

Changing Patterns and Driving Factors of Plankton Coupling Relationships in Lakes around the Yangtze River, China
Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Data Sources
2.3. Analysis Methods
3. Results
3.1. Trophic Level of Lakes in the Lake Basins
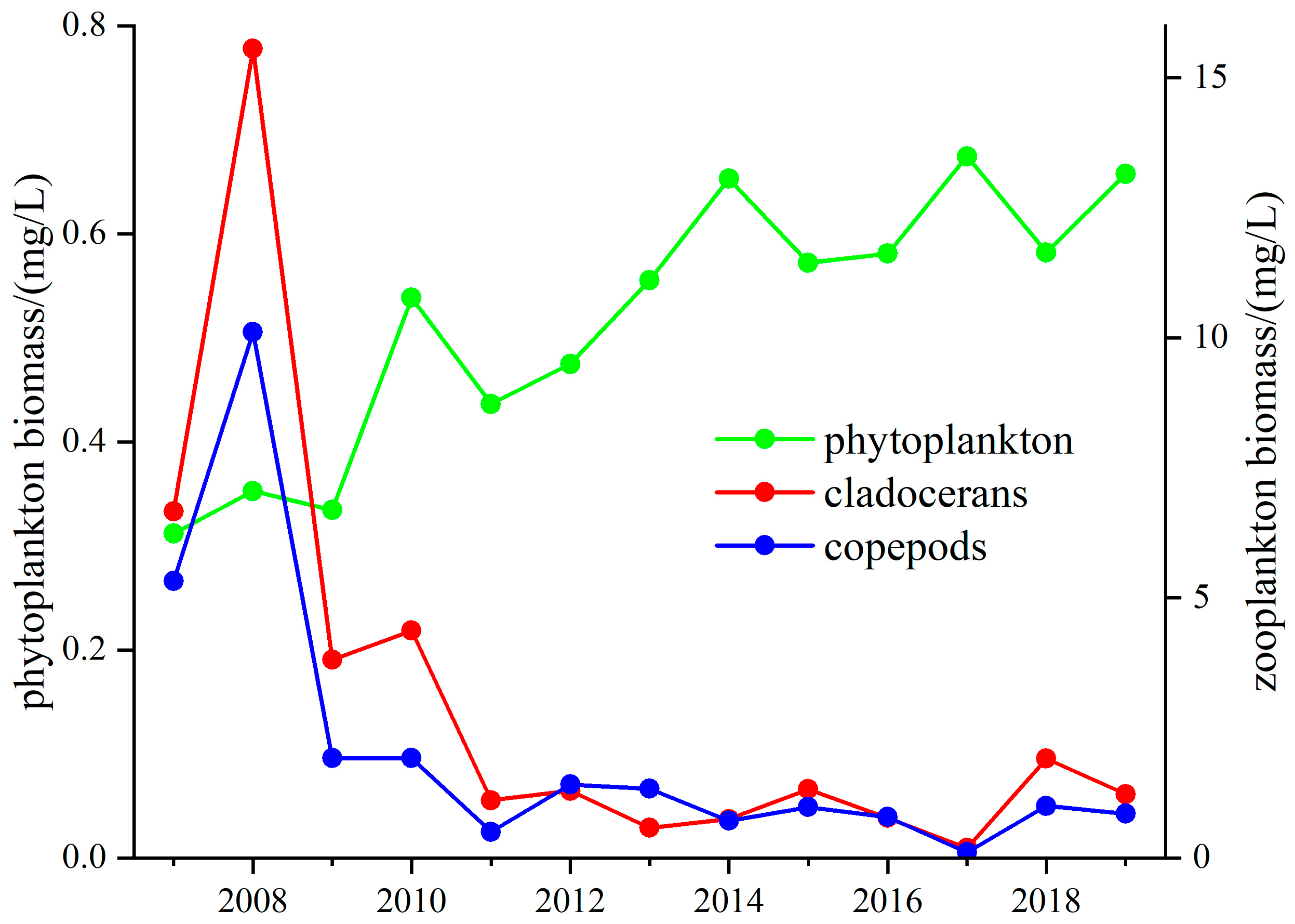
3.2. Changes in Plankton Coupling Relationships in Lakes around the Yangtze River
3.2.1. Changes in the Phytoplankton Community Structure
3.2.2. Changes in the Zooplankton Community Structure
3.2.3. Changes in the Z/P Ratio
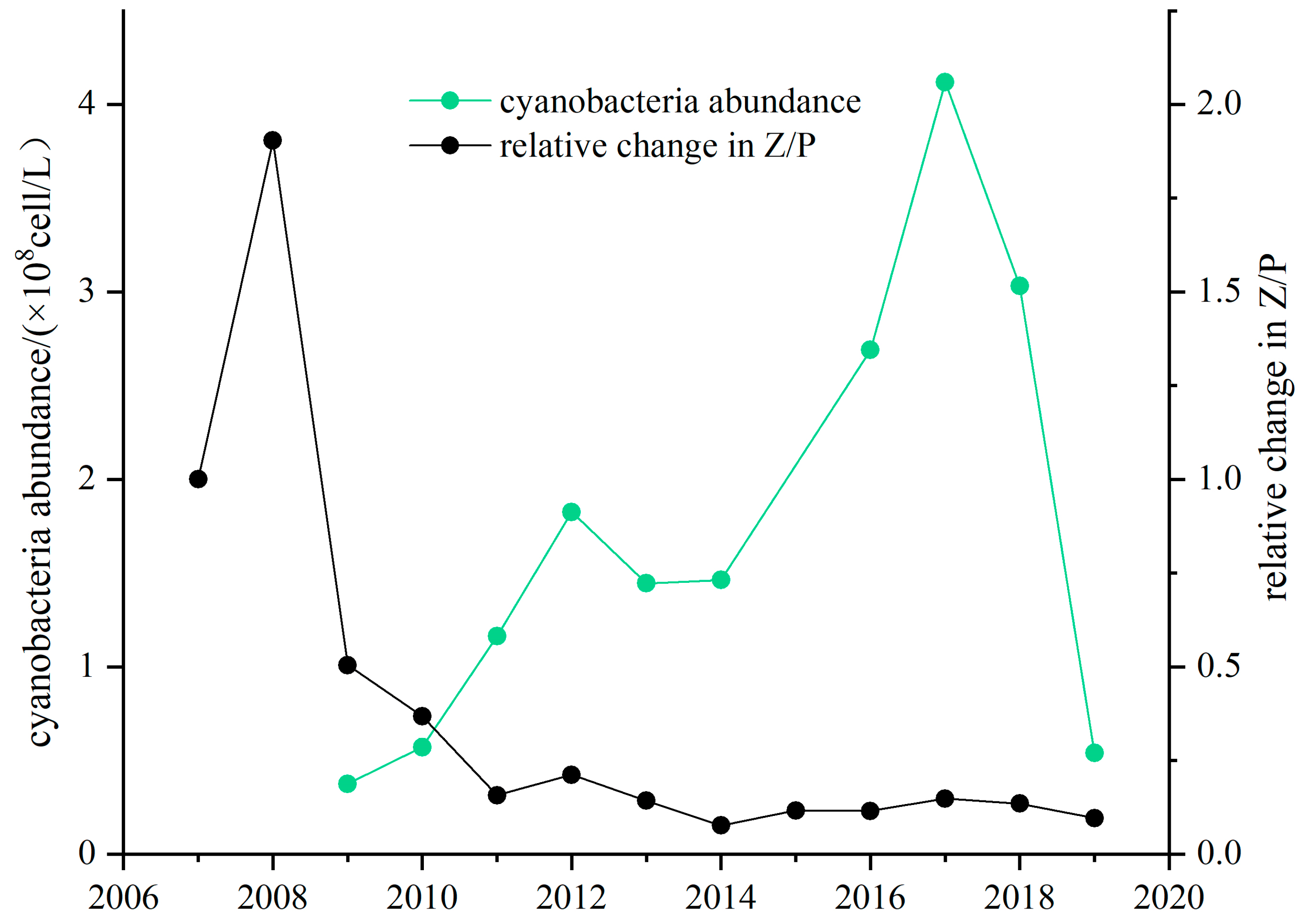
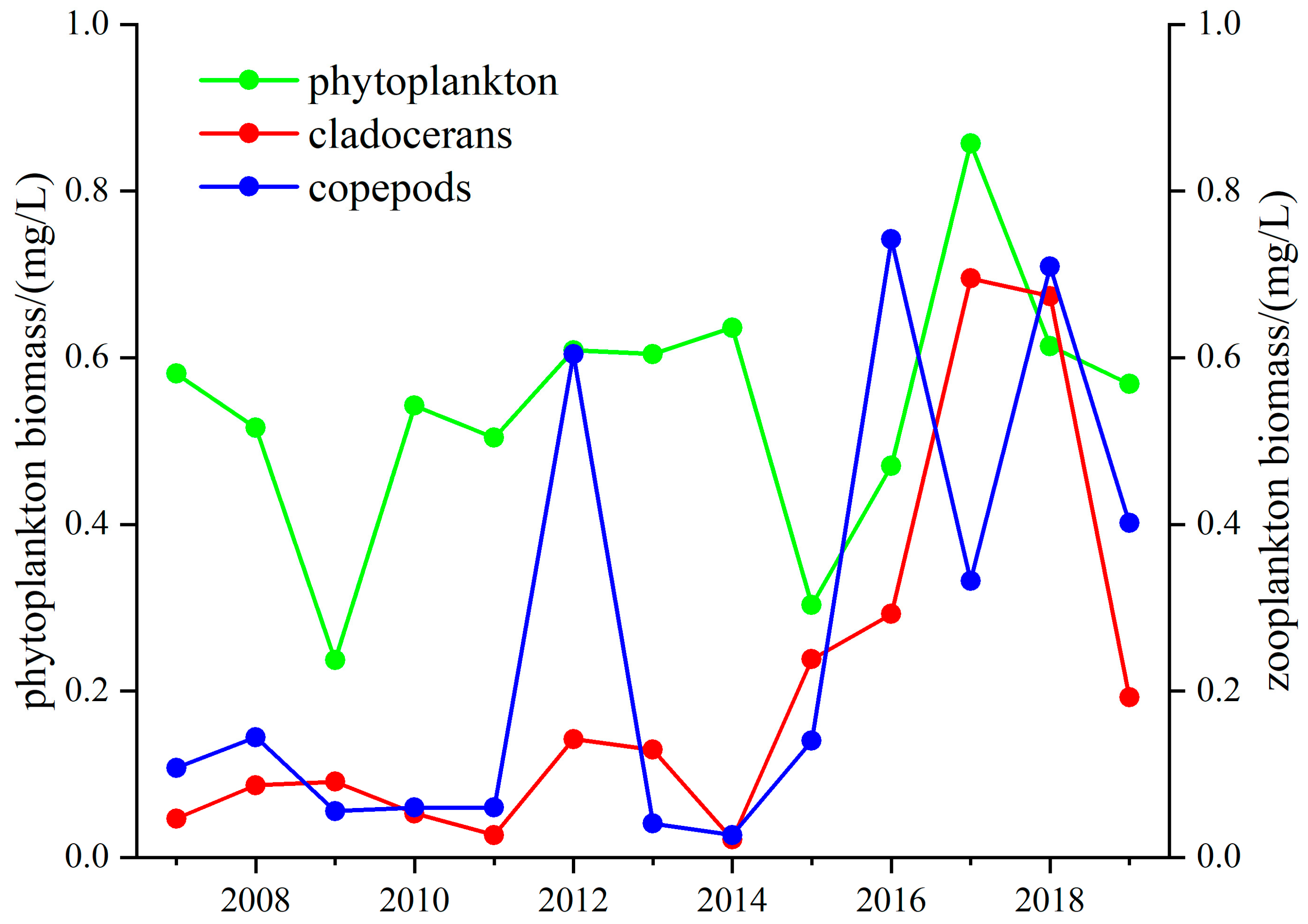
3.3. Long-Term Change in the Z/P Ratio in Lake Taihu and Lake Donghu
3.3.1. Lake Taihu
3.3.2. Lake Donghu
4. Discussion
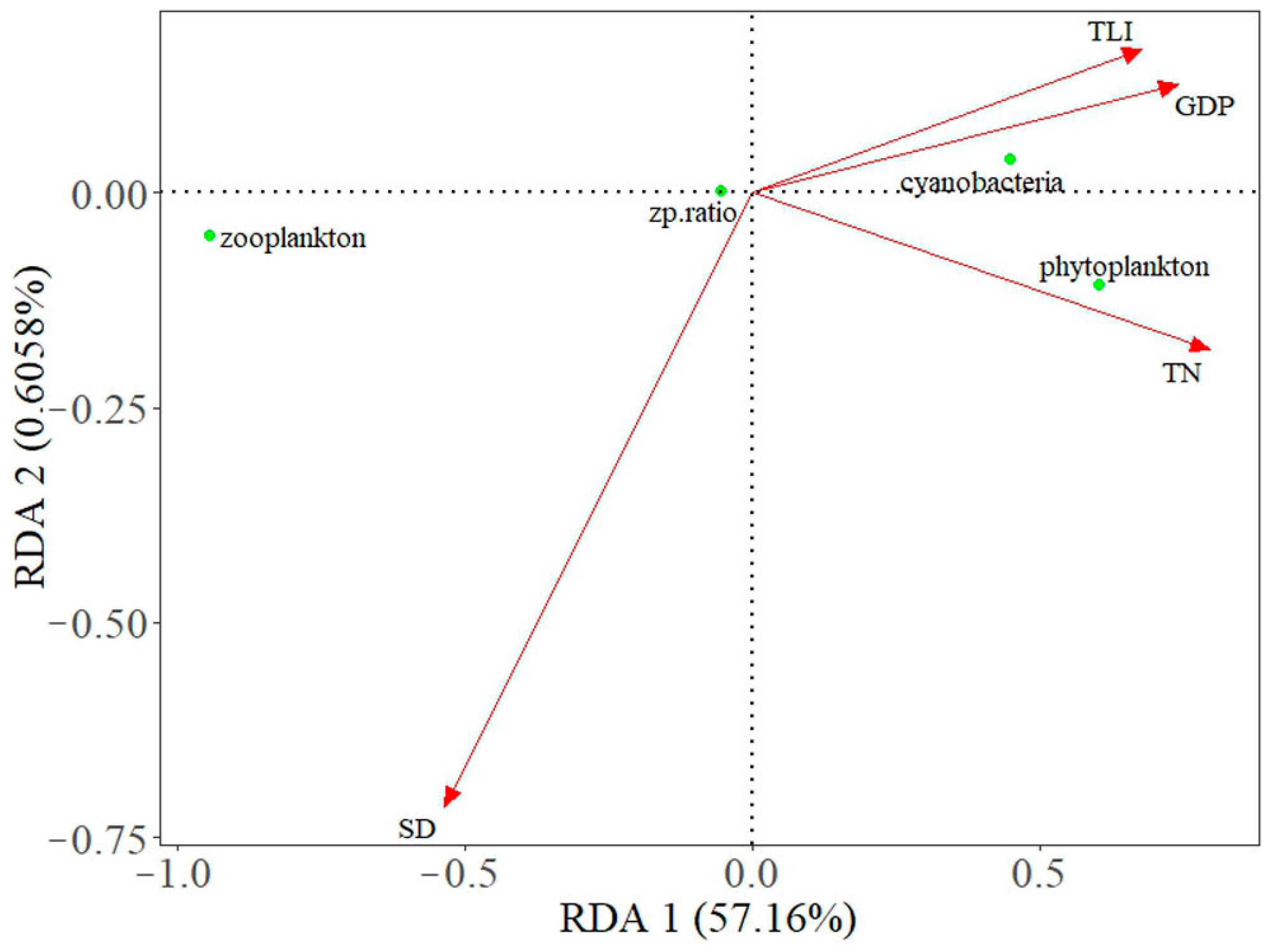
4.1. Drivers of Plankton Coupling Relationships in Lakes around the Yangtze River
4.2. Differences in Plankton Coupling Relationships between Lake Taihu and Lake Donghu
4.3. Suggestions for Lake Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiang, L.; Li, Y.W.; Liu, B.L.; Zhao, H.M.; Li, H.; Cai, Q.Y.; Mo, C.H.; Wong, M.H.; Li, Q.X. High ecological and human health risks from microcystins in vegetable fields in southern China. Environ. Int. 2019, 133, 105142. [Google Scholar] [CrossRef]
- Zhu, G.W.; Qin, B.Q.; Zhang, Y.L.; Xu, H.; Zhu, M.Y.; Yang, H.W.; Li, K.Y.; Min, S.; Shen, R.J.; Zhong, C.N. Variation and driving factors of nutrients and chlorophyll-a concentrations in northern region of Lake Taihu, China, 2005–2017. J. Lake Sci. 2018, 30, 279–295. [Google Scholar]
- Filstrup, C.T.; Hillebrand, H.; Heathcote, A.J.; Harpole, W.S.; Downing, J.A. Cyanobacteria dominance influences resource use efficiency and community turnover in phytoplankton and zooplankton communities. Ecol. Lett. 2014, 17, 464–474. [Google Scholar] [CrossRef]
