
Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future



Abstract
:1. Introduction
2. Crop Allelopathy
3. Allelopathic Practices Involved in Weed Management
3.1. Crop Rotation
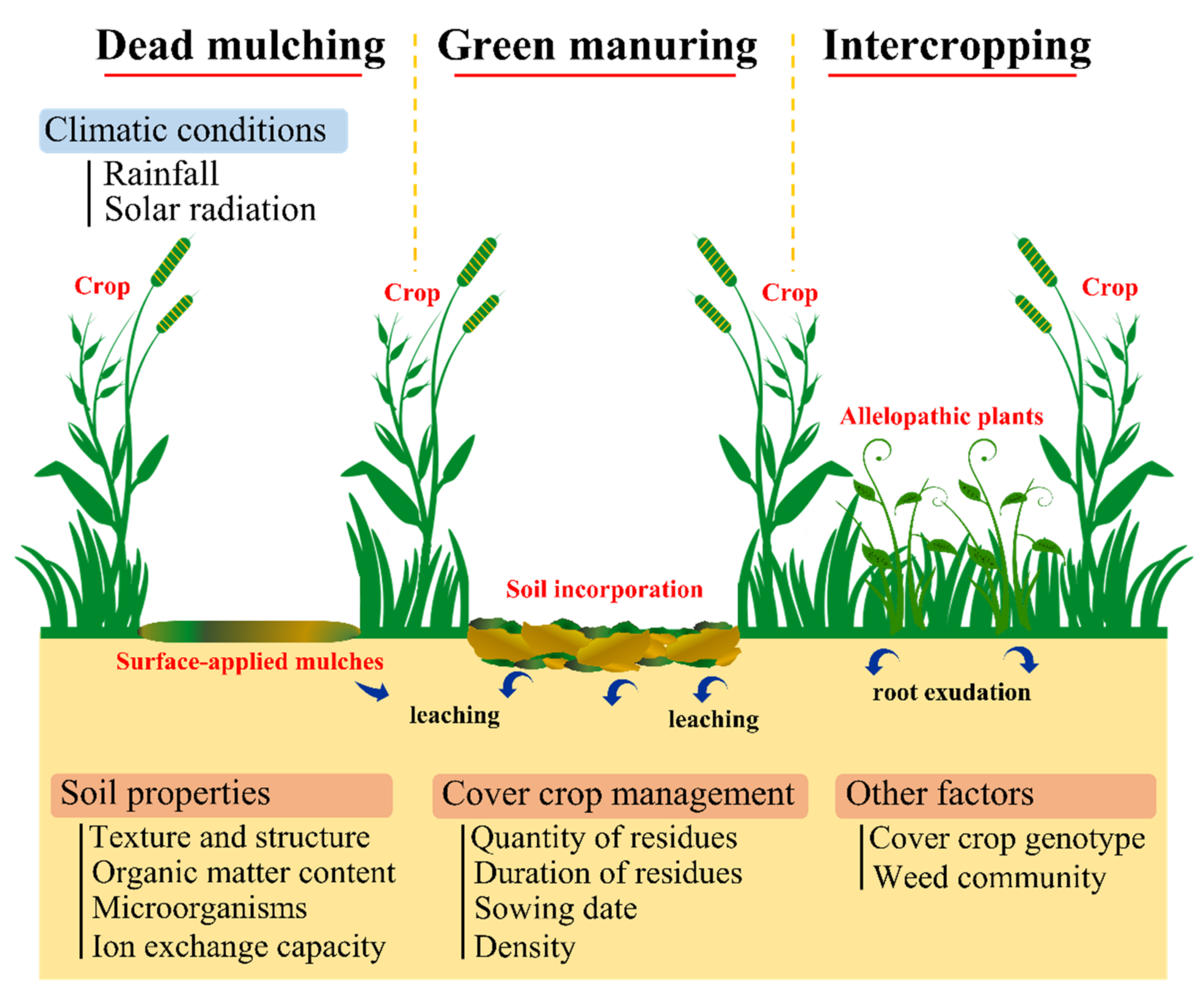
3.2. Cover Cropping
3.2.1. Dead Mulching
3.2.2. Green Manuring
3.2.3. Intercropping
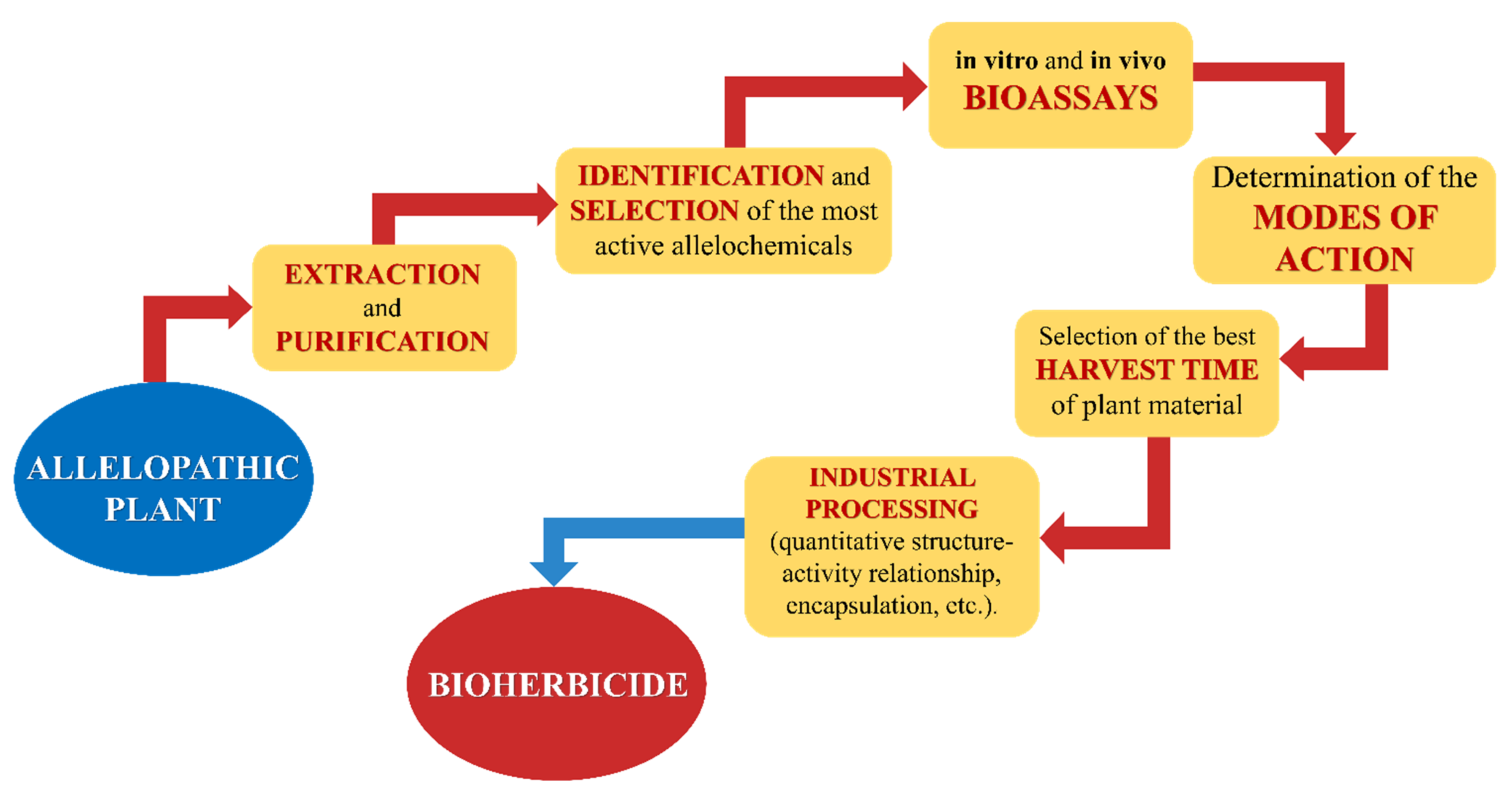
3.3. Bioherbicides
4. Biotechnologies in Crop Allelopathy
4.1. Screening and Selection of Allelopathic Crop Cultivars

{kind=link}
{kind=link}
| Biotechnology | Main Effect | Description | Reference |
|---|---|---|---|
| Genotype selection | Screening allelopathic cultivars | Crop genotypes differ from each other in their allelochemicals’ concentration and allelopathic activity. Screening and selecting genotypes allow obtaining a more allelopathic crop. | [117] |
| Stress induction | Increase in allelochemicals production | Induction of biotic and abiotic stress factors, or a combination of them, stimulates the synthesis of allelochemicals in donor plants. | [118] |
| Tissue culture | Increase in allelochemicals production Isolation from external factors during the study of allelopathic effects | Plant organ cultures such as hairy root cultures, both via normal callogenesis or using Agrobacterium spp. strains, may be applied to increase some competitive traits (e.g., rooting ability) and the production of allelochemicals, as well as to facilitate allelopathic studies. | [119] |
| Traditional breeding | Increase of crops’ allelopathic potential or introduction of allelopathy de novo | Breeding programs can improve the allelopathic potential of crops just as they improved crop yields. However, poligeneticity and the low economic added value make this approach very difficult. | [120] |
| QTL analysis | Identification of genetic markers encoding allelopathic-related traits | The genetic analysis of quantitative trait loci (QTL) is very useful to identify the genes encoding the synthesis of allelochemicals. | [121] |
| Green chemistry | Increase in allelochemicals production | Improving allelochemicals’ biotransformation by overexpressing the nitroreductase enzyme NfsB in Escherichia coli strains as a whole-cell biocatalyst. | [122] |
4.2. Stress Induction
4.3. Genetic Engineering
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Willer, H.; Schlatter, B.; Trávníček, J.; Kemper, L.; Lernoud, J. The World of Organic Agriculture Statistics and Emerging Trends 2020; Research Institute of Organic Agriculture (FiBL): Frick, Switzerland, 2020; p. 337. [Google Scholar]
- Scavo, A.; Restuccia, A.; Mauromicale, G. Allelopathy: General principles and basic aspects for agroecosystem control. In Sustainable Agriculture Reviews; Gaba, S., Smith, B., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2018; Volume 28, pp. 47–101. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984. [Google Scholar]
- Macías, F.A.; Mejías, F.J.R.; Molinillo, J.M.G. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Khanh, T.D.; Chung, M.I.; Xuan, T.D.; Tawata, S. The exploitation of crop allelopathy in sustainable agricultural production. J. Agron. Crop. Sci. 2005, 191, 172–184. [Google Scholar] [CrossRef]
- Farooq, M.; Jabran, K.; Cheema, Z.A.; Wahid, A.; Siddique, K.H.M. The role of allelopathy in agricultural pest management. Pest Manag. Sci. 2011, 67, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, P.C.; Inderjit. Challenges and opportunities in implementing allelopathy for natural weed management. Crop. Prot. 2003, 22, 661–671. [Google Scholar] [CrossRef]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop. Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Willis, R.J. The History of Allelopathy; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar] [CrossRef]
- Weidenhamer, J.D.; Romeo, J.T. Allelopathic properties of Polygonella myriophylla. J. Chem. Ecol. 1989, 15, 1957–1970. [Google Scholar] [CrossRef]
- Macías, F.A.; Oliveros-Bastidas, A.; Marín, D.; Chinchilla, N.; Castellano, D.; Molinillo, J.M.G. Evidence for an allelopathic interaction between rye and wild oats. J. Agric. Food Chem. 2014, 62, 9450–9457. [Google Scholar] [CrossRef]
- Macías, F.A.; Molinillo, J.M.G.; Varela, R.M.; Galindo, J.C. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef]
- Duke, S.O. Proving allelopathy in crop–weed interactions. Weed Sci. 2015, 63, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Qasem, J.R.; Foy, C.L. Weed allelopathy, its ecological impacts and future prospects: A review. J. Crop. Prot. 2001, 4, 43–119. [Google Scholar] [CrossRef]
- Tesio, F.; Ferrero, A. Allelopathy, a chance for sustainable weed management. Int. J. Sustain. Develop. World Ecol. 2010, 17, 377–389. [Google Scholar] [CrossRef]
- Sharangi, A.B. In search of allelopathy from common alliaceae crops for managing weeds in coriander: An overview. Int. J. Agric. Res. 2011, 6, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Funke, G.L. The influence of Artemisia absinthium on neighbouring plants. Blumea 1943, 5, 281–293. [Google Scholar]
