

Antimicrobial Resistance of Staphylococcus spp. from Human Specimens Submitted to Diagnostic Laboratories in South Africa, 2012–2017



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Data Extraction
2.2. Isolation, Identification, and Storage of Staphylococcus Strains
2.3. Data Management
2.4. Data Analysis
2.5. Ethical Considerations
3. Results
3.1. Descriptive Statistics
3.1.1. Isolates Examined in This Study
3.1.2. Resistance Observed against Different Antimicrobial Agents
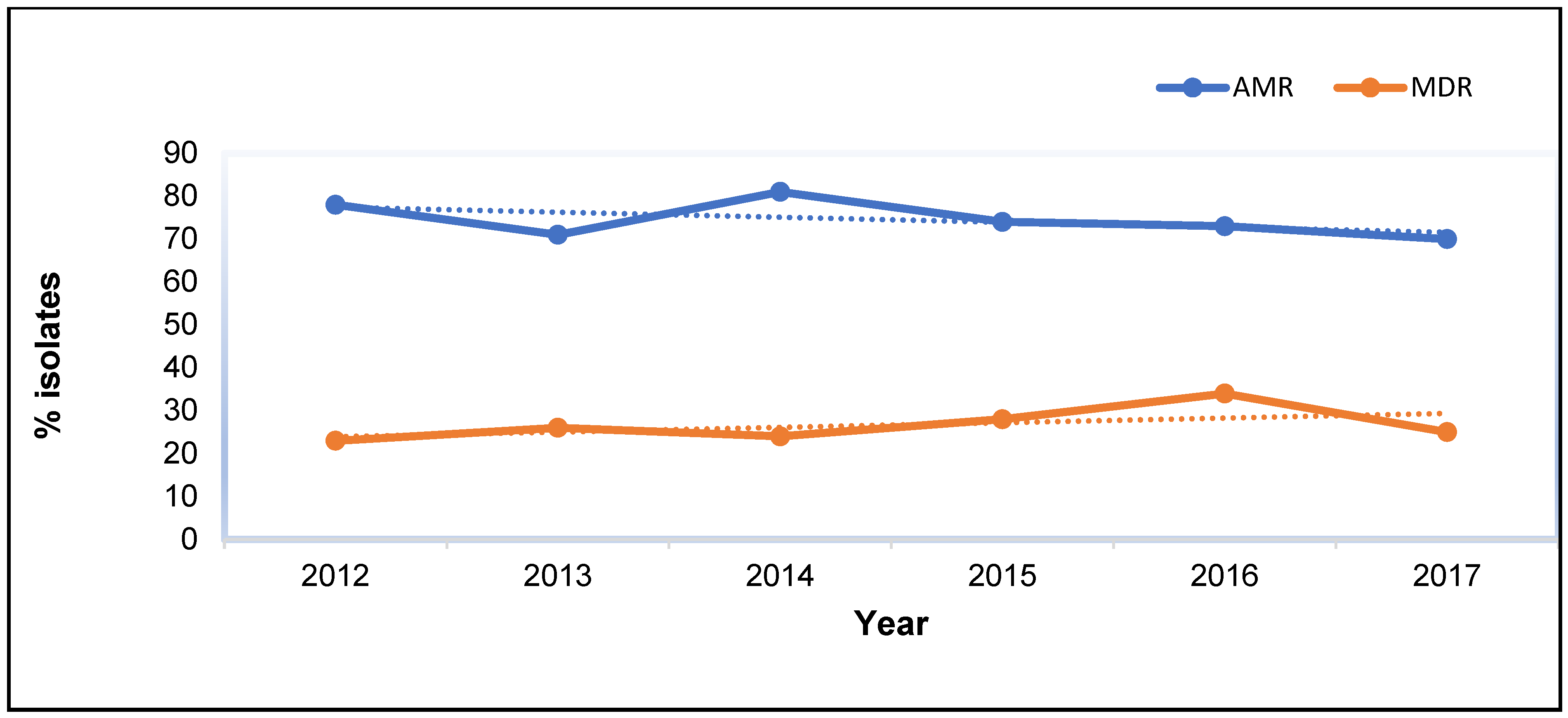
3.1.3. Temporal Trends
3.2. Inferential Statistics
3.2.1. Predictors for Antimicrobial Drug Resistance
3.2.2. Predictors for Multidrug Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musumeci, T.; Puglisi, G. Antimicrobial agents. Drug-Biomembr. Interact. Stud. Appl. Calorim. Tech. 2013, 1, 305–333. [Google Scholar] [CrossRef]
- Sakwinska, O.; Giddey, M.; Moreillon, M.; Morisset, D.; Waldvogel, A.; Moreillon, P. Staphylococcus aureus host range and human-bovine host shift. Appl. Environ. Microbiol. 2011, 77, 5908–5915. [Google Scholar] [CrossRef] [PubMed]
- Praxis, Covariance Structure Analysis of Health-Related Indicators for Elderly People Living at Home with a Focus on Subjective Sense of Health. 2022, Volume 2. Available online: https://www.jstage.jst.go.jp/ (accessed on 9 September 2023).
