
Decreased Gene Expression of Antiangiogenic Factors in Endometrial Cancer: qPCR Analysis and Machine Learning Modelling
 , , , and
, , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of Publicly Available Datasets
2.2. Validation Study on Endometrial Cancer Tissue
2.2.1. Endometrial Tissue
2.2.2. RNA Isolation and Reverse Transcription
2.2.3. Quantitative Real-Time PCR
2.2.4. Statistics
2.3. Machine Learning Modelling
- TCGA and study data were normalised for merging;
- TCGA data (22 samples) were merged with 22 stratified randomly selected samples of study data; the remaining 14 samples were assigned to the test dataset;
- An automated machine learning (AutoML) approach was used to create the models on the training dataset;
- The models were tested on the test dataset.
2.3.1. Merging and Normalisation
- The best fitting distribution for data was empirically selected by trying to fit the data to one of the common standard distributions (Normal, Log-normal, Poisson, Beta, Gamma).
- For distributions that require positive data, the data were right-shifted to ensure that the smallest value was positive.
- The best fitting distribution for most columns (Gamma) was then fitted for all columns, and distribution parameters were calculated, together with the correlation coefficient, significance, and estimated lower and upper bounds at the 95% confidence level using the MATLAB “corrcoef” function. It is worth noting at this point that the Gamma distribution has previously been linked to gene expression in multiple studies [28].
- The original values were then transformed to the value of the cumulative distribution function (CDF) of the fitted distribution at the original value, thus obtaining a value between 0 and 1, indicating the relative (expected) ratio of the population with a value lower than the original value [29].
- The training dataset was created by taking all 22 normalised samples from the TCGA dataset and combining them with 22 randomly selected samples from the study dataset, where a stratified random sampling approach was used to ensure the final dataset had a balanced distribution of the output variable (EC grade).
- The remaining samples from the study dataset represented the test dataset.
2.3.2. Modelling and Testing
- A model utilising the data combining the tumour tissue, adjacent tissue data, and calculated ratios between the tumour tissue and adjacent tissue measurements;
- A model utilising only the tumour tissue data;
- A model utilising only the adjacent tissue data.
3. Results
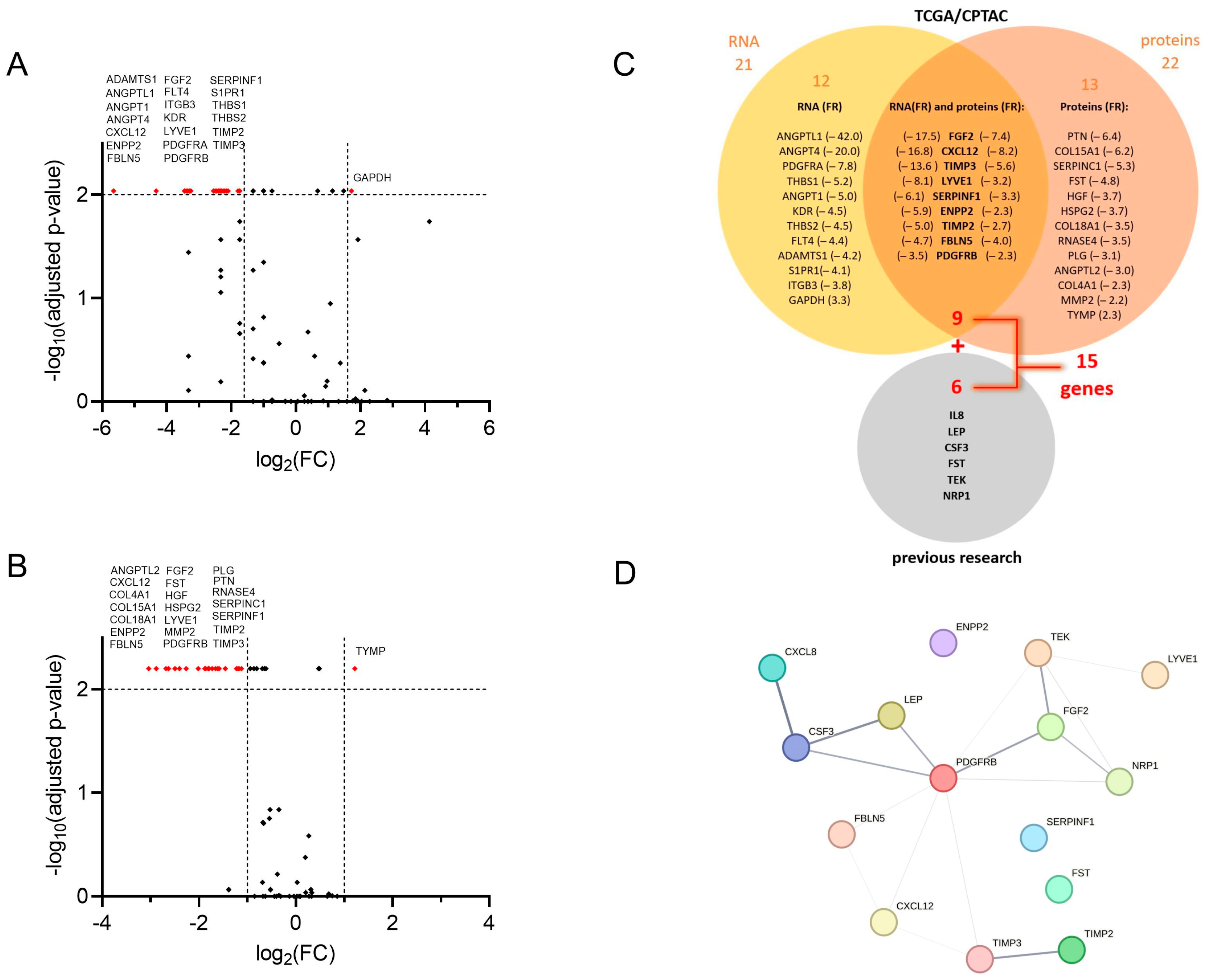
3.1. Public Databases Examination Revealed Twenty-One AF-Encoding Genes and Twenty-Two AF Proteins That Fulfilled Selection Criteria; Nine Gene/Protein Pairs Were in the Intersection
3.2. Validation of Findings on the Clinical Cohort
3.2.1. Clinical Characteristics of Enrolled Patients
3.2.2. Thirteen Genes Encoding AFs Are Differentially Expressed in Tumour Tissue Compared to Adjacent Control Tissue in EC Patients
3.2.3. Relationships of Gene Expression with Clinical Characteristics
In Early Stages and Lower Grades of EC, but Not in More Advanced or Aggressive Forms of EC, Genes for AFs Tend to Be Differentially Expressed in Tumour Tissue Compared to Adjacent Control Tissue
Genes for AFs Are Differentially Expressed between Tumour and Adjacent Control Tissue Only in Patients without DMI or LVI
There Is Much Broader Angiogenesis-Related Gene Involvement in Postmenopausal Women with EC than in Women of Reproductive Age