- Rial, C.; Tomé, S.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Phytochemical study of safflower roots (Carthamus tinctorius) on the induction of parasitic plant germination and weed control. J. Chem. Ecol. 2020, 46, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Satoh, A. Antimicrobial and nematicidal substances from the root of chicory (Cichorium intybus). In Allelochemicals: Biological Control of Plant Pathogens and Diseases. Disease Management of Fruits and Vegetables; Inderjit, Mukerji, K., Eds.; Springer: Dordrecht, The Netherlands, 2006; Volume 2, pp. 177–180. [Google Scholar] [CrossRef]
- Rial, C.; Novaes, P.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Phytotoxicity of cardoon (Cynara cardunculus) allelochemicals on standard target species and weeds. J. Agric. Food Chem. 2014, 62, 6699–6706. [Google Scholar] [CrossRef]
- Scavo, A.; Rial, C.; Molinillo, J.M.G.; Varela, R.M.; Mauromicale, G.; Macías, F.A. The extraction procedure improves the allelopathic activity of cardoon (Cynara cardunculus var. altilis) leaf allelochemicals. Ind. Crop. Prod. 2019, 128, 479–487. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Mauromicale, G. Leaf extracts of cultivated cardoon as potential bioherbicide. Sci. Hortic. 2020, 261, 109024. [Google Scholar] [CrossRef]
- Macías, F.A.; Fernández, A.; Varela, R.M.; Molinillo, J.M.G.; Torres, A.; Alves, P.L.C.A. Sesquiterpene lactones as allelochemicals. J. Nat. Prod. 2006, 69, 795–800. [Google Scholar] [CrossRef]
- Pan, L.; Sinden, M.R.; Kennedy, A.H.; Chai, H.; Watson, L.E.; Graham, T.L.; Kinghorn, A.D. Bioactive constituents of Helianthus tuberosus (Jerusalem artichoke). Phytochem. Lett. 2009, 2, 15–18. [Google Scholar] [CrossRef]
- Tesio, F.; Weston, L.A.; Ferrero, A. Allelochemicals identified from Jerusalem artichoke (Helianthus tuberosus L.) residues and their potential inhibitory activity in the field and laboratory. Sci. Hortic. 2011, 129, 361–368. [Google Scholar] [CrossRef]
- Chon, S.U.; Jang, H.G.; Kim, D.K.; Kim, Y.M.; Boo, H.O.; Kim, Y.J. Allelopathic potential in lettuce (Lactuca sativa L.) plants. Sci. Hortic. 2005, 106, 309–317. [Google Scholar] [CrossRef]
- Rehman, S.; Shahzad, B.; Bajwa, A.A.; Hussain, S.; Rehman, A.; Cheema, S.A.; Abbas, T.; Ali, A.; Shah, L.; Adkins, S.; et al. Utilizing the allelopathic potential of Brassica species for sustainable crop production: A review. J. Plant Growth Regul. 2018, 38, 343–356. [Google Scholar] [CrossRef]
- Tlili, N.; Nasri, N.; Saadaoui, E.; Khaldi, A.; Triki, S. Carotenoid and tocopherol composition of leaves, buds and flowers of Capparis spinosa grown wild in Tunisia. J. Agric. Food Chem. 2009, 57, 5381–5385. [Google Scholar] [CrossRef] [PubMed]
- Ladhari, A.; Omezzine, F.; Dellagreca, M.; Zarrelli, A.; Haouala, R. Phytotoxic activity of Capparis spinosa L. and its discovered active compounds. Allelopath. J. 2013, 32, 175–190. [Google Scholar]
- Ahmed, M.; Ji, M.; Peiwen, Q.; Liu, Y.; Gu, Z. Phytochemical screening, total phenolic and flavonoids contents and antioxidant activities of Citrullus colocynthis L. and Cannabis sativa L. Appl. Ecol. Environ. 2019, 17, 6961–6979. [Google Scholar] [CrossRef]
- Soni, B.; Tseng, T.P.M.; Yue, Z. Identification and quantification of allelochemicals from selected sweet potato (Ipomoea batatas (L.) Lam.) cultivars. Am. J. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Hao, Z.P.; Wang, Q.; Christie, P.; Li, X.L. Allelopathic potential of watermelon tissues and root exudates. Sci. Hortic. 2007, 112, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.Q.; Ye, S.F.; Zhang, M.F.; Hu, W.H. Effects of root exudates and aqueous root extracts of cucumber (Cucumis sativus) and allelochemicals, on photosynthesis and antioxidant enzymes in cucumber. Biochem. Syst. Ecol. 2003, 31, 129–139. [Google Scholar] [CrossRef]
- Li, X.; Ding, C.; Hua, K.; Zhang, T.; Zhang, Y.; Zhao, L. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy. Soil Biol. Biochem. 2014, 78, 149–159. [Google Scholar] [CrossRef]
- Ren, X.; Yan, Z.; He, X.; Li, X.; Qin, B. Allelochemicals from rhizosphere soils of Glycyrrhiza uralensis Fisch: Discovery of the autotoxic compounds of a traditional herbal medicine. Ind. Crop. Prod. 2017, 97, 302–307. [Google Scholar] [CrossRef]
- Chon, S.U.; Kim, J.D. Biological activity and quantification of suspected allelochemicals from alfalfa plant parts. J. Agron. Crop. Sci. 2002, 188, 281–285. [Google Scholar] [CrossRef]
- Chon, S.U.; Jennings, J.A.; Nelson, C.J. Alfalfa (Medicago sativa L.) autotoxicity: Current status. Allelopath. J. 2006, 18, 58–80. [Google Scholar]
- Asaduzzaman, M.; Asao, T. Autotoxicity in beans and their allelochemicals. Sci. Hortic. 2012, 134, 26–31. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Isolation and identification of an allelopathic substance in Pisum sativum. Phytochemistry 2003, 62, 1141–1144. [Google Scholar] [CrossRef]
- Biswas, S.M.; Chakraborty, N. Strong sour tamarind flavor of methyl-2, 3, 4-trihydroxyhexanoate, a new compound isolated from leaves of Tamarindus indica L. plays a role in plant. Ann. Trop. Res. 2012, 34, 26–44. [Google Scholar] [CrossRef] [Green Version]
- Syed, S.; Ahmed, Z.I.; Al-Haq, M.I.; Mohammad, A.; Fujii, Y. The possible role of organic acids as allelochemicals in Tamarindus indica L. leaves. Acta Agric. Scand. 2014, 64, 511–517. [Google Scholar] [CrossRef]