- Zhong, F.; Gao, Y.N.; Yu, T.; Zhang, Y.Y.; Xu, D.; Xiao, E.R.; He, F.; Zhou, Q.H.; Wu, Z.B. The management of undesirable cyanobacteria blooms in channel catfish ponds using a constructed wetland: Contribution to the control of off-flavor occurrences. Water Res. 2011, 45, 6479–6488. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, J.; Qin, B.Q.; Quan, Q.M.; Li, Y.X. Long-term Variation Characteristics of Zooplankton Community Structure in Meiliang Bay, Lake Taihu. Environ. Sci. 2020, 41, 1246–1255. [Google Scholar]
- Li, Y.; Wang, R.; Su, H.; Wang, J.; Xie, P.; Chen, F. Eutrophication and predation mediate zooplankton diversity and network structure. Limnol. Oceanogr. 2021, 67, 133–145. [Google Scholar] [CrossRef]
- Jiang, X.D.; Xie, J.H.; Xu, Y.; Zhong, W.F.; Zhu, X.; Zhu, C.D. Increasing dominance of small zooplankton with toxic cyanobacteria. Freshw. Biol. 2017, 62, 429–443. [Google Scholar] [CrossRef]
- He, H.; Ning, X.Y.; Chen, K.Q.; Li, Q.S.; Li, K.Y.; Liu, Z.W.; Jeppesen, E. Intraguild predation dampens trophic cascades in shallow aquatic mesocosms in the subtropics: Implications for lake restoration by biomanipulation. Freshw. Biol. 2021, 66, 1571–1580. [Google Scholar] [CrossRef]
- Peng, Y.H.; Li, Y.J.; Zhang, T.H. Global bifurcation in a toxin producing phytoplankton-zooplankton system with prey-taxis. Nonlinear Anal.-Real World Appl. 2021, 61, 103326. [Google Scholar] [CrossRef]
- Amorim, C.A.; Moura, A.D. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef]
- Heathcote, A.J.; Filstrup, C.T.; Kendall, D.; Downing, J.A. Biomass pyramids in lake plankton: Influence of Cyanobacteria size and abundance. Inland Waters 2016, 6, 250–257. [Google Scholar] [CrossRef]
- Muylaert, K.; Declerck, S.; Van Wichelen, J.; De Meester, L.; Vyverman, W. An evaluation of the role of daphnids in controlling phytoplankton biomass in clear water versus turbid shallow lakes. Limnologica 2006, 36, 69–78. [Google Scholar] [CrossRef]
- Symons, C.C.; Arnott, S.E.; Sweetman, J.N. Grazing rates of crustacean zooplankton communities on intact phytoplankton communities in Canadian Subarctic lakes and ponds. Hydrobiologia 2012, 694, 131–141. [Google Scholar] [CrossRef]
