
A New Bacillus velezensis Strain CML532 Improves Chicken Growth Performance and Reduces Intestinal Clostridium perfringens Colonization



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Identification of Bacillus Strains
2.2. Complete Genome Sequencing and Analysis of Bacillus velezensis CML532
2.3. Fermentation and Preparation of Bacterial Powder
2.4. Dietary Supplementation of B. velezensis CML532 in Chickens under Normal Feeding Conditions
2.5. Dietary Supplementation of B. velezensis CML532 under the C. perfringens Challenge Condition in Chickens
2.6. Bird Management
2.7. Organ Index
2.8. Serum Parameter Assay
2.9. Intestinal Morphological Structure
2.10. Intestinal Lesion Score
2.11. Intestinal Disaccharidase Activity Measurement
2.12. Total RNA Isolation and Quantitative Real-Time PCR
2.13. DNA Extraction and Quantification, Amplicon Sequencing, and Data Processing
2.14. DNA Extraction and Enumeration of Intestinal C. perfringens
2.15. Statistical Analysis
3. Results
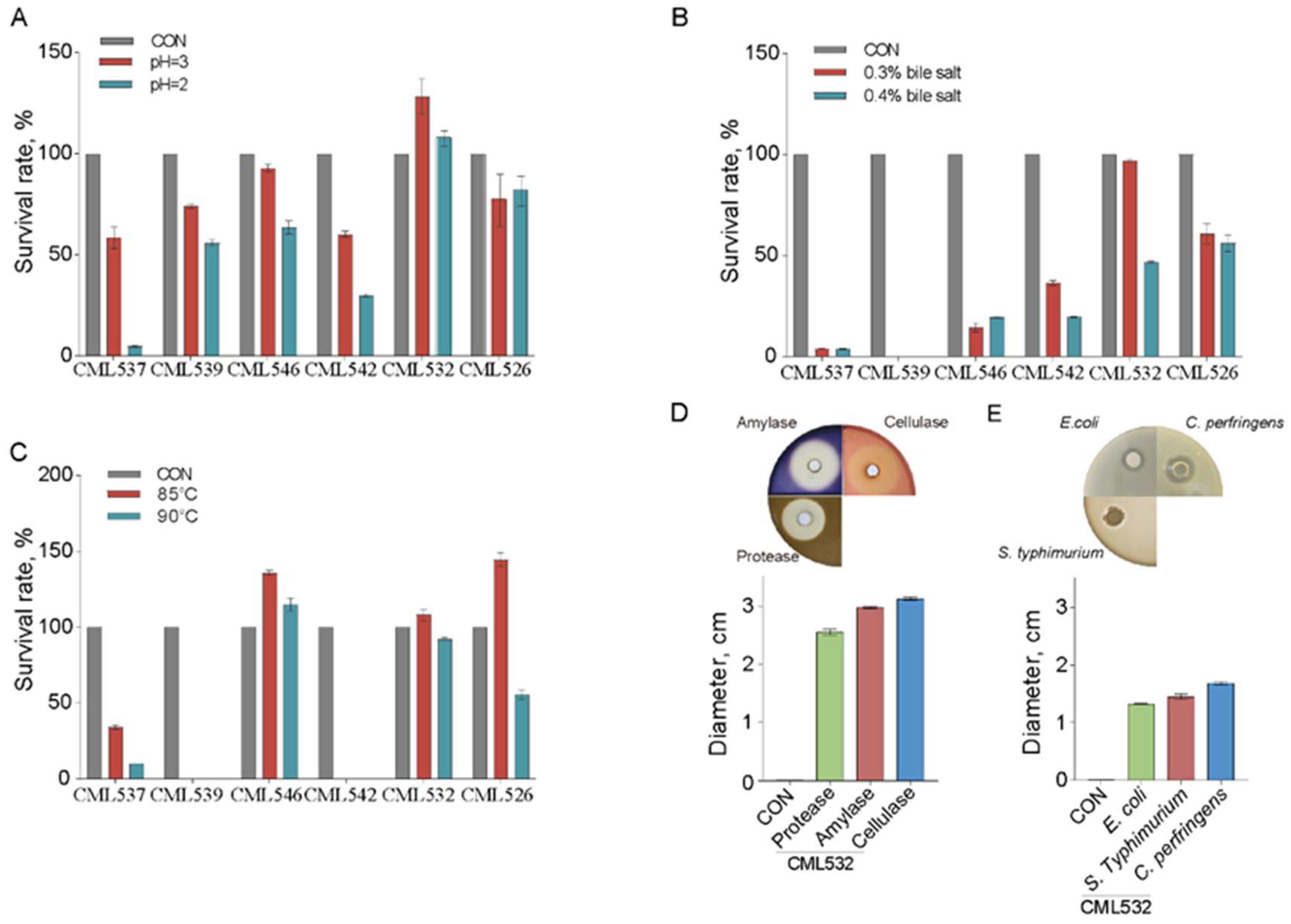
3.1. Isolation and Characterization of B. velezensis Strains from Chinese Local-Breed Chickens
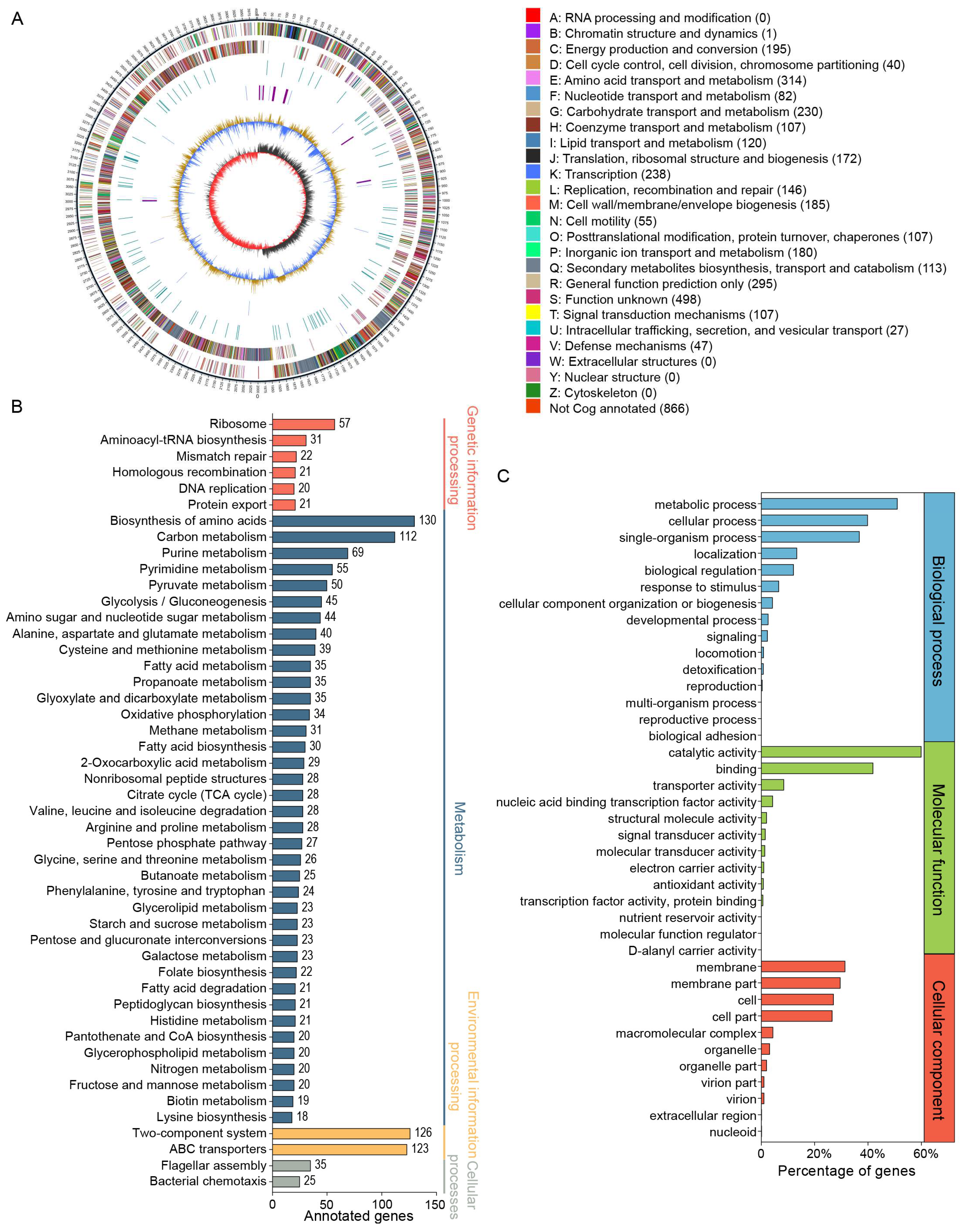
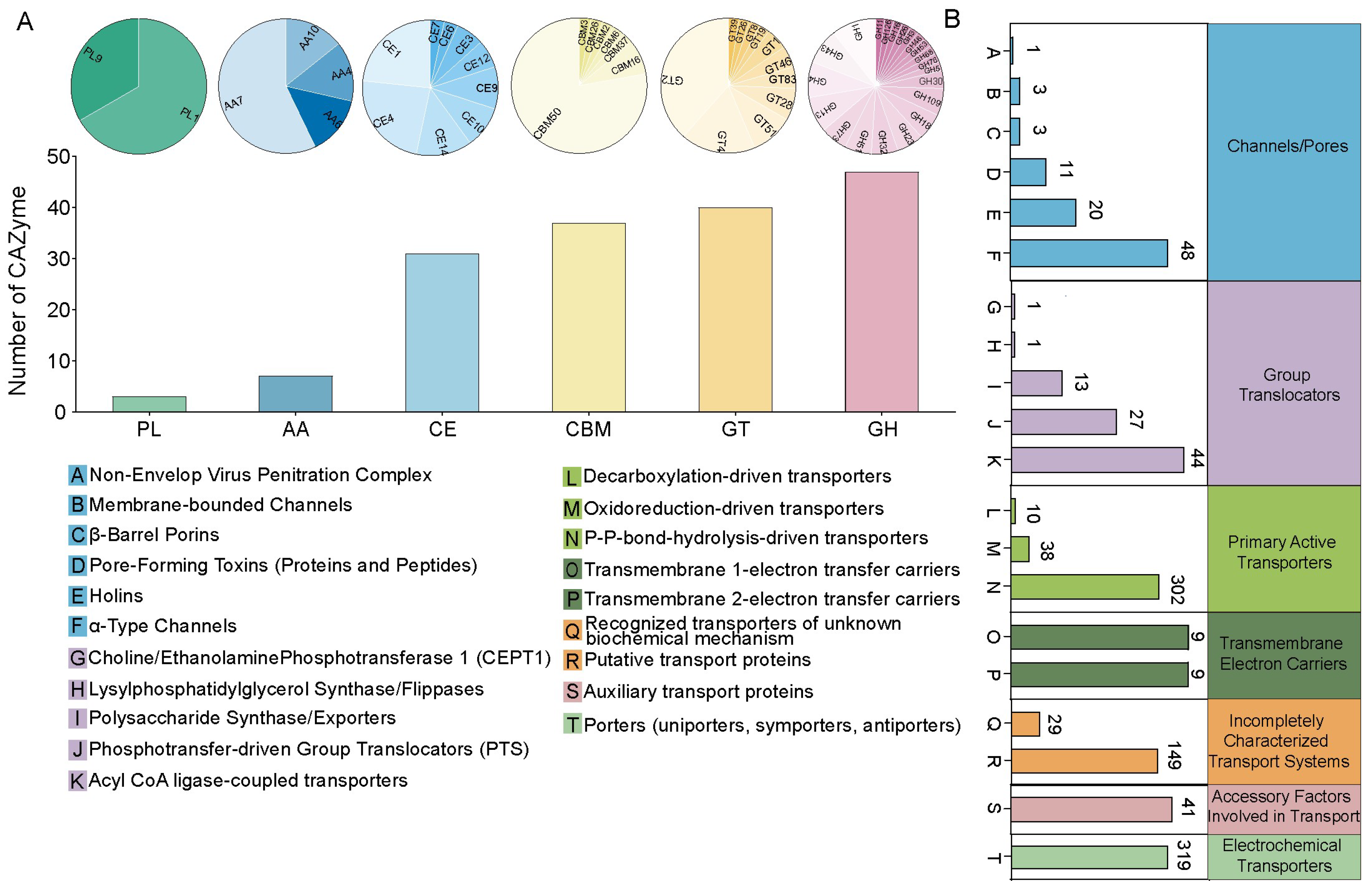
3.2. Genomic Analysis of B. velezensis CML532
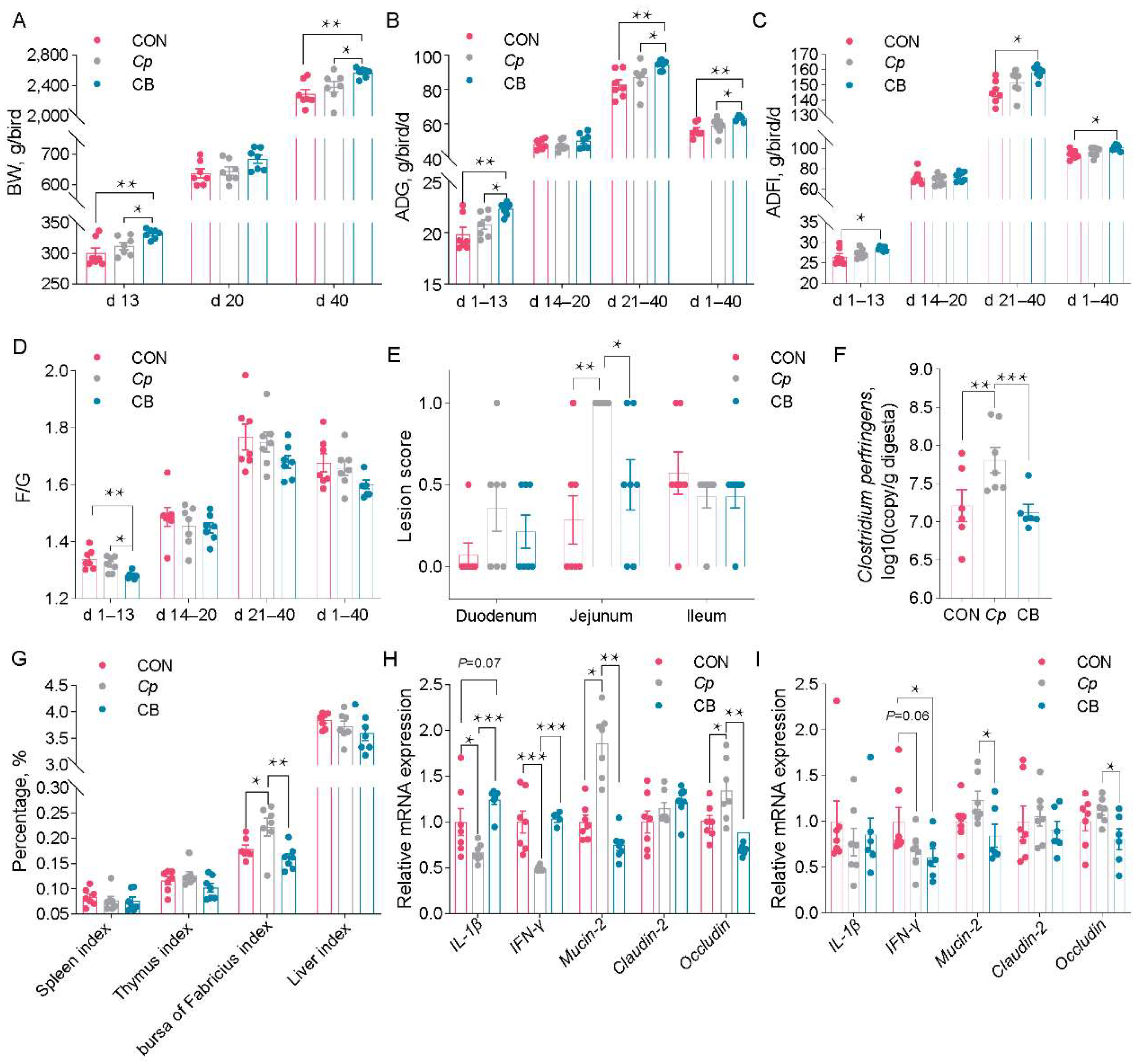
3.3. B. velezensis CML532 Improves the Chicken’s Growth Performance and Gut Function
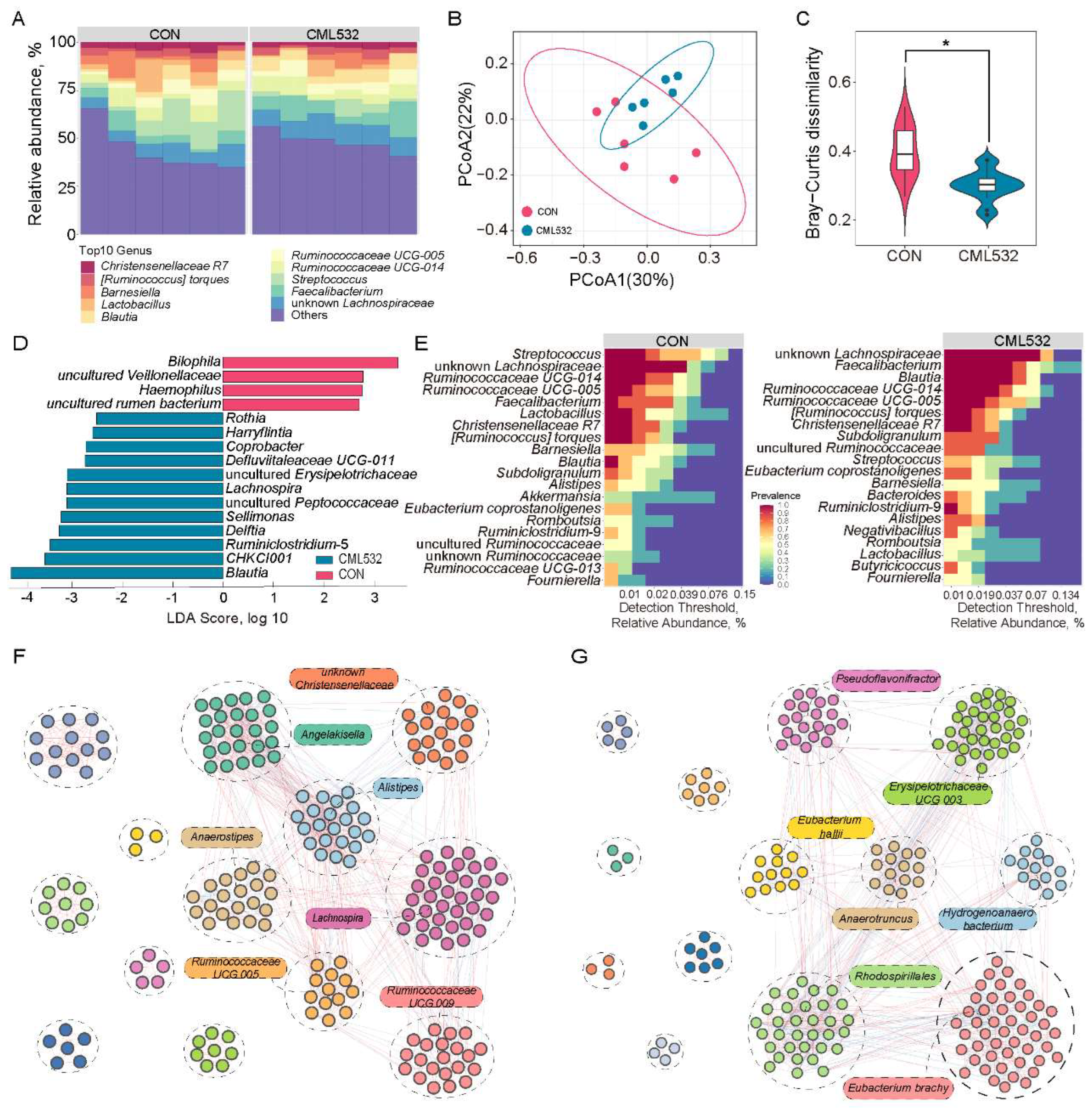
3.4. B. velezensis CML532 Alters the Chicken Cecal Microbial Community Structure
3.5. B. velezensis CML532 Suppresses Intestinal C. perfringens Colonization and Attenuates Intestinal Mucosal Injury