- Solar, A.; Colarič, M.; Usenik, V.; Stampar, F. Seasonal variations of selected flavonoids, phenolic acids and quinones in annual shoots of common walnut (Juglans regia L.). Plant Sci. 2006, 170, 453–461. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, Z.; Hu, T.; Rehman, H.; Chen, H.; Li, Z.; Ding, B.; Hu, H. Allelopathic activity and chemical constituents of walnut (Juglans regia) leaf litter in walnut–winter vegetable agroforestry system. Nat. Prod. Res. 2014, 28, 2017–2020. [Google Scholar] [CrossRef]
- Chen, F.; Peng, S.; Chen, B.; Ni, G.; Liao, H. Allelopathic potential and volatile compounds of Rosmarinus officinalis L. against weeds. Allelopath. J. 2013, 32, 57–66. [Google Scholar]
- Appiah, K.S.; Mardani, H.K.; Omari, R.A. Involvement of carnosic acid in the phytotoxicity of Rosmarinus officinalis leaves. Toxins 2018, 10, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladhari, A.; Gaaliche, B.; Zarrelli, A.; Ghannem, M.; Mimoun, M.B. Allelopathic potential and phenolic allelochemicals discrepancies in Ficus carica L. cultivars. S. Afr. J. Bot. 2020, 130, 30–44. [Google Scholar] [CrossRef]
- Puig, C.G.; Reigosa, M.J.; Valentão, P.; Andrade, P.B.; Pedrol, N. Unravelling the bioherbicide potential of Eucalyptus globulus Labill: Biochemistry and effects of its aqueous extract. PLoS ONE 2018, 13, e0192872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chon, S.U.; Kim, Y.M. Herbicidal potential and quantification of suspected allelochemicals from four grass crop extracts. J. Agron. Crop. Sci. 2004, 190, 145–150. [Google Scholar] [CrossRef]
- De Bertoldi, C.; De Leo, M.; Braca, A.; Ercoli, L. Bioassay-guided isolation of allelochemicals from Avena sativa L.: Allelopathic potential of flavone C-glycosides. Chemoecology 2009, 19, 169–176. [Google Scholar] [CrossRef]
- Kremer, R.J.; Ben-Hammouda, M. Allelopathic Plants. 19. Barley (Hordeum vulgare L.). Allelopath. J. 2009, 24, 225–242. [Google Scholar]
- Kong, C.H.; Li, H.B.; Hu, F.; Xu, X.H.; Wang, P. Allelochemicals released by rice roots and residues in soil. Plant Soil 2006, 288, 47–56. [Google Scholar] [CrossRef]
- Weston, L.A.; Alsaadawi, I.S.; Baerson, S.R. Sorghum allelopathy–From ecosystem to molecule. J. Chem. Ecol. 2013, 39, 142–153. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T. Allelopathy in wheat (Triticum aestivum). Ann. Appl. Biol. 2001, 139, 1–9. [Google Scholar] [CrossRef]
- Stochmal, A.; Kus, J.; Martyniuk, S.; Oleszek, W. Concentration of benzoxazinoids in roots of field-grown wheat (Triticum aestivum L.) varieties. J. Agric. Food Chem. 2006, 54, 1016–1022. [Google Scholar] [CrossRef]
- Fragasso, M.; Iannucci, A.; Papa, R. Durum wheat and allelopathy: Toward wheat breeding for natural weed management. Front. Plant Sci. 2013, 4, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.H.; Waller, G.R. Possible allelopathic constituents of Coffea arabica. J. Chem. Ecol. 1980, 6, 643–654. [Google Scholar] [CrossRef]
- Geng, G.D.; Zhang, S.Q.; Cheng, Z.H. Allelopathy and allelochemicals of root exudates in hot pepper. Acta Hortic. Sin. 2009, 36, 873–878. [Google Scholar]
- Sun, H.; Wang, Y. Potential allelopathic effects of allelochemicals in aqueous extracts of leaves and root exudates of Capsicum annuum on vegetable crops. Allelopath. J. 2015, 35, 11–22. [Google Scholar]
- Baek, J.M.; Kawecki, O.J.; Lubin, K.D.; Song, J.; Wiens, O.A.; Wu, F. Plant warfare: Allelopathic effects of Nicotiana tabacum on the germination of Vigna radiata and Triticum aestivum. WURJ Health Nat. Sci. 2017, 8. [Google Scholar] [CrossRef]
- Rial, C.; Gómez, E.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Ecological relevance of the major allelochemicals in Lycopersicon esculentum roots and exudates. J. Agric. Food Chem. 2018, 66, 4638–4644. [Google Scholar] [CrossRef] [PubMed]
- Rawat, L.S.; Maikhuri, R.K.; Bahuguna, Y.M.; Jha, N.K.; Phondani, P.C. Sunflower allelopathy for weed control in agriculture systems. J. Crop. Sci. Biotech. 2017, 20, 45–60. [Google Scholar] [CrossRef]
- Pandino, G.; Mauromicale, G. Globe artichoke and cardoon forms between traditional and modern uses. Acta Hortic. 2020, 1284, 1–18. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Pandino, G.; Onofri, A.; Mauromicale, G. Allelopathic effects of Cynara cardunculus L. leaf aqueous extracts on seed germination of some Mediterranean weed species. Ital. J. Agron. 2018, 13, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.; Pandino, G.; Restuccia, A.; Lombardo, S.; Pesce, G.R.; Mauromicale, G. Allelopathic potential of leaf aqueous extracts from Cynara cardunculus L. on the seedling growth of two cosmopolitan weed species. Ital. J. Agron. 2019, 14, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Dayan, F.E.; Rimando, A.M.; Pan, Z.; Baerson, S.R.; Gimsing, A.L.; Duke, S.O. Sorgoleone. Phytochemistry 2010, 71, 1032–1039. [Google Scholar] [CrossRef]
- Scavo, A.; Mauromicale, G. Integrated weed management in herbaceous field crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping system niches. Agron. J. 2005, 97, 322–332. [Google Scholar] [CrossRef]
- Buhler, D.D.; Liebman, M.; Obrycki, J.J. Theoretical and practical challenges to an IPM approach to weed management. Weed Sci. 2000, 48, 274–280. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Shahzad, M.; Farooq, M.; Jabran, K.; Hussain, M. Impact of different crop rotations and tillage systems on weed infestation and productivity of bread wheat. Crop. Prot. 2016, 89, 161–169. [Google Scholar] [CrossRef]
