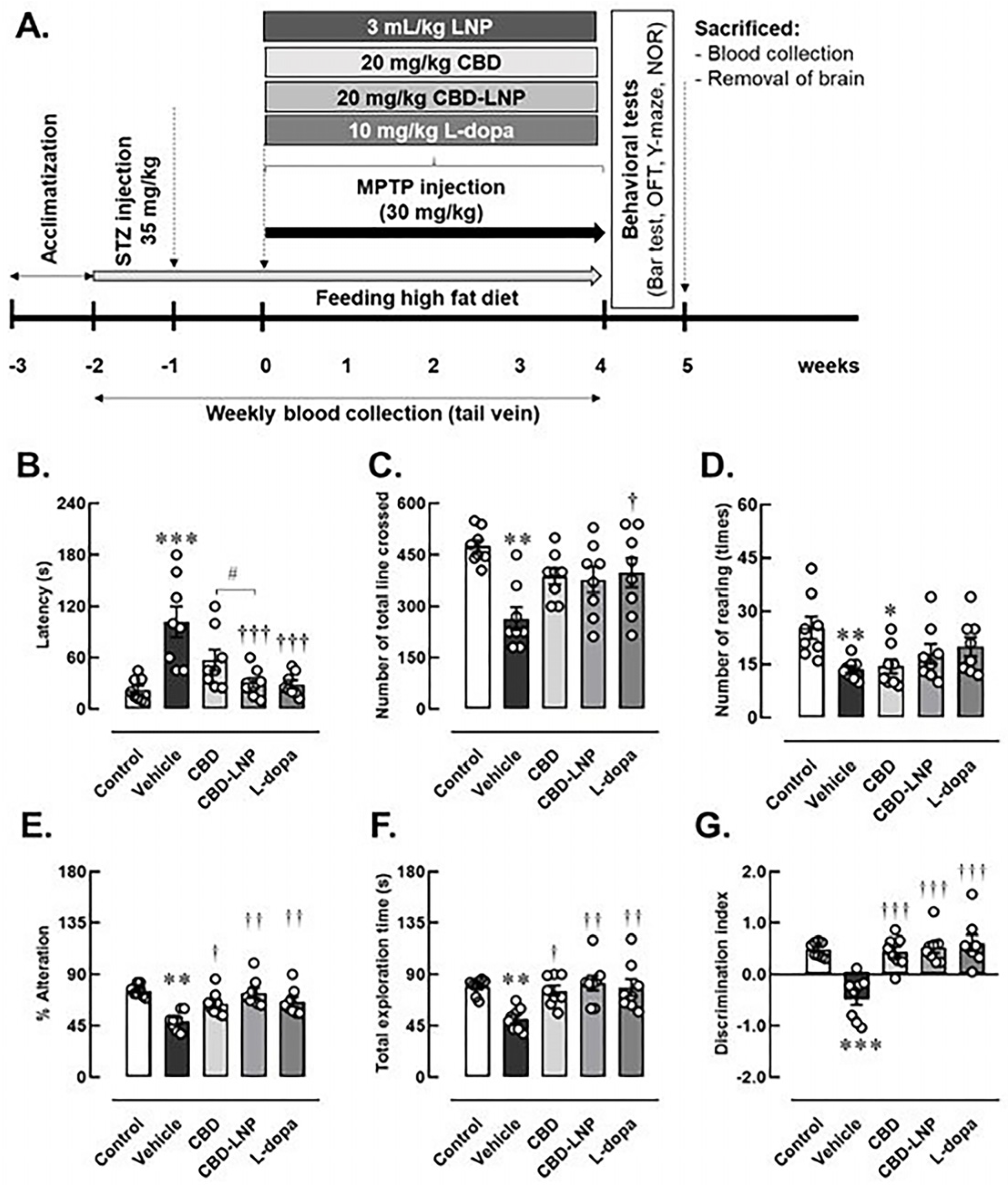
2.4. Induction of Diabetes and Parkinson’s Diseases (DP)
2.5. Administration of CBD-LNP
2.6. Administration of Antiparkinsonian Agent
2.7. Assessment of Muscular Rigidity-like Symptom
2.8. Assessment of Memory-like Behaviors
2.8.1. Y-Maze Test
2.8.2. Novel Object Recognition (NOR) Test
2.9. Assessment of Locomotor and Exploratory Activities
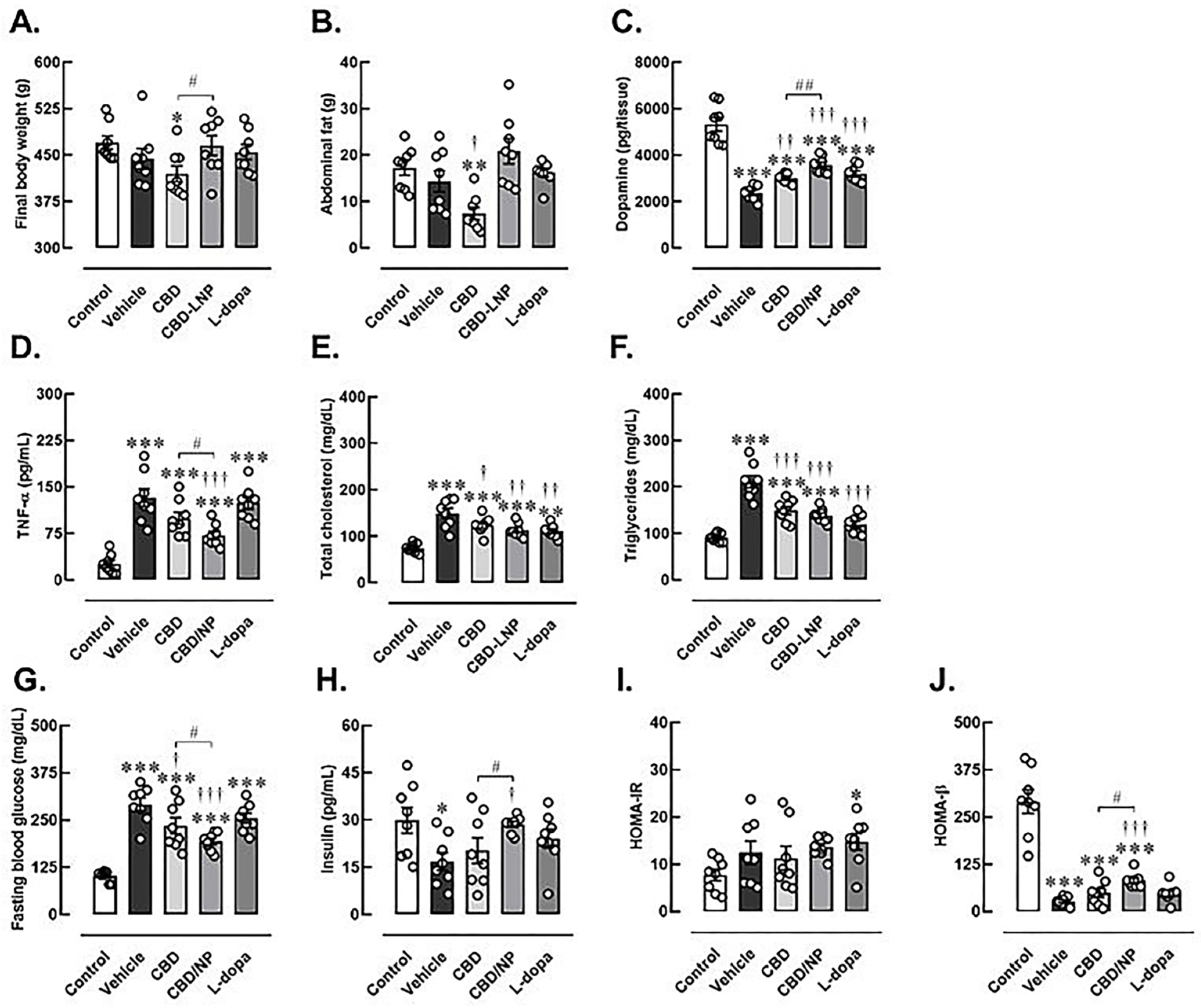
2.10. Assessment of Metabolic Parameters
2.11. Assessment of Insulin and Insulin Resistance
2.12. Assessment of Striatal Dopamine and Serum Inflammatory Cytokine Levels
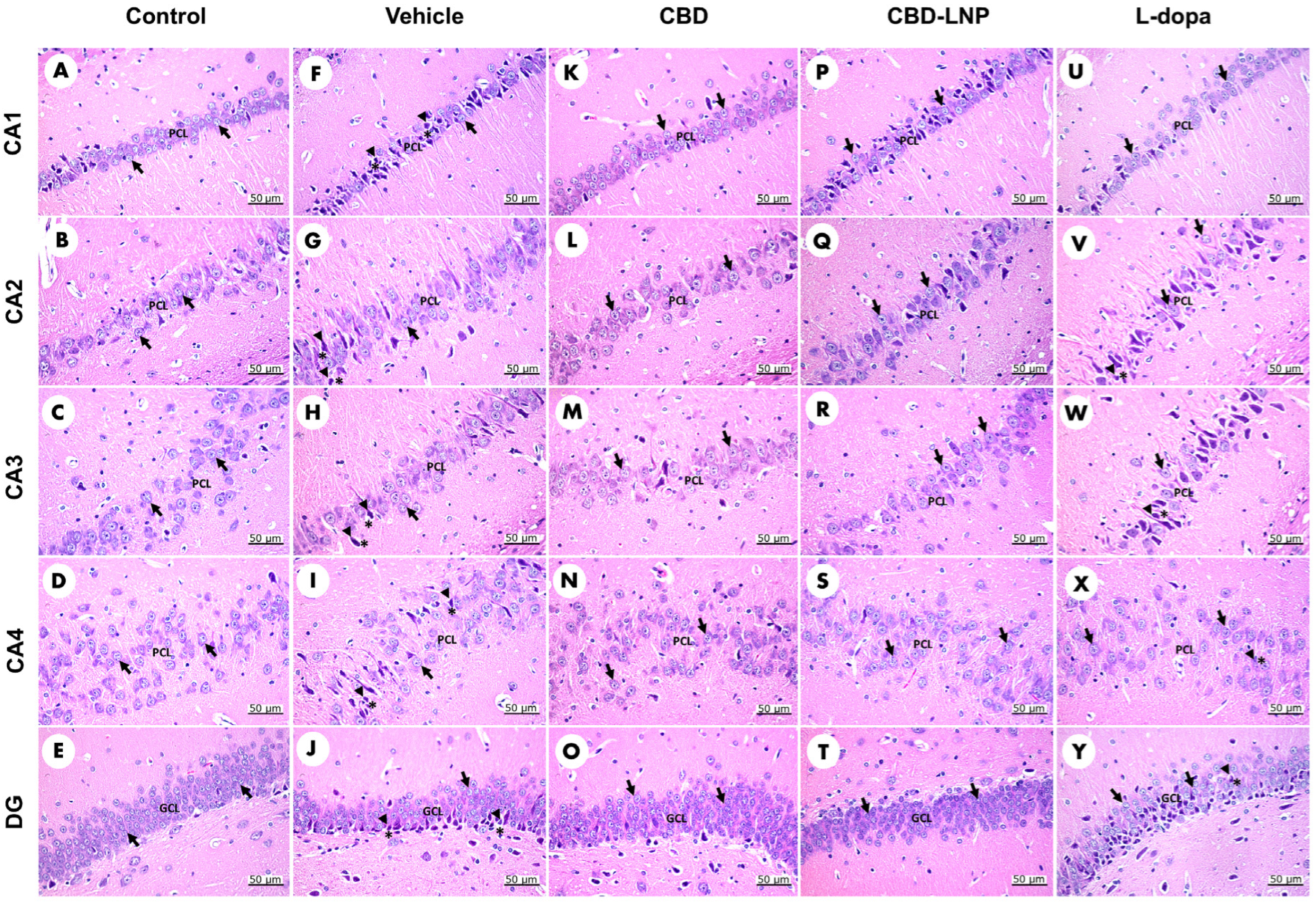
2.13. Assessment of Hippocampal Histomorphological Changes
2.14. Statistical Analysis
3. Results
3.1. Successful Preparation, Stability, and Cytotoxicity of CBD-LNP
3.2. Successful and Validation of the DP Model
3.3. The Potential Therapeutic Effects of CBD-LNP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhong, Q.; Wang, S. Association between diabetes mellitus, prediabetes and risk, disease progression of Parkinson’s disease: A systematic review and meta-analysis. Front. Aging Neurosci. 2023, 15, 1109914. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Schlenstedt, C.; Muthuraman, M.; Witt, K.; Weisser, B.; Fasano, A.; Deuschl, G. Postural control and freezing of gait in Parkinson’s disease. Park. Relat. Disord. 2016, 24, 107–112. [Google Scholar] [CrossRef]
- Bridi, J.C.; Hirth, F. Mechanisms of α-synuclein induced synaptopathy in Parkinson’s disease. Front. Neurosci. 2018, 12, 80. [Google Scholar] [CrossRef]
- Burke, R.E.; O’Malley, K. Axon degeneration in Parkinson’s disease. Exp. Neurol. 2013, 246, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Shadrina, M.I.; Slominsky, P.A.; Limborska, S.A. Molecular mechanisms of pathogenesis of Parkinson’s disease. Int. Rev. Cell Mol. Biol. 2010, 281, 229–266. [Google Scholar] [PubMed]
- Kim, E.J.; Baek, J.H.; Shin, D.J.; Park, H.M.; Lee, Y.B.; Park, K.H.; Shin, D.H.; Noh, Y.; Sung, Y.H. Correlation of sleep disturbance and cognitive impairment in patients with Parkinson’s disease. J. Mov. Disord. 2014, 7, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Castrioto, A.; Thobois, S.; Carnicella, S.; Maillet, A.; Krack, P. Emotional manifestations of PD: Neurobiological basis. Mov. Disord. 2016, 31, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Cereda, E.; Barichella, M.; Cassani, E.; Caccialanza, R.; Pezzoli, G. Clinical features of Parkinson disease when onset of diabetes came first: A case-control study. Neurology 2012, 78, 1507–1511. [Google Scholar] [CrossRef]
- Duarte, J.M.; Agostinho, P.M.; Carvalho, R.A.; Cunha, R.A. Caffeine consumption prevents diabetes-induced memory impairment and synaptotoxicity in the hippocampus of NONcZNO10/LTJ mice. PLoS ONE 2012, 7, e21899. [Google Scholar] [CrossRef] [PubMed]