- Pei, Y.Z.; Lv, Y.F.; Li, C.G. Evolutionary consequences of harvesting for a two-zooplankton one-phytoplankton system. Appl. Math. Model. 2012, 36, 1752–1765. [Google Scholar] [CrossRef]
- Peng, G.G.; Zhou, X.J.; Xie, B.; Huang, C.; Uddin, M.M.; Chen, X.X.; Huang, L.F. Ecosystem stability and water quality improvement in a eutrophic shallow lake via long-term integrated biomanipulation in Southeast China. Ecol. Eng. 2021, 159, 106119. [Google Scholar] [CrossRef]
- Li, Y.; Geng, M.D.; Yu, J.L.; Du, Y.X.; Xu, M.; Zhang, W.Z.; Wang, J.J.; Su, H.J.; Wang, R.; Chen, F.Z. Eutrophication decrease compositional dissimilarity in freshwater plankton communities. Sci. Total Environ. 2022, 821, 153434. [Google Scholar] [CrossRef]
- Ding, N.; Zhou, Y.F.; Song, J.T.; Jiang, S.L.; Ge, Y.; Jing, B.; You, Y. Community characteristics of phytoplankton in Yangcheng Lake and relationships with water environmental factors. J. Shanghai Ocean. Univ. 2015, 24, 702–711. [Google Scholar]
- Liu, Q. The Relationship between Community Structure of Micro-Zooplankton and Eutrophication in Yangcheng Lake. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2011. [Google Scholar]
- Guo, C.X.; Wang, D.M.; Hu, X.D.; Wu, P.P. Phytoplankton Community Structure and Its Relationship to Environmental Factors in Shijiu Lake (Jiangsu section). Acta Hydrobiol. Sin. 2016, 37, 23–29. [Google Scholar]
- Su, Y.Y.; Zhao, X.P.; Guo, L.C. Spatial and temporal dynamic changes and water quality biological evaluation of zooplankton community structure in Jiangsu section of Shijiu Lake. Jiangsu Water Resour. 2020, 11, 43–47. [Google Scholar]
- Chen, Y.; Peng, K.; Zhang, Q.J.; Cai, Y.J.; Zhang, Y.Z.; Gong, Z.J.; Xiang, X.L. Spatio-temporal Distribution Characteristics and Driving Factors of Zooplankton in Hongze Lake. Environ. Sci. 2021, 42, 3753–3762. [Google Scholar]