- Scherner, A.; Melander, B.; Kudsk, P. Vertical distribution and composition of weed seeds within the plough layer after eleven years of contrasting crop rotation and tillage schemes. Soil Tillage Res. 2016, 161, 135–142. [Google Scholar] [CrossRef]
- Hunt, N.D.; Hill, J.D.; Liebman, M. Reducing freshwater toxicity while maintaining weed control, profits, and productivity: Effects of increased crop rotation diversity and reduced herbicide usage. Environ. Sci. Technol. 2017, 51, 1707–1717. [Google Scholar] [CrossRef]
- Weisberger, D.; Nichols, V.; Liebman, M. Does diversifying crop rotations suppress weeds? A meta-analysis. PLoS ONE 2019, 14, e0219847. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.; Restuccia, A.; Abbate, C.; Mauromicale, G. Seeming field allelopathic activity of Cynara cardunculus L. reduces the soil weed seed bank. Agron. Sustain. Dev. 2019, 39, 41. [Google Scholar] [CrossRef]
- Karkanis, A.; Alexiou, A.; Katsaros, C.; Petropoulos, S. Allelopathic activity of spearmint (Mentha spicata L.) and peppermint (Mentha × piperita L.) reduces yield, growth, and photosynthetic rate in a succeeding crop of maize (Zea mays L.). Agronomy 2019, 9, 461. [Google Scholar] [CrossRef] [Green Version]
- Mauromicale, G.; Occhipinti, A.; Mauro, R. Selection of shade-adapted subterranean clover species for cover cropping in orchards. Agron. Sustain. Dev. 2010, 30, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Gerhards, R.; Schappert, A. Advancing cover cropping in temperate integrated weed management. Pest Manag. Sci. 2020, 76, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Mennan, H.; Jabran, K.; Zandstra, B.H.; Pala, F. Non-chemical weed management in vegetables by using cover crops: A review. Agronomy 2020, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Nadeem, M.A.; Tanveer, A.; Ali, H.H.; Farooq, N. Role of allelopathic crop mulches and reduced doses of tank-mixed herbicides in managing herbicide-resistant Phalaris minor in wheat. Crop. Prot. 2018, 110, 245–250. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Lombardo, S.; Fontanazza, S.; Abbate, C.; Pandino, G.; Anastasi, U.; Onofri, A.; Mauromicale, G. Improving soil health, weed management and nitrogen dynamics by Trifolium subterraneum cover cropping. Agron. Sustain. Dev. 2020, 40, 18. [Google Scholar] [CrossRef]
- Sturm, D.J.; Kunz, C.; Gerhards, R. Inhibitory effects of cover crop mulch on germination and growth of Stellaria media (L.) Vill., Chenopodium album L. and Matricaria chamomilla L. Crop. Prot. 2016, 90, 125–131. [Google Scholar] [CrossRef]
- Puig, C.G.; Revilla, P.; Barreal, M.E.; Reigosa, M.J.; Pedrol, N. On the suitability of Eucalyptus globulus green manure for field weed control. Crop. Prot. 2019, 121, 57–65. [Google Scholar] [CrossRef]
- Masilionyte, L.; Maiksteniene, S.; Kriauciuniene, Z.; Jablonskyte-Rasce, D.; Zou, L.; Sarauskis, E. Effect of cover crops in smothering weeds and volunteer plants in alternative farming systems. Crop. Prot. 2017, 91, 74–81. [Google Scholar] [CrossRef]
- Álvarez-Iglesias, L.; Puig, C.G.; Revilla, P.; Reigosa, M.J.; Pedrol, N. Faba bean as green manure for field weed control in maize. Weed Res. 2018, 58, 437–449. [Google Scholar] [CrossRef]
- Alonso-Ayuso, M.; Gabriel, J.L.; García-González, I.; Del Monte, J.P.; Quemada, M. Weed density and diversity in a long-term cover crop experiment background. Crop. Prot. 2018, 112, 103–111. [Google Scholar] [CrossRef]
- Blaise, D.; Manikandan, A.; Verma, P.; Nalayini, P.; Chakraborty, M.; Kranthi, K.R. Allelopathic intercrops and its mulch as an integrated weed management strategy for rainfed Bt-transgenic cotton hybrids. Crop. Prot. 2020, 135, 105214. [Google Scholar] [CrossRef]
- Pouryousef, M.; Yousefi, A.R.; Oveisi, M.; Asadi, F. Intercropping of fenugreek as living mulch at different densities for weed suppression in coriander. Crop. Prot. 2015, 69, 60–64. [Google Scholar] [CrossRef]
- Cheriere, T.; Lorin, M.; Corre-Hellou, G. Species choice and spatial arrangement in soybean-based intercropping: Levers that drive yield and weed control. Field Crop. Res. 2020, 256, 107923. [Google Scholar] [CrossRef]
- Vrignon-Brenas, S.; Celette, F.; Piquet-Pissaloux, A. Intercropping strategies of white clover with organic wheat to improve the trade-off between wheat yield, protein content and the provision of ecological services by white clover. Field Crop. Res. 2018, 224, 160–169. [Google Scholar] [CrossRef]
- Lemessa, F.; Wakjira, M. Mechanisms of ecological weed management by cover cropping: A review. J. Biol. Sci. 2014, 14, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Restuccia, A.; Scavo, A.; Lombardo, S.; Pandino, G.; Fontanazza, S.; Anastasi, U.; Abbate, C.; Mauromicale, G. Long-term effect of cover crops on species abundance and diversity of weed flora. Plants 2020, 9, 1506. [Google Scholar] [CrossRef] [PubMed]
- Westerman, P.R.; Luijendijk, C.D.; Wevers, J.D.A.; van der Werf, W. Weed seed predation in a phenologically late crop. Weed Res. 2011, 51, 157–164. [Google Scholar] [CrossRef]
