
EMP2 Serves as a Functional Biomarker for Chemotherapy-Resistant Triple-Negative Breast Cancer

 , , , , , , ,
, , , , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Tumor Specimens
2.2. Cell Lines and Cell Culture
2.3. Creation of Taxane Resistant Cells
2.4. Anti-EMP2 mAbs
3. Results
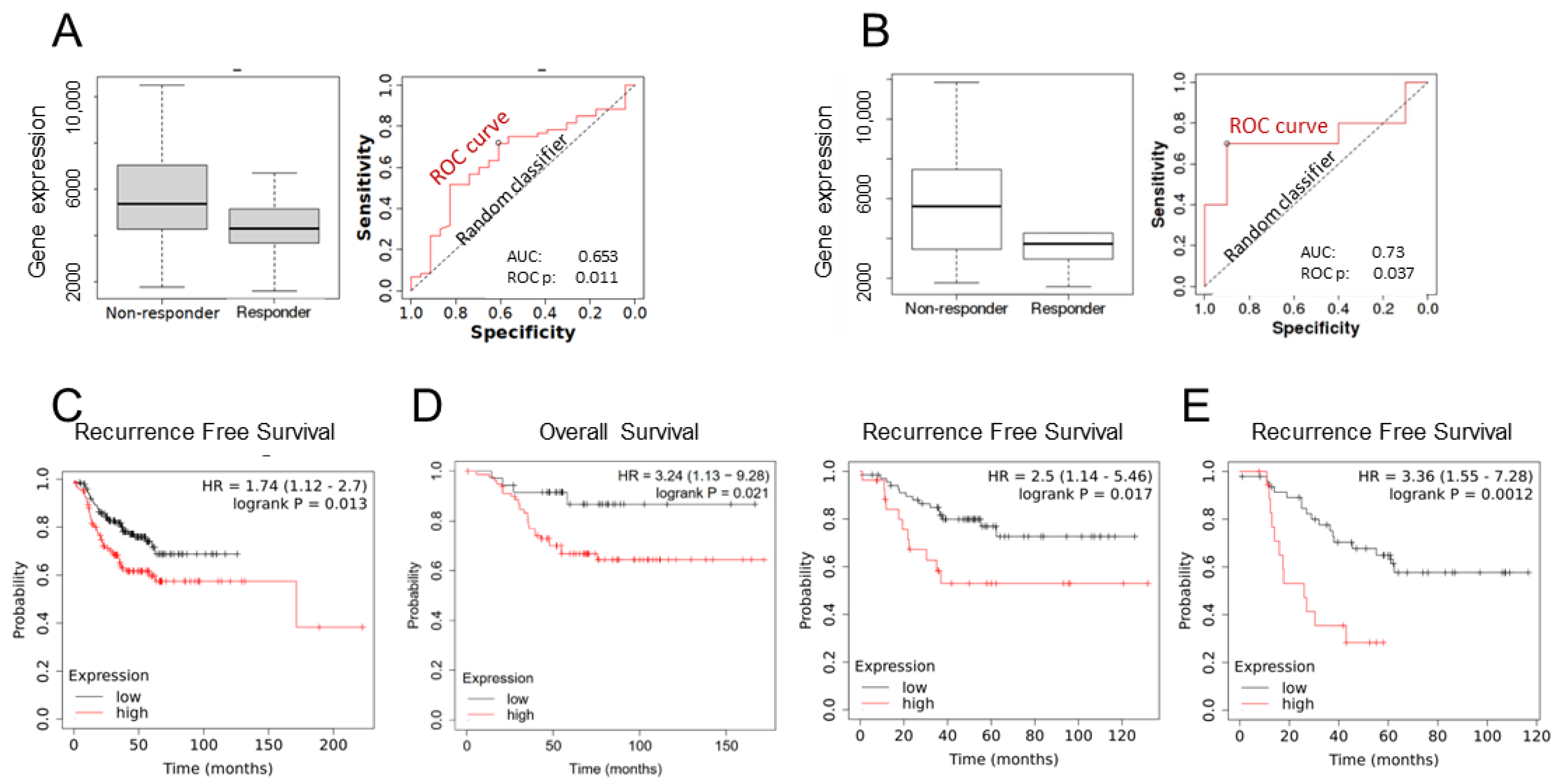
3.1. EMP2 Transcript Expression in Chemotherapy Resistant TNBC Tumors
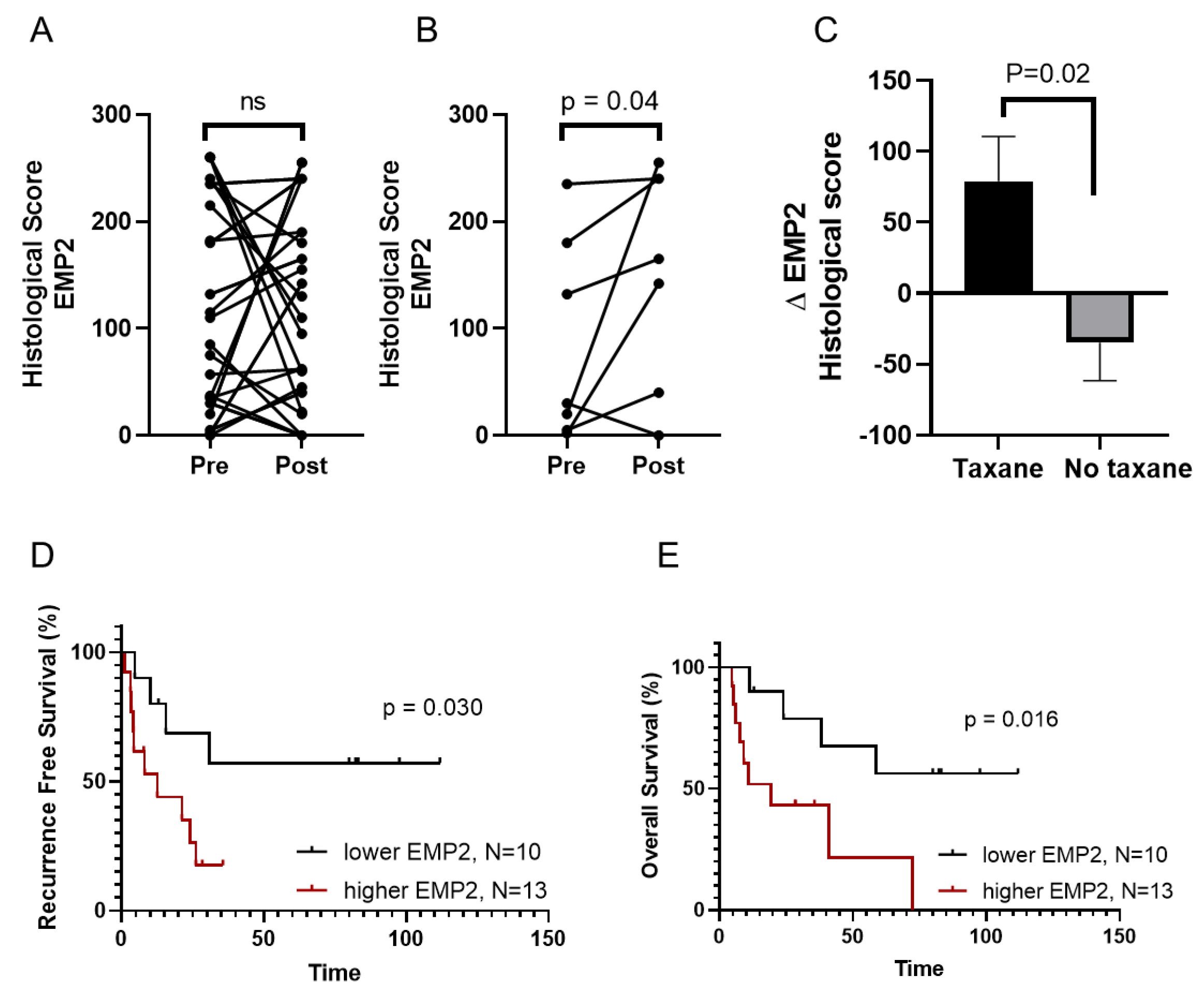
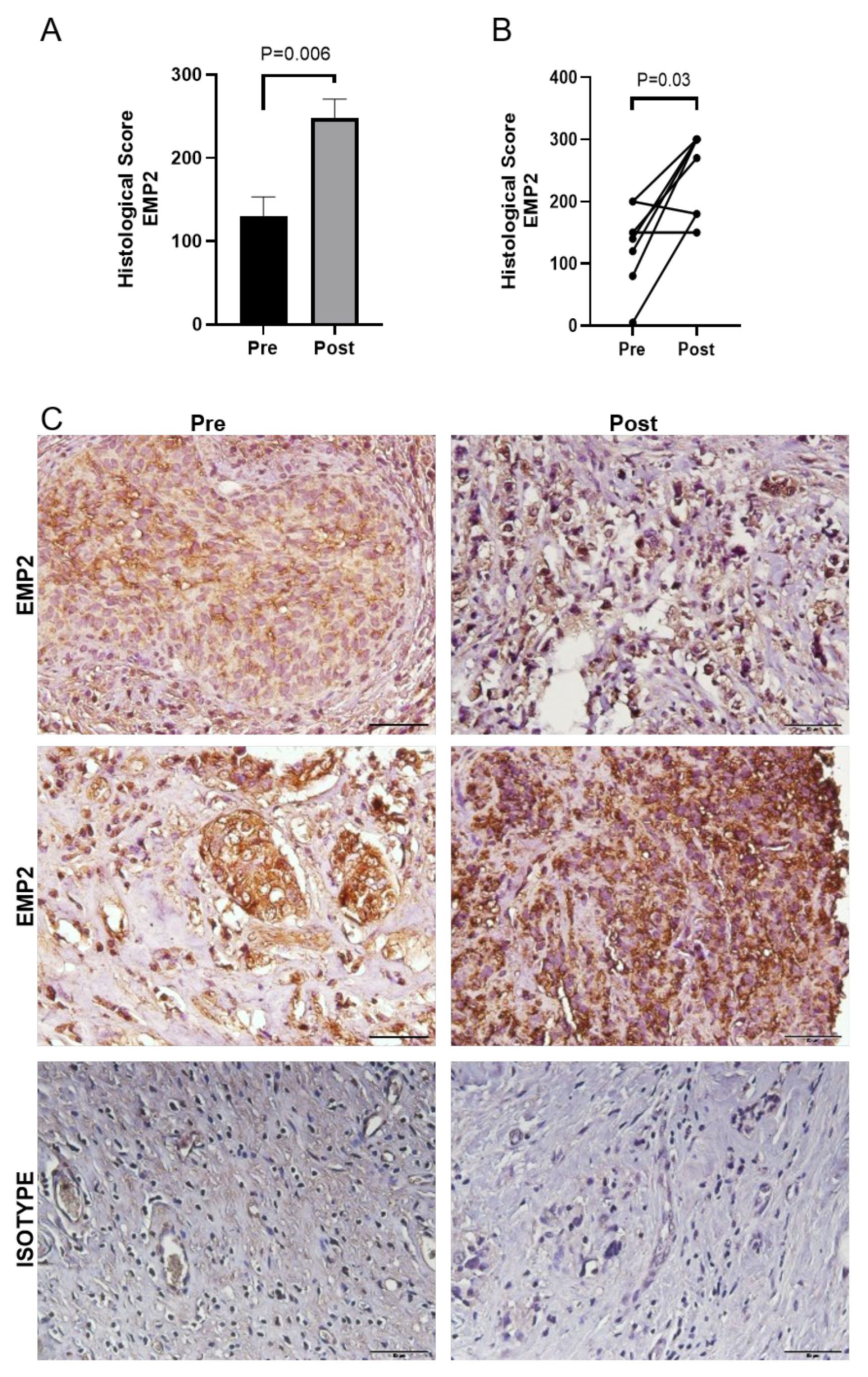
3.2. EMP2 Protein Expression in Chemotherapy Resistant TNBC Tumors
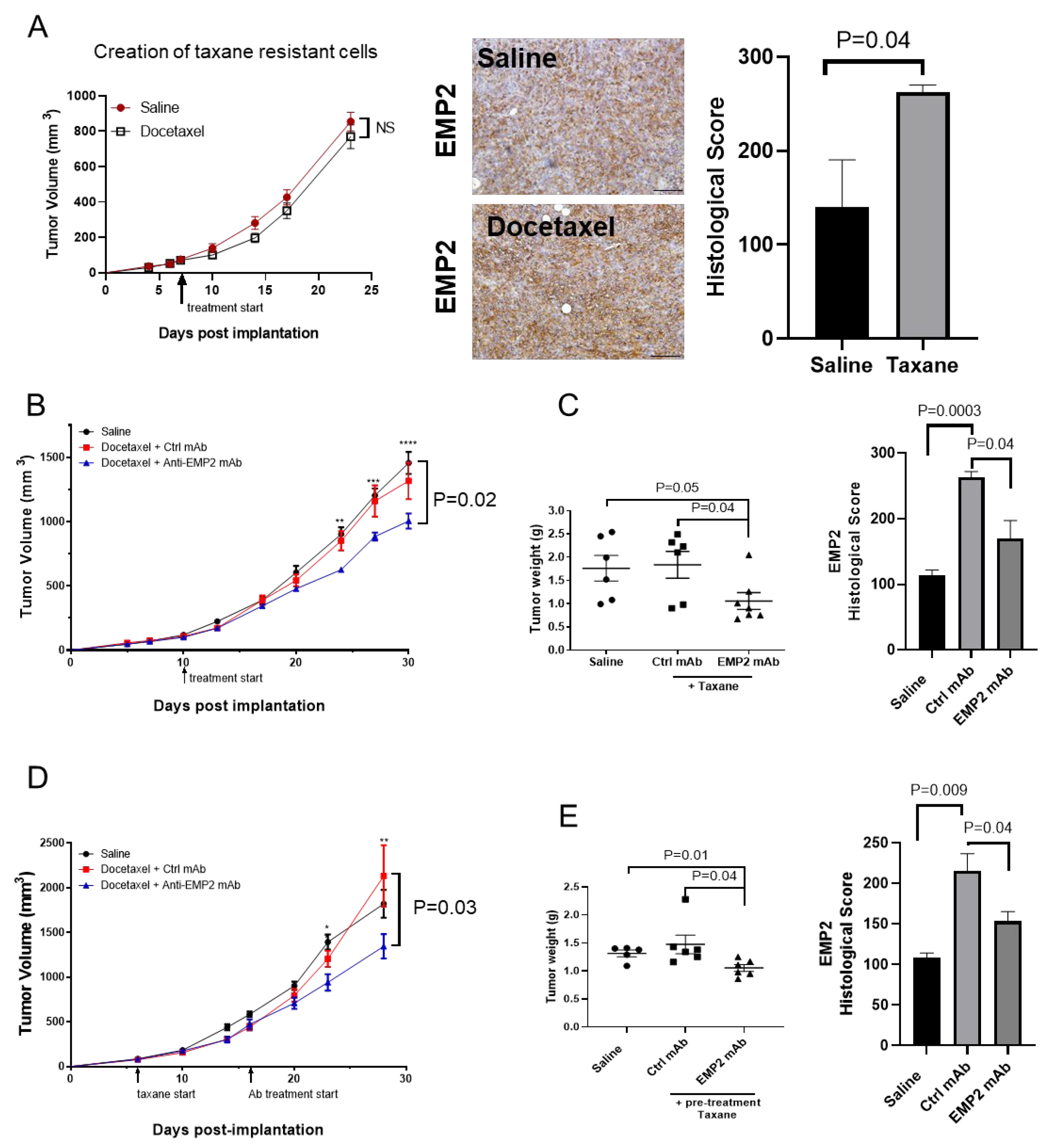
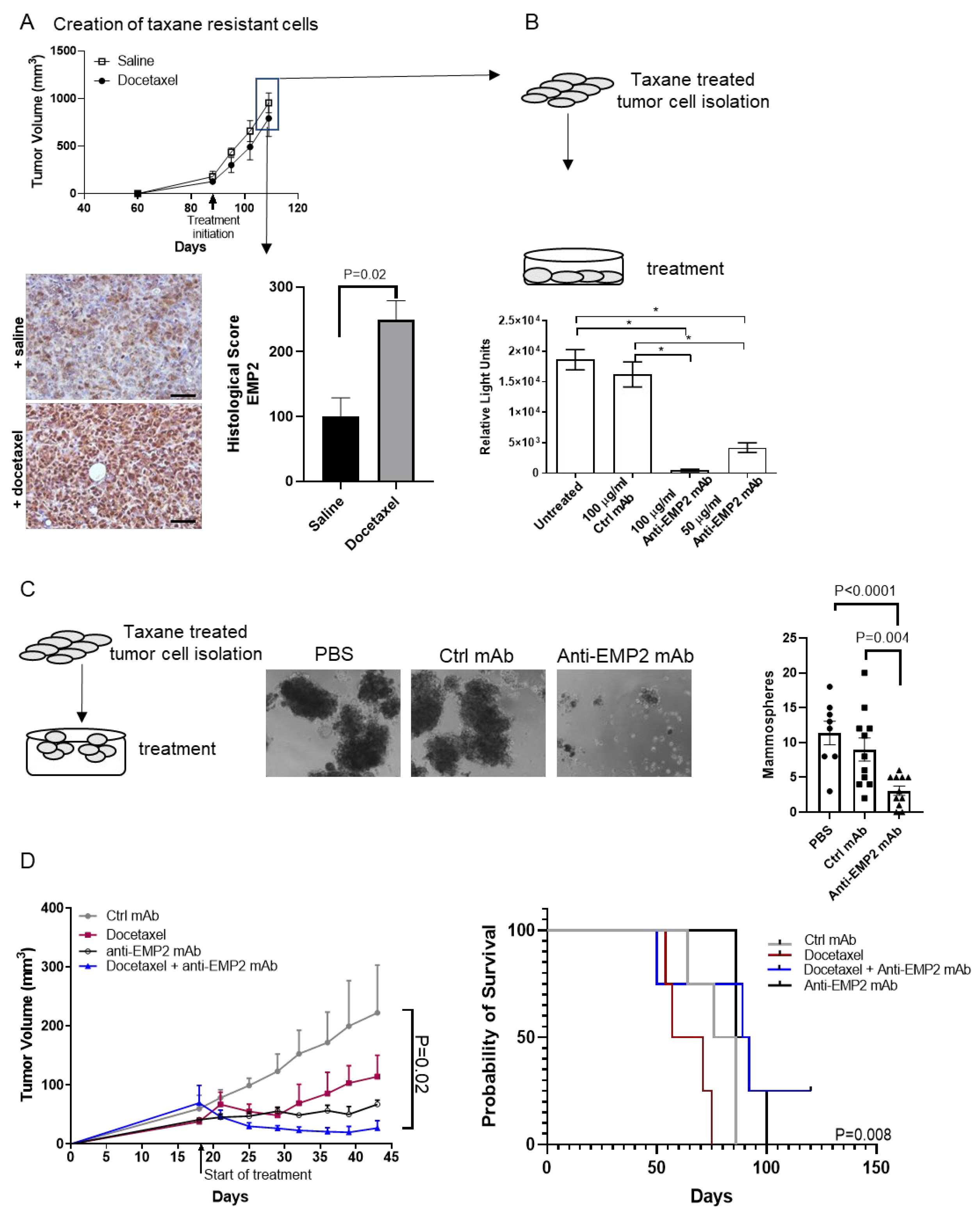
3.3. Modeling Chemo-Resistance Using Syngeneic Mouse Models
3.4. In Vivo Effects Using Human Xenograft Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Cancer Institute. SEER*Explorer: An interactive website for SEER cancer statistics. In Surveillance Research Program; National Cancer Institute: Rockville, ML, USA, 2021. [Google Scholar]
- Leon-Ferre, R.A.; Goetz, M.P. Advances in systemic therapies for triple negative breast cancer. BMJ 2023, 381, e071674. [Google Scholar] [CrossRef]
- Tsai, J.; Bertoni, D.; Hernandez-Boussard, T.; Telli, M.L.; Wapnir, I.L. Lymph node ratio analysis after neoadjuvant chemotherapy is prognostic in hormone receptor-positive and triple-negative breast cancer. Ann. Surg. Oncol. 2016, 23, 3310–3316. [Google Scholar] [CrossRef]
- Lai, J.-I.; Chao, T.-C.; Liu, C.-Y.; Huang, C.-C.; Tseng, L.-M. A systemic review of taxanes and their side effects in metastatic breast cancer. Front. Oncol. 2022, 12, 940239. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.A.; Vogel, C.L.; Irwin, D.H.; Kirshner, J.J.; Patel, R. Multicenter phase II trial of weekly paclitaxel in women with metastatic breast cancer. J. Clin. Oncol. 2001, 19, 4216–4223. [Google Scholar] [CrossRef]