3.2.4. Co-Expression Patterns of the Genes: Higher Number of Strong Correlations Was Identified in EC Patients with Present LVI
3.3. Machine Learning Modelling Succeeded in Creating a Relatively Robust EC-Grade Prediction Model Based on the Tumour Gene Expressions
3.3.1. Data Normalisation Results
3.3.2. Comparison of Training and Test Datasets
3.3.3. Modelling Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crosbie, E.J.; Kitson, S.J.; McAlpine, J.N.; Mukhopadhyay, A.; Powell, M.E.; Singh, N. Endometrial cancer. Lancet 2022, 399, 1412–1428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gong, T.T.; Liu, F.H.; Jiang, Y.T.; Sun, H.; Ma, X.X.; Zhao, Y.H.; Wu, Q.J. Global, Regional, and National Burden of Endometrial Cancer, 1990–2017: Results from the Global Burden of Disease Study, 2017. Front. Oncol. 2019, 9, 1440. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Cheung, M.K.; Shin, J.Y.; Husain, A.; Teng, N.N.; Berek, J.S.; Kapp, D.S.; Osann, K.; Chan, J.K. Prognostic factors for uterine cancer in reproductive-aged women. Obstet. Gynecol. 2007, 109, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Wilczyński, M.; Danielska, J.; Wilczyński, J. An update of the classical Bokhman’s dualistic model of endometrial cancer. Przegląd Menopauzalny 2016, 15, 63–68. [Google Scholar] [CrossRef]
- Murali, R.; Soslow, R.A.; Weigelt, B. Classification of endometrial carcinoma: More than two types. Lancet Oncol. 2014, 15, e268–e278. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network; Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar]
- Concin, N.; Matias-Guiu, X.; Vergote, I.; Cibula, D.; Mirza, M.R.; Marnitz, S.; Ledermann, J.; Bosse, T.; Chargari, C.; Fagotti, A.; et al. ESGO/ESTRO/ESP guidelines for the management of patients with endometrial carcinoma. Int. J. Gynecol. Cancer 2021, 31, 12–39. [Google Scholar] [CrossRef]
- Rodolakis, A.; Scambia, G.; Planchamp, F.; Acien, M.; Di Spiezio Sardo, A.; Farrugia, M.; Grynberg, M.; Pakiž, M.; Pavlakis, K.; Vermeulen, N.; et al. ESGO/ESHRE/ESGE Guidelines for the fertility-sparing treatment of patients with endometrial carcinoma. Int. J. Gynecol. Cancer 2023, 33, 208–222. [Google Scholar] [CrossRef]
- Helpman, L.; Kupets, R.; Covens, A.; Saad, R.S.; Khalifa, M.A.; Ismiil, N.; Ghorab, Z.; Dubé, V.; Nofech-Mozes, S. Assessment of endometrial sampling as a predictor of final surgical pathology in endometrial cancer. Br. J. Cancer 2014, 110, 609–615. [Google Scholar] [CrossRef]
- Risau, W. Mechanisms of angiogenesis. Nature 1997, 386, 671–674. [Google Scholar] [CrossRef]
- Abulafia, O.; Sherer, D.M. Angiogenesis of the endometrium. Obstet. Gynecol. 1999, 94, 148–153. [Google Scholar] [PubMed]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Roškar, L.; Roškar, I.; Rižner, T.L.; Smrkolj, Š. Diagnostic and Therapeutic Values of Angiogenic Factors in Endometrial Cancer. Biomolecules 2021, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Roškar, L.; Pušić, M.; Roškar, I.; Kokol, M.; Pirš, B.; Smrkolj, Š.; Rižner, T.L. Models including preoperative plasma levels of angiogenic factors, leptin and IL-8 as potential biomarkers of endometrial cancer. Front. Oncol. 2022, 12, 972131. [Google Scholar] [CrossRef] [PubMed]
- Karkia, R.; Wali, S.; Payne, A.; Karteris, E.; Chatterjee, J. Diagnostic Accuracy of Liquid Biomarkers for the Non-Invasive Diagnosis of Endometrial Cancer: A Systematic Review and Meta-Analysis. Cancers 2022, 14, 4666. [Google Scholar] [CrossRef] [PubMed]
- Obradović, D.D.; Milić, N.M.; Miladinović, N.; McClements, L.; Oprić, D.M. Loss of Expression of Antiangiogenic Protein FKBPL in Endometrioid Endometrial Carcinoma: Implications for Clinical Practice. Medicina 2022, 58, 1330. [Google Scholar] [CrossRef] [PubMed]