- Ho, N.; Sommers, M.S.; Lucki, I. Effects of diabetes on hippocampal neurogenesis: Links to cognition and depression. Neurosci. Biobehav. Rev. 2013, 37, 1346–1362. [Google Scholar] [CrossRef] [PubMed]
- Kerti, L.; Witte, A.V.; Winkler, A.; Grittner, U.; Rujescu, D.; Flöel, A. Higher glucose levels associated with lower memory and reduced hippocampal microstructure. Neurology 2013, 81, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- di Biase, L.; Pecoraro, P.M.; Carbone, S.P.; Caminiti, M.L.; di Lazzaro, V. Levodopa-induced dyskinesias in Parkinson’s disease: An pverview on pathophysiology, clinical manifestations, therapy management strategies and future directions. J. Clin. Med. 2023, 12, 4427. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorganic Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- di Marzo, V.; Piscitelli, F.; Mechoulam, R. Cannabinoids and endocannabinoids in metabolic disorders with focus on diabetes. In Diabetes-Perspectives in Drug Therapy; Springer: Berlin/Heidelberg, Germany, 2011; Volume 203, pp. 75–104. [Google Scholar]
- Lehmann, C.; Fisher, N.B.; Tugwell, B.; Szczesniak, A.; Kelly, M.; Zhou, J. Experimental cannabidiol treatment reduces early pancreatic inflammation in type 1 diabetes. Clin. Hemorheol. Microcirc. 2016, 64, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Zorzenon, M.R.T.; Santiago, A.N.; Mori, M.A.; Piovan, S.; Jansen, C.A.; Perina Padilha, M.E.; Ciotta, S.R.; Cezar de Freitas Mathias, P.; Guimarães, F.S.; Weffort de Oliveira, R.M.; et al. Cannabidiol improves metabolic dysfunction in middle-aged diabetic rats submitted to a chronic cerebral hypoperfusion. Chem. Biol. Interact. 2019, 312, 108819. [Google Scholar] [CrossRef] [PubMed]
- Palrasu, M.; Wright, L.; Patel, M.; Leech, L.; Branch, S.; Harrelson, S.; Khan, S. Perspectives on challenges in cannabis drug delivery systems: Where are we? Med. Cannabis Cannabinoids 2022, 25, 102–119. [Google Scholar] [CrossRef] [PubMed]
- Stella, B.; Baratta, F.; Della Pepa, C.; Arpicco, S.; Gastaldi, D.; Dosio, F. Cannabinoid formulations and delivery systems: Current and future options to treat pain. Drugs 2021, 81, 1513–1557. [Google Scholar] [CrossRef] [PubMed]
- Elbassuoni, E.A.; Ahmed, R.F. Mechanism of the neuroprotective effect of GLP-1 in a rat model of Parkinson’s with pre-existing diabetes. Neurochem. Int. 2019, 131, 104583. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.; Viswanad, B.; Asrat, L.; Kaul, C.L.; Ramarao, P. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res. 2005, 52, 313–320. [Google Scholar] [CrossRef]