- Bachelot, T.; Ciruelos, E.; Schneeweiss, A.; Puglisi, F.; Peretz-Yablonski, T.; Bondarenko, I.; Paluch-Shimon, S.; Wardley, A.; Merot, J.-L.; Du Toit, Y. Preliminary safety and efficacy of first-line pertuzumab combined with trastuzumab and taxane therapy for HER2-positive locally recurrent or metastatic breast cancer (PERUSE). Ann. Oncol. 2019, 30, 766–773. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Rodrigues-Ferreira, S.; Moindjie, H.; Haykal, M.M.; Nahmias, C. Predicting and Overcoming Taxane Chemoresistance. Trends Mol. Med. 2021, 27, 138–151. [Google Scholar] [CrossRef]
- Abuhadra, N.; Stecklein, S.; Sharma, P.; Moulder, S. Early-stage Triple-negative Breast Cancer: Time to Optimize Personalized Strategies. Oncologist 2022, 27, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Romeo, V.; Accardo, G.; Perillo, T.; Basso, L.; Garbino, N.; Nicolai, E.; Maurea, S.; Salvatore, M. Assessment and Prediction of Response to Neoadjuvant Chemotherapy in Breast Cancer: A Comparison of Imaging Modalities and Future Perspectives. Cancers 2021, 13, 3521. [Google Scholar] [CrossRef]
- Won, K.A.; Spruck, C. Triple-negative breast cancer therapy: Current and future perspectives (Review). Int. J. Oncol. 2020, 57, 1245–1261. [Google Scholar] [CrossRef]