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agus, A.; Planchais, J.; Sokol, H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Ramlucken, U.; Roets, Y.; Ramchuran, S.O.; Moonsamy, G.; van Rensburg, C.J.; Thantsha, M.S.; Lalloo, R. Isolation, selection and evaluation of bacillus spp. As potential multi-mode probiotics for poultry. J. Gen. Appl. Microbiol. 2020, 66, 228–238. [Google Scholar] [CrossRef]
- Ahiwe, E.U.; Tedeschi Dos Santos, T.T.; Graham, H.; Iji, P.A. Can probiotic or prebiotic yeast (Saccharomyces cerevisiae) serve as alternatives to in-feed antibiotics for healthy or disease-challenged broiler chickens?: A review. J. Appl. Poult. Res. 2021, 30, 100164. [Google Scholar] [CrossRef]
- Xu, C.; Wei, F.; Yang, X.; Feng, Y.; Liu, D.; Hu, Y. Lactobacillus salivarius cml352 isolated from chinese local breed chicken modulates the gut microbiota and improves intestinal health and egg quality in late-phase laying hens. Microorganisms 2022, 10, 726. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.; Ibrahim, H.I.M. Enterococcus faecium from chicken feces improves chicken immune response and alleviates Salmonella infections: A pilot study. J. Anim. Sci. 2023, 101, skad016. [Google Scholar] [CrossRef]
- Hernández-Granados, M.J.; Ortiz-Basurto, R.I.; Jiménez-Fernández, M.; García-Munguía, C.A.; Franco-Robles, E. Dietary encapsulated bifidobacterium animalis and agave fructans improve growth performance, health parameters, and immune response in broiler chickens. Anim. Biosci. 2022, 35, 587–595. [Google Scholar] [CrossRef]
- Wolfenden, R.E.; Pumford, N.; Morgan, M.J.; Shivaramaiah, S.; Wolfenden, A.D.; Tellez, G. Evaluation of a screening and selection method for bacillus isolates for use as effective direct-fed microbials in commercial poultry. Int. J. Poult. Sci. 2010, 9, 317–323. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.H. Bacillus velezensis: A valuable member of bioactive molecules within plant microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [PubMed]