- Wu, T.H.; Liu, J.S.; Deng, J.M.; Dai, X.L.; Tang, R.G.; Peng, K.; Zou, W.; Cai, Y.J.; Gong, Z.J. Community structure of phytoplankton and bioassessment of water quality in a large water-carrying lake, Lake Hongze. J. Lake Sci. 2019, 31, 440–448. [Google Scholar]
- Ding, H. The Current Situation, Problems and Countermeasures of Luoma Wetland. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2010. [Google Scholar]
- Peng, K.; Li, T.M.; Liu, L.; Zou, W.; Wu, T.H.; Gong, Z.J. Community structure of phytoplankton and bio-assessment of water quality in Lake Luoma, northern Jiangsu Province. J. Lake Sci. 2018, 30, 183–191. [Google Scholar]
- Chen, Q.Y.; Wu, L. Relationship between Zooplankton Functional Groups and Phytoplankton in Summer in Chaohu Lake. J. Anhui Univ. Sci. Technol. (Nat. Sci.) 2021, 41, 42–50. [Google Scholar]
- Li, H.G.; Yang, C.M.; Wang, Y.L. Water Quality and Community Structure Characteristics of Plankton in Chaohu Lake. J. Anhui Agric. Sci. 2017, 45, 13–16. [Google Scholar]
- Wu, L.; Zhou, M.H.; Shen, Z.J.; Geng, M.; Feng, W.S. Characteristics of Zooplankton Community and Water Quality Assessment in Chaohu Lake and Its Tributaries. Chin. J. Zool. 2017, 52, 792–811. [Google Scholar]
- Liu, X.H.; Zhao, X.X.; Gao, P.; Han, F.Y.; Zhou, F.; Zhou, Z.Z.; Xu, H.Q. Annual dynamics of phytoplankton abundance and community structure (2010) in Lake Caizi, Anhui Province. J. Lake Sci. 2012, 24, 771–779. [Google Scholar]
- Liu, Z.Z. Effects of Aquatic Vegetation Restored after the Removal of Purse Seine Net on the Community Structure of Metazoan Zooplankton in Caizi Lake. Master’s Thesis, Anhui University, Heifei, China, 2022. [Google Scholar]
- Chen, J.Q.; Zhao, K.; Cao, Y.; Wu, B.; Pang, W.T.; You, Q.M.; Wang, Q.X. Zooplankton community structure and its relationship with environmental factors in Poyang Lake. Acta Ecol. Sin. 2020, 40, 6644–6658. [Google Scholar]
- Qian, K.M.; Liu, B.G.; Chen, Y.W. Long term dynamics of phytoplankton functional groups in Lake Poyang during 2009–2016. J. Lake Sci. 2019, 31, 1035–1044. [Google Scholar]