- Sieluk, J.; Haiderali, A.; Huang, M.; Yang, L.; Tryfonidis, K.; Hirshfield, K.M. SEER-Medicare study of early-stage triple-negative breast cancer: Real-world treatment patterns, survival, and expenditures 2010 to 2016. J. Clin. Oncol. 2020, 38 (Suppl. S15), e12512. [Google Scholar] [CrossRef]
- Schuster, E.; Taftaf, R.; Reduzzi, C.; Albert, M.K.; Romero-Calvo, I.; Liu, H. Better together: Circulating tumor cell clustering in metastatic cancer. Trends Cancer 2021, 7, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Dillard, C.; Kiyohara, M.; Mah, V.; McDermott, S.P.; Bazzoun, D.; Tsui, J.; Chan, A.M.; Haddad, G.; Pellegrini, M.; Chang, Y.L.; et al. EMP2 Is a Novel Regulator of Stemness in Breast Cancer Cells. Mol. Cancer Ther. 2020, 19, 1682–1695. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Maresh, E.L.; Helguera, G.F.; Kiyohara, M.; Qin, Y.; Ashki, N.; Daniels-Wells, T.R.; Aziz, N.; Gordon, L.K.; Braun, J.; et al. Rationale and preclinical efficacy of a novel anti-EMP2 antibody for the treatment of invasive breast cancer. Mol. Cancer Ther. 2014, 13, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Balko, J.M.; Giltnane, J.M.; Wang, K.; Schwarz, L.J.; Young, C.D.; Cook, R.S.; Owens, P.; Sanders, M.E.; Kuba, M.G.; Sánchez, V.; et al. Molecular profiling of the residual disease of triple-negative breast cancers after neoadjuvant chemotherapy identifies actionable therapeutic targets. Cancer Discov. 2014, 4, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, A.; Carton, M.; Guiu, S.; Augereau, P.; Uwer, L.; Mouret-Reynier, M.A.; Levy, C.; Eymard, J.C.; Ferrero, J.M.; Leheurteur, M.; et al. Efficacy of taxanes rechallenge in first-line treatment of early metastatic relapse of patients with HER2-negative breast cancer