- Kamsrijai, U.; Wongchitrat, P.; Nopparat, C.; Satayavivad, J.; Govitrapong, P. Melatonin attenuates streptozotocin-induced Alzheimer-like features in hyperglycemic rats. Neurochem. Int. 2020, 132, 104601. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lv, H.; Cui, X.; Di, W.; Cheng, X.; Liu, J.; Tripathi, A. Exercise attenuates mitochondrial autophagy and neuronal degeneration in MPTP induced Parkinson’s disease by regulating inflammatory pathway. Folia Neuropathol. 2023, 61, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Trovato, A.E.; Comelli, F.; Giagnoni, G.; Colleoni, M. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur. J. Pharmacol. 2007, 556, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Szulc, M.; Kujawski, R.; Pacholak, A.; Poprawska, M.; Czora-Poczwardowska, K.; Geppert, B.; Mikołajczak, P.Ł. Cannabidiol as a modulator of the development of alcohol tolerance in rats. Nutrients 2023, 15, 1702. [Google Scholar] [CrossRef]
- Kääriäinen, T.M.; Käenmäki, M.; Forsberg, M.M.; Oinas, N.; Tammimäki, A.; Männistö, P.T. Unpredictable rotational responses to L-dopa in the rat model of Parkinson’s disease: The role of L-dopa pharmacokinetics and striatal dopamine depletion. Basic Clin. Pharmacol. Toxicol. 2012, 110, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Chesler, K.C.; Motz, C.T.; Bales, K.L.; Allen, R.A.; Vo, H.K.; Pardue, M.T. Voluntary oral dosing for precise experimental compound delivery in adult rats. Lab. Anim. 2022, 56, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Schmidt, W.J. Rotenone destroys dopaminergic neurons and induces parkinsonian symptoms in rats. Behav. Brain Res. 2002, 136, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Galeano, P.; Martino Adami, P.V.; Do Carmo, S.; Blanco, E.; Rotondaro, C.; Capani, F.; Castaño, E.M.; Cuello, A.C.; Morelli, L. Longitudinal analysis of the behavioral phenotype in a novel transgenic rat model of early stages of Alzheimer’s disease. Front. Behav. Neurosci. 2014, 8, 321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yi, H.; Bai, W.; Tian, X. Dynamic trajectory of multiple single-unit activity during working memory task in rats. Front. Comput. Neurosci. 2015, 9, 117. [Google Scholar] [CrossRef] [PubMed]
- Wietrzych, M.; Meziane, H.; Sutter, A.; Ghyselinck, N.; Chapman, P.F.; Chambon, P.; Krezel, W. Working memory deficits in retinoid X receptor gamma-deficient mice. Learn. Mem. 2005, 12, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Lapmanee, S.; Bhubhanil, S.; Sriwong, S.; Yuajit, C.; Wongchitrat, P.; Teerapornpuntakit, J.; Suntornsaratoon, P.; Charoenphandhu, J.; Charoenphandhu, N. Oral calcium and vitamin D supplements differentially alter exploratory, anxiety-like behaviors and memory in male rats. PLoS ONE 2023, 18, e0290106. [Google Scholar] [CrossRef] [PubMed]