- CDC. Antibiotic Resistance Threats in The United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- Oliveira, D.C.; De Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.A.; Enright, M.C. Evolutionary Models of the Emergence of Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 3926–3934. [Google Scholar] [CrossRef]
- Huttner, A.; Harbarth, S.; Carlet, J.; Cosgrove, S.; Goossens, H.; Holmes, A.; Jarlier, V.; Voss, A.; Pittet, D. Antimicrobial resistance: A global view from the 2013 World Healthcare-Associated Infections Forum. Antimicrob. Resist. Infect. Control 2013, 2, 31. [Google Scholar] [CrossRef]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Hariyanto, H.; Yahya, C.Q.; Cucunawangsih, C.; Pertiwi, C.L.P. Antimicrobial Resistance and Mortality. Afr. J. Infect. Dis. 2022, 16, 13–20. [Google Scholar] [CrossRef]
- Herron-Olson, L.; Fitzgerald, J.R.; Musser, J.M.; Kapur, V. Molecular correlates of host specialization in Staphylococcus aureus. PLoS ONE 2007, 2. [Google Scholar] [CrossRef]
- Allen, H.K.; Donato, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Adams, D.A.; Thomas, K.R.; Jajosky, R.A.; Foster, L.; Baroi, G.; Sharp, P.; Onweh, D.H.; Schley, A.W.; Anderson, W.J. Summary of Notifiable Infectious Diseases and Conditions—United States, 2015. Morb. Mortal. Wkly. Rep. 2017, 64, 1–143. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, N.D.; Dunavan, C.P.; Diamond, J. Origins of major human infectious diseases. Nature 2007, 447, 279–283. [Google Scholar] [CrossRef]
- Bramble, M.; Morris, D.; Tolomeo, P.; Lautenbach, E. Potential role of pet animals in household transmission of Methicillin-resistant staphylococcus aureus: A narrative review. Vector-Borne Zoonotic Dis. 2011, 11, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Vanderhaeghen, W.; Dewulf, J. Antimicrobial use and resistance in animals and human beings. Lancet Planet. Health 2017, 1, e307–e308. [Google Scholar] [CrossRef]
- Graveland, H.; Wagenaar, J.A.; Heesterbeek, H.; Mevius, D.; van Duijkeren, E.; Heederik, D. Methicillin resistant staphylococcus aureus ST398 in veal calf farming: Human MRSA carriage related with animal antimicrobial usage and farm hygiene. PLoS ONE 2010, 5, 4–9. [Google Scholar] [CrossRef]
- Perreten, V.; Kadlec, K.; Schwarz, S.; Andersson, U.G.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A.; et al. Clonal spread of methicillin-resistant Staphylococcus pseudintermedius in Europe and North America: An international multicentre study. J. Antimicrob. Chemother. 2010, 65, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Tsubakishita, S.; Tanaka, Y.; Sakusabe, A.; Ohtsuka, M.; Hirotaki, S.; Kawakami, T.; Fukata, T.; Hiramatsu, K. Multiplex-PCR method for species identification of coagulase-positive staphylococci. J. Clin. Microbiol. 2010, 48, 765–769. [Google Scholar] [CrossRef]
- Statistics South Africa and StatsSA. Statistical Release P0302: Mid-Year Population Estimates. 2020; pp. 1–22. Available online: http://www.statssa.gov.za/publications/P0302/P03022017.pdf (accessed on 6 September 2023).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- StataCorp. Stata Statistical Software: Release 17; StataCorp LLC.: College Station, TX, USA, 2023. [Google Scholar]
- Khanna, T.; Friendship, R.; Dewey, C.; Weese, J.S. Methicillin resistant Staphylococcus aureus colonization in pigs and pig farmers. Vet. Microbiol. 2008, 128, 298–303. [Google Scholar] [CrossRef]
- Soe, P.E.; Han, W.W.; Sagili, K.D.; Satyanarayana, S.; Shrestha, P.; Htoon, T.T.; Tin, H.H. High prevalence of methicillin-resistant staphylococcus aureus among healthcare facilities and its related factors in myanmar (2018–2019). Trop. Med. Infect. Dis. 2021, 6, 70. [Google Scholar] [CrossRef]
- Gandra, S.; Alvarez-Uria, G.; Turner, P.; Joshi, J.; Limmathurotsakul, D.; van Doorn, H.R. Antimicrobial Resistance Surveillance in Low- and Middle-Income Countries: Progress and Challenges in Eight South. Clin. Microbiol. Rev. 2020, 33, e00048-19. [Google Scholar] [CrossRef] [PubMed]