- Yang, X.; Ma, J.S.; Zhang, H.; Zhou, Q. Community structure and the water quality during different hydrological periods in Poyang Lake. Acta Hydrobiol. Sin. 2021, 45, 1093–1103. [Google Scholar]
- Gong, L.J.; Zhang, S.P.; Xiong, B.X.; Cao, J.; Zhang, J.; Li, J.Z.; Liu, D.Z. Identification and Comparative Analysis of Plankton in Aquiculture Lakes in Summer in Wuhan City. J. Hydroecol. 2009, 30, 9–14. [Google Scholar]
- Gong, L.J.; Zhang, S.P.; Xiong, B.X.; Liu, D.Z.; Li, J.Z.; Cao, J. Study on plankton of different categories of lakes in summer by means of principal component analysis, factor analysis and cluster analysis. J. Hydroecol. 2010, 34, 43–50. [Google Scholar]
- Xu, L.J.; Lu, X.X.; Wang, S.Z. The biodiversity of crustacean zooplankton in Lake Liangzi. Acta Ecol. Sin. 2009, 29, 6419–6428. [Google Scholar]
- Gong, S.S.; Li, X.M.; Wu, J.W.; Chai, Y.; Yang, D.G.; Tan, F.X.; Zhu, T.B. Characteristics and main influencing factors of plankton functional groups in summer and autumn of Changhu Lake. Acta Hydrobiol. Sin. 2022, 46, 707–717. [Google Scholar]
- Jiang, J.J.; Hu, W.; Ye, C.; Song, D.; Wang, Z.Y.; Li, C.H.; Li, J.; Tang, C. Succession and driving factors of Lake Dianchi aquatic ecosystem in the past 60 years. J. Environ. Eng. Technol. 2023, 13, 541–551. [Google Scholar]
- Liu, C.Y.; Yu, Y.H.; Wang, R.; Song, D.; Yang, S.L.; Wang, Z.L. The Plankton Diversity of Dianchi Lake. J. West China For. Sci. 2016, 45, 74–80. [Google Scholar]
- Sun, C.Q. The Study on the Community Structure and the Variation of Population Quantity in Dianchi Lake. Master’s Thesis, Yunnan University, Kunming, China, 2010. [Google Scholar]
- Wang, M.C.; Liu, X.Q.; Zhang, J.H. Evaluate method and classification standard on lake eutrophication. Environ. Monit. China 2002, 18, 47–49. [Google Scholar]