- Kokol, P.; Kokol, M.; Zagoranski, S. Machine learning on small size samples: A synthetic knowledge synthesis. Sci. Prog. 2022, 105, 00368504211029777. [Google Scholar] [CrossRef]
- Roškar, L.; Klančič, T.; Knific, T.; Rižner, T.L.; Smrkolj, Š. Tie-2, G-CSF, and Leptin as Promising Diagnostic Biomarkers for Endometrial Cancer: A Pilot Study. J. Clin. Med. 2021, 10, 765. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Mills Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Dou, Y.; Kawaler, E.A.; Cui Zhou, D.; Gritsenko, M.A.; Huang, C.; Blumenberg, L.; Karpova, A.; Petyuk, V.A.; Savage, S.R.; Satpathy, S.; et al. Proteogenomic Characterization of Endometrial Carcinoma. Cell 2020, 180, 729–748. [Google Scholar] [CrossRef] [PubMed]
- Pavlič, R.; Vidic, S.; Anko, M.; Knific, T.; Büdefeld, T.; Marton, K.; Sinreih, M.; Poschner, S.; Jäger, W.; Frković-Grazio, S.; et al. Altered Profile of E1-S Transporters in Endometrial Cancer: Lower Protein Levels of ABCG2 and OSTβ and Up-Regulation of SLCO1B3 Expression. Int. J. Mol. Sci. 2021, 22, 3819. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Zhou, Y.-H. Leveraging Scheme for Cross-Study Microbiome Machine Learning Prediction and Feature Evaluations. Bioengineering 2023, 10, 231. [Google Scholar] [CrossRef]
- Lee, A.J.; Park, Y.S.; Doing, G.; Hogan, D.A.; Greene, C.S. Correcting for experiment-specific variability in expression compendia can remove underlying signals. GigaScience 2020, 9, giaa117. [Google Scholar] [CrossRef]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Friedman, N.; Cai, L.; Xie, X.S. Linking stochastic dynamics to population distribution: An analytical framework of gene expression. Phys. Rev. Lett. 2006, 97, 168302. [Google Scholar] [CrossRef]
- Deisenroth, M.P.; Faisal, A.A.; Ong, C.S. Mathematics for Machine Learning; Cambridge University Press: London, UK, 2021. [Google Scholar]
- Piońska, A.; Pioński, P. MLJAR: State-of-the-art Automated Machine Learning Framework for Tabular Data. Version 0.10.3; MLJAR: Bialystok, Poland, 2021. [Google Scholar]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn Res. 2011, 12, 2825–2830. [Google Scholar]
- The Pandas Development Team. Pandas-Dev/Pandas: Pandas. Zenodo 2020, 21, 1–9. [Google Scholar]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605. [Google Scholar] [CrossRef] [PubMed]
- Amant, F.; Mirza, M.R.; Koskas, M.; Creutzberg, C.L. Cancer of the corpus uteri. Int. J. Gynecol. Obstet. 2018, 143, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Voss, M.A.; Ganesan, R.; Ludeman, L.; McCarthy, K.; Gornall, R.; Schaller, G.; Wei, W.; Sundar, S. Should grade 3 endometrioid endometrial carcinoma be considered a type 2 cancer-A clinical and pathological evaluation. Gynecol. Oncol. 2012, 124, 15–20. [Google Scholar] [CrossRef]
- Setiawan, V.W.; Yang, H.P.; Pike, M.C.; McCann, S.E.; Yu, H.; Xiang, Y.B.; Wolk, A.; Wentzensen, N.; Weiss, N.S.; Webb, P.M.; et al. Type I and II endometrial cancers: Have they different risk factors? J. Clin. Oncol. 2013, 31, 2607–2618. [Google Scholar] [CrossRef]
- Yetkin-Arik, B.; Kastelein, A.W.; Klaassen, I.; Jansen, C.H.J.R.; Latul, Y.P.; Vittori, M.; Biri, A.; Kahraman, K.; Griffioen, A.W.; Amant, F.; et al. Angiogenesis in gynecological cancers and the options for anti-angiogenesis therapy. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188446. [Google Scholar] [CrossRef]