- Lapmanee, S.; Bhubhanil, S.; Sriwong, S.; Khongkow, M.; Namdee, K.; Wongchitrat, P.; Pongkorpsakol, P. Venlafaxine and synbiotic attenuated learned fear-like behavior and recognition memory impairment in immobilized-stressed rats. Physiol. Pharmacol. 2023, 27, 171–181. [Google Scholar] [CrossRef]
- Haffner, S.M.; Miettinen, H.; Stern, M.P. The homeostasis model in the San Antonio Heart Study. Diabetes Care 1997, 20, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and abuse of HOMA modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Liu, K.; Li, Y.; Ma, S.; Ji, X.; Liu, L. Necrotic pyknosis is a morphologically and biochemically distinct event from apoptotic pyknosis. J. Cell Sci. 2016, 129, 3084–3090. [Google Scholar] [CrossRef] [PubMed]
- Presnell, J.K.; Schreibman, M.P. Humason’s Animal Tissue Techniques, 5th ed.; Johns Hopkins University Press: Baltimore, MD, USA, 1997. [Google Scholar]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Lapmanee, S.; Supkamonseni, N.; Bhubhanil, S.; Treesaksrisakul, N.; Sirithanakorn, C.; Khongkow, M.; Namdee, K.; Surinlert, P.; Tipbunjong, C.; Wongchitrat, P. Stress-induced changes in cognitive function and intestinal barrier integrity can be ameliorated by venlafaxine and synbiotic supplementations. PeerJ 2024, 12, e17033. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, P.; Karthivashan, G.; Park, S.Y.; Kim, J.; Choi, D.K. Microfluidization trends in the development of nanodelivery systems and applications in chronic disease treatments. Int. J. Nanomed. 2018, 13, 6109–6121. [Google Scholar] [CrossRef] [PubMed]
- Prakash, G.; Shokr, A.; Willemen, N.; Bashir, S.M.; Shin, S.R.; Hassan, S. Microfluidic fabrication of lipid nanoparticles for the delivery of nucleic acids. Adv. Drug Deliv. Rev. 2022, 184, 114197. [Google Scholar] [CrossRef]
- Vogelaar, A.; Marcotte, S.; Cheng, J.; Oluoch, B.; Zaro, J. Use of microfluidics to prepare lipid-based nanocarriers. Pharmaceutics 2023, 15, 1053. [Google Scholar] [CrossRef] [PubMed]
- Akhoond Zardini, A.; Mohebbi, M.; Farhoosh, R.; Bolurian, S. Production and characterization of nanostructured lipid carriers and solid lipid nanoparticles containing lycopene for food fortification. J. Food Sci. Technol. 2018, 55, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, J.; Wang, Y.; Gao, H.; Wei, G.; Huang, Y.; Yu, H.; Gan, Y.; Wang, Y.; Mei, L.; et al. Recent progress in drug delivery. Acta Pharm. Sin. B 2019, 9, 1145–1162. [Google Scholar] [CrossRef]
- Grifoni, L.; Vanti, G.; Donato, R.; Sacco, C.; Bilia, A.R. Promising nanocarriers to enhance solubility and bioavailability of cannabidiol for a plethora of therapeutic opportunities. Molecules 2022, 27, 6070. [Google Scholar] [CrossRef] [PubMed]