- Wu, T.; Zhu, G.; Zhu, M.; Xu, H.; Zhang, Y.; Qin, B. Use of conductivity to indicate long-term changes in pollution processes in Lake Taihu, a large shallow lake. Environ. Sci. Pollut. Res. 2020, 27, 21376–21385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, P.; Tao, M.; Guo, L.; Chen, J.; Li, L.; Zhang, X.; Zhang, L. The impact of fish predation and cyanobacteria on zooplankton size structure in 96 subtropical lakes. PLoS ONE 2013, 8, e76378. [Google Scholar] [CrossRef]
- Rohwer, R.R.; Hale, R.J.; Vander Zanden, M.J.; Miller, T.R.; McMahon, K.D. Species invasions shift microbial phenology in a two-decade freshwater time series. Proc. Natl. Acad. Sci. USA 2023, 120, e2211796120. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qian, K.M.; Chen, Y.W. Effects of water level fluctuations on phytoplankton in a Changjiang River floodplain lake (Poyang Lake): Implications for dam operations. J. Great Lakes Res. 2015, 41, 770–779. [Google Scholar] [CrossRef]
- Baranyi, C.; Hein, T.; Holarek, C.; Keckeis, S.; Schiemer, F. Zooplankton biomass and community structure in a Danube River floodplain system: Effects of hydrology. Freshw. Biol. 2002, 47, 473–482. [Google Scholar] [CrossRef]
- Wu, Z.S.; Ma, T.T.; Xiong, L.L.; Deng, Y.Q.; Li, K.Y. How does phytoplankton respond to hygrophyte decomposition during the inundation period? Hydrobiologia 2023, 850, 51–63. [Google Scholar] [CrossRef]
- Cao, J.; Hou, Z.; Li, Z.; Zheng, B.; Chu, Z. Spatiotemporal dynamics of phytoplankton biomass and community succession for driving factors in a meso-eutrophic lake. J. Environ. Manag. 2023, 345, 118693. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.G.; He, F.; Qi, T.C.; Sun, Z.; Shen, M.; Cao, Z.G.; Meng, D.; Duan, H.T.; Luo, J.H. Thirty-Four-Year Record (1987–2021) of the Spatiotemporal Dynamics of Algal Blooms in Lake Dianchi from Multi-Source Remote Sensing Insights. Remote Sens. 2022, 14, 4000. [Google Scholar] [CrossRef]
- Shi, H.B.; Chen, G.J.; Lu, H.B.; Wang, J.Y.; Huang, L.P.; Wang, L.; Zhao, S.Y.; Liu, X.H. Regional pattern of Bosmina responses to fish introduction and eutrophication in four large lakes from Southwest China. J. Plankton Res. 2016, 38, 443–455. [Google Scholar] [CrossRef]