- Demir, R.; Yaba, A.; Huppertz, B. Vasculogenesis and angiogenesis in the endometrium during menstrual cycle and implantation. Acta Histochem. 2010, 112, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Grzechocińska, B.; Dąbrowski, F.; Cyganek, A.; Panek, G.; Wielgoś, M. The role of metalloproteinases in endometrial remodelling during menstrual cycle. Ginekol. Pol. 2017, 88, 337–342. [Google Scholar] [CrossRef]
- Sun, X.; Cheng, G.; Hao, M.; Zheng, J.; Zhou, X.; Zhang, J.; Taichman, R.S.; Pienta, K.J.; Wang, J. CXCL12/CXCR4/CXCR7 chemokine axis and cancer progression. Cancer Metastasis Rev. 2010, 29, 709–722. [Google Scholar] [CrossRef]
- Liu, P.; Long, P.; Huang, Y.; Sun, F.; Wang, Z. CXCL12/CXCR4 axis induces proliferation and invasion in human endometrial cancer. Am. J. Transl. Res. 2016, 8, 1719. [Google Scholar]
- Gelmini, S.; Mangoni, M.; Castiglione, F.; Beltrami, C.; Pieralli, A.; Andersson, K.L.; Fambrini, M.; Taddei, G.L.; Serio, M.; Orlando, C. The CXCR4/CXCL12 axis in endometrial cancer. Clin. Exp. Metastasis 2009, 26, 261–268. [Google Scholar] [CrossRef]
- Sahoo, S.; Zhang, X.; Hondermarck, H.; Tanwar, P. The Emerging Role of the Microenvironment in Endometrial Cancer. Cancers 2018, 10, 408. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Wang, Y.; Liu, J.; Mok, S.C.; Xue, F.; Zhang, W. CXCL12/CXCR4: A symbiotic bridge linking cancer cells and their stromal neighbors in oncogenic communication networks. Oncogene 2015, 35, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Felix, A.S.; Weissfeld, J.; Edwards, R.; Linkov, F. Future directions in the field of endometrial cancer research: The need to investigate the tumor microenvironment. Eur. J. Gynaecol. Oncol. 2010, 31, 139–144. [Google Scholar] [PubMed]
- Jackson, H.W.; Defamie, V.; Waterhouse, P.; Khokha, R. TIMPs: Versatile extracellular regulators in cancer. Nat. Rev. Cancer 2017, 17, 38–53. [Google Scholar] [CrossRef]
- Graesslin, O.; Cortez, A.; Fauvet, R.; Lorenzato, M.; Birembaut, P.; Daraï, E. Metalloproteinase-2, -7 and -9 and tissue inhibitor of metalloproteinase-1 and -2 expression in normal, hyperplastic and neoplastic endometrium: A clinical-pathological correlation study. Ann. Oncol. 2006, 17, 637–645. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, W.; Zhang, S.; Zhang, Y.; Liu, P.; Li, X.; Zhi, W.; Yang, D.; Li, M.; Lu, Y. Pan-cancer analysis of osteogenesis imperfecta causing gene SERPINF1. Intractable Rare Dis. Res. 2022, 11, 15–24. [Google Scholar] [CrossRef]
- Brook, N.; Brook, E.; Dass, C.R.; Chan, A.; Dharmarajan, A. Pigment Epithelium-Derived Factor and Sex Hormone-Responsive Cancers. Cancers 2020, 12, 3483. [Google Scholar] [CrossRef]
- Risques, R.A.; Kennedy, S.R. Aging and the rise of somatic cancer-associated mutations in normal tissues. PLoS Genet. 2018, 14, e1007108. [Google Scholar] [CrossRef]
- Milholland, B.; Auton, A.; Suh, Y.; Vijg, J. Age-related somatic mutations in the cancer genome. Oncotarget 2015, 6, 24627–24635. [Google Scholar] [CrossRef]
- Catasus, L.; Pons, C.; Muñoz, J.; Espinosa, I.; Prat, J. Promoter hypermethylation contributes to TIMP3 down-regulation in high stage endometrioid endometrial carcinomas. Histopathology 2013, 62, 632–641. [Google Scholar] [CrossRef]
- Shaw, E.; Farris, M.; McNeil, J.; Friedenreich, C. Obesity and Endometrial Cancer. In Obesity and Cancer. Recent Results in Cancer Research; Pischon, T., Nimptsch, K., Eds.; Springer: Cham, Switzerland, 2016; pp. 107–136. [Google Scholar]