- Khalid, F.; Khalid, A.; Fu, Y.; Hu, Q.; Zheng, Y.; Khan, S.; Wang, Z. Potential of Bacillus velezensis as a probiotic in animal feed: A review. J. Microbiol. 2021, 59, 627–633. [Google Scholar] [CrossRef]
- Soni, R.; Keharia, H.; Bose, A.; Pandit, N.; Doshi, J.; Rao, S.V.R.; Paul, S.S.; Raju, M.V.L.N. Genome assisted probiotic characterization and application of Bacillus velezensis zbg17 as an alternative to antibiotic growth promoters in broiler chickens. Genomics 2021, 113, 4061–4074. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, S.; Dang, G.; Jia, R.; Chen, S.; Deng, X.; Liu, G.; Beckers, Y.; Cai, H. Screening and characterization of Bacillus velezensis lb-y-1 toward selection as a potential probiotic for poultry with multi-enzyme production property. Front. Microbiol. 2023, 14, 1143265. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Wei, C.; Khalid, A.; Hu, Q.; Yang, R.; Dai, B.; Cheng, H.; Wang, Z. Effect of Bacillus velezensis to substitute in-feed antibiotics on the production, blood biochemistry and egg quality indices of laying hens. BMC Vet. Res. 2020, 16, 400. [Google Scholar] [CrossRef]
- Cheng, H.; Concepcion, G.T.; Feng, X.; Zhang, H.; Li, H. Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat. Methods 2021, 18, 170–175. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The kegg resource for deciphering the genome. Nucleic Acids Res. 2004, 32, 277–280. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The gene ontology consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.; Forslund, K.; Szklarczyk, D.; Trachana, K.; Roth, A.; Huerta-Cepas, J.; Gabaldón, T.; Rattei, T.; Creevey, C.; Kuhn, M.; et al. Eggnog v4.0: Nested orthology inference across 3686 organisms. Nucleic Acids Res. 2014, 42, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. Resfinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. Vfdb 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The carbohydrate-active enzymes database (cazy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, 233–238. [Google Scholar] [CrossRef]
- Saier, M.H., Jr.; Tran, C.V.; Barabote, R.D. Tcdb: The transporter classification database for membrane transport protein analyses and information. Nucleic Acids Res. 2006, 34, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Wagner, U.; Burkhardt, E.; Failing, K. Evaluation of canine lymphocyte proliferation: Comparison of three different colorimetric methods with the 3h-thymidine incorporation assay. Vet. Immunol. Immunopathol. 1999, 70, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, J.P.; Hoehler, D.; Wilkie, D.C.; Van Kessel, A.G.; Drew, M.D. Dietary glycine concentration affects intestinal Clostridium perfringens and lactobacilli populations in broiler chickens. Poult. Sci. 2005, 84, 1875–1885. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using qiime 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The unite database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nat. Rev. Microbiol. 2019, 47, 259–264. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The silva ribosomal rna gene database project: Improved data processing and web-based tools. Nat. Rev. Microbiol. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Gokul, J.K.; Hodson, A.J.; Saetnan, E.R.; Irvine-Fynn, T.D.L.; Westall, P.J.; Detheridge, A.P.; Takeuchi, N.; Bussell, J.; Mur, L.A.J.; Edwards, A. Taxon interactions control the distributions of cryoconite bacteria colonizing a high arctic ice cap. Mol. Ecol. 2016, 25, 3752–3767. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Guo, Y. Dietary supplementation of essential oils and lysozyme reduces mortality and improves intestinal integrity of broiler chickens with necrotic enteritis. Anim. Sci. J. 2021, 92, e13499. [Google Scholar] [CrossRef]