previously treated with a (neo)adjuvant taxanes regimen: A multicentre retrospective observational study. Breast 2022, 65, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.M.; Olafsen, T.; Tsui, J.; Salazar, F.B.; Aguirre, B.; Zettlitz, K.A.; Condro, M.; Wu, A.M.; Braun, J.; Gordon, L.K.; et al. 89Zr-Imm unoPET for the Specific Detection of EMP2-Positive Tumors. Mol. Cancer Ther. 2024. [Google Scholar] [CrossRef] [PubMed]
- Gradishar, W.J. Taxanes for the treatment of metastatic breast cancer. Breast Cancer 2012, 6, 159–171. [Google Scholar] [CrossRef]
- Fekete, J.T.; Győrffy, B. ROCplot.org: Validating predictive biomarkers of chemotherapy/hormonal therapy/anti-HER2 therapy using transcriptomic data of 3104 breast cancer patients. Int. J. Cancer 2019, 145, 3140–3151. [Google Scholar] [CrossRef]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef]
- Pienta, K.J. Preclinical mechanisms of action of docetaxel and docetaxel combinations in prostate cancer. Semin. Oncol. 2001, 28 (Suppl. S15), 3–7. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Grasselly, C.; Denis, M.; Bourguignon, A.; Talhi, N.; Mathe, D.; Tourette, A.; Serre, L.; Jordheim, L.P.; Matera, E.L.; Dumontet, C. The Antitumor Activity of Combinations of Cytotoxic Chemotherapy and Immune Checkpoint Inhibitors Is Model-Dependent. Front. Immunol. 2018, 9, 2100. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, E.; Nasrollahzadeh, J.; Fatemi, R.; Barzegar-Yar Mohamadi, L.; Saliminejad, K.; Amiri, Z.; Kimiagar, M.; Houshyari, M.; Tavakoli, M.; Idali, F. Effects of Combined Soy Isoflavone Extract and Docetaxel Treatment on Murine 4T1 Breast Tumor Model. Avicenna J. Med. Biotechnol. 2015, 7, 16–21. [Google Scholar] [PubMed]