- Reeves, G.K.; Pirie, K.; Beral, V.; Green, J.; Spencer, E.; Bull, D. Cancer incidence and mortality in relation to body mass index in the Million Women Study: Cohort study. Br. Med. J. 2007, 335, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Onstad, M.A.; Schmandt, R.E.; Lu, K.H. Addressing the role of obesity in endometrial cancer risk, prevention, and treatment. J. Clin. Oncol. 2016, 34, 4225–4230. [Google Scholar] [CrossRef] [PubMed]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-mass index and incidence of cancer: A systematic review and meta-analysis of prospective observational studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Daley-Brown, D.; Oprea-Ilies, G.M.; Lee, R.; Pattillo, R.; Gonzalez-Perez, R.R. Molecular cues on obesity signals, tumor markers and endometrial cancer. Horm. Mol. Biol. Clin Investig. 2015, 21, 89–106. [Google Scholar] [CrossRef]
- Mantzoros, C.S.; Magkos, F.; Brinkoetter, M.; Sienkiewicz, E.; Dardeno, T.A.; Kim, S.Y.; Hamnvik, O.P.; Koniaris, A. Leptin in human physiology and pathophysiology. Am. J. Physiol. Metab. 2011, 301, E567–E584. [Google Scholar] [CrossRef]
- Bouloumié, A.; Drexler, H.C.; Lafontan, M.; Busse, R. Leptin, the product of Ob gene, promotes angiogenesis. Circ. Res. 1998, 83, 1059–1066. [Google Scholar] [CrossRef]
- Sierra-Honigmann, M.R.; Nath, A.K.; Murakami, C.; García-Cardeña, G.; Papapetropoulos, A.; Sessa, W.C.; Madge, L.A.; Schechner, J.S.; Schwabb, M.B.; Polverini, P.J.; et al. Biological action of leptin as an angiogenic factor. Science 1998, 281, 1683–1686. [Google Scholar] [CrossRef]
- Ellis, P.E.; Barron, G.A.; Bermano, G. Adipocytokines and their relationship to endometrial cancer risk: A systematic review and meta-analysis. Gynecol. Oncol. 2020, 158, 507–516. [Google Scholar] [CrossRef]
- Wang, P.P.; He, X.Y.; Wang, R.; Wang, Z.; Wang, Y.G. High leptin level is an independent risk factor of endometrial cancer: A meta-analysis. Cell Physiol. Biochem. 2014, 34, 1477–1484. [Google Scholar] [CrossRef]
- Hazelwood, E.; Sanderson, E.; Tan, V.Y.; Ruth, K.S.; Frayling, T.M.; Dimou, N.; Gunter, M.J.; Dossus, L.; Newton, C.; Ryan, N.; et al. Identifying molecular mediators of the relationship between body mass index and endometrial cancer risk: A Mendelian randomization analysis. BMC Med. 2022, 20, 125. [Google Scholar] [CrossRef]
- Boroń, D.; Nowakowski, R.; Grabarek, B.O.; Zmarzły, N.; Opławski, M. Expression pattern of leptin and its receptors in endometrioid endometrial cancer. J. Clin. Med. 2021, 10, 2787. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Park, H.S.; Kawada, T.; Kim, J.H.; Lim, D.; Hubbard, N.E.; Kwon, B.S.; Erickson, K.L.; Yu, R. Circulating levels of MCP-1 and IL-8 are elevated in human obese subjects and associated with obesity-related parameters. Int. J. Obes. 2006, 30, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Ciortea, R.; Mihu, D.; Mihu, C.M. Association between visceral fat, IL-8 and endometrial cancer. Anticancer Res. 2014, 34, 379–383. [Google Scholar] [PubMed]
- Koch, A.E.; Polverini, P.J.; Kunkel, S.L.; Harlow, L.A.; DiPietro, L.A.; Elner, V.M.; Elner, S.G.; Strieter, R.M. Interleukin-8 as a Macrophage-Derived Mediator of Angiogenesis. Science 1992, 258, 1798–1801. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, J.; Aoki, I.; Khatun, S.; Toyoki, H.; Tamaya, T. Clinical implications of expression of interleukin-8 related to myometrial invasion with angiogenesis in uterine endometrial cancers. Ann. Oncol. 2002, 13, 430–434. [Google Scholar] [CrossRef]
- Kotowicz, B.; Fuksiewicz, M.; Jonska-Gmyrek, J.; Berezowska, A.; Radziszewski, J.; Bidzinski, M.; Kowalska, M. Clinical significance of pretreatment serum levels of VEGF and its receptors, IL- 8, and their prognostic value in type I and II endometrial cancer patients. PLoS ONE 2017, 12, e0184576. [Google Scholar] [CrossRef]