- Nøhr-Meldgaard, K.; Struve, C.; Ingmer, H.; Agersø, Y. Intrinsic tet(l) sub-class in Bacillus velezensis and Bacillus amyloliquefaciens is associated with a reduced susceptibility toward tetracycline. Front. Microbiol. 2022, 13, 966016. [Google Scholar] [CrossRef]
- Latorre, J.D.; Hernandez-Velasco, X.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Menconi, A.; Bielke, L.R.; Hargis, B.M.; Tellez, G. Evaluation and selection of bacillus species based on enzyme production, antimicrobial activity, and biofilm synthesis as direct-fed microbial candidates for poultry. Front. Vet. Sci. 2016, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian. Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Giongo, J.L.; Lucas, F.S.; Casarin, F.; Heeb, P.; Brandelli, A. Keratinolytic proteases of bacillus species isolated from the amazon basin showing remarkable de-hairing activity. World J. Microbiol. Biotechnol. 2007, 23, 375–382. [Google Scholar] [CrossRef]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Li, F.; Wang, Z. Characteristics and application of a novel species of bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.A.; Taha, A.E.; Soliman, S.M.; Ahmed, A.E.; El-Kott, A.F.; et al. Alternatives to antibiotics for organic poultry production: Types, modes of action and impacts on bird’s health and production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.-P.; Nelson, L.A.; Huang, F.S.; Warner, B.W. Intestinal adaptation: Structure, function, and regulation. Semin. Pediat. Surg. 2001, 10, 56–64. [Google Scholar] [CrossRef]