- Charafe-Jauffret, E.; Ginestier, C.; Iovino, F.; Wicinski, J.; Cervera, N.; Finetti, P.; Hur, M.-H.; Diebel, M.E.; Monville, F.; Dutcher, J.; et al. Breast Cancer Cell Lines Contain Functional Cancer Stem Cells with Metastatic Capacity and a Distinct Molecular Signature. Cancer Res. 2009, 69, 1302–1313. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Y.; Deng, L.; Wang, D.; He, X.; Zhou, L.; Wicha, M.S.; Bai, F.; Liu, S. Transcriptional profiles of different states of cancer stem cells in triple-negative breast cancer. Mol. Cancer 2018, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Ehmsen, S.; Pedersen, M.H.; Wang, G.; Terp, M.G.; Arslanagic, A.; Hood, B.L.; Conrads, T.P.; Leth-Larsen, R.; Ditzel, H.J. Increased Chol-esterol Biosynthesis Is a Key Characteristic of Breast Cancer Stem Cells Influencing Patient Outcome. Cell Rep. 2019, 27, 3927–3938.e3926. [Google Scholar] [CrossRef] [PubMed]
- Flamant, L.; Notte, A.; Ninane, N.; Raes, M.; Michiels, C. Anti-apoptotic role of HIF-1 and AP-1 in paclitaxel exposed breast cancer cells under hypoxia. Mol. Cancer 2010, 9, 191. [Google Scholar] [CrossRef]
- Daskalaki, I.; Gkikas, I.; Tavernarakis, N. Hypoxia and Selective Autophagy in Cancer Development and Therapy. Front. Cell Dev. Biol. 2018, 6, 104. [Google Scholar] [CrossRef]
- Goldman, A.; Majumder, B.; Dhawan, A.; Ravi, S.; Goldman, D.; Kohandel, M.; Majumder, P.K.; Sengupta, S. Temporally sequenced anticancer drugs overcome adaptive resistance by targeting a vulnerable chemotherapy-induced phenotypic transition. Nat. Commun. 2015, 6, 6139. [Google Scholar] [CrossRef]