- Morakul, B.; Junyaprasert, V.B.; Sakchaisri, K.; Teeranachaideekul, V. Cannabidiol-loaded nanostructured lipid carriers (NLCs) for dermal delivery: Enhancement of photostability, cell viability, and anti-inflammatory activity. Pharmaceutics 2023, 15, 537. [Google Scholar] [CrossRef] [PubMed]
- Mohite, P.; Singh, S.; Pawar, A.; Sangale, A.; Prajapati, B.G. Lipid-based oral formulation in capsules to improve the delivery of poorly watersoluble drugs. Front. Drug Deliv. 2023, 3, 1232012. [Google Scholar] [CrossRef]
- Mehta, M.; Bui, T.A.; Yang, X.; Aksoy, Y.; Goldys, E.M.; Deng, W. Lipid-based nanoparticles for drug/gene delivery: An overview of the production techniques and difficulties encountered in their industrial development. ACS Mater. Au 2023, 3, 600–619. [Google Scholar] [CrossRef] [PubMed]
- Kolesarova, M.; Simko, P.; Urbanska, N.; Kiskova, T. Exploring the potential of cannabinoid nanodelivery systems for CNS disorders. Pharmaceutics 2023, 15, 204. [Google Scholar] [CrossRef] [PubMed]
- Assadpour, E.; Rezaei, A.; Das, S.S.; Krishna Rao, B.V.; Singh, S.K.; Kharazmi, M.S.; Jha, N.K.; Jha, S.K.; Prieto, M.A.; Jafari, S.M. Cannabidiol-loaded nanocarriers and their therapeutic applications. Pharmaceuticals 2023, 16, 487. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.I.; Kotagal, V.; Müller, M.L.; Koeppe, R.A.; Scott, P.J.; Albin, R.L.; Frey, K.A.; Petrou, M. Diabetes mellitus is independently associated with more severe cognitive impairment in Parkinson disease. Park. Relat. Disord. 2014, 20, 1394–1398. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, R.; Vasudeva, N.; Sharma, S. Acute and chronic animal models for the evaluation of anti-diabetic agents. Cardiovasc. Diabetol. 2012, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.; Coppey, L.J.; Davidson, E.P.; Yorek, M.A. Rat models of diet-induced obesity and high fat/low dose streptozotocin type 2 diabetes: Effect of reversal of high fat diet compared to treatment with enalapril or menhaden oil on glucose utilization and neuropathic endpoints. J. Diabetes Res. 2015, 2015, 307285. [Google Scholar] [CrossRef] [PubMed]
- Sergi, D.; Renaud, J.; Simola, N.; Martinoli, M.G. Diabetes, a contemporary risk for Parkinson’s disease: Epidemiological and cellular evidences. Front. Aging Neurosci. 2019, 11, 302. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.Q.; Yuan, L.; Sun, Y.; Dou, H.W.; Su, J.H.; Hou, Z.P.; Li, J.Y.; Li, W. Long-term hyperglycemia aggravates α-synuclein aggregation and dopaminergic neuronal loss in a Parkinson’s disease mouse model. Transl. Neurodegener. 2022, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Khalili, D.; Khayamzadeh, M.; Kohansal, K.; Ahanchi, N.S.; Hasheminia, M.; Hadaegh, F.; Tohidi, M.; Azizi, F.; Habibi-Moeini, A.S. Are HOMA-IR and HOMA-B good predictors for diabetes and pre-diabetes subtypes? BMC Endocr. Disord. 