- Kruidhof, H.M.; Gallandt, E.R.; Haramoto, E.R.; Bastiaans, L. Selective weed suppression by cover crop residues: Effect of seed mass and timing of species sensitivity. Weed Res. 2011, 51, 177–186. [Google Scholar] [CrossRef]
- Kruidhof, H.M.; Bastiaans, L.; Kropff, M.J. Cover crop residue management for optimizing weed control. Plant Soil 2009, 318, 169–184. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef]
- Barto, E.K.; Weidenhamer, J.D.; Cipollini, D.; Rillig, M.C. Fungal superhighways: Do common mycorrhizal networks enhance below ground communication? Trends Plan Sci. 2012, 17, 633–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rad, S.V.; Valadabadi, S.A.R.; Pouryousef, M.; Saifzadeh, S.; Zakrin, H.R.; Mastinu, A. Quantitative and qualitative evaluation of Sorghum bicolor L. under intercropping with legumes and different weed control methods. Horticulturae 2020, 6, 78. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Emeran, A.A.; Rubiales, D. Inter-cropping with berseem clover (Trifolium alexandrinum) reduces infection by Orobanche crenata in legumes. Crop. Prot. 2010, 29, 867–871. [Google Scholar] [CrossRef] [Green Version]
- Florence, A.M.; Higley, L.G.; Drijber, R.A.; Francis, C.A.; Lindquist, J.L. Cover crop mixture diversity, biomass productivity, weed suppression, and stability. PLoS ONE 2019, 14, e0206195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, C.; Sturm, D.J.; Varnholt, D.; Walker, F.; Gerhards, R. Allelopathic effects and weed suppressive ability of cover crops. Plant Soil Environ. 2016, 62, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Sturm, D.J.; Peteinatos, G.; Gerhards, R. Contribution of allelopathic effects to the overall weed suppression by different cover crops. Weed Res. 2018, 58, 331–337. [Google Scholar] [CrossRef]
- Hoagland, R.E.; Boyette, C.D.; Weaver, M.A.; Abbas, H.K. Bioherbicides: Research and risks. Toxin Rev. 2007, 26, 313–342. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a natural products approach to herbicide discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowska, A. Allelochemicals as bioherbicides—Present and perspectives. In Herbicides—Current Research and Case Studies in Use; Price, A.J., Kelton, J.A., Eds.; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Jamil, M.; Cheema, Z.A.; Mushtaq, M.N.; Farooq, M.; Cheema, M.A. Alternative control of wild oat and canary grass in wheat fields by allelopathic plant water extracts. Agron. Sustain. Dev. 2009, 29, 475–482. [Google Scholar] [CrossRef]
- Anjum, T.; Bajwa, R. The effect of sunflower leaf extracts on Chenopodium album in wheat fields in Pakistan. Crop. Prot. 2007, 26, 1390–1394. [Google Scholar] [CrossRef]
- Ullah, R.; Aslam, Z.; Maitah, M.; Bashir, S.; Hassan, W. Sustainable weed control and enhancing nutrient use efficiency in crops through brassica (Brassica campestris L.) allelopathy. Sustainability 2020, 12, 5763. [Google Scholar] [CrossRef]
- Carrubba, A.; Labruzzo, A.; Comparato, A.; Muccilli, S.; Spina, A. Use of plant water extracts for weed control in durum wheat (Triticum turgidum L. subsp. durum Desf.). Agronomy 2020, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Caser, M.; Demasi, S.; Caldera, F.; Dhakar, N.K.; Trotta, F.; Scariot, V. Activity of Ailanthus altissima (Mill.) Swingle extract as a potential bioherbicide for sustainable weed management in horticulture. Agronomy 2020, 10, 965. [Google Scholar] [CrossRef]
- Barros, V.M.D.S.; Pedrosa, J.L.F.; Gonçalves, D.R.; De Medeiros, F.C.L.; Carvalho, G.R.; Gonçalves, A.H.; Teixeira, P.V.V.Q. Herbicides of biological origin: A review. J. Hortic. Sci. Biotech. 2021, 96, 288–296. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Alqarawi, A.A.; Abd Allah, E.F. Bioherbicides: Current knowledge on weed control mechanism. Ecotoxicol. Environ. Saf. 2018, 158, 131–138. [Google Scholar] [CrossRef]
- Alsaadawi, I.S.; Khaliq, A.; Farooq, M. Integration of allelopathy and less herbicides effect on weed management in field crops and soil biota: A Review. Plant Arch. 2020, 20, 225–237. [Google Scholar]
- Asaduzzaman, M.; Pratley, J.E.; An, M.; Luckett, D.J.; Lemerle, D. Metabolomics differentiation of canola genotypes: Toward an understanding of canola allelochemicals. Front. Plant Sci. 2015, 5, 765. [Google Scholar] [CrossRef] [Green Version]
- Ghimire, B.K.; Yu, C.Y.; Ghimire, B.; Seong, E.S.; Chung, I.M. Allelopathic potential of phenolic compounds in Secale cereale cultivars and its relationship with seeding density. Appl. Sci. 2019, 9, 3072. [Google Scholar] [CrossRef] [Green Version]
- Maver, M.; Miras-Moreno, B.; Lucini, L.; Trevisan, M.; Pii, Y.; Cesco, S.; Mimmo, T. New insights in the allelopathic traits of different barley genotypes: Middle Eastern and Tibetan wild-relative accessions vs. cultivated modern barley. PLoS ONE 2020, 15, e0231976. [Google Scholar] [CrossRef]
- Scavo, A.; Rial, C.; Varela, R.M.; Molinillo, J.M.G.; Mauromicale, G.; Macías, F.A. Influence of genotype and harvest time on the Cynara cardunculus L. sesquiterpene lactone profile. J. Agric. Food Chem. 2019, 67, 6487–6496. [Google Scholar] [CrossRef]
- Reigosa, M.J.; Souto, X.C.; González, L. Effect of phenolic compounds on the germination of six weeds species. Plant Growth Regul. 1999, 28, 83–88. [Google Scholar] [CrossRef]