- Abdoli Shadbad, M.; Hosseinkhani, N.; Asadzadeh, Z.; Derakhshani, A.; Karim Ahangar, N.; Hemmat, N.; Lotfinejad, P.; Brunetti, O.; Silvestris, N.; Baradaran, B. A Systematic Review to Clarify the Prognostic Values of CD44 and CD44(+)CD24(-) Phenotype in Triple-Negative Breast Cancer Patients: Lessons Learned and The Road Ahead. Front. Oncol. 2021, 11, 689839. [Google Scholar] [CrossRef]
- Gordon, L.K.; Kiyohara, M.; Fu, M.; Braun, J.; Dhawan, P.; Chan, A.; Goodglick, L.; Wadehra, M. EMP2 regulates angiogenesis in endometrial cancer cells through induction of VEGF. Oncogene 2013, 32, 5369–5376. [Google Scholar] [CrossRef]
- Qin, Y.; Fu, M.; Takahashi, M.; Iwanami, A.; Kuga, D.; Rao, R.G.; Sudhakar, D.; Huang, T.; Kiyohara, M.; Torres, K.; et al. Epithelial mem brane protein-2 (EMP2) activates Src protein and is a novel therapeutic target for glioblastoma. J. Clin. Investig. 2020, 130, 157–170. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Pietenpol, J.A.; Tan, A.R. Triple-Negative Breast Cancer: Molecular Subtypes and New Targets for Therapy. Am. Soc. Clin. Oncol. Educ. Book 2015, 35, e31–e39. [Google Scholar] [CrossRef]
- Mustacchi, G.; De Laurentiis, M. The role of taxanes in triple-negative breast cancer: Literature review. Drug. Des. Devel. Ther. 2015, 9, 4303–4318. [Google Scholar] [CrossRef]
- Zagami, P.; Carey, L.A. Triple negative breast cancer: Pitfalls and progress. NPJ Breast Cancer 2022, 8, 95. [Google Scholar] [CrossRef]