- Raudenska, M.; Sztalmachova, M.; Gumulec, J.; Fojtu, M.; Polanska, H.; Balvan, J.; Feith, M.; Binkova, H.; Horakova, Z.; Kostrica, R.; et al. Prognostic significance of the tumour-adjacent tissue in head and neck cancers. Tumor. Biol. 2015, 36, 9929–9939. [Google Scholar] [CrossRef]
- Kulinczak, M.; Sromek, M.; Panek, G.; Zakrzewska, K.; Lotocka, R.; Szafron, L.M.; Chechlinska, M.; Siwicki, J.K. Endometrial Cancer-Adjacent Tissues Express Higher Levels of Cancer-Promoting Genes than the Matched Tumors. Genes 2022, 13, 1611. [Google Scholar] [CrossRef]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive analysis of normal adjacent to tumor transcriptomes. Nat. Commun. 2017, 8, 1077. [Google Scholar] [CrossRef]
- Huang, X.; Stern, D.F.; Zhao, H. Transcriptional Profiles from Paired Normal Samples Offer Complementary Information on Cancer Patient Survival—Evidence from TCGA Pan-Cancer Data. Sci. Rep. 2016, 6, 20567. [Google Scholar] [CrossRef]
- Frost, H.R. Analyzing cancer gene expression data through the lens of normal tissue-specificity. PLoS Comput. Biol. 2021, 17, e1009085. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Downloaded from | Detail | Samples | References |
|---|---|---|---|---|
| cBioPortal | https://www.cbioportal.org/ accessed on 4 July 2022 | TCGA Pan-Cancer study | n (T) = up to 527 | [19,20] |
| UCSC Xena | https://xena.ucsc.edu/ accessed on 8 July 2022 | GDC TCGA Endometrioid Cancer (UCEC) study (TCGA data uniformly reanalysed at GDC by UCSC Xena group using the latest Human Genome Assembly hg38) | n (T) = up to 548; n (TA) = up to 35; n (paired samples) = up to 23. | [21] |
| NCI PDC server | https://proteomic.datacommons.cancer.gov/pdc/ accessed on 5 July 2022 | CPTAC UCEC Discovery Study—Proteome, PDC ID: PDC000125, study ID: c935c587-0cd1-11e9-a064-0a9c39d33490 | n (T) = up to 95; n (TA) = up to 25; n (paired samples) = up to 24. | [22] |
| Sample | Age | Menopausal Status | Histological Type/Grade | FIGO Stage | Gradus HG/LG | Depth of Myometrial Invasion | Lymphovascular Invasion |
|---|---|---|---|---|---|---|---|
| 5 | 39 | premenopausal | dedifferentiated | IB | HG | >50% | yes |
| 7 | 50 | premenopausal | endometrioid G1 | IB | LG | no | no |
| 8 | 83 | postmenopausal | dedifferentiated | IB | HG | >50% | no |
| 9 | 41 | premenopausal | endometrioid G1 | IA | LG | <50% | no |
| 10 | 53 | postmenopausal | endometrioid G1 | IA | LG | no | no |
| 13 | 64 | postmenopausal | endometrioid G1 | IV | LG | <50% | NA |
| 14 | 73 | postmenopausal | endometrioid G1 | IB | LG | >50% | no |
| 16 | 69 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 18 | 79 | postmenopausal | endometrioid G1 | IB | LG | >50% | no |
| 19 | 74 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 20 | 76 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 21 | 53 | premenopausal | endometrioid G2 | IA | LG | no | no |
| 23 | 45 | premenopausal | endometrioid G1 | IA | LG | no | no |
| 24 | 69 | postmenopausal | endometrioid G2 | IB | LG | >50% | yes |
| 25 | 54 | premenopausal | endometrioid G3 | IA | HG | <50% | no |
| 26 | 72 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 30 | 54 | premenopausal | endometrioid G1 | IA | LG | no | no |
| 33 | 77 | postmenopausal | endometrioid G3 | IB | HG | >50% | no |
| 34 | 57 | postmenopausal | mucinous | IA | LG | <50% | no |
| 40 | 71 | postmenopausal | serous | IA | HG | <50% | no |
| 44 | 73 | postmenopausal | serous | IB | HG | >50% | yes |