- Stanišić, M.; Ćosić, T.; Savić, J.; Krstić-Milošević, D.; Mišić, D.; Smigocki, A.; Ninković, S.; Banjac, N. Hairy root culture as a valuable tool for allelopathic studies in apple. Tree Physiol. 2019, 39, 888–905. [Google Scholar] [CrossRef]
- Duke, S.O.; Scheffler, B.E.; Boyette, C.D.; Dayan, F.E. Biotechnology in Weed Control. Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Shehzad, T.; Okuno, K. Genetic analysis of QTLs controlling allelopathic characteristics in sorghum. PLoS ONE 2020, 15, e0235896. [Google Scholar] [CrossRef] [PubMed]
- De la Calle, M.E.; Cabrera, G.; Cantero, D.; Valle, A.; Bolivar, J. A genetically engineered Escherichia coli strain overexpressing the nitroreductase NfsB is capable of producing the herbicide D-DIBOA with 100% molar yield. Microb. Cell Factories 2019, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Oueslati, O.; Ben-Hammouda, M.; Ghorbal, M.H.; Guezzah, M.; Kremer, R.J. Barley autotoxicity as influenced by varietal and seasonal variation. J. Agron. Crop. Sci. 2005, 191, 249–254. [Google Scholar] [CrossRef]
- Scavo, A.; Rial, C.; Molinillo, J.M.G.; Varela, R.M.; Mauromicale, G.; Macías, F.A. Effect of shading on the sesquiterpene lactone content and phytotoxicity of cultivated cardoon leaf extracts. J. Agric. Food Chem. 2020, 68, 11946–11953. [Google Scholar] [CrossRef]
- Xuan, T.D.; Minh, T.N.; Khanh, T.D. Allelopathic momilactones A and B are implied in rice drought and salinity tolerance, not weed resistance. Agron. Sustain. Dev. 2016, 36, 52. [Google Scholar] [CrossRef]
- Tranel, P.J.; Horvath, D.P. Molecular biology and genomics: New tools for weed science. Bioscience 2009, 59, 207–215. [Google Scholar] [CrossRef]
- Frey, M.; Chomet, P.; Glawischnig, E.; Stettner, C.; Grün, S.; Winklmair, A.; Eisenreich, W.; Bacher, A.; Meeley, R.B.; Briggs, S.P.; et al. Analysis of a chemical plant defense mechanism in grasses. Science 1997, 277, 696–699. [Google Scholar] [CrossRef]
- Jensen, L.B.; Courtois, B.; Shen, L.S.; Li, Z.K.; Olofsdotter, M.; Mauleon, R.P. Locating genes controlling allelopathic effects against barnyardgrass in upland rice. Agron. J. 2001, 93, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Pratley, J.; Ma, W.; Haig, T. Quantitative trait loci and molecular markers associated with wheat allelopathy. Theor. Appl. Genet. 2003, 107, 1477–1481. [Google Scholar] [CrossRef]
- Scaglione, D.; Reyes-Chin-Wo, S.; Acquadro, A. The genome sequence of the outbreeding globe artichoke constructed de novo incorporating a phase-aware low-pass sequencing strategy of F1 progeny. Sci. Rep. 2016, 6, 19427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eljounaidi, K.; Cankar, K.; Comino, C.; Moglia, A.; Hehn, A.; Bourgaud, F.; Bouwmeester, H.; Menin, B.; Lanteri, S.; Beekwilder, J. Cytochrome P450s from Cynara cardunculus L. CYP71AV9 and CYP71BL5, catalyze distinct hydroxylations in the sesquiterpene lactone biosynthetic pathway. Plant Sci. 2014, 223, 59–68. [Google Scholar] [CrossRef] [PubMed]


| Botanical Family | Binomial Name | Allelochemicals | References |
|---|---|---|---|
| Amaryllidaceae | Allium cepa L. | S-containing compounds (alliin, isoalliin, methiin, allicin, ajoene, sulfenic acid, methyl propenyl disulfate, methylpropyl trisulfate) and phenolic acids (ferulic, p-coumaric, p-hydroxybenzoic, syringic, vanillic) | [17] |
| A. sativum | |||
| A. ursinum | |||
| Asteraceae | Artemisia absinthium L. | Tannins, terpenes and alkaloids (absinthium) | [18] |
| Carthamus tinctorius L. | Sesquiterpene lactones (dehydrocostuslactone, costunolide) and strigolactones (solanacol, GR24 and abacyl acetate) | [19] | |
| Cichorium intybus L. | Sesquiterpene lactones (8α-angeloyloxycichoralexin, lactupicrin) and guaianolides (cichoralexin, 10α-hydroxycichopumilide) | [20] | |
| Cynara cardunculus L. | Sesquiterpene lactones (cynaropicrin, deacylcynaropicrin, 11,13-dihydro-deacylcynaropicrin, grosheimin, 11,13-dihydroxi-8-deoxygrosheimin, aguerin B, cynaratriol), pinoresinol and polyphenols (caffeoylquinic and dicaffeoylquinic acids, luteolin and apigenin derivatives) | [21,22,23] | |
| Helianthus annuus L. | Sesquiterpene lactones (helivypolide D, leptocarpin, helivypolide E, annuolide F, annuolide H, helivypolides F, helivypolides H, helivypolides J, helieudesmanolide A, 8β-angeloiloxicumambranolide), heliannuoles (heliannuol J), bisnorsesquiterpenes (annuionone D, (+)-dehydrovomifoliol), flavonoids (heliannone A, kukulkanine B, heliannone B, tambuline) and (+)-loliolide | [24] | |
| H. tuberosus | Sesquiterpene lactones (1,10-epoxidized heliangolides, 1-keto-2,3-unsaturated-furanoheliangolides, 4,15-isoatriplicolide angelate, 4,15-isoatriplicolide methylacrylate), diterpenes (ent-17-oxokaur-15(16)-en-19-oic acid, ent-17-hydroxykaur-15(16)-en-19-oic acid, ent-15β-hydroxykaur-16(17)-en-19-oic acid methyl ester and ent-15-nor-14-oxolabda-8(17),12E-dien-18-oic acid), phenolic compounds (p-hydroxybenzoic acid, p-hydroxybenzaldehyde, salicylic acid, coumarin, o-coumarinic acid, and p-coumaric acid) and (+)- pinoresinol | [25,26] | |
| Lactuca sativa L. | Phenolic acids (coumarin, trans-cinnamic acid, o-coumaric acid, p-coumaric acid and chlorogenic acid) | [27] | |