- Sharma, P.; López-Tarruella, S.; García-Saenz, J.A.; Khan, Q.J.; Gómez, H.L.; Prat, A.; Moreno, F.; Jerez-Gilarranz, Y.; Barnadas, A.; Picornell, A.C.; et al. Pathological Response and Survival in Triple-Negative Breast Cancer Following Neoadjuvant Carboplatin plus Docetaxel. Clin. Cancer Res. 2018, 24, 5820–5829. [Google Scholar] [CrossRef]
- Cortazar, P.; Zhang, L.; Untch, M.; Mehta, K.; Costantino, J.P.; Wolmark, N.; Bonnefoi, H.; Cameron, D.; Gianni, L.; Valagussa, P.; et al. Pathological complete response and long-term clinical benefit in breast cancer: The CTNeoBC pooled analysis. Lancet 2014, 384, 164–172. [Google Scholar] [CrossRef]
- Gooding, A.J.; Schiemann, W.P. Epithelial–Mesenchymal Transition Programs and Cancer Stem Cell Phenotypes: Mediators of Breast Cancer Therapy Resistance. Mol. Cancer Res. 2020, 18, 1257–1270. [Google Scholar] [CrossRef]
- Almansour, N.M. Triple-Negative Breast Cancer: A Brief Review About Epidemiology, Risk Factors, Signaling Pathways, Treatment and Role of Artificial Intelligence. Front. Mol. Biosci. 2022, 9, 836417. [Google Scholar] [CrossRef]
- Eyre, R.; Alférez, D.G.; Spence, K.; Kamal, M.; Shaw, F.L.; Simões, B.M.; Santiago-Gómez, A.; Sarmiento-Castro, A.; Bramley, M.; Absar, M.; et al. Patient-derived Mammosphere and Xenograft Tumour Initiation Correlates with Progression to Metastasis. J. Mammary Gland. Biol. Neoplasia 2016, 21, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Wadehra, M.; Sulur, G.G.; Braun, J.; Gordon, L.K.; Goodglick, L. Epithelial membrane protein-2 is expressed in discrete anatomical regions of the eye. Exp. Mol. Pathol. 