- Waterlow, N.R.; Cooper, B.S.; Robotham, J.V.; Knight, G.M. Antimicrobial resistance prevalence in bloodstream infection in 29 European countries by age and sex: An observational study. PLoS Med. 2024, 21, 1–24. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Mitchell, J.; Cooke, P.; Baral, S.; Arjyal, A.; Shrestha, A.; King, R. Gender and Antimicrobial Resistance: What Can We Learn From Applying a Gendered Lens to Data Analysis Using a Participatory Arts Case Study? Front. Glob. Women’s Health 2022, 3, 745862. [Google Scholar] [CrossRef] [PubMed]
- Gautron, J.M.C.; Thanh, G.T.; Barasa, V.; Voltolina, G. Using intersectionality to study gender and antimicrobial resistance in low- and middle-income countries. Health Policy Plan. 2023, 38, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, S.F.; Exner, M.; Signorelli, C.; Nath, K.J.; Scott, E.A. The chain of infection transmission in the home and everyday life settings, and the role of hygiene in reducing the risk of infection. Int. Sci. Forum Home Hyg. 2012, 1–140. Available online: https://ifh-homehygiene.org/wp-content/uploads/2019/03/IFHinfectiontransmissionreviewFINAL.pdf (accessed on 6 September 2023).
- Sigudu, T.T.; Oguttu, J.W.; Qekwana, D.N. Prevalence of Staphylococcus spp. from human specimens submitted to diagnostic laboratories in South Africa, 2012–2017. S. Afr. J. Infect. Dis. 2023, 38, 477. [Google Scholar] [CrossRef]
- Petinaki, E.; Spiliopoulou, I. Methicillin-resistant Staphylococcus aureus among companion and food-chain animals: Impact of human contacts. Clin. Microbiol. Infect. 2012, 18, 626–634. [Google Scholar] [CrossRef]
- Emeka-Nwabunnia, I.; Ejigeme, K.; Oguoma, O.I. Antibiotic resistance pattern of methicillin-resistant Staphylococcus aureus isolated from children under the age of five Years in Anambra State. Niger. J. Biotechnol. 2019, 36, 213. [Google Scholar] [CrossRef]
- Ai, X.; Gao, F.; Yao, S.; Liang, B.; Mai, J.; Xiong, Z.; Chen, X.; Liang, Z.; Yang, H.; Ou, Z.; et al. Prevalence, Characterization, and Drug Resistance of Staphylococcus Aureus in Feces From Pediatric Patients in Guangzhou, China. Front. Med. 2020, 7, 127. [Google Scholar] [CrossRef]
- Nielsen, M.V.; Sarpong, N.; Krumkamp, R.; Dekker, D.; Loag, W.; Amemasor, S.; Agyekum, A.; Marks, F.; Huenger, F.; Krefis, A.C.; et al. Incidence and characteristics of bacteremia among children in rural Ghana. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Martínez, J.L.; Lanza, V.F.; Rodríguez-Beltrán, J.; Galán, J.C.; San Millán, A.; Cantón, R.; Coque, T.M. Evolutionary Pathways and Trajectories in Antibiotic Resistance. Clin. Microbiol. Rev. 2021, 34, e00050-19. [Google Scholar] [CrossRef]
- Kardaś-Słoma, L.; Boëlle, P.Y.; Opatowski, L.; Brun-Buisson, C.; Guillemot, D.; Temime, L. Impact of antibiotic exposure patterns on selection of community-associated methicillin-resistant Staphylococcus aureus in hospital settings. Antimicrob. Agents Chemother. 2011, 55, 4888–4895. [Google Scholar] [CrossRef]
- Hayward, A.; Fragaszy, E.; Kovar, J.; Nguyen, V.; Beale, S.; Byrne, T.; Aryee, A.; Hardelid, P.; Wijlaars, L.; Fong, W.L.E.; et al. Risk factors, symptom reporting, healthcare-seeking behaviour and adherence to public health guidance: Protocol for Virus Watch, a prospective community cohort study. BMJ Open 2021, 11, e048042. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, I.N.; Lozano, C.; Zarazaga, M.; Saidenberg, A.B.S.; Stegger, M.; Torres, C. Clonal relatedness of coagulase-positive staphylococci among healthy dogs and dog-owners in Spain. Detection of multidrug-resistant-MSSA-CC398 and novel linezolid-resistant-MRSA-CC5. Front. Microbiol. 2023, 14, 1121564. [Google Scholar] [CrossRef] [PubMed]
- Denissen, J.; Reyneke, B.; Waso-Reyneke, M.; Havenga, B.; Barnard, T.; Khan, S.; Khan, W. Prevalence of ESKAPE pathogens in the environment: Antibiotic resistance status, community-acquired infection and risk to human health. Int. J. Hyg. Environ. Health 2022, 244, 114006. [Google Scholar] [CrossRef] [PubMed]
- Saputra, S. Antimicrobial Resistance in Companion Animal Pathogens in Australia and Assessment of Pradofloxacin on the Gut Microbiota. 2018. Available online: https://digital.library.adelaide.edu.au/dspace/handle/2440/127176 (accessed on 6 September 2023).