| 47 | 27 | premenopausal | dedifferentiated | IA | HG | <50% | no |
| 49 | 70 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 50 | 73 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 51 | 75 | postmenopausal | endometrioid G2 | IA | LG | >50% | yes |
| 52 | 75 | postmenopausal | endometrioid G2 | IA | LG | <50% | yes |
| 53 | 50 | postmenopausal | endometrioid G3 | IA | HG | <50% | yes |
| 54 | 71 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 56 | 55 | postmenopausal | endometrioid G1 | IA | LG | no | no |
| 57 | 43 | premenopausal | endometrioid G1 | IA | LG | no | no |
| 62 | 59 | postmenopausal | endometrioid G1 | IA | LG | no | no |
| 63 | 66 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 65 | 80 | postmenopausal | carcinosarcoma | IB | HG | >50% | yes |
| 66 | 72 | postmenopausal | endometrioid G1 | IA | LG | <50% | no |
| 68 | 45 | premenopausal | endometrioid G1 | II | LG | <50% | no |
| 71 | 48 | premenopausal | serous | IA | HG | <50% | no |
| Gene Symbol | Gene/AF Name | Assay ID |
|---|---|---|
| CSF3 | colony stimulating factor 3 | Hs99999083_m1 |
| CXCL12 | C-X-C motif chemokine ligand 12 | Hs00171022_m1 |
| ENPP2 | ectonucleotide pyrophosphatase/phosphodiesterase 2 | Hs00196470_m1 |
| FBLN5 | fibulin 5 | Hs00197064_m1 |
| FGF2 | fibroblast growth factor 2 | Hs00266645_m1 |
| FST | follistatin | Hs00246256_m1 |
| HPRT1 * | hypoxanthine-guanine phosphoribosyltransferase | Hs02758991_g1 |
| IL8 | C-X-C motif chemokine ligand 8 | Hs00174103_m1 |
| LEP | leptin | Hs00174877_m1 |
| LYVE1 | lymphatic vessel endothelial hyaluronan receptor 1 | Hs00272659_m1 |
| NRP1 | neuropilin 1 | Hs00826128_m1 |
| PDGFRB | platelet derived growth factor receptor beta | Hs00387364_m1 |
| POLR2A * | DNA-directed RNA polymerase II subunit RPB1 | Hs00426592_m1 |
| SERPINF1 | serpin family F member 1 | Hs00171467_m1 |
| TEK | TEK receptor tyrosine kinase; Tie-2 | Hs00176096_m1 |
| TIMP2 | TIMP metallopeptidase inhibitor 2 | Hs00234278_m1 |
| TIMP3 | TIMP metallopeptidase inhibitor 3 | Hs00165949_m1 |
| Genes | Mean FR | FR CI 95% | p-Value ‡ | ||
|---|---|---|---|---|---|
| IL8 | 4.75 | 2.46 | 7.04 | 0.0164 | * |
| CXCL12 | −18.21 | −31.95 | −4.46 | 0.0015 | ** |
| FGF2 | −7.39 | −10.51 | −4.27 | 0.0015 | ** |
| LEP | 4.68 | 2.87 | 6.49 | 0.0104 | * |
| LYVE1 | −10.98 | −16.52 | −5.44 | 0.0015 | ** |
| NRP1 | −3.67 | −5.11 | −2.22 | 0.0015 | ** |
| TIMP2 | −6.79 | −8.87 | −4.70 | 0.0015 | ** |
| TIMP3 | −14.77 | −21.77 | −7.77 | 0.0015 | ** |
| CSF3 | −10.43 | −20.51 | −0.35 | 0.8328 | ns |
| ENPP2 | −10.10 | −15.05 | −5.14 | 0.0015 | ** |
| FBLN5 | −8.27 | −12.70 | −3.83 | 0.0015 | ** |
| FST | −6.77 | −9.98 | −3.56 | 0.0668 | ns |
| PDGFRB | −4.50 | −6.08 | −2.92 | 0.0015 | ** |
| TEK | −5.20 | −7.50 | −2.91 | 0.0015 | ** |
| SERPINF1 | −8.68 | −12.71 | −4.66 | 0.0015 | ** |
| Tumour Tissue | Tumour-Adjacent Tissue | Tumour Tissue vs. Tumour-Adjacent Tissue | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene_T | Gene_T | r | p-Value | Gene_TA | Gene_TA | r | p-Value | Gene_T | Gene_TA | r | p-Value | |
| All EC Patients | PDGFRB | SERPINF1 | 0.906 | 3.3 × 10−14 | ||||||||
| TEK | SERPINF1 | 0.888 | 4.9 × 10−13 | |||||||||
| TIMP2 | TIMP3 | 0.879 | 1.7 × 10−12 | |||||||||
| FGF2 | TIMP3 | 0.871 | 4.9 × 10−12 | |||||||||
| PDGFRB | TEK | 0.866 | 9.2 × 10−12 | |||||||||
| EC Patients without LVI | PDGFRB | SERPINF1 | 0.878 | 7.9 × 10−10 | ENPP2 | TEK | 0.854 | 7.2 × 10−9 | ||||
| CXCL12 | TIMP3 | 0.875 | 1.1 × 10−9 | |||||||||
| FBLN5 | PDGFRB | 0.873 | 1.4 × 10−9 | |||||||||