2023, 23, 39. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.; St-Amour, I.; Vandal, M.; Julien, P.; Cicchetti, F.; Calon, F. High-fat diet exacerbates MPTP-induced dopaminergic degeneration in mice. Neurobiol. Dis. 2012, 45, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s disease: Mechanisms and therapeutic implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef] [PubMed]
- Lisco, G.; de Tullio, A.; Iovino, M.; Disoteo, O.; Guastamacchia, E.; Giagulli, V.A.; Triggiani, V. Dopamine in the regulation of glucose homeostasis, pathogenesis of type 2 diabetes, and chronic conditions of impaired dopamine activity/metabolism: Implication for pathophysiological and therapeutic purposes. Biomedicines 2023, 11, 2993. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, E.K.F.F.; Costa, A.B.; Salla, D.H.; Silva, M.R.D.; Mendes, T.F.; Silva, L.E.D.; Turatti, C.D.R.; Bitencourt, R.M.; Rezin, G.T. Cannabis sativa as a treatment for obesity: From anti-inflammatory indirect support to a promising metabolic re-establishment target. Cannabis Cannabinoid Res. 2022, 7, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Ramlugon, S.; Levendal, R.A.; Frost, C.L. Effect of oral cannabis administration on the fat depots of obese and streptozotocin-induced diabetic rats. Phytother. Res. 2023, 37, 1806–1822. [Google Scholar] [CrossRef] [PubMed]
- El-Hammadi, M.M.; Bourgon, J.V.; Dominguez, E.B.; Facorro, B.C.; Pereira, I.S.; Banderas, L.M. CBD-loaded NPs increases glucose uptake and attenuates palmitate-induced lipid accumulation in human HepG2 hepatocytes. RESCIFAR Rev. Española de Cienc. Farm. 2021, 2, 200–201. [Google Scholar]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.; Rodriguez-Torres, M.D.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and anti-inflammatory properties of cannabidiol. Antioxidants 2019, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Peres, F.F.; Levin, R.; Suiama, M.A.; Diana, M.C.; Gouvêa, D.A.; Almeida, V.; Santo, C.M.; Lungato, L.; Zuardi, A.W.; Hallak, J.E.; et al. Cannabidiol prevents motor and cognitive impairments induced by reserpine in rats. Front. Pharmacol. 2016, 7, 343. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Anwar, F.; Saleem, U.; Shahzadi, I.; Ahmad, B.; Mir, A.; Ismail, T. Parkinsonism attenuation by antihistamines via downregulating the oxidative stress, histamine, and inflammation. ACS Omega 2022, 7, 14772–14783. [Google Scholar] [CrossRef] [PubMed]