- França, A.; Gaio, V.; Lopes, N.; Melo, D.R. Virulence Factors in Coagulase-Negative Staphylococci. Pathogens 2021, 10, 170. [Google Scholar] [CrossRef]
- Owens, R.C. Antimicrobial stewardship: Concepts and strategies in the 21st century. Diagn. Microbiol. Infect. Dis. 2008, 61, 110–128. [Google Scholar] [CrossRef]
- Bru, J.P.; Garraffo, R. Role of intravenous cloxacillin for inpatient infections. Med. Mal. Infect. 2012, 42, 241–246. [Google Scholar] [CrossRef]
- Leong, H.N.; Kurup, A.; Tan, M.Y.; Kwa, A.L.H.; Liau, K.H.; Wilcox, M.H. Management of complicated skin and soft tissue infections with a special focus on the role of newer antibiotics. Infect. Drug Resist. 2018, 11, 1959–1974. [Google Scholar] [CrossRef]
- Lambert, P.A. Bacterial resistance to antibiotics: Modified target sites. Adv. Drug Deliv. Rev. 2005, 57, 1471–1485. [Google Scholar] [CrossRef]
- Tamma, P.D.; Rodriguez-Baňo, J. The use of noncarbapenem β-lactams for the treatment of extended-spectrum β-lactamase infections. Clin. Infect. Dis. 2017, 64, 972–980. [Google Scholar] [CrossRef]
- Lagacé-Wiens, P.; Walkty, A.; Karlowsky, J.A. Ceftazidime-avibactam: An evidence-based review of its pharmacology and potential use in the treatment of Gram-negative bacterial infections. Core Evid. 2014, 9, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Gad, G.F.; Mohamed, H.A.; Ashour, H.M. Aminoglycoside resistance rates, phenotypes, and mechanisms of gram-negative bacteria from infected patients in upper Egypt. PLoS ONE 2011, 6, e17224. [Google Scholar] [CrossRef] [PubMed]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and treatment of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa infections. Clin. Microbiol. Rev. 2019, 32, e00031-19. [Google Scholar] [CrossRef]
- Huber, H.; Koller, S.; Giezendanner, N.; Stephan, R.; Zweifel, C. Prevalence and characteristics of meticillin-resistant staphylococcus aureus in humans in contact with farm animals, in livestock, and in food of animal origin, Switzerland, 2009. Eurosurveillance 2010, 15, 7–10. [Google Scholar] [CrossRef]
- Scaglione, V.; Reale, M.; Davoli, C.; Mazzitelli, M.; Serapide, F.; Lionello, R.; La Gamba, V.; Fusco, P.; Bruni, A.; Procopio, D.; et al. Prevalence of Antibiotic Resistance Over Time in a Third-Level University Hospital. Microb. Drug Resist. 2022, 28, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: Causes and control strategies. Antimicrob. Resist. Infect. Control 2017, 6, 47. [Google Scholar] [CrossRef]
- Alhamami, T.; Chowdhury, P.R.; Gomes, N.; Carr, M.; Veltman, T.; Khazandi, M.; Mollinger, J.; Deutscher, A.T.; Turni, C.; Mahdi, L.; et al. Correction: Alhamami et al. First Emergence of Resistance to Macrolides and Tetracycline Identified in Mannheimia haemolytica and Pasteurella multocida Isolates from Beef Feedlots in Australia. Microorganisms 2021, 9, 1322, Erratum in Microorganisms 2022, 10, 825. [Google Scholar] [CrossRef]
- Dinos, G.P.; Athanassopoulos, C.M.; Missiri, D.A.; Giannopoulou, P.C.; Vlachogiannis, I.A.; Papadopoulos, G.E.; Papaioannou, D.; Kalpaxis, D.L. Chloramphenicol derivatives as antibacterial and anticancer agents: Historic problems and current solutions. Antibiotics 2016, 5, 20. [Google Scholar] [CrossRef]