| TIMP2 | SERPINF1 | 0.858 | 5.3 × 10−9 | |||||||||
| TIMP2 | TIMP3 | 0.857 | 5.8 × 10−9 | |||||||||
| EC Patients with LVI | PDGFRB | TEK | 1.000 | 4.9 × 10−5 | CXCL12 | SERPINF1 | 0.952 | 0.001 | TIMP3 | NRP1 | 0.929 | 0.002 |
| LYVE1 | NRP1 | 0.976 | 4.0 × 10−4 | CXCL12 | TIMP2 | 0.929 | 0.002 | ENPP2 | PDGFRB | 0.929 | 0.002 | |
| LYVE1 | PDGFRB | 0.976 | 4.0 × 10−4 | TIMP2 | SERPINF1 | 0.929 | 0.002 | FGF2 | PDGFRB | 0.905 | 0.005 | |
| LYVE1 | TEK | 0.976 | 4.0 × 10−4 | TIMP2 | TIMP3 | 0.905 | 0.005 | ENPP2 | NRP1 | 0.905 | 0.005 | |
| NRP1 | SERPINF1 | 0.976 | 4.0 × 10−4 | FBLN5 | SERPINF1 | 0.905 | 0.005 | FGF2 | NRP1 | 0.881 | 0.007 | |
| PDGFRB | SERPINF1 | 0.976 | 4.0 × 10−4 | FGF2 | TIMP3 | 0.857 | 0.011 | NRP1 | NRP1 | 0.881 | 0.007 | |
| TEK | SERPINF1 | 0.976 | 4.0 × 10−4 | FGF2 | ENPP2 | 0.857 | 0.011 | PDGFRB | NRP1 | 0.881 | 0.007 | |
| LYVE1 | SERPINF1 | 0.952 | 1.1 × 10−3 | TIMP3 | ENPP2 | 0.857 | 0.011 | TEK | NRP1 | 0.881 | 0.007 | |
| CXCL12 | FST | 0.922 | 2.6 × 10−3 | PDGFRB | TEK | 0.857 | 0.011 | |||||
| FGF2 | TIMP3 | 0.905 | 4.6 × 10−3 | |||||||||
| NRP1 | TIMP3 | 0.905 | 4.6 × 10−3 | |||||||||
| TIMP3 | PDGFRB | 0.905 | 4.6 × 10−3 | |||||||||
| TIMP3 | TEK | 0.905 | 4.6 × 10−3 | |||||||||
| IL8 | CSF3 | 0.881 | 7.2 × 10−3 | |||||||||
| CXCL12 | NRP1 | 0.881 | 7.2 × 10−3 | |||||||||
| CXCL12 | PDGFRB | 0.881 | 7.2 × 10−3 | |||||||||
| CXCL12 | TEK | 0.881 | 7.2 × 10−3 | |||||||||
| TIMP3 | ENPP2 | 0.881 | 7.2 × 10−3 | |||||||||
| TIMP3 | SERPINF1 | 0.881 | 7.2 × 10−3 | |||||||||
| CXCL12 | TIMP3 | 0.857 | 1.1 × 10−2 | |||||||||
| CXCL12 | SERPINF1 | 0.857 | 1.1 × 10−2 | |||||||||
| FGF2 | TIMP2 | 0.857 | 1.1 × 10−2 | |||||||||
| LYVE1 | TIMP3 | 0.857 | 1.1 × 10−2 | |||||||||
| ENPP2 | SERPINF1 | 0.857 | 1.1 × 10−2 | |||||||||
| Model | Accuracy | Precision | Recall | F1 | AUC | Sensitivity | Specificity |
|---|---|---|---|---|---|---|---|
| All Data | 93.2% | 100% | 86.4% | 92.7% | 1.00 | 86% | 100% |
| Tumour (Normalised T) | 90.9% | 100% | 81.8% | 90% | 0.99 | 82% | 100% |
| Adjacent (Normalised TA) | 95.5% | 91.7% | 100% | 95.7% | 0.99 | 100% | 91% |
| Model | Accuracy | Precision | Recall | F1 | AUC | Sensitivity | Specificity |
|---|---|---|---|---|---|---|---|
| All Data | 85.7% | 75% | 75% | 75% | 0.78 | 75% | 90% |
| Tumour (Normalised T) | 85.7% | 66.7% | 100% | 80% | 0.98 | 100% | 80% |
| Adjacent (Normalised TA) | 50% | 28.6% | 50% | 36.4% | 0.40 | 50% | 50% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roškar, L.; Kokol, M.; Pavlič, R.; Roškar, I.; Smrkolj, Š.; Rižner, T.L. Decreased Gene Expression of Antiangiogenic Factors in Endometrial Cancer: qPCR Analysis and Machine Learning Modelling. Cancers 2023, 15, 3661. https://doi.org/10.3390/cancers15143661
Roškar L, Kokol M, Pavlič R, Roškar I, Smrkolj Š, Rižner TL. Decreased Gene Expression of Antiangiogenic Factors in Endometrial Cancer: qPCR Analysis and Machine Learning Modelling. Cancers. 2023; 15(14):3661. https://doi.org/10.3390/cancers15143661
Chicago/Turabian StyleRoškar, Luka, Marko Kokol, Renata Pavlič, Irena Roškar, Špela Smrkolj, and Tea Lanišnik Rižner. 2023. "Decreased Gene Expression of Antiangiogenic Factors in Endometrial Cancer: qPCR Analysis and Machine Learning Modelling" Cancers 15, no. 14: 3661. https://doi.org/10.3390/cancers15143661
APA StyleRoškar, L., Kokol, M., Pavlič, R., Roškar, I., Smrkolj, Š., & Rižner, T. L. (2023). Decreased Gene Expression of Antiangiogenic Factors in Endometrial Cancer: qPCR Analysis and Machine Learning Modelling. Cancers, 15(14), 3661. https://doi.org/10.3390/cancers15143661