- Lallès, J.-P. Recent advances in intestinal alkaline phosphatase, inflammation, and nutrition. Nutr. Res. Rev. 2019, 77, 710–724. [Google Scholar] [CrossRef]
- Singh, K.M.; Shah, T.M.; Reddy, B.; Deshpande, S.; Rank, D.N.; Joshi, C.G. Taxonomic and gene-centric metagenomics of the fecal microbiome of low and high feed conversion ratio (fcr) broilers. J. Appl. Genet. 2014, 55, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Grant, A.; Gay, C.G.; Lillehoj, H.S. Bacillus spp. as direct-fed microbial antibiotic alternatives to enhance growth, immunity, and gut health in poultry. Avian. Pathol. 2018, 47, 339–351. [Google Scholar] [CrossRef]
- Li, A.; Wang, Y.; Pei, L.; Mehmood, K.; Li, K.; Qamar, H.; Iqbal, M.; Waqas, M.; Liu, J.; Li, J. Influence of dietary supplementation with bacillus velezensis on intestinal microbial diversity of mice. Microb. Pathog. 2019, 136, 103671. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia—A new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Guo, W.; Cui, S.; Zhang, Q.; Zhao, J.; Tang, X.; Zhang, H. Blautia producta displays potential probiotic properties against dextran sulfate sodium-induced colitis in mice. Food Sci. Hum. Wellness 2024, 13, 709–720. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, Y.; Mao, Y.; Gong, L.; Li, X.; Xu, S.; Wang, F.; Guo, Q.; Zhang, H.; Li, W. Dietary supplementation with Lactobacillus plantarum ameliorates compromise of growth performance by modulating short-chain fatty acids and intestinal dysbiosis in broilers under clostridium perfringens challenge. Front. Nutr. 2021, 8, 706148. [Google Scholar] [CrossRef]
- Zhu, X.; Tao, L.; Liu, H.; Yang, G. Effects of fermented feed on growth performance, immune organ indices, serum biochemical parameters, cecal odorous compound production, and the microbiota community in broilers. Poult. Sci. 2023, 102, 102629. [Google Scholar] [CrossRef]