- Livermore, D.M. Antibiotic resistance in staphylococci. Int. J. Antimicrob. Agents 2000, 16, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Food, E.; Authority, S. EU Summary Report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2013. EFSA J. 2015, 13, 4036. [Google Scholar] [CrossRef]
- Cantón, R.; Morosini, M.I. Emergence and spread of antibiotic resistance following exposure to antibiotics. FEMS Microbiol. Rev. 2011, 35, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, M.; Delgaty, K.L.; Ramotar, K.; Seetaram, C.; Toye, B. Prevalence and mechanisms of erythromycin resistance in group A and group B Streptococcus: Implications for reporting susceptibility results. J. Clin. Microbiol. 2004, 42, 5620–5623. [Google Scholar] [CrossRef] [PubMed]
- Muteeb, G.; Rehman, M.T.; Shahwan, M.; Aatif, M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals 2023, 16, 1615. [Google Scholar] [CrossRef]
- Mitcheltree, M.J. A Platform for the Discovery of New Lincosamide Antibiotics. Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 2018. [Google Scholar]
- Fiebelkorn, K.R.; Crawford, S.A.; McElmeel, M.L.; Jorgensen, J.H. Practical disk diffusion method for detection of inducible clindamycin resistance in Staphylococcus aureus and coagulase-negative staphylococci. J. Clin. Microbiol. 2003, 41, 4740–4744. [Google Scholar] [CrossRef]
- Roberts, J.A.; Pea, F.; Lipman, J. The clinical relevance of plasma protein binding changes. Clin. Pharmacokinet. 2013, 52, 1–8. [Google Scholar] [CrossRef]
- Malbruny, B.; Werno, A.M.; Murdoch, D.R.; Leclercq, R.; Cattoir, V. Cross-resistance to lincosamides, streptogramins A, and pleuromutilins due to the lsa(C) Gene in Streptococcus agalactiae UCN70. Antimicrob. Agents Chemother. 2011, 55, 1470–1474. [Google Scholar] [CrossRef]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef]
- Guardabassi, L.; Schwarz, S.; Lloyd, D.H. Pet animals as reservoirs of antimicrobial-resistant bacteria. J. Antimicrob. Chemother. 2004, 54, 321–332. [Google Scholar] [CrossRef]
- Shafran, S.D. The basis of antibiotic resistance in bacteria. J. Otolaryngol. 1990, 19, 158–168. [Google Scholar] [PubMed]
- Pacheco, T.; Bustos, R.H.; González, D.; Garzón, V.; García, J.C.; Ramírez, D. An approach to measuring colistin plasma levels regarding the treatment of multidrug-resistant bacterial infection. Antibiotics 2019, 8, 100. [Google Scholar] [CrossRef]
- Kakoullis, L.; Papachristodoulou, E.; Chra, P.; Panos, G. Mechanisms of antibiotic resistance in important antibiotic gram-positive and gram-negative pathogens and novel antibiotic solutions. Antibiotics 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Li, Y.; Yao, H. Plasmid-Mediated Transfer of Antibiotic Resistance Genes in Soil. Antibiotics 2022, 11, 525. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial resistance and virulence: A successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Levy, S.B. The impact of antibiotic use on resistance development and persistence. Drug Resist. Updat. 2000, 3, 303–311. [Google Scholar] [CrossRef]