- Zou, W.; Zhu, G.; Xu, H.; Zhu, M.; Guo, C.; Qin, B.; Zhang, Y. Atmospheric Stilling Promotes Summer Algal Growth in Eutrophic Shallow Lakes. Biology 2021, 10, 1222. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Sondergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B.; et al. Lake responses to reduced nutrient loading: An analysis of contemporary long-term data from 35 case studies. Freshw. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Zhang, K.; Jiang, F.Y.; Chen, H.; Dibar, D.T.; Wu, Q.L.; Zhou, Z.Z. Temporal and spatial variations in zooplankton communities in relation to environmental factors in four floodplain lakes located in the middle reach of the Yangtze River, China. Environ. Pollut. 2019, 251, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.J.; He, W.C.; Guo, L.G.; Gong, L.; Yang, Y.L.; Yang, J.J.; Ni, L.Y.; Chen, Y.S.; Jeppesen, E. Can top-down effects of planktivorous fish removal be used to mitigate cyanobacterial blooms in large subtropical highland lakes? Water Res. 2022, 218, 118483. [Google Scholar] [CrossRef] [PubMed]
- Belfiore, A.P.; Buley, R.P.; Fernandez-Figueroa, E.G.; Gladfelter, M.F.; Wilson, A.E. Zooplankton as an alternative method for controlling phytoplankton in catfish pond aquaculture. Aquac. Rep. 2021, 21, 100897. [Google Scholar] [CrossRef]
- Chen, J.; Liu, J.; Han, S.; Su, H.; Xia, W.; Wang, H.; Liu, Y.; Zhang, L.; Ke, Z.; Zhang, X.; et al. Nontraditional biomanipulation: A powerful ecotechnology to combat cyanobacterial blooms in eutrophic freshwaters. Innov. Life 2023, 1, 100038. [Google Scholar] [CrossRef]
- Liu, J.K.; Xie, P. Direct control of microsystis bloom through the use of Planktivorous Carp-closure experiments and lake fishery practice. Ecol. Sci. 2003, 22, 193–198. [Google Scholar]