2003, 74, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Wadehra, M.; Fisk, B.C.; Goodglick, L.; Braun, J. Epithelial membrane protein 2, a 4-transmembrane protein that suppresses B-cell lymphoma tumorigenicity. Blood 2001, 97, 3890–3895. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Lanczky, A.; Szallasi, Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EMP2 Increased after Chemotherapy (Yes/No) | p-Value | |
|---|---|---|
| Age | ||
| Median | 46 years | 0.47 §§ |
| Range | 29–69 years | |
| Stage | 1 § | |
| IIa | 1 | |
| IIb | 2 | |
| IIIb | 19 | |
| IIIc | 1 | |
| Lymph node spread | 0.068 § | |
| Present | 16 | |
| Absent | 7 | |
| Menopausal status | ||
| Pre-menopausal | 11 | 1 § |
| Post-menopausal | 12 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, A.M.; Aguirre, B.; Liu, L.; Mah, V.; Balko, J.M.; Tsui, J.; Wadehra, N.P.; Moatamed, N.A.; Khoshchehreh, M.; Dillard, C.M.; et al. EMP2 Serves as a Functional Biomarker for Chemotherapy-Resistant Triple-Negative Breast Cancer. Cancers 2024, 16, 1481. https://doi.org/10.3390/cancers16081481
Chan AM, Aguirre B, Liu L, Mah V, Balko JM, Tsui J, Wadehra NP, Moatamed NA, Khoshchehreh M, Dillard CM, et al. EMP2 Serves as a Functional Biomarker for Chemotherapy-Resistant Triple-Negative Breast Cancer. Cancers. 2024; 16(8):1481. https://doi.org/10.3390/cancers16081481
Chicago/Turabian StyleChan, Ann M., Brian Aguirre, Lucia Liu, Vei Mah, Justin M. Balko, Jessica Tsui, Navin P. Wadehra, Neda A. Moatamed, Mahdi Khoshchehreh, Christen M. Dillard, and et al. 2024. "EMP2 Serves as a Functional Biomarker for Chemotherapy-Resistant Triple-Negative Breast Cancer" Cancers 16, no. 8: 1481. https://doi.org/10.3390/cancers16081481