- Aidara-Kane, a. Containment of antimicrobial resistance due to use of antimicrobial agents in animals intended for food: WHO perspective Major WHO initiatives addressing the public health impact of the use in food-producing animals. Rev. Sci. Tech. 2012, 31, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Yaovi, A.B.; Sessou, P.; Tonouhewa, A.B.N.; Hounmanou, G.Y.M.; Thomson, D.; Pelle, R.; Farougou, S.; Mitra, A. Prevalence of antibiotic-resistant bacteria amongst dogs in Africa: A meta-analysis review. Onderstepoort J. Vet. Res. 2022, 89, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.L.; Papp-Wallace, K.M.; Hujer, A.M.; Domitrovic, T.N.; Hujer, K.M.; Hurless, K.N.; Tuohy, M.; Hall, G.; Bonomo, R.A. Unexpected challenges in treating multidrug-resistant gram-negative bacteria: Resistance to ceftazidime-avibactam in archived isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015, 59, 1020–1029. [Google Scholar] [CrossRef]
- Bitrus, A.A.; Peter, O.M.; Abbas, M.A.; Goni, M.D. Staphylococcus aureus: A Review of Antimicrobial Resistance Mechanisms. Vet. Sci. Res. Rev. 2018, 4, 43–54. [Google Scholar] [CrossRef]
- Diekema, D.J.; Pfaller, M.A.; Schmitz, F.J.; Smayevsky, J.; Bell, J.; Jones, R.N.; Beach, M. Survey of infections due to Staphylococcus species: Frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, Latin America, Europe, and the Western Pacific region for the SENTRY Antimicrobial Surveillanc. Clin. Infect. Dis. 2001, 32, S114–S132. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Variable | Total Isolates | AMR b Isolates | MDRc Isolates | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | % | 95% CI a | n | % | 95% CI a | n | % | 95% CI a | |
| Age groups | 404,217 | 219,086 | 54.2 | 153,602 | |||||
| ≥65 | 22,517 | 5.6 | 0.544–0.576 | 11,392 | 5.2 | 0.504–0.536 | 7321 | 4.8 | 0.046–0.049 |
| 60–64 | 2438 | 0.6 | 0.054–0.066 | 7449 | 3.4 | 0.324–0.364 | 693 | 0.5 | 0.004–0.005 |
| 55–59 | 7049 | 1.7 | 0.164–0.176 | 3067 | 1.4 | 0.124–0.156 | 2070 | 1.3 | 0.013–0.014 |
| 50–54 | 11,770 | 2.9 | 0.284–0.304 | 8983 | 4.1 | 0.384–0.436 | 3159 | 2.1 | 0.020–0.022 |
| 45–49 | 14,962 | 3.7 | 0.366–0.374 | 5258 | 2.4 | 0.224–0.264 | 3516 | 2.3 | 0.022–0.024 |
| 40–44 | 19,620 | 4.9 | 0.486–0.494 | 11,612 | 5.3 | 0.504–0.564 | 4458 | 2.9 | 0.028–0.030 |
| 35–39 | 26,031 | 6.4 | 0.636–0.644 | 14,679 | 6.7 | 0.646–0.694 | 5047 | 3.3 | 0.032–0.034 |
| 30–34 | 33,685 | 8.3 | 0.826–0.834 | 17,308 | 7.9 | 0.766–0.812 | 9699 | 6.3 | 0.062–0.065 |
| 25–29 | 34,788 | 8.6 | 0.856–0.864 | 14,241 | 6.5 | 0.626–0.674 | 9198 | 6.0 | 0.058–0.062 |
| 20–24 | 25,897 | 6.4 | 0.636–0.644 | 12,926 | 5.9 | 0.564–0.616 | 6442 | 4.2 | 0.041–0.043 |
| 15–19 | 9610 | 2.4 | 0.236–0.244 | 9202 | 4.2 | 0.364–0.476 | 1957 | 1.3 | 0.012–0.014 |
| 10–14 | 6144 | 1.5 | 0.146–0.154 | 6573 | 3.0 | 0.276–0.324 | 3128 | 2.0 | 0.019–0.021 |
| 5–9 | 13,953 | 3.5 | 0.346–0.354 | 11,173 | 5.1 | 0.476–0.546 | 4756 | 3.1 | 0.030–0.032 |