- Zhu, G.W.; Xu, H.; Zhu, M.Y.; Zou, W.; Guo, C.X.; Ji, P.F.; Da, W.Y.; Zhou, Y.Q.; Zhang, Y.L.; Qin, B.Q. Changing characteristics and driving factors of trophic state of lakes in the middle and lower reaches of Yangtze River in the past 30 years. J. Lake Sci. 2019, 31, 1510–1524. [Google Scholar]
- Lu, B.; Gu, H.B.; Jiang, H. Brief Analysis on Environmental Assessment for Dam Removal of Small Hydropower Project. Water Power 2019, 45, 12–16. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Lake Area (km2) | Lake Basin GDP in 2010s (billion CNY) | Data Sources |
|---|---|---|---|
| Lake Yangcheng | 120 | 634 | [17,18] |
| Lake Taihu | 2340 | 224 | CNERN |
| Lake Shijiu | 208 | 2080 | [19,20] |
| Lake Hongze | 2069 | 430 | [21,22] |
| Lake Luoma | 296 | 785 | [23,24] |
| Lake Chaohu | 780 | 441 | [25,26,27] |
| Lake Caizi | 226 | 94 | [28,29] |
| Lake Poyang | 3150 | 645 | [30,31,32] |
| Lake Donghu | 32 | 546 | CNERN |
| Lake Liangzi | 370 | 179 | [33,34,35] |
| Lake Changhu | 131 | 308 | [36] |
| Lake Dongting | 2579 | 365 | proprietary dataset |
| Lake Dianchi | 330 | 217 | [37,38,39] |
| Lake Erhai | 250 | 91 | proprietary dataset |
| chla | TP | TN | SD | CODMn | |
|---|---|---|---|---|---|
| 1 | 0.7056 | 0.6724 | 0.6889 | 0.6889 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Guo, X.; Liu, H.; Chu, Z.; Wu, T. Changing Patterns and Driving Factors of Plankton Coupling Relationships in Lakes around the Yangtze River, China. Microorganisms 2024, 12, 1698. https://doi.org/10.3390/microorganisms12081698
Dong C, Guo X, Liu H, Chu Z, Wu T. Changing Patterns and Driving Factors of Plankton Coupling Relationships in Lakes around the Yangtze River, China. Microorganisms. 2024; 12(8):1698. https://doi.org/10.3390/microorganisms12081698
Chicago/Turabian StyleDong, Chenhao, Xinchao Guo, Haiyan Liu, Zhaosheng Chu, and Tianhao Wu. 2024. "Changing Patterns and Driving Factors of Plankton Coupling Relationships in Lakes around the Yangtze River, China" Microorganisms 12, no. 8: 1698. https://doi.org/10.3390/microorganisms12081698