- Eskow Jaunarajs, K.L.; Dupre, K.B.; Ostock, C.Y.; Button, T.; Deak, T.; Bishop, C. Behavioral and neurochemical effects of chronic L-DOPA treatment on nonmotor sequelae in the hemiparkinsonian rat. Behav. Pharmacol. 2010, 21, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Grayson, B.; Leger, M.; Piercy, C.; Adamson, L.; Harte, M.; Neill, J.C. Assessment of disease-related cognitive impairments using the novel object recognition (NOR) task in rodents. Behav. Brain Res. 2015, 285, 176–193. [Google Scholar] [CrossRef]
- Cheong, J.L.Y.; de Pablo-Fernandez, E.; Foltynie, T.; Noyce, A.J. The association between type 2 diabetes mellitus and Parkinson’s disease. J. Park. Dis. 2020, 10, 775–789. [Google Scholar] [CrossRef]
- Ruiz-Pozo, V.A.; Tamayo-Trujillo, R.; Cadena-Ullauri, S.; Frias-Toral, E.; Guevara-Ramírez, P.; Paz-Cruz, E.; Chapela, S.; Montalván, M.; Morales-López, T.; Simancas-Racines, D.; et al. The molecular mechanisms of the relationship between insulin resistance and Parkinson’s disease pathogenesis. Nutrients 2023, 15, 3585. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, J.Y.; Seo, J.; Choi, I.S. Neuroprotective effect of cannabidiol against hydrogen peroxide in hippocampal neuron culture. Cannabis Cannabinoid Res. 2021, 6, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, S.; Kolishetti, N.; Arias, A.Y.; Vashist, A.; Nair, M. Cannabidiol for neurodegenerative disorders: A comprehensive review. Front. Pharmacol. 2022, 13, 989717. [Google Scholar] [CrossRef] [PubMed]
- Beckers, M.; Bloem, B.R.; Verbeek, M.M. Mechanisms of peripheral levodopa resistance in Parkinson’s disease. NPJ Park. Dis. 2022, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Chohan, H.; Senkevich, K.; Patel, R.K.; Bestwick, J.P.; Jacobs, B.M.; Bandres Ciga, S.; Gan-Or, Z.; Noyce, A.J. Type 2 diabetes as a determinant of Parkinson’s disease risk and progression. Mov. Disord. 2021, 36, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xie, J.; Li, Y.; Liu, H.; Liu, C.; Kan, D.; Geng, X.; Wei, S. Levodopa affects spike and local field synchronisation in the pedunculopontine nucleus of a rat model of Parkinson’s disease. Aging 2021, 13, 7314–7329. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrodynamic Size (nm) | Day 0 | Day 30 | ||
| Kept at 4 °C | Kept at 25 °C | Kept at 45 °C | ||
| Blank-LNP | 169.93 ± 34.41 | 163.50 ± 3.78 | 151.27 ± 2.15 | 153.80 ± 0.44 |
| CBD-LNP | 156.33 ± 1.24 | 158.17 ± 1.06 | 167.87 ± 0.81 | 166.83 ± 5.37 |
| Polydispersity | Day 0 | Day 30 | ||
| Kept at 4 °C | Kept at 25 °C | Kept at 45 °C | ||
| Blank-LNP | 0.41 ± 0.02 | 0.18 ± 0.02 | 0.10 ± 0.03 | 0.07 ± 0.03 |
| CBD-LNP | 0.11 ± 0.10 | 0.12 ± 0.02 | 0.10 ± 0.03 | 0.13 ± 0.02 |
| Zeta Potential (mV) | Day 0 | Day 30 | ||
| Kept at 4 °C | Kept at 25 °C | Kept at 45 °C | ||
| Blank-LNP | −3.53 ± 0.15 | −15.28 ± 0.09 | −17.89 ± 0.84 | −13.04 ± 0.63 |
| CBD-LNP | −16.57 ± 0.04 | −17.04 ± 0.90 | −14.47 ± 2.08 | −19.55 ± 0.70 |
| Parameters | Control Group | DP Group | |||