| 0–4 | 117,566 | 29.1 | 0.289–0.291 | 81,500 | 37.2 | 0.364–0.386 | 74,378 | 48.4 | 0.481–0.488 |
| Unknown | 58,187 | 14.4 | 0.138–0.150 | 29,138 | 13.3 | 0.130–0.136 | 17,779 | 11.6 | 0.114–0.118 |
| Sex | 404,217 | 219,086 | 153,602 | ||||||
| Male | 210,858 | 52.2 | 0.126–0.134 | 97,274 | 44.4 | 0.439–0.482 | 87,092 | 56.7 | 0.565–0.570 |
| Female | 181,270 | 44.8 | 0.443–0.453 | 91,359 | 41.7 | 0.413–0.420 | 60,673 | 39.5 | 0.392–0.397 |
| Unknown | 12,089 | 3.0 | 0.276–0.324 | 30,453 | 13.9 | 0.137–0.141 | 5837 | 3.8 | 0.037–0.039 |
| Variable | Total Isolates | AMR b Isolates | MDR c Isolates | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | % | 95% CI a | n | % | 95% CI a | n | % | 95% CI a | |
| Staph species | 404,217 | 219,086 | 153,602 | ||||||
| CoPS d | 284,973 | 70.5 | 142,406 | 65.0 | 77,108 | 50.2 | |||
| S. aureus | 270,421 | 66.9 | 0.665–0.672 | 131,233 | 59.9 | 0.598–0.600 | 69,474 | 90.1 | 0.896–0.906 |
| S. intermedius | 8084 | 2.0 | 0.188–0.212 | 5696 | 2.6 | 0.025–0.026 | 4168 | 6.0 | 0.562–0.638 |
| S. pseudintermedius | 6468 | 1.6 | 0.148–0.172 | 5477 | 2.5 | 0.024–0.025 | 163 | 3.9 | 0.362–0.418 |
| CoNS e | 71,546 | 17.7 | 50,390 | 23 | 47,309 | 30.8 | |||
| S. epidermidis | 45,272 | 11.2 | 0.109–0.144 | 23,004 | 10.5 | 0.836–0.846 | 34,678 | 73.3 | 0.729–0.737 |
| S. haemolyticus | 14,552 | 3.6 | 0.336–0.376 | 9640 | 4.4 | 0.434–0.448 | 5583 | 16.1 | 0.156–0.166 |
| S. hominis | 7276 | 1.8 | 0.168–0.192 | 11,393 | 5.3 | 0.522–0.538 | 402 | 7.2 | 0.682–0.758 |
| S. saprophyticus | 4446 | 1.1 | 0.108–0.112 | 6353 | 2.9 | 0.286–0.294 | 14 | 3.5 | 0.328–0.372 |
| CoPS/CoNS f | 47,698 | 11.8 | 26,290 | 12.0 | 29,185 | 19.0 | |||
| Unspeciated Staphylococcus | 36,784 | 9.1 | 0.902–0.920 | 15,117 | 6.9 | 0.686–0.694 | 25,945 | 88.9 | 0.884–0.894 |
| S. schleiferi | 5659 | 1.4 | 0.132–0.148 | 5258 | 2.4 | 0.234–0.244 | 1219 | 4.7 | 0.432–0.508 |
| S. hyicus | 5255 | 1.3 | 0.126–0.134 | 5915 | 2.7 | 0.264–0.274 | 78 | 6.4 | 0.602–0.678 |
| AMR b Isolates | MDR c Isolates | |||||
|---|---|---|---|---|---|---|
| n | % | 95% CI a | n | % | 95% CI a | |
| Beta-lactams: | 136,862 | 33.9 | 106,917 | 36.9 | ||
| Ceftazidime | 411 | 0.3 | 0.003–0.004 | 214 | 0.2 | 0.18–0.22 |
| Amoxicillin | 3832 | 2.8 | 0.027–0.029 | 1711 | 1.6 | 1.48–1.72 |
| Ampicillin | 3148 | 2.3 | 0.022–0.024 | 2352 | 2.2 | 2.08–2.32 |
| Cloxacillin | 96,147 | 70.3 | 0.701–0.706 | 79,332 | 74.2 | 73.96–74.44 |
| Penicillin | 19,982 | 14.6 | 0.144–0.148 | 14,755 | 13.7 | 13.68–13.92 |
| Piperacillin | 274 | 0.2 | 0.002–0.003 | 107 | 0.1 | 0.08–0.12 |
| Cefazolin | 274 | 0.2 | 0.001–0.003 | 107 | 0.1 | 0.08–0.12 |
| Cefepime | 411 | 0.3 | 0.002–0.003 | 214 | 0.2 | 0.18–0.22 |
| Cefotaxime | 821 | 0.6 | 0.005–0.006 | 428 | 0.4 | 0.38–0.42 |
| Cefoxitin | 10,128 | 7.4 | 0.073–0.075 | 6736 | 6.3 | 6.18–6.42 |
| Cefuroxime | 958 | 0.7 | 0.007–0.008 | 642 | 0.6 | 0.58–0.64 |
| Ertapenem | 67 | 0.0 | 0.000–0.000 | 100 | 0.1 | 0.00–0.02 |
| Imipenem | 274 | 0.2 | 0.001–0.002 | 214 | 0.2 | 0.18–0.22 |
| Meropenem | 137 | 0.1 | 0.001–0.001 | 107 | 0.1 | 0.08–0.12 |