| Brassicaceae | Brassica juncea (L.) Czern. | Glucosinolates [isothiocyanates (allyl-ITC, 2-phenylethyl, 3-butenyl, 4-pentenyl, 4-methylthiobutyl, 5-methylthiopentyl), nitriles (5-methylthiopentanenitrile, 6-methylthiohexanenitrile), oxazolidinethione (goitrin)] and brassinosteroids (brassinolide, 24-epibrassinolide, 28-homobrassinolide) | [28] |
| B. napus | |||
| B. nigra | |||
| B. oleracea | |||
| Capparis spinosa L. | Flavonoids (quercetin-3-O-β-D-glucopyranoside, quercetin, kaempferol 3-O-β-D-glucopyranoside, rhamnetin, isorhamnetin, rhamnozin, thomnocitirin), carotenoids (β-carotene, lutein, neoxanthin and violaxanthin), tocopherols (α- and γ-tocopherol) and glucosinates | [29,30] | |
| Cannabaceae | Cannabis sativa L. | Glycosides, alkaloids, flavonoids, flavones, steroids, tannins, phenols and saponins | [31] |
| Convolvulaceae | Ipomea batatas (L.) Lam. | Polyphenols (coumarin, trans-cinnamic acid, hydroxycinnamic acid, p- and o-coumaric acid, caffeic acid, chlorogenic acid) | [32] |
| Cucurbitaceae | Citrullus lanatus (Thunb.) Matsum and Nakai | Phenolic acids (ρ-hydrobenzoic, vanillic, syringic, ρ-coumaric and frulic acids) | [33] |
| Cucurbitaceae | Cucumis sativus L. | Phenolic acids (benzoic, p-hydroxybenzoic, 2,5-dihydroxybenzoic, 3-phenylpropionic, cinnamic, p-hydroxycinnamic, gallic, vanillic, caffeic, hydrocaffeic, p-coumaric, ferulic, sinapic, p-thiocyanatophenol and 2-hydroxybenzothiazole) and fatty acids (myristic, palmitic, and stearic) | [34] |
| Fabaceae | Arachis hypogaea L. | Phenolic acids (p-coumaric and benzoic) and fatty acids (tetradecanoic, hexadecanoic, octadecanoic) | [35] |
| Glycyrrhiza uralensis Fisch. | Flavonones (liquiritin, isoliquiritigenin) and triterpenes (glycyrrhizic acid, dodecanoic acid) | [36] | |
| Medicago sativa L. | Phenolic compounds (salicylic acid, p-hydroxybenzoic acid, trans-cinnamic acid, o-coumaric acid, p-coumaric acid, ferulic acid, vanillic acid, chlorogenic acid, caffeic acid, coumarin, rutin, quercetin, scopoletin, medicarpin, sativan, 4-methoxymedicarpin, 5-methoxy-sativan) | [37,38] | |
| Phaseolus vulgaris L. | Phenolic acids (benzoic, salicylic, and malonic) | [39] | |
| Pisum sativum L. | Phenolic acids (benzoic, cinnamic, p-hydroxybenzoic, 3,4-dihydrobenzoic, vanillic, p-coumaric, sinapic) and pisatin | [40] | |
| Vicia faba L. | Phenolic acids (lactic, benzoic, p-hydroxybenzoic, vanillic, adipic, succinic, malic, glycolic, p-hydroxyphenylacetic, salicylic) | [39] | |
| Tamarindus indica L. | Phenolic acids (caffeic), methyl-2,3,4-trihydroxyhexanoate and organic acids (citric, malic, oxalic, and tartaric) | [41,42] | |
| Juglandaceae | Juglans nigra L. | Naphthoquinones (juglone, 1,4-naphthoquinone, plumbagin, 2-methyl-1,4-naphthoquinone), triterpenoids (lupenone, lupeol, squalene) fatty acids (n-hexadecanoic, 9,12-octadecadienoic, 8-octadecenoic, palmitic, stearic), phenolic acids (chlorogenic, p-coumaric, o-coumaric, ferulic, tannic, caffeic, vanillic, syringic), flavonoides (catechin, epicatechin and myricetin), flavonoids (quercetin and quercetin derivatives), hydroxybenzoic acids (gallic, ellagic, protocatechuic), and steroids (γ-sitosterol, sitostenone) | [43,44] |
| J. regia | |||
| Labiateae | Rosmarinus officinalis Schleid. | Monoterpenoids (α-pinene, myrcene, α-terpinene, β-cymene, 1,8-cineole, camphene, α-limonene, sabinene) and polyphenols (caffeic, ferulic, gallic, rosmarinic, carnosic, and chlorogenic acids) | [45,46] |
| Moraceae | Ficus carica L. | Phenolic acids (3-O-caffeoylquinic acid, 5-O-caffeoylquinic acid, dihydroxybenzoic acid, caffeoylmalic acid) and flavonoids (rutin, isoquercetin, catechin and astragalin) | [47] |
| Myrtaceae | Eucalyptus globulus Labill. | Phenolic compounds (chlorogenic acid, ρ-coumaric acid, ellagic acid, hyperoside, rutin, quercitrin and kaempferol 3-O-glucoside) and organic acids (citric, malic, shikimic, succinic and fumaric) | [48] |
| E. urograndis | |||
| E. urophylla | |||
| Poaceae | Avena sativa L. | Flavonoids (2-O-glucoside, isovitexin 2’’-O-arabinoside), phenolic acids (caffeic, ferulic, coumaric, salicylic, coumarin, cynnamic and derivatives) and saponins (avenacoside A, avenacoside B, 26-desglucoavenacoside A, 26-desglucoavenacoside B) | [49,50] |
| Poaceae | Hordeum vulgare L. | Phenolic acids (benzoic, caffeic, chlorogenic, coumaric, coumarin, ferulic, p-hydroxybenzoic, ferulic, gentisic, salicylic, sinapic, syringic, vanillic, cinnamic, hydroxycinnamic, scopoletin), benzoxazinoids (DIBOA, DIMBOA), alkaloids (gramine, hordenine), flavonoids (saponarin, apigenin, lutonarin, catechin, cyanidin, isovitexin) and cyanoglucosides (heterodendrin, epidermin, epiheterodendrin, sutherlandin, osmaronin, dihydroosmaronin) | [51] |
| Oryza sativa L. | Diterpenes (momilactones and oryzalexins), phenolic acids (caffeic, ferulic, coumaric, salicylic, syringic, p-hydroxybenzoic, coumarin, cynnamic and derivatives), flavones (5,7,4′-trihydroxy-3′,5′-dimethoxyflavone) and cyclohexenones (3-isopropyl-5-acetoxycyclohexene-2-one-1) | [13,49,52] | |
| Secale cereale L. | Benzoxazinoids (DIBOA, HMBOA, BOA, DIMBOA, MBOA) and phenolic acids (caffeic, ferulic, coumaric, salicylic, coumarin, cynnamic and derivatives) | [49] | |