|---|---|---|---|---|---|
| Vehicle | Vehicle | CBD | CBD-LNP | L-Dopa | |
| Pyknotic index of the CA1 (%) | 7.70 ± 3.25 | 22.59 ± 12.91 ** | 11.50 ± 4.30 † | 9.78 ± 5.01 †† | 7.86 ± 0.84 †† |
| Pyknotic index of the CA2 (%) | 6.13 ± 1.67 | 40.32 ± 14.39 *** | 8.86 ± 3.94 †† | 12.45 ± 5.61 †† | 34.39 ± 10.39 ** |
| Pyknotic index of the CA3 (%) | 6.78 ± 0.97 | 38.96 ± 10.41 ** | 14.00 ± 0.86 † | 6.12 ± 2.41 †† | 14.21 ± 4.48 † |
| Pyknotic index of the CA4 (%) | 7.84 ± 3.82 | 55.95 ± 9.98 ** | 8.22 ± 2.24 †† | 14.12 ± 7.04 †† | 21.61 ± 15.31 † |
| Pyknotic index of the DG (%) | 6.39 ± 3.76 | 25.83 ± 8.51 ** | 10.04 ± 0.84 † | 11.05 ± 4.90 † | 14.51 ± 5.73 † |
| Thickness of the pyramidal cell layer in the CA1 (µm) | 47.28 ± 1.19 | 31.99 ± 1.72 *** | 47.47 ± 1.95 ††† | 48.51 ± 1.45 ††† | 45.8 ± 1.26 ††† |
| Thickness of the pyramidal cell layer in the CA2 (µm) | 55.69 ± 1.98 | 47.67 ± 2.27 *** | 51.28 ± 1.52 † | 52.89 ± 1.83 †† | 48.48 ± 1.44 * |
| Thickness of the pyramidal cell layer in the CA3 (µm) | 57.47 ± 1.15 | 39.97 ± 1.30 *** | 51.19 ± 1.32 *††† | 54.45 ± 1.71 ††† | 51.72 ± 1.36 *††† |
| Thickness of the pyramidal cell layer in the CA4 (µm) | 93.29 ± 3.33 | 70.16 ± 2.38 *** | 82.63 ± 3.56 † | 88.23 ± 3.42 ††† | 89.10 ± 2.17 ††† |
| Thickness of the granular cell layer in the DG (µm) | 64.07 ± 1.91 | 50.14 ± 1.73 *** | 58.36 ± 1.08 †† | 61.08 ± 1.62 ††† | 62.18 ± 1.15 ††† |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lapmanee, S.; Bhubhanil, S.; Wongchitrat, P.; Charoenphon, N.; Inchan, A.; Ngernsutivorakul, T.; Dechbumroong, P.; Khongkow, M.; Namdee, K. Assessing the Safety and Therapeutic Efficacy of Cannabidiol Lipid Nanoparticles in Alleviating Metabolic and Memory Impairments and Hippocampal Histopathological Changes in Diabetic Parkinson’s Rats. Pharmaceutics 2024, 16, 514. https://doi.org/10.3390/pharmaceutics16040514
Lapmanee S, Bhubhanil S, Wongchitrat P, Charoenphon N, Inchan A, Ngernsutivorakul T, Dechbumroong P, Khongkow M, Namdee K. Assessing the Safety and Therapeutic Efficacy of Cannabidiol Lipid Nanoparticles in Alleviating Metabolic and Memory Impairments and Hippocampal Histopathological Changes in Diabetic Parkinson’s Rats. Pharmaceutics. 2024; 16(4):514. https://doi.org/10.3390/pharmaceutics16040514
Chicago/Turabian StyleLapmanee, Sarawut, Sakkarin Bhubhanil, Prapimpun Wongchitrat, Natthawut Charoenphon, Anjaree Inchan, Thitaphat Ngernsutivorakul, Piroonrat Dechbumroong, Mattaka Khongkow, and Katawut Namdee. 2024. "Assessing the Safety and Therapeutic Efficacy of Cannabidiol Lipid Nanoparticles in Alleviating Metabolic and Memory Impairments and Hippocampal Histopathological Changes in Diabetic Parkinson’s Rats" Pharmaceutics 16, no. 4: 514. https://doi.org/10.3390/pharmaceutics16040514