| Aminoglycosides: | 53,081 | 13.1 | 40,494 | 14.1 | ||
| Gentamicin | 51,655 | 97.3 | 0.992–0.993 | 37,457 | 92.5 | 0.991–0.994 |
| Amikacin | 657 | 1.2 | 0.001–0.003 | 1701 | 4.2 | 0.004-.0005 |
| Streptomycin | 330 | 0.6 | 0.000–0.001 | 486 | 1.2 | 0.000–0.001 |
| Tobramycin | 439 | 0.8 | 0.002–0.009 | 850 | 2.1 | 0.002–0.003 |
| Sulphonamides: | ||||||
| Co-trimoxazole | 15,498 | 3.7 | 0.037–0.038 | 12,601 | 4.3 | 0.035–0.036 |
| Tetracyclines: | ||||||
| Tetracycline | 46,311 | 12.1 | 0.120–0.122 | 31,721 | 10.9 | 0.108–0.110 |
| Macrolides: | ||||||
| Erythromycin | 85,591 | 21.2 | 0.211–0.213 | 56,160 | 19.4 | 0.246–0.249 |
| Lincosamides: | ||||||
| Clindamycin | 56,036 | 15.5 | 0.155–0.156 | 34,759 | 12.1 | 0.190–0.192 |
| Phenicols: | ||||||
| Chloramphenicol | 10,490 | 2.5 | 0.024–0.025 | 7159 | 2.5 | 0.027–0.028 |
| Polymyxins: | ||||||
| Colistin | 348 | 0.1 | 0.001–0.001 | 193 | 0.1 | 0.000–0.001 |
| Variable | AMR c | ||
|---|---|---|---|
| AOR a | 95% CI b | p-Value | |
| Staph. species | |||
| S. aureus | 2.6 | 1.80–2.80 | <0.001 |
| S. intermedius | 0.5 | 0.30–0.90 | <0.001 |
| S. pseudintermedius | Reference | Reference | Reference |
| Specimen source | |||
| Skin | 1.6 | 1.20–2.10 | 0.243 |
| Urinary | 0.8 | 0.60–1.60 | 0.384 |
| Blood | 0.4 | 0.20–2.20 | 0.067 |
| Nasopharyngeal fluid | 0.9 | 0.60–1.80 | 0.672 |
| All other sites | Reference | Reference | Reference |
| Year | 0.97 | 0.94–1.10 | 0.022 |
| Variable | MDR c | ||
|---|---|---|---|
| AOR a | 95% CI b | p-Value | |
| Specimen source | <0.001 | ||
| Skin | 2.7 | 0.80–11.60 | 0.246 |
| Urinary | 6.0 | 2.20–17.60 | 0.407 |
| Blood | 13.0 | 3.40–55.20 | 0.001 |
| Nasopharyngeal fluid | 2.4 | 0.80–7.80 | 0.265 |
| Other sites (Reference) | 1 | - | - |
| Season | 0.022 | ||
| Autumn | 0.9 | 0.80–1.30 | 0.152 |
| Winter | 1.8 | 1.20–2.70 | 0.045 |
| Spring | 1.6 | 0.40–4.60 | 0.355 |
| Summer (Reference) | 1 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sigudu, T.T.; Oguttu, J.W.; Qekwana, D.N. Antimicrobial Resistance of Staphylococcus spp. from Human Specimens Submitted to Diagnostic Laboratories in South Africa, 2012–2017. Microorganisms 2024, 12, 1862. https://doi.org/10.3390/microorganisms12091862
Sigudu TT, Oguttu JW, Qekwana DN. Antimicrobial Resistance of Staphylococcus spp. from Human Specimens Submitted to Diagnostic Laboratories in South Africa, 2012–2017. Microorganisms. 2024; 12(9):1862. https://doi.org/10.3390/microorganisms12091862
Chicago/Turabian StyleSigudu, Themba Titus, James Wabwire Oguttu, and Daniel Nenene Qekwana. 2024. "Antimicrobial Resistance of Staphylococcus spp. from Human Specimens Submitted to Diagnostic Laboratories in South Africa, 2012–2017" Microorganisms 12, no. 9: 1862. https://doi.org/10.3390/microorganisms12091862
APA StyleSigudu, T. T., Oguttu, J. W., & Qekwana, D. N. (2024). Antimicrobial Resistance of Staphylococcus spp. from Human Specimens Submitted to Diagnostic Laboratories in South Africa, 2012–2017. Microorganisms, 12(9), 1862. https://doi.org/10.3390/microorganisms12091862