- Kelsall, B.L. Innate and adaptive mechanisms to control of pathological intestinal inflammation. J. Pathol. 2008, 214, 242–259. [Google Scholar] [CrossRef]
- McAuley, J.L.; Linden, S.K.; Png, C.W.; King, R.M.; Pennington, H.L.; Gendler, S.J.; Florin, T.H.; Hill, G.R.; Korolik, V.; McGuckin, M.A. Muc1 cell surface mucin is a critical element of the mucosal barrier to infection. J. Clin. Investig. 2007, 117, 2313–2324. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Lee, S.H.; Gadde, U.D.; Oh, S.T.; Lee, S.J.; Lillehoj, H.S. Allium hookeri supplementation improves intestinal immune response against necrotic enteritis in young broiler chickens. Poult. Sci. 2018, 97, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Lindén, S.K.; Florin, T.H.; McGuckin, M.A. Mucin dynamics in intestinal bacterial infection. PLoS ONE 2008, 3, e3952. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Wang, B.; Zhou, Y.; Tang, L.; Zeng, Z.; Zhang, H.; Li, W. Protective effects of Lactobacillus plantarum 16 and paenibacillus polymyxa 10 against Clostridium perfringens infection in broilers. Front. Immunol. 2021, 11, 628374. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, Z.; Ramzan, R.; Abdelazez, A.; Amjad, A.; Afzaal, M.; Zhang, S.; Pan, S. Assessment of the antimicrobial potentiality and functionality of Lactobacillus plantarum strains isolated from the conventional inner mongolian fermented cheese against foodborne pathogens. Pathogens 2019, 8, 71. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Sensitivity 1 | Antibiotic | Sensitivity | Antibiotic | Sensitivity | |||
|---|---|---|---|---|---|---|---|---|
| F1 2 | F5 3 | F1 | F5 | F1 | F5 | |||
| Piperacillin/tazobactam | S | S | Meropenem | S | S | Rifampin | S | S |
| Ampicillin | S | S | Vancomycin | S | S | Tetracycline | R | R |