| Sorghum bicolor (L.) Moench S. bicolor × S. sudanense S. halepense | Benzoxazinoids (DIBOA, HMBOA, BOA, DIMBOA, MBOA), benzoquinones (sorgoleone), cyanogenic glycosides (dhurrin), phenolic acids (p-hydroxybenzoic, p-hydroxybenzaldehyde, coumaric, ferulic) | [53] | |
| Triticum aestivum L. T. durum | Benzoxazinoids (DIBOA, DIMBOA-Glc, HMBOA, BOA, DIMBOA, MBOA), phenolic acids (trans-ρ-coumaric, cis-ρ-coumaric ferulic, vanillic, syringic, ρ-hydroxybenzoic), fatty acids (acetic, propionic and butyric), triterpenoids (cycloart-5-ene-3β,25-diol and cycloart-3β,25-diol) and steroids (cholesterol, ergosterol, campesterol, stigmasterol, sitosterol, spinasterol and stigmastanol) | [54,55,56] | |
| Rubiaceae | Coffea arabica L. | Alkaloids (caffeine, theobromine, theophylline, paraxanthine), coumarins (scopoletin) and phenolic acids (chlorogenic, ferulic, p-coumaric, p-hydroxybenzoic, caffeic and vanillic) | [57] |
| Solanaceae | Capsicum annuum L. | Phthalate esters (N-phenyl-2-naphthylamine, dibutyl phthalate, butyl cyclohexyl ester), dicarboxylic acids (phthalic acid, 1,2-benzenedicarboxylic acid) and phenols anilines (diphenylamine, 4,4′-(1-methylethylidene) bis-phenol, 1-naphthalenamine, n-phenyl-1-naphthylamine) | [58,59] |
| Nicotiana tabacum L. | Alkaloids (nicotine), sucrose esters (fluoro derivatives of sucrose) and diterpenes (duvatrienediol and duvatrienediol derivatives) | [60] | |
| Solanum lycopersicum L. | Alkaloids (α-tomatine), steroids (stigmasterol), furocoumarins (bergapten) and strigolactones (7-oxoorobanchylacetate, solanacol, orobanchol, strigol, fabacyl acetate, orobanchyl acetate and 5-deoxysrtrigol) | [61] |
| Cover Cropping Type | Allelopathic Cover Crop | Main Crop | Target Weeds | References |
|---|---|---|---|---|
| Dead mulching | Helianthus annuus L., Zea mays L., Oryza sativa L., Sorghum bicolor L. | Wheat | Phalaris minor Retz. | [80] |
| Trifolium subterraneum L. | Apricot | Several monocots and dicots | [81] | |
| Fagopyrum esculentum Moench., Sinapis alba L., T. subterraneum, H. annuus, Linum usitatissimum L., Raphanus sativus L., Vicia sativa L., Avena strigosa Schreb., Cannabis sativa L. and mixtures | Sugar beet | Stellaria media (L.) Vill., Chenopodium album L., Matricaria chamomilla L. | [82] | |
| Green manuring | Eucalyptus globulus Labill. | Corn | Digitaria sanguinalis (L.) Scop., C. album | [83] |
| S. alba and S. alba + F. esculentum | Red clover, wheat, pea, barley rotation with red clover as a undersown crop | Cirsium arvense (L.) Scop., Sonchus arvensis L., Galium aparine L., Lamium purpureum L., Fallopia convolvulus (L.) Á. Löve, C. album, S. media | [84] | |
| V. faba | Corn | A. retroflexus, C. album, Solanum nigrum, D. sanguinalis, Cyperus rotundus L. | [85] | |
| Hordeum vulgare L., V. sativa | Corn, sunflower | Xanthium spinosum L. and other broadleaf species | [86] | |
| Intercropping | Crotalaria juncea L. | Cotton | C. rotundus, Alternanthera paronychioides A. St.-Hil. | [87] |
| Trigonella foenum-graecum L. | Coriander | Several monocots and dicots | [88] | |
| F. esculentum, Lens culinaris Medik., S. bicolor, H. annuus | Soybean | C. album, Polygonum persicaria L. | [89] | |
| T. repens | Wheat | A. fatua, S. media, M. recutica | [90] |
| Donor Plant (Dose) | Extract Concentration | Main Crop (Yield) | Target Weeds | Weed Control | References |
|---|---|---|---|---|---|
| Sorghum (0.0006 L m−2) | 10% | Wheat (+39%) | Avena fatua L., Phalaris minor Retz. | −31% and −32% of DW | [106] |
| Sorghum + sunflower (0.0012 L m−2) | Wheat (+49.5%) | −52% and −45.5% of DW | |||
| Sorghum + sunflower (0.0006 L m−2) | Wheat (+62%) | −31.5% and −32.5% of DW | |||
| Sunflower (0.1 L m−2) | 10% | Wheat (no yield losses) | Chenopodium album L. | −70% of biomass | [107] |
| Chinese cabbage (0.002 L m−2) | 10% | Mung bean | Trianthema portulacastrum L., Cyperus rotundus L. | −14.6% of density and DW | [108] |
| Tree wormwood (4 L m−2) | 18.82% | Wheat (−52.9%) | Several monocots and dicots, mainly A. fatua and P. paradoxa | ~30% of weed suppression | [109] |
| Sicilian sumac (4 L m−2) | 8.75% | Wheat (+9%) | 50.8% of weed suppression | ||
| Common thyme (4 L m−2) | 22.33% | Wheat (−7.2%) | ~35% of weed suppression | ||
| Common lantana (4 L m−2) | 6.14% | Wheat (+16.5%) | 16% of weed suppression | ||
| Mediterranean spurge (4 L m−2) | 2.27% | Wheat (−2.3%) | ~40% of weed suppression | ||
| Tree of heaven (0.001–0.002 g L−1) | 20% | Sage, rosemary, carnation | Lepidium sativum L., Raphanus sativus L. | 0% weed presence in sage and rosemary, ~24% in carnation | [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scavo, A.; Mauromicale, G. Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy 2021, 11, 2104. https://doi.org/10.3390/agronomy11112104
Scavo A, Mauromicale G. Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy. 2021; 11(11):2104. https://doi.org/10.3390/agronomy11112104
Chicago/Turabian StyleScavo, Aurelio, and Giovanni Mauromicale. 2021. "Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future" Agronomy 11, no. 11: 2104. https://doi.org/10.3390/agronomy11112104
APA StyleScavo, A., & Mauromicale, G. (2021). Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy, 11(11), 2104. https://doi.org/10.3390/agronomy11112104