| Ciprofloxacin | S | S | Neomycin | S | S | Trimethoprim/ Sulfamethoxazole | S | S |
| Penicillin | S | S | Ceftriaxone | S | S | Amikacin | S | S |
| Erythromycin | S | S | Cephalosporin | S | S | Ceftazidime | S | S |
| Chloromycetin | S | S | Cefazolin | S | S | Cephalothin | S | S |
| Azithromycin | S | S | Cefotaxime | S | S | Cefoperazone | S | S |
| Clindamycin | S | S | Spectinomycin | S | S | Gentamicin | S | S |
| Doxycycline | S | S | Cefuroxime | S | S | Oxacillin | S | S |
| Clarithromycin | S | S | Minocycline | S | S | Nitrofurantoin | S | S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu La, A.L.T.; Wen, Q.; Xiao, Y.; Hu, D.; Liu, D.; Guo, Y.; Hu, Y. A New Bacillus velezensis Strain CML532 Improves Chicken Growth Performance and Reduces Intestinal Clostridium perfringens Colonization. Microorganisms 2024, 12, 771. https://doi.org/10.3390/microorganisms12040771
Zhu La ALT, Wen Q, Xiao Y, Hu D, Liu D, Guo Y, Hu Y. A New Bacillus velezensis Strain CML532 Improves Chicken Growth Performance and Reduces Intestinal Clostridium perfringens Colonization. Microorganisms. 2024; 12(4):771. https://doi.org/10.3390/microorganisms12040771
Chicago/Turabian StyleZhu La, A La Teng, Qiu Wen, Yuxuan Xiao, Die Hu, Dan Liu, Yuming Guo, and Yongfei Hu. 2024. "A New Bacillus velezensis Strain CML532 Improves Chicken Growth Performance and Reduces Intestinal Clostridium perfringens Colonization" Microorganisms 12, no. 4: 771. https://doi.org/10.3390/microorganisms12040771
APA StyleZhu La, A. L. T., Wen, Q., Xiao, Y., Hu, D., Liu, D., Guo, Y., & Hu, Y. (2024). A New Bacillus velezensis Strain CML532 Improves Chicken Growth Performance and Reduces Intestinal Clostridium perfringens Colonization. Microorganisms, 12(4), 771. https://doi.org/10.3390/microorganisms12040771